![]()
![]()
![]()
by
Alexandre Magno Sebbenn
Antonio Carlos Scatena Zanatto
Lígia de Castro Ettori
Omar Jorge Di Dio
Junior[24]
INTRODUCTION
The São Paulo Forest Institute, Brazil (SPIF) has since 1979 been conserving a number of Brazilian forest tree species which are endangered or subject to genetic depletion caused by the growing industrialization and unplanned agricultural and urban development. Vitor (1975) gives an idea of the worrying situation in the natural forests in São Paulo state, reporting that at the beginning of the nineteenth century, 70.5% of the area of the State was covered with natural forests but that in 1975 only 7% of these forests remained. The FAO Panel of Experts on Forest Gene Resources lists the following species of Brasil as being in need of attention: Aspidosperma polyneuron, Araucaria angustifolia, Balfourodendron riedelianum, Cariniana legalis, Dipterix alata, Hymenaea coubaryl, Myracrodruon urundeuva (FAO, 1998, 2000). An interesting historical reference is the one made in the middle of 1900 by the Swedish naturalist Albert Löfgren:“The devastation of the forests is destroying what took nature millions of years to create; the price of this destruction will be very high; in the name of life itself and our future, we need to halt this devastation “ (SPIF, 1994).
Today the lack of a forest policy to stimulate genetic conservation of the remaining forests, reforestation and recuperation of degraded areas and sustainable forest management is keenly felt. While in 1886, more than 70% of the Atlantic Forest remained, this area is now only 7%.
The ex situ genetic conservation program of native Brazilian species at SPIF aims to conserve the genetic variability existing among and within the populations of forest species, for present or future use, to meet economic social and environmental needs. Objectives of the program also include the study of the genetic structure, silvicultural behavior, heredity of traits of economic interest and improved seed production for reforestation of degraded and altered areas.
THE SÃO PAULO FOREST INSTITUTE
The SPIF is a branch of the Environmental Agency of the Government of the State of São Paulo, founded in 1888 by the Swedish naturalist Alberto Löfgren and was originally known as the Horto Botânico. In 1911 the Horto Botânico was transformed into the São Paulo State Forest Service and in 1970 it became the São Paulo Forest Institute (SPIF, 1994). The Institute has several ecological stations, parks, forests, experimental stations, nurseries and state forest reserves, distributed throughout the State of São Paulo, and occupying approximately 3.4% of the area[25]. Of the total area administered by the SPIF, 94% (0.8 million hectares) are used for conservation activities. The largest of the last remnants of Atlantic Forest in São Paulo State are found in these areas, providing in situ conservation and sources of seed and progagules for future rehabilitation of the natural forests of the State. The value of these areas is therefore incalculable. The SPIF also ensures ex situ conservation of Brazilian native and exotic forest species. Presently, the ex situ conservation programme includes 24 native forest tree species and 45 introduced tree species. The latter include 23 species of Eucalyptus, 14 species of Pinus and eight species of other genera: Liriodendron, Toona, Grevillea, Cupressus, Cryptomeria, Cordia and Araucaria (see Gurgel Garrido, 1997).
EX SITU CONSERVATION
The ex situ conservation activities were initiated by Dr. Ana Cristina Machado de Franco Siqueira, who was one of the first scientists to acknowledge the problems of accelerated loss of forest genetic resources in São Paulo State and Brazil and to take corresponding action.
Table 1 gives information on the native forest species included in ex situ conservation areas. These areas have been established since 1979. They include, however, also two provenance trials of Araucaria angustifolia established in 1973, which were subsequently incorporated into the ex situ conservation program. Presently, the program includes 24 species and 41 experimental areas. Some experiments are replicated in two sites. The ex situ areas include progeny and progeny/populations tests, initially established as pure plantations, following experimental designs such as randomized blocks, complete randomized blocks and compact family block design, always with six blocks and five plants per plot, totalling 30 plants per progeny/site. Since 1990, recognizing the importance of respecting the successional stages for better adaptation and growth of the majority of the native forest tree species in Brazil, the experiments have been established mixing several successional phase species in the blocks, including pioneers, secondary and climax species. The bracket with a number in table 1 indicates that this species is mixed with species with the same number in bracket.
Table 1: Ex situ genetic conservation experiments with native forest species in the SPIF, Brazil. Compiled by Gurgel Garrido (1997)[26]
|
Species
|
Experiment |
Establishment |
Number of provenances
|
Total number of plants
|
||
|
Type |
Design |
Year |
Sites |
|||
|
Anadenathera falcata |
pt |
IC |
1994 |
1 |
1 |
20 |
|
Araucaria angustifolia |
pt |
BC |
1974 |
2 |
1 |
32 |
|
Araucaria angustifolia |
Or |
BC |
1985 |
1 |
1 |
19 |
|
Araucaria angustifolia |
p/p |
BFC |
1973 |
2 |
5 |
115 |
|
Araucaria angustifolia |
pt |
BC |
1981 |
1 |
7 |
70 |
|
Araucaria angustifolia |
p/p |
BFC |
1980 |
1 |
15 |
121 |
|
Aspidosperma polyneuron |
pt |
BC (1) |
1992 |
1 |
1 |
23 |
|
Astronium graveolens |
pt |
BC |
1990 |
1 |
1 |
23 |
|
Balfourodendron riedelianum |
p/p |
BFC |
1984 |
1 |
3 |
48 |
|
Cariniana legalis |
p/p |
BFC |
1992 |
2 |
3 |
50 |
|
Cordia trichotoma |
pt |
BC |
1986 |
1 |
1 |
32 |
|
Cordia trichotoma |
pt |
BC |
1986 |
1 |
1 |
25 |
|
Dipteryx alata |
p/p |
BFC |
1980 |
1 |
2 |
38 |
|
Dipteryx alata |
p/p |
BFC |
1986 |
1 |
2 |
42 |
|
Dipteryx alata |
pt |
IC |
1981 |
1 |
1 |
25 |
|
Dipteryx alata |
pt |
BC |
1986 |
1 |
1 |
25 |
|
Enterolobium contortisiliquum |
pt |
BC |
1990 |
1 |
1 |
23 |
|
Esenbeckia leiocarpa |
pt |
BC (2) |
1992 |
1 |
1 |
25 |
|
Gallesia gorarema |
pt |
BC |
1985 |
1 |
1 |
25 |
|
Gallesia gorarema |
p/p |
CFB |
1982 |
2 |
3 |
65 |
|
Genipa americana |
pt |
BC (3) |
1997 |
1 |
1 |
21 |
|
Guazuma ulmifolia |
pt |
BC (3) |
1997 |
1 |
1 |
20 |
|
Hymenaea courbaryl |
p/p |
IC |
1985 |
1 |
3 |
68 |
|
Machaerium villosum |
p/p |
BC (1) |
1992 |
1 |
1 |
13 |
|
Machaerium villosum |
p/p |
CFB |
1981 |
1 |
4 |
26 |
|
Myracrodruon urundeuva |
pt |
BC (2) |
1993 |
1 |
1 |
23 |
|
Myracrodruon urundeuva |
p/p |
BFC |
1981 |
1 |
5 |
91 |
|
Myracrodruon urundeuva |
pt |
BC |
1986 |
1 |
1 |
42 |
|
Myracrodruon urundeuva |
pt |
BC |
1988 |
1 |
1 |
28 |
|
Myroxylon peruiferum |
p/p |
IC |
1984 |
1 |
4 |
45 |
|
Parapiptadenia rigida |
pt |
BC (1) |
1991 |
1 |
1 |
25 |
|
Peltophorum dubium |
pt |
BC (2) |
1992 |
1 |
1 |
26 |
|
Peltophorum dubium |
p/p |
BFC |
1985 |
2 |
2 |
28 |
|
Peltophorum dubium |
pt |
BC |
1985 |
1 |
1 |
18 |
|
Persea condata |
pt |
BC (1) |
1992 |
1 |
1 |
9 |
|
Prunus sellowii |
pt |
IC |
1989 |
1 |
1 |
17 |
|
Pterogyne nitens |
p/p |
BFC |
1989 |
1 |
4 |
36 |
|
Pterogyne nitens |
p/p |
BFC |
1989 |
1 |
5 |
93 |
|
Tabebuia heptaphylla |
pt |
BC |
1984 |
1 |
1 |
19 |
|
Tabebuia heptaphylla |
pt |
BC |
1985 |
1 |
1 |
15 |
|
Tabebuia vellosoi |
p/p |
BFC |
1986 |
1 |
1 |
18 |
RESULTS OF GENETIC STUDIES
Table 2 gives results from the analysis of genetic structure of conserved populations, estimated using methodology outlined by Hamrick (1976). The greatest part of genetic variability for all species was found within populations, notably among individuals within progenies. This is in agreement with most of the population structure studies carried out based on quantitative traits in tropical and temperate forest species (Hamrick 1976; Khalil, 1985; Li et al., 1993; Moraes, 1993; Buliuckas et al. 1999, Rehfeldt, 1999; Sebbenn et al. 1999), as well as with isoenzyme data (Hamrick & Godt, 1990). It is believed that the low genetic variation found among populations is associated with the small number of the populations of each species included in the experiments, and points to the need to include a greater number of populations in ex situ conservation banks to conserve the variability among populations. The absence of observed molecular level genetic variability among the population seems to indicate that, for tree breeding purposes, advantage in the selection of one or other population may be limited. If this observation is confirmed, selection of materials to be included in seed orchards can be made based on the performance of progenies across populations.
Genetic variability within the populations was also assessed, estimating heritability, the genetic variation coefficient, and phenotypic variation among the progeny. The heritability in height and diameter was generally low. Inconsistencies found in estimates of genetic parameters are possibly associated with the use of a low number of progeny per population, and flawed basic assumptions in quantitative genetics for the estimation of genetic parameters (see e.g. Nogueira et al., 1986b, Siqueira et al., 1993, Sebbenn et al., 2000). In this regard, Moraes (1993) studying the reproductive systems in populations of Myracrodruon urundeuva based on isoenzymes, observed that the outcrossing rate of the species was 0.49. Later it was found that this species has mixed reproductive behaviour (outbreeding, inbreeding) underlining the need for basic information on reproductive systems, as suggested by Ritland (1989) or Surles et al. (1990). Sebbenn (2000), when comparing the estimate of genetic parameters in Cariniana legalis populations using (i) the classical model, and (ii) incorporating principles of Ritland, observed over-estimates in the heritability coefficients of up to 33% when using the classical models. There is strong evidence from the literature that the majority of the native Brazilian forest species have mixed reproductive behaviour (see e.g. Paiva, 1993; Lepsch-Cunha, 1996; Sebbenn et al., 1998; Souza, 1997; Moraes, 1997; Oliveira, 2000). Thus, it is clear that genetic parameters in native forest species cannot be uncritically estimated based on the classical quantitative genetic models.
Table 2. Proportion of phenotypic variation within
progenies ( ), genetic variation among
progenies within populations (
), genetic variation among
progenies within populations ( ) and
genetic variation among populations (
) and
genetic variation among populations (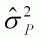 )
for some tropical tree species under ex situ conservation at SPIF,
Brazil.
)
for some tropical tree species under ex situ conservation at SPIF,
Brazil.
|
Species |
Trait |
Pop. |
|
|
|
Reference |
|
Balfourodendron riedelianum
|
DAP |
3 |
84.1 |
12.7 |
3.1 |
Siqueira et al. (2000)
|
|
ALT |
3 |
81.5 |
11.8 |
6.7 |
||
|
Pterogyne nitens |
DAP |
4 |
91.9 |
7.7 |
0.4 |
Sebbenn et al. (2000) |
|
ALT |
4 |
84.7 |
8.7 |
6.7 |
||
|
Peltophorum dubium |
DAP |
2 |
80.9 |
17.8 |
1.2 |
Sebbenn et al. (2000) |
|
ALT |
2 |
74.4 |
24.2 |
1.5 |
||
|
Cariniana legalis |
FF |
3 |
91.6 |
2.4 |
6.0 |
Sebbenn (2000) |
|
DAP |
3 |
96.5 |
3.4 |
0.1 |
||
|
ALT |
3 |
97.3 |
2.6 |
0.1 |
||
|
VC |
3 |
85.7 |
1.5 |
12.8 |
The Atlantic Forest of Brazil is one of the greatest pools of forest biological diversity on the planet, and many areas contain more than 400 tree species per hectare; 90% of the species are found at frequencies of only one individual per hectare (Kageyama et al, 1997). However, as reported above, the SPIF ex situ conservation areas include only 24 species, and less than five populations have been included for most species. All the ex situ areas and experiments were set up with resources from the SPIF but there has been a gradual reduction in the financial investments from the State Government in the institution and the inclusion of new species has been drastically reduced in recent years: 73% of the 41 experiments were established before 1990.
There is a need to increase the number of populations of the species already established in the ex situ stands to improve the coverage of genetic variation conserved. There is, further, a need to continue to assess, periodically, the material contained in the ex situ conservation banks, to gain better understanding of the genetic characteristics and silvicultural behavior of the species. Thirdly, there is a need to consider transforming the experiments into seed orchards, using low intensity selection to maintain large effective population sizes. Finally, there is a need to increase the number of species under ex situ conservation, and to include a range of populations of each new species included. A sample of 30 populations, each represented by 25 progenies and 30 plants per progeny established in at least three localities, would likely meet conservation objectives and to guarantee the conservation also of rare alleles (frequency < 5%).
New candidate species to be included have been identified based on two criteria: (i) occurrence in São Paulo State, and (ii) economic value. These species are listed in table 3 according to the successional groups they belong to.
Table 3: List of new candidate species to be included in the ex situ conservation program.
|
Species |
Common name |
Family |
|
Pioneer species |
|
|
|
Cecropia glaziovi Snethl. |
Embaúbavermelha |
Cacropiaceae |
|
Cecropia hololeuca Miquel |
Imbaúba prateada |
Cacropiaceae |
|
Clethra scabra Pers. |
Carne de vaca |
Clethraceae |
|
Croton floribundus Spreng |
Capixingui |
Euphorbiaceae |
|
Piptadenia paniculata Bentham |
Angico |
Mimosaceae |
|
Secondary species |
|
|
|
Cabraela canjerana |
Canjarana |
Meliaceae |
|
Cariniana estrellensis |
Jequitibá branco |
Lecithidacea |
|
Cedrela fissilis Vellozo |
Cedro |
Meliaceae |
|
Centrolobium robustum (Vell.) Martius ex Bentham
|
Araribá rosa |
Fabaceae |
|
Centrolobium tomentosum Guillem ex Bentham
|
Arauva |
Fabaceae |
|
Chlorophora tinctoria (Linnaeus) Gaudichaud
|
Taiúva |
Moraceae |
|
Chorisia speciosa (A. S. Hil.) Dawson |
Paineira |
Bombacaceae |
|
Cinnamomum glaziovii (Mez) Kosterm |
Garuva |
Lauraceae |
|
Copaifera langsdorffii Desf. |
Bálsamo |
Caesalpinaceae |
|
Copaifera trapezifolia Hayne |
Copaíba |
Caesalpinaceae |
|
Cryptocarya moschata Nees & Martius ex Nees
|
Canela branca |
Lauraceae |
|
Dalbergia brasiliensis Vogel |
Jacaranda |
Fabaceae |
|
Erytrina falcata Benth. |
Corticeira |
Fabaceae |
|
Inga striata |
Inga |
Mimosoidae |
|
Joannesia princeps Vell. |
Boleira |
Euphorbiaceae |
|
Lafoensia pacari A. Saint-Hilaire |
Dedaleiro |
Lythraceae |
|
Luehea divaricata Martius & Zuccarini
|
Açoito a cavalo |
Tiliaceae |
|
Machaerium scleroxylon Tulasne |
Caviuna |
Fabaceae |
|
Nectandra lanceolata Nees et Matius ex Nees.
|
Canela-amarela |
Lauraceae |
|
Platypodium elegans Vog. |
Amendoin do campo |
Fabaceae |
|
Podocarpus lambertii Klotzsch ex Endl.
|
Pinheiro bravo |
Podocarpaceae |
|
Talauma ovata A. Saint-Hilaire |
Baguaçu |
Magnoliaceae |
|
Climax species |
|
|
|
Calophyllum brasiliensis Combessedes |
Guanandi |
Clusiaceae |
|
Casealpinia leiostachya Ducke |
Pauferro |
Caesalpiniaceae |
|
Dalbergia nigra (Vell.) Freire Allemão ex
Bantham |
Jacaranda da Bahia |
Fabaceae |
|
Holocalyx balansae Micheli |
Alecrim |
Fabaceae |
|
Laplacea fruticosa (Schrader) Kobuski |
Santa Rira |
Theaceae |
|
Ocotea catharinensis Mez |
Canela-preta |
Lauraceae |
|
Ocotea odorifera (Vell.) Rohwer |
Canela-sassafrás |
Lauraceae |
|
Ocotea porosa (Nees ex Martius) Libarato Barroso
|
Imbuia |
Lauraceae |
|
Plathymenia reticulata Benth. |
Amarelinho |
Mimosaceae |
|
Tabebuia impetiginosa |
Ipê-rosa |
Bignoniaceae |
The authors thank the heads/directors of the SPIF Experimental Stations who have carried out the maintenance of the conservation banks; and the field assessment teams, who have ensured high quality and efficiency in data collection. We are also grateful to Girlei Costa da Cunha, Forest Engineer for reading the manuscript, and for valuable suggestions of new species to be included in the SPIF conservation bank.
REFERENCES
Buliuckas, V.; Ekberg, I.; Eriksson, G. & Norell, L. Genetic variation among and within populations of four Swedish hardwood species assessed in a nursery trial. Silvae Genetica, Frankfurt, 48(1): 17-25, 1999.
Ettori, L.C.; Siqueira, A.C.M.F.; Nogueira, J.C.B.; Ferreira, A.B. & Zanatto, A.C.S. Conservação ex situ dos recursos genéticos do Ipê-amarelo (Tabebuia vellosoi Tol.) através de testes de procedências e progênies. Rev. Inst. Flor., São Paulo, 7(2): 157-168., 1995.
Ettori, L.C.; Siqueira, A.C.M.F.; Sato, A.S. & Campos, O.R. Variabilidade genética em populações de Ipé-roxo - Tabebuia heptaphylla (Vell.) Tol. - para conservação ex situ. Rev. Inst. Flor., São Paulo, 8(1): 61-70, 1996.
Ettori, L.C.; Siqueira, A.C.M.F.; Zanatto, A.C.S. & Boas, O.V. Variabilidade genética em duas populações de Cordia trichotoma. Rev. Inst. Flor., São Paulo, 11(2): 179-187, 1999.
FAO. 1998. FAO Panel of Experts on Forest Gene Resources: Tenth Session, Rome, Italy, 9-11 September 1997. FAO, Rome, 68 p.
FAO. 2000. FAO Panel of Experts on Forest Gene Resources: Eleventh Session, Rome, Italy, 29 September to 1 October 1999. FAO, Rome, 90 p.
Giannotti, E.; Timoni, J.L.; Mariano, G.; Coelho, L.C.C.; Fontes, M.A. & Kageyama, P.Y. Variação genética entre procedências e progênies de Araucaria angustifolia (Bert.) O. Ktze. In: Congresso National sobre Essëncias Nativas, Campos do Jordão, 1982. Anais... Silvicultura em São Paulo, São Paulo, 16A: 970-975. Pt. 2. (Edição especial).
Gurgel Garrido, L.M.A.; Siqueira, A.C.M.F.; Cruz, S.F.; Romanelli, R.C.; Ettori, L.C.; Crestana, C.S.M.; Silva, A.A.; Morais, E.; Zanatto, A.C.S. & Sato, A.S. Programa de melhoramento genético florestal do Instituto Florestal de São Paulo (Acervo). IF Série Registros, São Paulo, 18: 1-53, 1997.
HamricK, J.L. & Godt, M.J.W. Allozyme diversity in plant species. In: Brown, A.H. D.; Clegg, M.T.; Kahler, A.L. & Weir, B.S. (Eds) Plant population genetics, breeding and genetic resources. Sinauer, Sunderland, Massachusetts, p. 43-63, 1990.
HamricK, J.L. Variation and selection in western montane species II. Variation withing and between populations of White Firon an elevation transect. Theor. Appl. Genet, 47: 27-34, 1976.
IFSP. Intituto Florestal de São Paulo. São Paulo, Brasil, IF-Série Registros No. 12, 1994, 31 p.
Kageyama, P.Y. Gandara, F. & Borges, A. Pesquisa e recuperação de áreas degradadas na Mata Atlântica - Avaliação bibliográfica. In: Recuperação de áreas degradadas na Mata Atlântica. CNBR, São Paulo, p. 11 a 16. 1997.
Khalil, M.A. Genetics of wood characteres of black spruce [Picea mariana (Mill.) B.S.P.] in Newfoundland, Canada. Silvae Genetica, Frankfurt, 34(6): 221-230, 1985.
Lepsch-Cunha, N. Variabilidade Genética Intrapopulacional de Duas Espécies de Baixa Densidade na Amazônia Central: Couratari multiflora (J.E. Smith) Eyma e Couratari guianensis Aublet. Item 1. de Resultados da Dissertação “Estrutura Genética e Fenologia de Espécies Raras de Couratari spp. (Lecythidaceae) na Amazônia Central”. ESALQ/USP, Piracicaba, SP. 1996, 121p. (Dissertação de mestrado)
Li, P.; Beaulieu, J.; Corriveau, A. & Bousquet, J. Genetic variation in juvenile growth and phenology in a White Spruce provenance-progeny test. Silvae Genetica, Frankfurt, 42 (1): 52-60, 1993.
Moraes, M.L.T. Variabilidade genéica por isoenzimas e caracteres quantitativos em duas populações naturais de aroeira Myracrodruon urundeuva F.F. & M.F. Allemão Anacardiaceae (Syn: Astronium urundeuva (Fr. Allemão) Engler. ESALQ/USP, Piracicaba, SP, 1993, 139 p. (Tese de doutorado)
Moraes, P.L.R. Estrutura genética de populações de Cryptocarya moschata Nees & Martius ex Nees (Lauraceae). UNESP, Rio Claro, SP, 1997, 190 p. (Tese de doutorado)
Mori, E.S. Estrutura genética de populações narurais de canafístula Peltophorum dubium (Spreng.) Taub. UNESP, Botucatu, São Paulo, 145 p. 2000. (Tese de livre docência).
Nogueira, J.C.B.; Siqueira, A.C.M.F. & Zanatto, A.C.S. Teste de progênies e procedências do pau d”alho Gallesia gorarema Vell. Moq. Boletim Técnico do IF, São Paulo, 40A: 344-356, 1986a.
Nogueira, J.C.B.; Siqueira, A.C.M.F.; Moares, E. & Ywane, M.S.S. Teste de progênies e procedências da aroeira Astronium urundeuva (Fr. All.) Engl. Boletim Técnico IF, São Paulo, 40A: 367-377, 1986b.
Oliveira, A.F. Estrutura genética de populações naturais de Copaifera langsdorffii Desf. a partir de isoenzimas. UFLA, Lavras, MG, 2000, 115 P. (Dissertação de mestrado)
Paiva, J.R. Variabilidade isoenzimática em populações naturais de seringueira [Hevea brasiliensis (Willd) ex Adr. de Juss.] Müell. Arg. ESALQ/USP, Piracicaba, 1992, 144 p. (Tese de doutorado)
Rehfeldt, G.E. Systematics and genetic structure of Washoe Pine: Applications in Conservation genetics. Silvae Genetica, Frankfurt, 48(3-4): 167-173, 1999.
Ritland, K. Correlated matings in the partial selfer Mimulus guttatus. Evolution, 43(4): 848-859, 1989.
Sebbenn, A. M.; Durigan, J.; Siqueira, A. C. M. F.; Pontinha, A. A. S. Teste de procedências de Grevilha robusta A. Cunn.. Rev. do Inst. Flor, SP, 11(1): 65-73, 1999.
Sebbenn, A. M.; Kageyama, P. Y. & Vencosvky, R. Variabilidade genética, sistema reprodutivo e estrutura genética espacial em Genipa americana L. através de marcadores isoenzimáticos. Scientia Forestalis, 53: 15-30, 1998b.
Sebbenn, A. M.; Siqueira, A. C. M. F.; Kageyama, P. Y. & Di Dio J., O. P. Estrutura genética de Pterogine nitens Tul. (Caesalpinaceae) através de caracteres quantitativos. Scientia Forestalis, Piracicaba, São Paulo, 2000. (no prelo)
Sebbenn, A.M. Estudo de populações de Cariniana legalis (Mart.) O. Ktze, por caracteres quantitatívos e isoenzimas. ESALQ/USP, Piracicaba, SP, 209 p. 2000. (Tese de doutorado)
Sebbenn, A.M.; Siqueira, A.C.M. F.& Gurgel Garrido, L. M. A. Interação progênies x locais e variabilidade genética em Jequitibá-Rosa - Cariniana legalis (Mart.) O. Ktze. Rev. do Inst. Flor. SP, 12(1): 13-23, 2000.
Sebbenn, A.M.; Siqueira, A.C.M.F. Vencovsky, R. & Machado, J.A.R. Interação genótipo ambiente na conservação ex situ de Peltophorum dubium, em duas regiões do Estado de São Paulo. Rev. Inst. Flor, SP, 11(1): 75-89, 1999.
Siqueira, A.C.M.F.; Nogueira, J.C.B. & Kageyama, P.Y. Conservação de recursos genéticos ex situ de cumbaru (Dipteryx alata Vog.) - Leguminosae. Rev. do Inst. Flor., São Paulo, 5(2): 231-43, 1993.
Siqueira, A.C.M.F.; Sebbenn, A.M.; Garrido, L.M.A.G.; Castollen, M.S. & Zanatto, A.C.E. Comportamento silvicultural e genético de duas espécies arbóreas tropicais secundárias. Rev. do Inst. Flor., SP, 11(1): 53-64, 1999.
Siqueira, A.C.M.F.; Sebbenn, A.M.; Kageyama, P.Y. & Ettori, L.C. Distribuição da variação genética entre e dentro de populações de Balfourodendron riedelianum (Engler) Engler para a conservação ex situ. Rev. do Inst. Flor, São Paulo, 2000. (no prelo)
Souza, L.M.F.I. Estrutura genética de populações naturais de Chorisia speciosa St. Hil. (Bombacaceae) em fragmentos florestais na região de Bauru (SP) - Brasil. ESALQ/USP, Piracicaba, SP, 1997, 76 p. (Dissertação de mestrado).
Surles, S.E.; Arnold, J.; Schnabel, A.; Hamrick, J.L. & Bongarten, B.C. Genetic relatedness in open-pollinated families of two leguminous tree species, Robinia pseudoacacia L. and Gleditsia triacanthos L. Theor. Appl. Genet, 80: 49-56, 1990.
Victor, M. A devastação florestal. Sociedade Brasileira de Silvicultura, São Paulo, 28-32, 1975.
![]()
![]()
![]()
 (%)
(%)