![]()
![]()
![]()
Bunvong Thaiutsa et Orman Granger
BUNVONG THAIUTSA est membre de la faculté de sylviculture de l'université Kasetsart, Bangkok (Thailande). ORMAN GRANGER est membre du Département de géographie de l'université de Californie, Berkeley (Etats-Unis).
L'importance de la couverture morte en tant qu'élément constituant de l'écosystème forestier a été reconnue depuis longtemps. Une des principales activités dont elle est le siège est la a décomposition», terme général qui s'applique à tout changement dans les caractères biochimiques, l'apparence et le poids. Wood (1974) définit la décomposition des feuilles d'eucalyptus par la perte de poids due à un certain nombre de facteurs, dont l'enlèvement ou la consommation de tissus foliaires par les invertébrés phyllophages. le lessivage, la dégradation biochimique par les micro-organismes ou par passage dans l'intestin des invertébrés. La décomposition est active dans la couverture morte, où les bactéries et les champignons sont, semble-t-il, les premiers colonisateurs de la litière fraîche.
Les écologistes qui ont étudié les formations végétales naturelles se sont particulièrement intéressés à la décomposition de la litière en relation avec le cycle des éléments nutritifs et la productivité du sol. Cela tient manifestement à ce que cette décomposition a un effet marqué sur la disponibilité des éléments nutritifs, qui est un facteur essentiel de la croissance des arbres et de la production ligneuse.
L'étude de la décomposition de la litière fait appel à trois techniques principales: (1) mesure de l'évolution du bioxyde de carbone ou de l'absorption d'oxygène (Reiners, 1968; Wiant Jr., 1967a; 1967b; Witkamp, 1966; Witkamp et Van Der Drift, 1961); (2) observation de la disparition du disque foliaire et d'un constituant spécifique tel que cellulose et lignine (Edwards et Heath, 1963; Heath et al., 1964; 1966; Madge, 1965); (3) mesure de l'activité de la macrofaune et de la microfaune dans les conditions particulières de substrat et de milieu écologique.
Le feuillage des résineux se décompose en général plus lentement que celui des essences décidues (Daubemire et Prusso, 1963; Gosz et al., 1973; Mikola, 1960; Ovington, 1954), du fait que la litière de feuillus contient davantage de potassium et de phosphore, moins de lignine, et presque toujours une fraction soluble à l'éther moins importante que celle des résineux. Les différences sont importantes également entre espèces feuillues (Edwards et Heath, 1963; Heath et al., 1966). Pradhan (1973) indique que la décomposition de litière de teck est plus rapide que celle d'Acacia arabica. En outre, la litière de feuilles disparaît beaucoup plus rapidement que les brindilles et les branches (John, 1973; Rochow, 1974), et les feuilles d'Ombre qui sont plus tendres disparaissent plus rapidement que les feuilles de lumière (Williams et Gray, 1974).
D'une manière générale, la décomposition est le plus rapide chez les espèces ayant une teneur maximale en cendres et en azote, et une teneur en lignine et un rapport C/N minimaux. Elle est la plus lente chez les espèces relativement pauvres en cendres et en azote et qui montrent la teneur en lignine et le rapport C/N les plus élevés. Les espèces ayant une teneur moyenne en cendres, en azote et en lignine, et un rapport C/N moyen semblent avoir une rapidité de décomposition intermédiaire (Singh, 1969). Broadfoot et Pierre (1939) ont trouvé une corrélation significative entre la décomposition de la litière et chacune des cinq variables indépendantes suivantes: excédent basique, matière organique hydrosoluble, azote total, cendres totales et calcium total. Le coefficient de corrélation multiple entre le pourcentage de décomposition et les trois variables, azote total, matière organique hydrosoluble et excédent basique, était de 0,86. Kucera (1959) indique lui aussi une corrélation positive entre rapidité de décomposition d'une part, teneur en cendres et matières solubles à l'eau chaude d'autre part. Etant donné que la composition chimique de la litière influe sur sa vitesse de décomposition. on suppose qu'elle a également une importance déterminante dans la libération des éléments nutritifs.
Les facteurs dominants influant sur l'activité microbienne dans la litière de feuilles sont l'espèce végétale et le climat. L'influence de l'espèce végétale sur la microflore est déterminée par les propriétés physiques et chimiques des feuilles, qui se reflètent dans le rapport C/N. Crossley et Hoglund (1962) ont trouvé une corrélation élevée entre le nombre de micro-arthropodes dans des échantillons de litière et la vitesse de décomposition de la litière. Au cours de l'hiver il y avait peu d'animaux sur les disques foliaires et moins de tissu foliaire consommé qu'en été, où davantage de litière était consommée par des animaux plus nombreux à la surface du sol.
FORÊT TROPICALE AUX CARAIBES comment se transforme-t-elle en humus? à quel rythme?
Le nombre d'arthropodes dans la litière dépend également de sa teneur en humidité. C'est ainsi que Madge (1965) conclut qu'il y a davantage d'animaux sur les disques foliaires à la saison des pluies qu'à la saison sèche. Lorsqu'on a totalement éliminé les animaux pendant 9 mois, il ne s'est produit aucune désintégration visible des feuilles de chêne et de hêtre (Edwards et Heath, 1963). Ces mêmes chercheurs indiquent que les vers de terre enlèvent les disques foliaires trois fois plus vite que les invertébrés plus petits, dont les plus importants sont les podures, les enchytrées et les larvers de diptères. Le carbofuran, insecticide carbamate courant, appliqué aux doses recommandées, réduit le taux de décomposition des feuilles d'érable de Virginie («plaine rouge») à 0,991,26 g/m2/jour. Cela tient surtout à ce que ce type d'insecticides est hautement toxique pour les vers de terre (Weary et Merriam, 1978).
1 ETr = évapotranspiration réelle.
2 ETp = évapotranspiration potentielle.
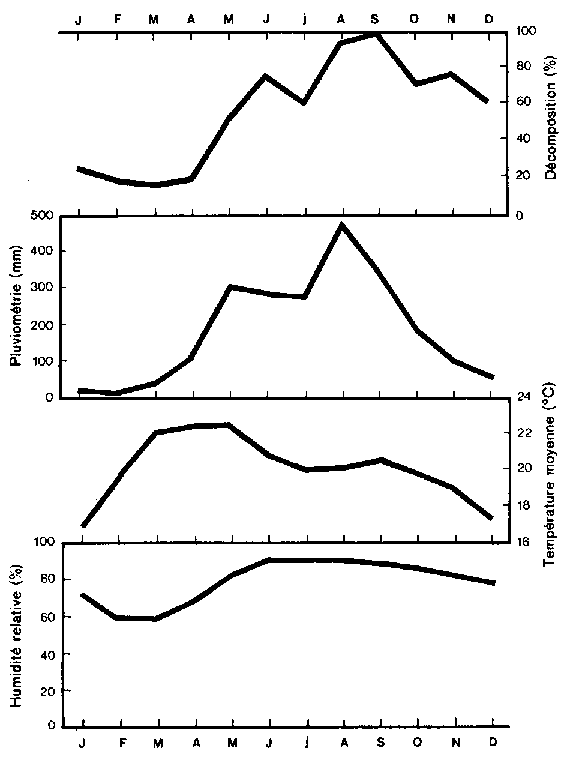
DÉCOMPOSITION DE LA LITIÈRE D 'UNE FORÊT SEMPERVIRENTE DE MONTAGNE EN FONCTION DE LA PLUVIOSITÉ, DE LA TEMPÉRATURE MOYENNE ET DE L'HUMIDITÉ RELATIVE
Conditions climatiques
Le climat modifie de façon marquée la nature et la rapidité de décomposition des débris végétaux à la surface du sol, et exerce de ce fait une forte influence sur le caractère et l'abondance de la matière organique. L'humidité et la température sont parmi les variables les plus déterminantes (Brinson, 1977; Singh, 1969), du fait qu'elles affectent à la fois le développement de la couverture végétale et les activités de micro-organismes qui sont des agents très importants dans la formation des sols. Kononova (1975), citant plusieurs autres publications, conclut que la plus forte intensité de décomposition de la matière organique s'observe dans des conditions de température modérée (de l'ordre de 30°C) et avec une teneur en humidité du sol d'environ 60-80 pour cent de sa capacité maximale de rétention en eau. Un accroissement ou une diminution simultanés de la température et de l'humidité par rapport aux niveaux optimaux ralentissent la décomposition de la matière organique. C'est pourquoi des études sur la décomposition de la litière ne doivent être comparées entre elles que si les conditions de température et d'humidité sont identiques.
Wood (1974) présente des régressions linéaires entre (1) décomposition microbienne et pluviométrie (r = 0,80), (2) décomposition et carbone organique (r = 0,75), (3) décomposition et pH (r = - 0,60). Le résultat obtenu pour le pH traduit l'interaction entre pluviométrie et température, révélant une décomposition lente aussi bien à pH bas (températures basses et pluviométrie élevée) qu'à pH élevé (températures élevées et pluviométrie faible), et une décomposition rapide avec des valeurs de pH intermédiaires.
Des différences dans les vitesses de décomposition à diverses altitudes, dues aux variations de température, sont signalées par Williams et Gray (1974). Shanks et Olson (1961) comparent les vitesses de décomposition de la litière sous des peuplements naturels à diverses altitudes et concluent qu'il y a une diminution de près de 2 pour cent chaque fois que la température moyenne s'abaisse de 1°C; Ce résultat est analogue à ceux de Mikola (1960), qui constate que la vitesse de décomposition diminue lorsque la latitude augmente. L'influence de la température sur la décomposition de la lignine est particulièrement marquée; à 37°C, la lignine se décompose rapidement, 50-60 pour cent disparaissant en l'espace de 9 mois (Waksman et Gerretsen, 1931). Plus récemment, Meentemeyer (1978) a utilisé l'évapotranspiration réelle annuelle comme variable fournissant un indice de prédiction de la vitesse de décomposition.
Reiners (1968) calcule la corrélation entre facteurs du milieu et évolution du bioxyde de carbone, et constate une corrélation élevée entre températures moyennes dans le sol à 5 et 15 cm, et production de bioxyde de carbone. Le taux de production de bioxyde de carbone double chaque fois que la température s'élève de 10°C entre 20 et 40°C (Wiant, Jr., 1967a). Aucune production de bioxyde de carbone n'a été détectée à 10°C, et à 50°C et plus elle diminue fortement. L'évolution du bioxyde de carbone est par ailleurs fortement influencée par la teneur en humidité. Dans une matière organique non incorporée au sol et dont la teneur en humidité tombait au-dessous de 20 pour cent, ou dans un sol minéral où elle était tombée à seulement 5 pour cent, on n'a pu détecter aucune production de bioxyde de carbone (Wiant, Jr., 1967b).
Cependant, on a rapporté certaines conclusions contradictoires sur les effets du climat. Van Der Drift (1963) souligne que les précipitations influent davantage que la température sur la décomposition de la litière, bien qu'un hiver doux et un printemps chaud, associés à une humidité du sol proche de la normale, puissent l'accélérer, de même que l'humidité pénétrant à travers les détritus. L'eau de pluie peut, en percolant, lessiver les excréments et les débris organiques vers les horizons inférieurs, où d'autres microbes spécialisés attaqueront les molécules organiques subsistantes. C'est pourquoi la décomposition de la litière est plus lente lors des années sèches qu'en année moyenne. Par contre, selon Jenny et al. (1949) et Witkamp (1966) c'est la température qui est le facteur le plus important. suivie par la pluviométrie.
Etant donné que le climat est le principal facteur qui commande la décomposition, la vitesse de désintégration varie avec la saison. Gosz et al. (1973) et Loomis (1975) ont constaté que la décomposition des résidus végétaux était rapide en été, tandis que Lang (1974) estime la décomposition d'une litière de feuilles à3,75 g/m2/jour durant les mois d'automne et à 0,80 g/m2/jour pendant le reste de l'année. Boonyawat et Ngampongsai (1974) confirment les résultats de Lang en constatant que la décomposition la plus active, pour une litière de forêt à feuilles persistantes de montagne, se situe à la fin de la saison des pluies et au début de l'hiver (0,36 t/ha/mois), et qu'elle est au niveau le plus bas en été (0,14 t/ha/mois). Madge (1965) conclut que la disparition de la litière, au Nigéria, a lieu surtout durant la saison humide, en raison de l'activité des acariens et des collemboles.
Bien que les différences dans les vitesses de décomposition entre régions tempérées et régions tropicales aient été étudiées il y a quelques dizaines d'années par Jenny et al. (1949), on possède peu de données concernant le Sud-Est asiatique. La plupart des études antérieures sur la production et la décomposition de la litière forestière en Thaïlande (Aksornkoae et. al. 1972; Boonyawat et Ngampongsai 1974; Paovongsar, 1976; Thaiutsa et al., 1978) notaient les données climatiques, mais négligeaient l'interprétation analytique. L'objectif de la présente étude, par conséquent, est d'élaborer des modèles de régression permettant de prédire la vitesse de décomposition à partir des données climatiques.
L'étude a été réalisée à la station de recherche sur les bassins versants de Kogma, située à 18°45' de latitude N. 98°54' de longitude E. dans le nord de la Thaïlande. Les pentes sont inférieures à20 pour cent et l'altitude est comprise entre 1250 et 1500 m. Le sol s'est formé sur roche-mère de granit, de gneiss et de quartzite, et est classé dans le grand groupe des sols latéritiques brun rougeâtre. Du point de vue texture, c'est un sable limoneux faiblement argileux; il est fortement lessivé, ce qui a donné naissance à un horizon B profond. Selon Spurr et Barnes (1973), le type de forêt sempervirente que l'on y trouve correspond à la subdivision désignée sous le terme de «forêt ombrophile montagnarde». Les principales essences que l'on y rencontre appartiennent à la famille des fagacées, comprenant Castanopsis acuminatissima, Lithocarpus polystachyus, Quercus kerrii et Q. kingiana. Le degré de couverture est d'environ 85 pour cent.
Tableau 2. Equations de régression linéaire entre pourcentage de décomposition (D) et humidité relative (HR, %) pluviométrie (P. mm), température moyenne (T,°C), température maximale (Tmax, °C), évapotranspiration réelle mensuelle (ETrm, mm), indice de siccité (IS), et facteur hydrique (fH) dans une forêt sempervirente montagnarde, Chiang Mai, Thaïlande
|
Equations |
r |
r2 |
|
D = 2,286 HR - 125,481 |
0,89 |
0,79 |
|
D = 30,096 + 0,149 P |
0,77 |
0,59 |
|
D = 242,712-7,860 Tmax |
- 0,52 |
0,27 |
|
D = 26,267 + 14,122 IS |
0,85 |
0,72 |
|
D = 0,891 fH - 2,103 |
0,75 |
0,56 |
|
D = 1,626 HR - 0,466 ETrm + 0,899 P - 53,948 |
0,93 |
0,86 |
|
D = 2,284 HR - 0,910 ETrm + 6,222 T + 0,607P - 192,715 |
0,94 |
0,88 |
Tableau 3. Matrice de corrélation et tableau résumé de corrélation multiple par étapes
|
Variable |
X1 |
X2 |
X3 |
X4 |
|
% |
% |
mm |
mm |
|
|
X1 |
1,00 |
0,89 |
0,17 |
0,77 |
|
X2 |
|
1,00 |
0,35 |
0,80 |
|
X3 |
|
|
1,00 |
0,61 |
|
X4 |
|
|
1,00 |
|
|
Étape n° |
Variable entrée |
r |
r2 |
Accroissement r2 |
Seuil de signification |
|
1 |
X2 |
0,89 |
0,79 |
0,79 |
0,000 |
|
2 |
X3 |
0,90 |
0,81 |
0,02 |
0,001 |
|
3 |
X4 |
0,93 |
0,86 |
0,05 |
0,001 |
On disposa de façon aléatoire sur le sol 156 trappes en bois dans tout le bassin versant de 0,65 km2 afin de mesurer les entrées et sorties de litière. Chaque trappe mesurait 0,75 x 0,75 m. La décomposition de la litière fut suivie dans douze d'entre elles. La litière fut récoltée chaque mois, de janvier 1968 à décembre 1972. Après séchage à 70°C pendant 24 h elle était pesée, la perte de poids étant supposée représenter le taux de décomposition mensuel. Température, pluviométrie et humidité relative étaient notées.
Les éléments du bilan hydrique furent calculés par ordinateur selon le programme proposé par Stone, Jr. (1971, «Water Balance Computer Programme»), en se basant sur les tables de Thornthwaite et Mather (1957), qui donnent les valeurs de l'évapotranspiration potentielle non corrigées en fonction de la température moyenne et des valeurs de I (indice thermique). Ces tables fournissent également la durée moyenne mensuelle d'insolation en fonction de la latitude.
Deux variables secondaires furent déduites des données climatiques de base. Le «facteur hydrique» est calculé par l'équation empirique indiquée par Kononova (1975)
fH = 43,2 log P - T
dans laquelle fH est le facteur hydrique, P la pluviométrie mensuelle en millimètres, et T la température moyenne en degrés centigrades. L'autre variable, le «taux de siccité» (TS), est calculée approximativement par l'équation

dans laquelle P est la pluviométrie mensuelle en millimètres, ETp l'évapotranspiration potentielle en millimètres.
Des régressions simples et multiples entre pourcentage de décomposition de la litière forestière et variables climatiques primaires et secondaires furent établies. Dans le cas des régressions multiples, on employait une technique de sélection par étapes, consistant à établir par ordinateur, pour chaque relation, une série d'équations de régression multiple variant par degrés. A chaque degré, la variable ajoutée est celle qui entraîne la plus grande réduction de la somme des carrés des écarts, ou celle qui a la corrélation partielle la plus élevée avec la variable dépendante lorsqu'on donne une valeur fixe à la variable qui a déjà été ajoutée, ou encore celle qui, si on l'avait ajoutée, aurait eu la valeur F la plus élevée. La série d'interactions est fractionnée au moyen de critères prédéterminés basés sur certaines valeurs F et sur un certain degré de tolérance.
Les données climatiques originales et dérivées et les taux de décomposition mensuels dans la forêt sempervirente montagnarde sont présentés dans le tableau 1. Les données climatiques primaires sont l'humidité relative, la pluviométrie, la température maximale et la température moyenne. On constate que le pourcentage de décomposition s'accroît lorsque la pluviométrie et l'humidité augmentent. La tendance est inverse pour la température. La figure montre que le taux de désintégration suit étroitement l'intensité des pluies, avec un retard d'environ un mois. Par exemple, un premier maximum de décomposition se produit en juin alors que le maximum de pluviométrie se situe en mai. On constate un second maximum de décomposition et de pluviométrie en septembre et en août respectivement.
Un élément climatique important dérivé de la pluviométrie et de la température observées est l'évapotranspiration. L'«évapotranspiration réelle», c'est-à-dire la quantité d'eau effectivement évaporée et transpirée, est le seul terme gui traduise la quantité d'énergie et d'humidité simultanément disponible pour les écosystèmes naturels. L'«évapotranspiration potentielle», par contre, représente une perte d'eau théorique dans l'hypothèse d'une disponibilité en eau illimitée. Si la température n'est pas considérée sous les tropiques comme un facteur climatique limitant c'est parce qu'elle joue un rôle dans l'évapotranspiration potentielle, qui est constamment élevée. C'est cette particularité de la température qui fait de la pluviométrie le plus important des facteurs climatiques limitants.
Meentemeyer (1974) cite une centaine d'études sur le rôle de l'évapotranspiration dans la décomposition de la litière, et établit des régressions positives entre le pourcentage de dé composition et l'évapotranspiration réelle. Toutefois, les résultats de la présente étude ne font pas apparaître une relation aussi forte entre la décomposition de la litière forestière et l'évapotranspiration réelle. Le coefficient de corrélation trouvé n'est que de 0,17, alors que dans les études antérieures on avait des valeurs supérieures à 0,90. Il est généralement reconnu que la formule de Thornthwaite sous-estime l'évapotranspiration réelle dans les climats de régime tropical, ce qui peut avoir influencé les résultats. Il est possible également que la valeur de la capacité de rétention en eau du sol utilisée dans les calculs d'ordinateur (10 inches = 254 mm) était trop basse pour les sols tropicaux.
Tableau 4. Taux de décomposition de la litière forestière au cours de la première année dans différentes localités
|
Type de foret ou essence |
Localité |
Composition 1re année (%) |
Références |
|
Sempervirente montagnarde |
Thaïlande |
56 |
Présente étude |
|
Plantation de teck |
Thaïlande |
95 |
Aksornkoae et al., 1972 |
|
Peuplement de teck |
Nigéria |
100 |
Egunjobi, 1974 |
|
Forêt tropicale humide |
Trinité |
100 |
Cornforth, 1970 |
|
Forêt tropicale humide |
Ghana |
100 |
Nye, 1961 |
|
Hêtre et chêne |
Angleterre |
75 |
Edwards & Heath, 1963 |
|
Végétation annuelle |
New Jersey |
58 |
Lang, 1973 |
|
Graminées pérennes et buissons |
New Jersey |
35 |
Lang, 1973 |
|
Chêne, érable de Virginie, genévrier de Virginie |
New Jersey |
39-48 |
Lang, 1973 |
|
Chêne |
New Jersey |
85 |
Lang, 1974 |
|
Erable de Virginie |
Canada |
100 |
Weary & Merriam, 1978 |
|
Eucalyptus delegatensis |
Australie |
35-97 |
Wood, 1974 |
Les équations de régression simple et multiple établies dans cette étude sont résumées dans le tableau 2. On a constaté que les pourcentages de décomposition dans la forêt sempervirente montagnarde de la Thaïlande septentrionale étaient en corrélation significative avec quatre composantes climatiques: humidité relative, pluviométrie, indice de siccité et facteur hydrique. La température maximale montre un coefficient de corrélation (r = - 0,52) bien meilleur que la température moyenne (r = - 0,10). On a constaté par ailleurs un accroissement de désintégration de la litière de 2,29 pour cent chaque fois que l'humidité relative s'élève de 1 pour cent. De même, pour le site étudié, le taux de décomposition s'accroît de 0,15 pour cent par millimètre de pluviométrie supplémentaire.
Les facteurs climatiques qui influent le plus sur la décomposition ont été isolés par application de la technique de régression par étapes et sont présentés dans le tableau 3. L'humidité intervient à elle seule pour 79 pour cent de la variance totale dans les taux de décomposition observés. L'évapotranspiration réelle mensuelle ajoute 2 pour cent, la pluviométrie 5 pour cent. Les valeurs de r2 pour les deux régressions linéaires multiples montrent que l'emploi de facteurs climatiques multiples permet une meilleure prédiction que l'emploi de variables simples. Ni la température moyenne ni la température maximale n'ont l'influence importante sur la décomposition de la litière que les résultats des recherches antérieures auraient pu faire attendre. Il apparaît donc clairement que les précipitations jouent ici un plus grand rôle que les températures.
Il ne faut pas, toutefois, perdre de vue que la hauteur de pluie mesurée au pluviomètre peut être moins significative que l'eau du sol, notamment l'humidité du sol superficiel et de la litière. L'eau du sol joue un rôle plus direct dans l'activité de la faune du sol que la pluviométrie mesurée, en partie parce que ce rôle est indépendant de l'excédent hydrique ou du ruissellement de surface. On a tenté dans cette expérience de montrer la relation entre ruissellement superficiel et décomposition de la litière; on a obtenu l'équation
Y = 30,869 + 2,644 X,
dans laquelle Y est le pourcentage de décomposition et X le ruissellement en centimètres (le coefficient de corrélation r est égal à 0,90). Cette corrélation semble toutefois empirique, car au cours des mois de ruissellement maximal (août et septembre) la disparition de litière peut être due pour la plus grande part à l'entraînement par l'eau plutôt qu'à la décomposition. C'est d'ailleurs la principale limitation de cette technique des casiers employée pour évaluer la perte de poids de la litière. L'humidité du sol et l'humidité de la litière seront donc vraisemblablement plus importantes que la pluviométrie totale. On peut penser qu'elles représentent les «précipitations efficaces», ou l'humidité totale disponible après déduction du ruissellement superticiel immédiat.
Etant donné que la vitesse de décomposition varie selon l'espèce végétale, le climat et les micro-organismes présents, la durée nécessaire pour la minéralisation complète est variable. Le tableau 4 fournit des données sur le taux de décomposition de résidus végétaux au cours de la première année, pour diverses stations. La décomposition au cours des 12 premiers mois (56,23 pour cent de la litière produite) semble moins élevée que ne l'indiquaient les études antérieures en zone tropicale. Cependant, en utilisant une équation proposée par Olson (1963) dans le cas d'un état stable
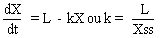
on trouve pour ce bassin versant une valeur de k égale à 2,28. Or le paramètre de décomposition k pour les foréts tropicales varie, selon Olson, entre un peu plus de 1 et un peu plus de 4 dans n'importe quel peuplement; la valeur de 2,28 trouvée ici se situe donc dans la fourchette. A ce taux, la minéralisation complète se produirait en l'espace de 1,8 année. Des différences dans la durée de ce cycle influent sur le développement des écosystèmes naturels, en particulier dans les zones tropicales où la litière forestière a une influence sur les feux de brousse et sur les phénomènes de lessivage. C'est pourquoi des méthodes appropriées de contrôle de la litière sont nécessaires.
Une étude des effets de la pluviométrie, des températures et de certaines composantes modifiées du climat sur la décomposition de la litière dans une forêt sempervirente montagnarde du nord de la Thailande a montré que 56,23 pour cent de la litière formée annuellement se décomposent en 12 mois, avec un paramètre de décomposition k de 2,28. La minéralisation complète de cet écosystème se produirait en 1,8 année. Le pourcentage de décomposition montre une corrélation significative avec l'humidité relative, la pluviométrie, l'indice de siccité et le facteur hydrique. On a trouvé une relation négative entre pourcentage de décomposition et température maximale. Une régression sélective par étapes indique que 86 pour cent de la décomposition au cours de la première année résultent de l'humidité, de la pluviométrie et de l'évapotranspiration réelle. Lorsqu'on y ajoute la température, moyenne ou maximale, comme quatrième variable indépendante, ce pourcentage n'est pas modifié de façon significative. On a par ailleurs constaté que les précipitations jouent un plus grand rôle dans la décomposition de la litière que la température. La désintégration de la litière s'accroît de 0,15 pour cent chaque fois que la pluviométrie augmente de 1 mm. L'eau du sol superficiel et la teneur en humidité de la litière ne devront pas être négligées dans les recherches futures.
L'établissement de modèles mathématiques pour la décomposition de la litière en relation avec les facteurs climatiques pourra requérir certaines études de laboratoire, toutefois seules des recherches sur le terrain permettront une compréhension pratique de l'évolution des écosystèmes. Cette expérimentation sur le terrain devra couvrir toute la large gamme de conditions climatiques. Il faudra tenir compte de l'eau du sol superficiel et de la teneur en humidité de la litière. Des études détaillées sur la production et la décomposition de la litière forestière dans divers types de foréts de toute la Thaïlande sont prévues dans un avenir proche.
AKSORNKOAE, &, KHENMARK, C. & KAEWLA IAD, T. 1972. [Study on organic matter in teak plantation]. Kasetsart University Forestry Research Bulletin No. 23, 36 p. (En thaïlandais avec sommaire en anglais)
BOONYAWAT, S. & NGAMPONGSAI, C. 1974. An analysis of accumulation and decomposition of litter fall in hill evergreen forest, Doi Pui, Chiangmai. Kasetsart University Kogma Watershed Research Bulletin No. 17. 21 p. (En thaïlandais avec sommaire en anglais)
BRINSON, M.M. 1977. Decomposition and nutrient exchange of litter in an alluvial swamp forest. Ecology, 58:601-609.
BROADFOOT, W.M. & PIERRE, W.H. 1939. Forest soil studies: I. Relation of rate of decomposition of tree leaves to their acid-base balance and other chemical properties. Soil Sci., 48:329-348.
CORNFORTH, I.S. 1970. Leaf-fall in a tropical rain forest. J. appl. Ecol., 7:603-608.
CROSSLEY, D.A. & HOGLUND, M.P. 1962. A litter-bag method for the study of microarthropods inhabiting leaf litter. Ecology, 43:571-574.
DAUBEMIRE, R. & PRUSSO, D.C. 1963. Studies of the decomposition rates of tree litter. Ecology, 44:589-592.
EDWARDS, C.A. & HEATH, G.W. 1963. The role of soil animals in breakdown of leaf material. Dans Soil organisms, ed. by J. Doeksen and J. van der Drift, p. 76-84. Amsterdam. North-Holland.
EGUNJOBI J.K. 1974. Litter fall and mineralization in a teak (Tectona grandis) stand. Oikos, 25:222-226.
Gosz, J.R., LIKENS, G.E. & BORMANN, F.M. 1973. Nutrient release from decomposing leaf and branch litter in the Hubbard Brook Forest, New Hampshire. Ecol. Monogr., 43:173-191.
HEATH, G.W., ARNOLD, M.K. & EDWARDS, C.A. 1966. Studies in leaf litter breakdown. I. Breakdown rates of leaves of different species. Pedobiol., 6:1-12.
HEATH, G.W., EDWARDS, C.A. & ARNOLD M.K. 1964. Some methods for assessing the activity of soil animals in the breakdown of leaves. Pedobiol., 4:80-87.
JENNY, H., GESSEL, S.P. & BINGHAM, F.T. 1949. Comparative study of decomposition rates of organic matter in temperate and tropical regions. Soil Sci. 68:419-432.
JOHN, D.M. 1973. Accumulation and decay of litter and net production of forest in tropical West Africa. Oikos 24:430-435.
KONONOVA, M.M. 1975. Humus of virgin and cultivated soils. Dans Soil components, Vol. 1, ed by J.E. Gieseking p. 475-526. New York, Springer-Verlag.
KUCERA, C.L. 1959. Weathering characteristics of deciduous leaf litter. Ecology, 40:485-487.
LANG, G.E. 1973. Litter accumulation through ecosystem development. Rutgers University, New Brunswick, New Jersey. 105 p. (Thèse)
LANG, G.E. 1974. Litter dynamics in a mixed oak forest on the New Jersey Piedmont. Bull. Torrey Bot, Club, 101:277-286.
LOOMIS, R.M. 1975. Animal changes in forest floor weights under a southeast Missouri oak stand. USFS Res. Note NC - 184. 3 p.
MADGE, D.S. 1965. Leaf fall and litter disappearance in a tropical forest. Pedobiol., 5:273-288.
MEENTEMEYER, V. 1974. Climatic water budget approach to forest problems. II. The prediction of regional differences in decomposition rates of organic debris. Publ. Climatol., 27(1): 35-74.
MEENTEMEYER, V. 1978. Macroclimate and lignin control of litter decomposition rates. Ecology, 59:465-472.
MIMOLA, P. 1960. Comparative experiment on decomposition rates of forest litter in southern and northern Finland. Oikos, 11:161-166.
NYE, P.H. 1961. Organic matter and nutrient cycles under moist tropical forest. Plant & Soil, 13:333-346.
OLSON, J.S. 1963. Energy storage and the balance of producers and decomposers in ecological systems. Ecology, 44:322-331.
OVINGTON, J.D. 1954. Studies of development of woodland conditions under different trees. II. The forest floor. 1. Ecol., 42:71-80.
FAOVONGSAR, S. 1976. Litter fall and mineral nutrient content of litter in dry dipterocarps forest. (Thèse)
PRADHAN, I.P. 1973. Preliminary study of interception through leaf litter. Indian For., 99:440-445.
REINERS, W.A. 1968. Carbon dioxide evolution from the floor of there Minnesota forests. Ecology, 49:471-483.
ROCHOW, J.J. 1974. Litter fall relations in a Missouri forest. Oikos, 25:80-85.
SHANKS, R.E. & OLSON, J.S. 1961. Firstyear breakdown of leaf litter in southern Appalachian forests. Science, 134:194-195.
SINGH, K.P. 1969. Studies in decomposition of leaf litter of important trees of tropical deciduous forest at Varanasi. Trop. Ecol., 10:292-311.
SPURR S.H. & BARNES, B.V. 1973. Forest ecology. New York. Ronald Press. 571 p.
STONE, L.A. JR. 1971. Water balance computer program using consecutive monthly data. Dans Computer program for the climatic water balance, ed. by L.A. Stone, Jr., J.C. Albrecht, and G.A. Yoshioka. Publ. Climatol., 24(3): 1-10.
THAIUTSA, B., SUWANNAPINUNT & KAITPRANEET, W. 1978. Production and chemical composition of forest litter in Thailand. Kasetsart University Forest Research Bulletin No. 52. 32 p.
THORNTHWAITE, C.W. & MATHER, J.R. 1957. Instructions and tables for computing potential evapotranspiration and the water balance. Publ. Climatol., 10(3): 185-311.
VAN DER DRIFT, J. 1963. The disappearance of litter in mull and mor in connection with weather conditions and the activity of the macrofauna. Dans Soil organisms, ed. by J. Doeksen and J. van der Drift, p. 124-133. Amsterdam, North Holland.
WAKSMAN, S.A. & GERRETSEN, F.C. 1931. Influence of temperature and moisture up on the nature and extent of de composition of plant residues by microorganisms. Ecology, 12:33-60.
WEARY, G.C. & MERRIAM, H.G. 1978. Litter decomposition in a red maple woodlot under natural conditions and under insecticide treatment. Ecology, 59:180-184.
WIANT, H.V. JR. 1967a. Influence of temperature on the rate of soil respiration. 1. For. 65:489-490.
WIANT, H.V. JR. 1967b. Influence of moisture content on soil respiration. J. For., 63:902-903.
WILLIAMS, S.T. & GRAY, T.R.G. 1974. Decomposition of litter on soil surface. Dans Biology of litter decomposition, Vol. 2, ed. by C.H. Dickinson and G.J.E. Pugh, p. 611-630. New York, Academic Press.
WITKAMP, M. 1966. Decomposition of litter in relation to environmental factors. Plant & Soil, 15:295-311.
WOOD, T.G. 1974. Field investigations on the decomposition of leaves of Eucalyptus delegatensia in relation to environmental factors. Pedobiol. 14:343-371.
La forêt au service de la collectivité
La documentation qu'il vous faut et que vous pouvez vous procurer auprès de
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}