![]()
![]()
![]()
BENT SØEGAARD
Todo científico consciente concede importancia a la resistencia de los árboles contra el ataque de insectos. Existe una imperiosa necesidad de intensificar la labor en este sentido, precisándose una estrecha colaboración entre entomólogos, fisiólogos y genetistas.
Uno de los ejemplos mejor conocidos de resistencia contra insectos lo proporciona el gorgojo de pino blanco, Pissodes strobi, y su relación con Pinus strobus; tanto el gorgojo como el pino han sido muy estudiados. Otros ejemplos son el gorgojo de la reproducción del pino, Cylindrocopturus eatoni y su ataque a Pinus jeffreyi y P. coulteri y el híbrido retrocruzado Pinus jeffreyi x (jeffreyi x coulteri), que ha mostrado una resistencia notable. También la mosca de la resina, Retinodiplosis sp., y los escarabajos de la corteza Dendroctonus han sido estudiados en pinos en California. En el abeto de Douglas, Pseudotsuga taxifolia, los estudios sobre resistencia a Chermes cooleyi han conducido a resultados interesantes.
Entre las especies frondosas Be hace mención a la resistencia de Robinia pseudoacacia al perforador de la robinia, Cyllene robiniae y en los chopos ha sido reseñada resistencia al escarabajo japonés de la corteza.
El término «resistencia» Be refiere a árboles que son menos perjudicados o están menos infestados que otros bajo condiciones ambientales comparables. La resistencia puede ser debida a las siguientes razones:
1. El árbol puede no ser preferido para la postura, cobijo o alimentación.2. Puede poseer un componente de la resistencia llamado antibiosis y puede afectar a la biología del insecto.
3. El árbol resistente puede ser tolerante y sobrevivir bajo niveles de infestación que matarían o perjudicarían severamente a otro susceptible.
La variación en la resistencia a los insectos es la base para la selección y mejora. Hay ejemplos que demuestran esta variación en pino blanco, alerce y abeto de Douglas. La razón de la resistencia ha sido investigada para el gorgojo de la reproducción del pino y es atribuida o a una capa de corcho que rodea la punción de la puesta en la corteza primaria o a alguna propiedad de la resina. Otros ejemplos tienen que ver con la resina como factor de resistencia. Los métodos para investigar y mejorar la resistencia se basan en la selección. La comparación de la selección directa e indirecta aconseja el empleo de un sistema de selección mixto que combine las ventajas de ambos métodos. En el ensayo del abeto de Douglas para resistencia al Chermes cooleyi, el invernadero proporciona condiciones de «laboratorio». La producción en masa del material resistente puede hacerse por el injertado, enraizamiento de estacas o sirviéndose de los huertos para semilla.
Los estudios sobre la genética y biología de Ips confusas y Dendroctonus pseudotsugae Be extienden también a la forma de vuelo.
Finalmente, se llama la atención sobre la importancia de establecer la colaboración internacional.
BENT SØEGAARD es genetista forestal en el Arboreto del Colegio Real de Veterinaria y Agricultura de Hørsholm Dinamarca. Fue ayudado en la preparación de la versión final de este capítulo por H. D. Gerhold (Estados Unidos). Otros miembros del equipo de trabajo fueron E. J. Schreiner (Estados Unidos), E. Björkman (Suecia), R. T. Bingham (Estados Unidos) y O. Fugalli (FAO).
En los pasados 20 años ha habido un claro interés creciente por la resistencia de las plantas a los ataques de insectos, afectando no sólo a la investigación agrícola sino también a la forestal. El empleo de insecticidas y el subsiguiente desarrollo de resistencia por parte de los insectos a muchos de aquellos ha hecho comprender a los entomólogos y genetistas que es preciso emplear toda clase de medios posibles en esta lucha. Painter (1958) sugiere que debe ser parte importante de todo proyecto bien preparado de lucha contra los insectos la investigación de las causas de resistencia a ellos y el empleo de variedades y cultivares que la posean, para así reducir sus poblaciones y los daños que causan. También Schreiner (1960) insiste en la importancia de mejorar la resistencia contra los insectos de las especies forestales.
Se ha trabajado muy poco sobre la clase de mejora genética que nos ocupa en el ámbito forestal, tanto por parte de los entomólogos como por parte de los genetistas. Es esencial una colaboración estrecha con los que se ocupan de la resistencia a enfermedades, ya que los problemas en esencia son los mismos para ambos. La cooperación entre entomólogos, fisiólogos y mejoradores de especies forestales es muy necesaria para una mejor comprensión de las causas de la resistencia.
Los insectos afectan al crecimiento de los árboles principalmente comiendo los tejidos de las plantas, chupando los flúidos de los tejidos, haciendo las puestas y provocando la formación de agallas. También influyen en el crecimiento actuando como vectores de algunas de las más importantes enfermedades de hongos, bacterias y virus de los árboles. Graham (1925) recalca que no hay etapa en el ciclo de la vida de un árbol que esté libre del ataque de insectos. Las semillas pueden ser atacadas aún antes de su recolección y también pueden serlo más tarde en el almacén. Los brinzales y pimpollos son atacados y a menudo muertos por una gran variedad de insectos. En general parece que los árboles adquieren alguna resistencia cuando han pasado del estado de bajo latizal. Sin embargo, cuando se aproximan a la madurez y su actividad fisiológica declina, se vuelven de nuevo susceptibles a muchos ataques de insectos.
El gorgojo de pino blanco, Pissodes strobi, es considerado desde hace mucho tiempo como la plaga más importante del Pinus strobus a lo largo de toda su área. Los daños al árbol consisten en la pérdida de la guía durante la alimentación larval. El primer ataque serio del gorgojo ocurre normalmente cuando la planta tiene cinco años y 1 metro (3 pies) o menos de altura; luego el ataque aumenta en severidad hasta que el árbol alcanza una altura de 8 a 10 metros (25 a 30 pies). Sullivan (1959) investigó la relación entre las actividades del gorgojo y las características climáticas de su habitat, y cree que un 50 por ciento de la luz incidente marca la línea divisoria entre los daños severos y moderados. Wright y Gabriel (1959) afirman que se ha comprobado suficiente resistencia, como para servir de punto de partida de cultivares resistentes al gorgojo con características de crecimiento buenas y que vale la pena emprender la producción de un Pinus strobus que sea resistente genéticamente al Pissodes strobi. Gerhold (1962) analiza lo que debiera ser objeto de selección y afirma que es necesario medir con exactitud el grado de resistencia al gorgojo y clasificar exactamente los tipos de resistencia inherente en descendencias de pino blanco.
Se hicieron injertos de cabeza con púas de Pinus strobus seleccionados, sobre plantas jóvenes de éste, en una zona muy infectada de gorgojo, al mismo tiempo que se injertaron como testigos púas procedentes de pinos fuertemente atacados por el gorgojo. Los injertos de púas procedentes de una plantación de origen norteño fueron atacados al sur de Ontario con severidad claramente menor que los injertos de púas procedentes de árboles locales que padecían la plaga.
Los árboles de corteza delgada y guías menores son menos susceptibles según Barnes (1928), Prebble (1951), Kriebel (1954) y Sullivan (1957); mientras Fowler (1957) sugirió que un flujo fuerte de resina en el Pinus peuce puede ser la causa de la recuperación del ataque en esta especie. Los experimentos de Anderson y Fisher (1956, 1960) y las observaciones de Barnes (1928) y de MacAloney (1930) sugieren que actúan olores atrayentes y repelentes relacionados específicamente con el hospedante.
Ha sido comunicada también la presencia del Pissodes strobi sobre Pinus resinosa y P. banksiana (Heimburger, 1963). Incluso el Picea abies es atacado (Holst, 1955).
En Berkeley y Placerville, California, E.U.A., se investigó la resistencia de varias especies de pino y de sus híbridos al ataque y a los daños del gorgojo de la reproducción del pino Cylindrocopturus eatoni. Miller (1950) demostró que el Pinus coulteri y el híbrido retrocruzado Pinus jeffreyi x (jeffreyi X coulteri) poseían una resistencia notable al ataque y daños del gorgojo de la reproducción en ensayos en jaulas bajo condiciones de vivero; en contraste, el P. jeffreyi fue severamente atacado. Hall (1959) informó que las dos plantaciones experimentales dañadas por el gorgojo han demostrado con seguridad casi total que el híbrido retrocruzado Pinus jeffreyi X (jeffreyi x coulteri) tiene un alto grado de resistencia al ataque y daños por el gorgojo de la reproducción bajo condiciones de plantación en el monte.
Austin et al. (1945) describieron el daño causado al Pinus ponderosa por la mosca de la resina (Retinodiplosis sp.) y hallaron que los árboles con brotes glaucos (recubiertos de vello) y glabros (secos y lisos) padecían daños pequeños o ninguno, mientras que los árboles con brotes pegajosos (untados y cubiertos de resina) los padecieron mayores. Se sugiere que los árboles susceptibles al ataque de este insecto sean rápidamente señalados para la extracción del monte, eligiéndolos por el aspecto superficial de los brotes.
Aparte de lograr árboles que resistan al gorgojo de la reproducción del pino, ha sido muy estudiada en California la cuestión de obtener pinos resistentes a los escarabajos de la corteza, Dendroctonus. Se han conseguido algunas orientaciones prometedoras sobre la identificación de la resistencia y su causa.
Las investigaciones hechas en Dinamarca sobre la resistencia de Pseudotsuga taxifolia a los ataques de Chermes cooleyi incluyeron el cultivo de clones bajo condiciones uniformes en un invernadero especial, donde había gran posibilidad de infección por Chermes cooleyi. Los clones variaron mucho en susceptibilidad a juzgar por los depósitos de cera. Las ninfas se encontraron en 32 clones que variaban mucho en susceptibilidad a juzgar por los depósitos de cera. Solamente un clon permaneció inatacado a lo largo de los tres años que duró la experiencia (Petersen y Søegaard, 1958).
También ha sido estudiada la resistencia de especies frondosas a los ataques de insectos. Así, Hall (1942) afirma que durante su estudio sobre el perforador de la robinia (Cyllene robiniae), se observaron zonas donde ciertas variedades de Robinia pseudoacacia (black locust) con excelente forma parecían ser muy resistentes a los daños del perforador. Dos ejemplos destacados de esta resistencia aparente se dan en las robinias conocidas como «Shipmast»de Long Island y «Higbee» del sur de Indiana. En el último caso, la resistencia aparente ha sido comprobada durante un período de cuatro años por aplicación artificial a ciertos árboles de más de 2.000 larvas del perforador, de las cuales sólo 20 sobrevivieron hasta el estado adulto.
En los Populus señales claras de variación en resistencia por clones. Schreiner (1949) afirma que la variación extremadamente grande en susceptibilidad entre individuos híbridos de los mismos árboles genitores, tiene gran importancia para la mejora genética. Tales diferencias no podían esperarse para un insecto que, como el escarabajo japonés (Popillia japonica), se alimenta de numerosas especies de plantas. Si los resultados de 1947 fuesen confirmados durante los próximos años, se justificaría la mejora intensiva de la resistencia contra otros insectos forestales.
Los expuestos constituyen sólo unos pocos ejemplos de diferentes especies forestales que muestran algún grado de resistencia. Antes de pasar adelante será conveniente definir. qué es lo que se entiende cuando hablamos de «resistencia». Arboles resistentes son aquellos menos dañados o menos infestados que otros bajo condiciones de ambiente comparables. De acuerdo con Painter (1958), la resistencia puede ser debida a tres factores fundamentales:
1. Los árboles pueden no ser preferidos para la postura, cobijo o alimento.2. Pueden poseer un factor de resistencia llamado antibiosis, esto es, el árbol puede influir en la biología del insecto.
3. Los árboles resistentes pueden ser tolerantes, sobreviviendo bajo niveles de infestación que matarían o perjudicarían severamente a los árboles susceptibles.
En muchos de los ejemplos mencionados anteriormente han sido observadas variaciones en la resistencia a los insectos. Esta variación forma la base de la selección y mejora genética para obtener plantas capaces de una gran producción. Heimburger (1963) informa que los tipos de Pinus strobus de copa ancha sobreviven mejor que los de copa estrecha de crecimiento más lento, incluso cuando están muy atacados por Pissodes strobi. Esto es un ejemplo del grupo (3) de Painter, árboles «tolerantes». Heimburger op. cit., también observó diferencias relativas a la resistencia al ataque del gorgojo entre procedencias. Al desplazar los árboles desde el Sur hacia el Norte encontró que los árboles cuyo origen estaba situado al sur del lugar de plantación eran víctimas del gorgojo con más frecuencia e intensidad que los de origen local. Heimburger sacó en conclusión que el Pinus strobus aceptable en las plantaciones del este de Norteamérica, deberá ser. resistente a la roya vesicular Cronartium ribicola y a los ataques medianamente fuertes del gorgojo. La selección de individuos de copa estrecha y guías esbeltas, que son menos atractivas al gorgojo, ofrece posibilidades bajo condiciones de ataque moderado. El cribado por resistencia al ataque del gorgojo es más difícil y requiere más tiempo que el cribado por resistencia a la roya vesicular del pino blanco. Por esto, es más fácil escoger primero las plantitas en las almácigas por su resistencia a la roya vesicular y luego escoger el material superviviente por la resistencia al ataque del gorgojo.
Wright y Gabriel (1959) descubrieron diferencias en resistencia al gorgojo entre razas geográficas de Pinus strobus en un ensayo pequeño de procedencias en la zona de Tug Hill de Nueva York. Pauley, Spurr y Whitmore (1955) encontraron que tres procedencias de Pinus strobus de Ontario padecieron menos daños de Pissodes que los árboles oriundos de Massachusetts, Nueva Hampshire, o Nueva York y atribuyeron estas diferencias a la diversa intensidad de crecimiento.
Syrach Larsen (1956) demostró que el Larix decidua es susceptible a una especie de Chermes mientras que éste no se encuentra sobre Larix leptolepis. Del ensayo de especies e híbridos Callaham (1960) sacó la conclusión de que existía una variación racial hereditaria considerable de resistencia a los gorgojos dentro del Pinus ponderosa. Holst (1963) hace un reconocimiento de la susceptibilidad de las especies de Pinus de los grupos Lariciones¹ y Australes a daños por la mariposa europea del brote de pino, Rhyacionia buoliana, y por la mariposa de las puntas de Nantucket, Rhyacionia frustrana. En cada grupo se clasificaron las especies por la resistencia a estas mariposas. En la serie Lariciones, el Pinus nigra ofrece muchas posibilidades de proporcionar material muy resistente. El Pinus resinosa muestra una resistencia significativamente más baja que los otros pinos de la serie. En la serie Australes el pino palustre (Pinus palustris) y el pino «slash» (Pinus elliottii) son muy resistentes. Las especies con producciones altas de resina son muy resistentes a ambas mariposas, y dentro de una especie o grupo, los buenos productores de resina han mostrado más resistencia que los malos.
¹ Serie Lariciones = serie Sylvestres. La nomenclatura aquí seguida es la empleada por Wright (1962) en el Capítulo 3.
Syrach Larsen (1953) y Heitmüller (1954) han informado sobre probable resistencia completa a los ataques de Chermes en casos aislados de Pseudotsuga taxifolia. Petersen y Søegaard (1958) afirmaron la presencia de diferencias significativas entre ejemplares de Pseudotsuga taxifolia en la resistencia al ataque por el Chermes cooleyi. Un solo clon mostró inmunidad completa contra el ataque del Chermes, continuando en 1963 libre de ataques, a pesar de haber sido injertado en varios sitios de condiciones ambientales diferentes.
Aspecto importante en el problema de la resistencia es que un buen sistema de tratamiento de los árboles para un cierto país o localidad puede no ser satisfactorio para otro u otra, debido a cambios en la relación hospedante/insecto. Holst (1963) proporciona ejemplos de esto en su trabajo sobre posibilidades de mejorar genéticamente la resistencia de especies de Pinus a las mariposas Rhyacionia. La mariposa europea de los brotes de pino, Rhyacionia buoliana fue descubierta por primera vez en Canadá en 1915. El insecto está ahora estabilizado a lo largo de la isoterma mínima de invierno de - 6,7° C (+ 20° F), aunque en Europa se extiende por zonas con inviernos muy severos. El clima de verano con su influencia sobre la producción de resina e incubación de la plaga es importante tanto en Europa como en América del Norte. El estudio de Voute (1946) sobre la mariposa europea de los brotes de pino en el Pinus sylvestris en Holanda por un período de 40 años, pone de manifiesto grandes daños después de los años con mes de julio cálido, asociado normalmente con tiempo seco. En los años secos los árboles producen poca resina por lo que son incapaces de repeler las larvas de la mariposa. Neugebauer (1952) dio una explicación algo semejante para su mapa en el que siempre aparecieron las poblaciones de la mariposa europea de los brotes en zonas en que el período de mayo a junio fue seco. Las zonas con poblaciones constantemente altas de mariposa eran aquellas con bajas precipitaciones de verano, que causaban baja de turgencia y por consiguiente baja asimilación, producción de resina y presión de la savia.
Harris (1960) puso de manifiesto para el Pinus sylvestris del este de Inglaterra que en un verano cálido emergen los adultos, se aparean, ponen huevos, de éstos salen larvas, y las larvas se desarrollan hasta la tercera fase en cinco semanas, mientras que en un verano fresco un desarrollo equivalente requiere catorce semanas. Por este motivo en los veranos cálidos, las larvas en tercera fase atacan las yemas antes de que estén bien protegidas, mientras que en un verano fresco atacan cuando las yemas están protegidas por canales de resina bien desarrollados.
En cualquier estudio sobre resistencia se plantean siempre desde el principio los problemas que tienen que ver con la base o causa de la relación insecto/hospedante. A menudo la primera cuestión a contestar es si los árboles que sobreviven a los ataques severos son efectivamente resistentes. Para poder responder a esto es necesario ensayar tales árboles «resistentes» bajo condiciones reguladas para compararlos con árboles testigos. Una etapa posterior en el estudio de las causas de la resistencia consiste en el examen detallado de los factores diversos que intervienen en la relación insecto/hospedante. Unos pocos ejemplos pueden ilustrar la cuestión.
Miller (1950) se ocupó de la resistencia de las especies de Pinus al gorgojo de la reproducción del pino, Cylindrocopturus eatoni. Halló que se desarrollaba un tejido necrótico alrededor de la punción de la puesta de huevos en la corteza primaria, aislando este tejido. En los árboles que murieron, las pequeñas larvas atravesaron el tejido necrótico y una capa de corcho y alcanzaron al cambium. En los árboles supervivientes las larvas fueron incapaces de atravesar el tejido corchoso. Miller atribuyó la resistencia ya sea a esta capa que se desarrolla alrededor del tejido necrótico ya a alguna propiedad de la resina.
Salman y Bongberg (1942) y Keen y Salman (1942) idearon un sistema de graduación por riesgo para clasificar árboles maduros y supermaduros de Pinus ponderosa y P. jeffreyi de acuerdo con su susceptibilidad relativa al ataque del escarabajo de la corteza Dendroctonus. Los árboles clasificados de esta forma sirvieron para uno de los estudios más a fondo sobre el papel de la resina en la resistencia. La resina es considerada como un producto secundario de la planta, y Fraenkel (1959) opina, con gran razón, que los productos secundarios tienen mucha importancia para la selección, susceptibilidad y resistencia en las relaciones insecto/hospedante. Mergen et al. (1955) encontraron que ciertas características de la resina estaban condicionadas en alto grado por la genética.
La resina fué teóricamente encartada por primera vez en la resistencia del Pinus ponderosa al Dendroctonus monticolae por Hopkins (1902), pero en los 25 años siguientes se hizo muy poco para explorar esta posibilidad. Person (1931), trabajando con Pinus ponderosa y Dendroctonus brevicomis, sentó que el éxito de los primeros ataques a un árbol depende de la disposición del escarabajo para superar el flujo de resina. Gordon (1932) halló que ciertos constituyentes de la resina eran tóxicos para los escarabajos de la corteza, mientras Callaham (1955) mostró que el flujo de una herida tipo se prolongaba en árboles con pequeño riesgo (que lo tienen menor de morir por el ataque de los escarabajos de la corteza). Atribuyó el fallo de los ataques del escarabajo sobre los árboles de riesgo pequeño al flujo continuo de resina, pero no fue capaz de asociar el flujo total con la susceptibilidad de acuerdo con la clasificación por riesgo. Smith (1961a) ideó una técnica para determinar el efecto tóxico de los vapores saturados de las resinas frescas de pino en los escarabajos de la corteza adultos. Empleando un número limitado de especies de Pinus y tres especies de escarabajos de la corteza, formuló la hipótesis de que los Dendroctonus podían tolerar los vapores saturados de resina (esto es, los terpenos volátiles) de los pinos hospedantes, pero no de los pinos que no lo eran (Smith 1961b).
Hechas las observaciones relativas a la variación en la resistencia a ataques de insectos para una determinada especie, la cuestión inmediata es cómo seleccionar árboles que posean las cualidades deseadas. Un buen método ha sido descrito por Gerhold y Stroh (1963) para la selección de Pinus strobus por la resistencia al Pissodes strobi.
En sus consideraciones teóricas relativas al empleo de la selección directa o indirecta afirman que la última podría ser más efectiva que la primera en determinadas circunstancias. Si deseamos mejorar el carácter A, normalmente es mejor seleccionar directamente por A que no por el carácter B correlacionado con A; sin embargo, la selección indirecta podría ser más efectiva que la directa siempre que uno o varios de los factores siguientes compensase la incompleta correlación de B con respecto a A:
1. Menor esfuerzo o coste es necesario para seleccionar por el carácter B que por el A.2. Menor tiempo por generación es necesario para mejorar B que para A, como por ejemplo si la selección es posible en una edad más temprana.
3. Es mayor la precisión en el reconocimiento del fenotipo B que del fenotipo A, como puede ser el caso en ausencia de la plaga con suficiente intensidad.
4. Existe una mejor correlación de las medias de las descendencias con las medias parentales para el carácter B que para A.
Gerhold y Stroh recomiendan valorar las circunstancias anteriores dando el debido peso a cada una, en un programa particular de mejora genética y continúan: «La situación cuando hay que decidir entre la selección directa o indirecta por resistencia a los insectos ha sido desde luego simplificada excesivamente. La resistencia a los insectos es muchas veces una característica compleja en la que intervienen varios mecanismos (Painter, 1958). En realidad, pues, es probable que la selección indirecta debiera afectar a varias características y no solamente a la B. Descuidar las otras será aumentar el riesgo de que el insecto se adapte al material mejorado por una sola característica. Sin embargo, es indudable que la selección por varias características multiplica el trabajo, y pudiera uno preguntarse si la ganancia genética resultante de seleccionar características no sería sólo el 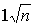 de la ganancia seleccionando por característica única. De hecho, la última objeción no es válida ya que la selección por la característica compleja A incluye indirectamente todos sus componentes y la selección actúa sobre una combinación de éstos cuya expresión es una mayor resistencia.»
de la ganancia seleccionando por característica única. De hecho, la última objeción no es válida ya que la selección por la característica compleja A incluye indirectamente todos sus componentes y la selección actúa sobre una combinación de éstos cuya expresión es una mayor resistencia.»
Grafius (1956, 1959), Grafius y Kiesling (1958), y Grafius y Adams (19,60), han desarrollado una técnica para el manejo de un número grande de caracteres, aunque no estén relacionados especialmente con la resistencia a los insectos. Se preparó un modelo basado en la acción cuantitativa de los genes para predecir las mejores combinaciones parentales para producir descendencias cuyas medias se aproximen a un ideal para un número grande de caracteres. Los caracteres complejos se descomponen en sus componentes para hacer mínimas las interacciones epistáticas. En apoyo de este nuevo método escribe Grafius (comunicación personal de febrero 1963) «Tenemos ahora datos sobre aproximadamente 50 cruces de cebada, y los datos concuerdan en forma excelente con las previsiones - esto para una docena de características diferentes».
Gerhold y Stroh (1963) discuten luego la selección directa y afirman que dos atributos del ataque de gorgojos hacen difícil la selección directa del Pinus strobus por resistencia al insecto. En primer lugar, el porcentaje de árboles dañados cada año es normalmente muy bajo sólo cuando la infestación es excepcionalmente fuerte excede del 50 por ciento. En segundo lugar, los casos atacados se distribuyen al azar o poco menos; estas características quitan mucha precisión a la selección. Proponen un procedimiento de selección preliminar del Pinus strobus, que consiste en una variante de una técnica propuesta por Wright y Gabriel (1959). Las masas o plantaciones en las que se ha de practicar la selección deberán ser bastante uniformes, de 5 a 12 metros de altura y estar muy atacadas por el gorgojo desde 6 años atrás por lo menos. Primeramente, se cuenta el número de casos de gorgojo por árbol y por año, en cada uno de 100 pies de árbol aproximadamente. Luego se determina el ritmo anual de casos y se calcula la distribución de frecuencias previstas, según el número de casos por árbol desarrollando la expresión binómica.
(p1 + q1) (p2 + q2).... (pn + qn)
en la cual p = probabilidad de ataque y q = probabilidad de escapar al ataque en cada año. Las distribuciones de frecuencia reales y previstas se comparan, y calculan por chi al cuadrado. La selección deberá practicarse sólo en las masas en las que la frecuencia de árboles no atacados es significativamente mayor que la prevista y en las que es alta la razón del número real al número previsto de árboles no atacados. Se discute la selección directa de otros pinos blancos que se considera impracticable en el nordeste, con la posible excepción del Pinus monticola.
En la discusión de la selección indirecta, Gerhold y Stroh (1963) afirman que varios caracteres están correlacionados con la resistencia del Pinus strobus al gorgojo, sirviendo de ejemplos tal como se mencionó anteriormente los árboles con corteza más delgada y guías menores que resultaron ser menos susceptibles, y la disminución de susceptibilidad por encima de un límite superior del diámetro de la guía. Los caracteres más prometedores, a partir de sus investigaciones, son el grosor de la corteza y la profundidad hasta los canales corticales de resina en la grúa. La medición de tres caracteres en muestras de corteza dio cuenta del 20 al 49 por ciento de la variación de los daños en diversas plantaciones. La obtención de estos datos requiere de 4 a 9 minutos por árbol, según su altura. Si valiese la pena el ensayo químico, éste requeriría alrededor de otros 20 minutos por árbol, además del coste del equipo de análisis y materiales. Se citan estas cifras como demostración de que la selección indirecta es varias veces más costosa que la selección directa, que requiere solamente de 1 a 3 minutos por árbol en muchos casos.
Basándose en los conocimientos limitados actuales parece que sería demasiado costoso sustituir la selección directa por la indirecta; sin embargo, Gerhold y Stroh proponen la utilización de un sistema mixto de selección en el cual se complementan entre sí las ventajas potenciales de ambos métodos.
En el estudio de la resistencia a los ataques de insectos el empirismo precedió a los ensayos de laboratorio. Sin embargo, la investigación llevada a cabo por Petersen y Søegaard (1958) se aproxima mucho a las condiciones de laboratorio. Colocaron juntos injertos de árboles seleccionados en una estufa especial, donde era posible dar a los injertos condiciones uniformes e ideales para el desarrollo de la plaga. Tales ensayos de «laboratorio x> alcanzan su máxima aplicación en los casos en que puede ser practicada la propagación vegetativa, pues entonces es posible combinar directamente las investigaciones de laboratorio con las pruebas en el campo.
La apreciación de la resistencia para árboles que se propagan por semilla ha sido discutida por Wright y Gabriel (1959) que proponen el siguiente programa duradero hasta el final de la segunda generación para la resistencia al Pissodes strobi: selección fenotípica de 500 individuos aparentemente resistentes; determinación de la aptitud combinatoria general de cada uno de estos 500 reproductores por ensayos 1-parentales de descendencia; determinación de la aptitud combinatoria específica de los 32 mejores reproductores por ensayos de descendencia 2-parentales dialelos; selección fenotípica en las plantaciones de ensayo F1; determinación de la aptitud combinatoria general de las 500 mejores plantas F1 por ensayos de descendencia 1-parentales; y determinación de la aptitud combinatoria especifica de los 32 mejores reproductores por ensayos de descendencia 2-parentales dialelos.
La investigación del origen de la resistencia ha precedido al estudio de su herencia y al trabajo de mejora genética, estrechamente relacionado con ella. Con respecto al trabajo de mejora para la defensa de los pinos contra el gorgojo de la reproducción del pino, Smith (1960) informa haber sido descubiertas 11 especies variedades o híbridos de Pinus resistentes entre 30 que fueron observadas en el lapso precedente de 15 años en que ha sido estudiada la relación del insecto con el hospedante. Encontró hechos que sugieren que el árbol genitor femenino tiene mayor influencia que el masculino en la transmisión de la resistencia a la descendencia. Los hallazgos de la investigación hasta la fecha muestran ser considerables las posibilidades de hibridación de los pinos para producir árboles resistentes a los gorgojos.
Otro ejemplo relativo al Pinus lo proporcionan los trabajos de mejora genética de Smith y Eaton (1963), quienes produjeron 2 híbridos de mucho interés, el P. jeffreyi X ponderosa y el P. jeffreyi X coulteri. En cada uno se trata de un cruce de especie no hospedante y hospedante con respecto a las dos especies, Dendroctonus brevicomis y D. monticolae. Los vapores de resina de estos híbridos fueron normalmente tóxicos a D. brevicomis pero no a D. monticolae.
Cuando el mejorador ha logrado producir árboles resistentes al ataque de insectos, su preocupación pasa a ser la posibilidad de reproducir en gran escala el material obtenido, para su empleo en el monte. Si la especie se propaga vegetativamente (como sucede con Populus y Cryptomeria japonica) el material obtenido puede utilizarse directamente por medio de injertos o estacas. Si la especie se propaga por semilla, un método para producir en masa el cultivar resistente es el establecimiento de huertos para semilla como se describen en los capítulos 7 y 10.
Smith y Eaton (1963) informan que el Servicio Forestal de los Estados Unidos comenzó hace 5 años un programa en gran escala para producir material resistente al gorgojo para empleo en el norte de California (véase Libby, 1958). El programa de mejora emplea híbridos F1 P. jeffreyi X coulteri y el híbrido natural. Se procede regularmente al cribado de gran número de Pinus jeffreyi por su compatibilidad con Pinus coulteri. Los ejemplares de Pinus jeffreyi destinados a la producción de híbridos son injertados en un huerto para subsiguiente polinización dirigida. Actualmente la producción anual de híbridos resistentes al gorgojo se cuentan por decenas de millares pero el objetivo es llegar a varios millones.
En los estudios sobre resistencia se concede gran importancia a la relación insecto/hospedante, así como al estudio de los dos factores encartados, el árbol y el insecto. Varios ejemplos mencionados anteriormente se referían a observaciones y estudios sobre los árboles y Smith y Eaton (1963) insisten sobre la importancia del insecto y han comenzado a trabajar sobre la capacidad variable del escarabajo de la corteza para atacar a los pinos, «... lo que podría ser tan importante como la capacidad variable del árbol para resistir a los escarabajos».
Gara (1963) estudió al Ips confusus y halló que su forma de vuelo observada desde la iniciación hasta que el insecto se posa, estaba influenciada por la velocidad y dirección del viento, intensidad de substancias de atracción, y cubierta del suelo. Las respuestas a las substancias de atracción tienen carácter inmediato, indicando que el ejercicio del vuelo no es un requisito previo para la respuesta en esta especie. La capacidad de respuesta a las substancias de atracción se vio que variaba de escarabajo a escarabajo, con el sexo, entre individuos de la misma puesta y entre poblaciones. Saca en conclusión que las diferencias en respuesta entre individuos y entre puestas son debidas a su condición fisiológica.
Vité et al. (1963) informan haber descubierto en sus estudios la existencia de una substancia volátil responsable de la atracción en masa del Ips confusus a las trampas, substancia obtenido de la parte posterior del tubo digestivo de los machos adultos, de sus excrementos y por condensación del aire extraído de recipientes conteniendo secciones de troncos infestados del escarabajo. Se vio que la substancia de atracción sólo era producida después de que el macho emergente había comido durante varias horas del nuevo material hospedante y el tubo digestivo, vacío durante el vuelo, se había llenado. En algunos de los experimentos de laboratorio las substancias parecían funcionar más por retención que por atracción.
Rudinsky (1963) ha estudiado la respuesta de Dendroctonus pseudotsugae a materiales volátiles emanantes de troncos infestados de Pseudotsuga taxifolia. Inmediatamente después de la emergencia de los insectos comenzó el vuelo de dispersión al azar que más tarde pasó a vuelo concentrado como respuesta a un factor de atracción causado por las primeras invasiones. La substancia atractiva ha sido concentrada en el laboratorio y cuando fue esparcida en el campo causó respuesta en el vuelo de la población. Esto sugiere su posible empleo en la prevención y lucha contra ataques violentos de Dendroctonus pseudotsugae.
La colaboración internacional ha sido establecida para el estudio de las enfermedades. Aunque en escala algo limitada, se ha iniciado la colaboración entre entomólogos forestales de Dinamarca, Finlandia, Noruega y Suecia. El número reducido de ocho investigadores ocupados por la entomología forestal en estos países nórdicos hace que tal colaboración sea de la máxima conveniencia para cada país, permitiendo al grupo acometer a veces proyectos de investigación a escala mucho mayor de lo que hubiese sido posible de trabajar aislados. Además, esta colaboración permite un mayor grado de especialización, el estudio de zonas geográficas y climáticas mucho mayores y redunda a menudo en un muestreo más representativo de la variable a investigar.
Si tal colaboración pudiera ampliarse de forma que, por un lado fuese posible al grupo mencionado cooperar con grupos semejantes establecidos en otras regiones geográficas de tamaño y volumen de trabajo razonables, y por otro permitiese a cada grupo, dentro de su región respectiva, organizar una colaboración más estrecha con los genetistas y mejoradores forestales de la región, se habría conseguido una colaboración internacional que reportaría grandes ventajas.
ANDERSON, J. M. & FISHER, K. C. 1956. Repellency and host specificity in the white pine weevil. Physiol. Zool., 29: 314324.
ANDERSON, J. M. & FISHER, K. C. 1960 .The response of the white pine weevil to naturally occurring repellents. Canada J. Zool., 38: 547-564.
AUSTIN, L., YUILL, J. S. & BRECHEEN K. G. 1945. Use of shoot characters in selecting Ponderosa pines resistant to resin midge. Ecol., 26 (3): 288-296.
BARNES, T. C. 1928. A biological study of the white pine weevil with special reference, to anatomy, flight, phenology, parasitism, behaviour, and injuries to young trees. Unpub. doctoral dies., Harvard Univ., Cambridge, Mass.
CALLAHAM, R. Z. 1955. Oleoresin production in the resistance of Ponderosa pine to bark beetles. Ph. D. thesis, Univ. of California, Berkeley, Calif. 120 p.
CALLAHAM, R. Z. 1960. Observations on pine susceptibility to weevils. U.S. Forest Serv. Pac. Southwest Forest and Range Expt. Sta. Tech. Paper 51. 12 p.
FOWLER, D. P. 1967. Discussion of tree breeding at Maple, Ontario. Proc. 5th Northeast. For. Tree Impr. Conf., p. 26.
FRAENKEL, G. S. 1959. The raison d'être of secondary plant substances. Science, 129 (3361): 1466-1470.
GARA, R. I. 1963. Studies on the fight behaviour of Ips confusus (Lec.) (Coleoptera: Scolytidae) in response to attractive material. Contrib. Boyce Thompson Inst., Jan-March.
GERHOLD, H. D. 1962. Testing white pines for weevil resistance. Proc. 9th Northeast. For. Tree Impr. Conf., 1961: 44-50.
GERHOLD, H. D. & STROH, R. C. 1963. Integrated selection for white pine weevil resistance and its components. FAO/FORGEN 63-6b/1.
GORDON, A. 1932. Tree injection experiments in white fir and ecological studies of insects attacking yellow pine. U.S. Dept. Agric., Bur. Ent., Forest Insect Lab., Berkeley, Calif. Unpub. Prog. 10 p.
GRAFIUS, J. E. 1956. Components of yield in oats: a geometrical interpretation. Agron. Jour., 48: 419-423.
GRAFIUS, J. E. 1959. Heterosis in barley. Agron. J., 51: 551-554.
GRAFIUS, J. E. & KIESLING, R. C. 1958. Vector representation of biologic fields of force. Agron. J., 50: 757-760.
GRAFIUS J. E. & ADAMS, M. W. 1960. Eugenics in crops. Agron. J., 52: 519-523.
GRAHAM, S. A. 1952. Forest entomology. New York, McGraw-Hill.
HALL, R. C. 1942. Control of the locust borer. U.S. Dept. Agric., Circ. 626. 19 p.
HALL, R. C. 1959. Field tests of the resistance of hybrid pinees to the pine reproduction weevil. U.S. Forest Serv., Pac. Southwest For. and Range Expt. Sta., Tech. Paper 33. 21 p.
HARRIS, P. 1960. Natural mortality of the pine shoot moth, Rhyacionia buoliana (Schiff.) (Lepidoptera: Olethreutidae), in England. Can. J. Zool., 38: 755-768.
HEIMBURGER, C. 1963. The breeding of white pine for resistance to weevil. FAO/FORGEN, 63-6b/5.
HEITMÜLLER, H. H. 1954. Beobachtungen über individuelle. Resistenz gegen Gilletteella cooleyi Gill. an Douglasie. Z. Forstgenet., 3: 99-100.
HOLST, M. J. 1955. An observation of weeril damage in Norway spruce. For. Res. Div., Tech. Note No. 4, Ottawa.
HOLT M. J. 1963. Breeding resistance in pines to Rhyacionia moths. Canada; Dept. of Forestry, Forest. Res. Branch Contr. (In Press)
HOPKINS, A. D. 1902. Insect enemies of the pine in the Black Hills forest reserve. U.S. Div. Ent. Bull. 32. 24 p.
KEEN, F. P. & SALMAN, K. A. 1942. Progress in pine beetle control through tree selection. J. For., 40 (11): 854-858.
KRIEBEL, H. B. 1954. Bark thickness as a factor in resistance to white pine weevil injury. J. For., 54 (2): 133-34.
LIBBY, W. J. 1958. The backcross hybrid Jeffrey X (Jeffrey X Coulter) pine. J. For., 56: 840-842.
MACALONEY H. J. 1930. The white pine weevil (Pissodes strobi Peck). Its biology and control. N. Y. State Coll. of Forestry, Tech. Publ. 28. 87 p.
MERGEN, F., HOEKSTRA P. E. & ECHOLS, R. M. 1955. Genetic control of oleoresin yield and viscosity in slash pine. For. Sci., 1 (1): 19-30.
MILLER, J. M. 1950. Resistance of pine hybrids to the pine reproduction weevil. U.S. Forest Serv., Calif. Forest and Range Expt. Sta. Res. Note 68. 17 p.
NEUGEBAUER, W. 1952. Die Bekämpfung des Kieferntriebwicklers. Forstarchiv, 23: 159-165.
PAINTER, R. H. 1958. Resistance of plants to insects. Ann. Rev. Entomology, 3: 267-290.
PAULEY, S. S., SPURR, S. H. & WHITMORE, F.W. 1955 .Seed source trials of eastern white pine. For. Sci., 1: 244-256.
PERSON, H. L. 1931. Theory in explanation of the selection of certain trees by the western pine beetle. J. For., 29 (5): 696699.
PETERSEN, B. B. & SOEGAARD, B. 1958. Studies on resistance to attacks of Chermes cooleyi (Gill.) on Pseudotsuga taxifolia (Poir.) Britt. Forstl. Foresogsv. Danm., 25: 37-45.
PREBBLE, M. L. 1951. Summary of investigations of attack by the white pine weevil in plots at the Petawawa Forest Exp. Sta. 1936-1948. For. Insect Lab. Sault St. Marie, Ontario.
RUDINSKY, J. A. 1963. Response of Dendroctonus pseudotsugae, Hopkins to volatile attractants. Contrib. Boyce Thompson Inst., Jan-March.
SALMAN, K. A. & BONGBERG, J. W. 1942. Logging high-risk trees to control insects in pine stands in northeastern California. J. For., 40 (7): 533-539.
SCHREINER, E. J. 1949. Poplars can be bred to order. U.S. Dept. Agric. Yearbook, p. 153-157.
SCHREINER, E. J. 1960. Objectives of pest-resistance improvement in forest trees and their possible attainment. Proc. 5th World For. Congr., Seattle, U.S.A., 2: 721-727.
SMITH R. H. 1960. Resistance of pines to the pine reproduction weevil Cylindrocepturus eatoni. J. Econ. Ent., 53: 144148.
SMITH, R. H. 1961. Techniques for determining the toxicity of pine resin vapors to Dendroctonus brevicomis and D. jeffreyi. J. Econ. Ent., 54 (2): 359-365.
SMITH, R. H. 1961b. The fumigant toxicity of three pine resins to Dendroctonus brevicomis and D. jeffreyi J. Econ. Ent., 54 (2): 365-369.
SMITH, R. H. & EATON, C. B. 1963. Studies on resistance of pines to insects. FAO/FORGEN, 63-66/2.
SULLIVAN, C. R. 1957. A biological study of the white pine weevil, Pissodes strobi Peck, with special reference to the EFFECT of physical factors on its actitiviy and behavior. Ph. D. thesis, McGill Univ.
SULLIVAN, C. R. 1961. The effect of weather and the physical attributes of white pine leaders on the behaviour and survival of the white pine weevil, Pissodes strobi Peck. in mixed stands. Canad. Ent., 53: 721-724.
SYRACH LARSEN, C. 1953. Studies of diseases in clones of forest trees. Hereditas, 39: 179-192.
SYRACH LARSEN, C. 1956. Genetics in silviculture. Edinburgh, Oliver and Boyd.
VITÉ, J. P., GARA, R. I. & KLIEFORTH, R. A. 1963. Collection and bioassay of a volatile fraction attractive to Ips confusus (Lec.) (Coleoptera: Scolytidea). Contrib. Boyce Thompson Inst., Jan-March.
VOUTE, A. D. 1946. De oorzaak van het optreden van gradaties van de dennenlotrups (Evetria buoliana [Schiff.]). en de mogelijkheid deze te voorkomen [Cause of the occurrence of the pine-shoot moth Evetria buoliana and the possibility of controlling it]. Ned. Boschb. Tijdschr., 18: 104-111, 128-135.
WRIGHT, J. W. & GABRIEL, W. J. 1959. Possibilities of breeding weevil-resistance white pine strains. Northeast. For. Expt. Sta. Paper 115. 35 p.
![]()
![]()
![]()