![]()
![]()
![]()
Helmut Lieth
HELMUT LIETH, an ecologist specialized in global patterns of primary productivity and seasonal ecosystems, is a professor at the University of North Carolina in the United States.
Converting tropical forests into crop lands is more tempting than ever for agriculturists. But the author's studies indicate that moving from temperate to tropical zones causes a decrease in biological productivity. He supports his arguments with a series of predictive computer models.
Natural resources in the temperate regions are being exploited toward their limits. It is therefore logical that the world economy is turning to the developing countries of the tropics to see what untapped resources are still available and how they can be utilized.
Experience in Africa and South America shows that this is not so easy, especially in the large humid areas of the tropics with soils that originated from sedimentary rock - conditions which apply to large parts of the Amazon basin in Brazil.
A wise plan for using the resources of the tropics and the development of appropriate management models for sustaining yield are therefore urgently needed.
A fundamental input in such models is the primary (biological) productivity pattern. Present knowledge of productivity patterns is best in the temperate zone and poorest in tropical areas. Productivity studies in the tropical rain forest area are therefore highly desirable.
The table of net primary productivity of major vegetation units lists the most likely productivity levels tin different parts of the world (see page 30). It shows productivity values averaged for major vegetation formations. The formations are distinct in species composition and structure of their vegetation body. Column 1 gives types of vegetation, column 2 the size of the area, column 3 its location in or outside the tropics, column 4 its approximate mean annual productivity level per square metre, and column 5 the annual average total productivity for that entire area.
The Amazon area is covered mainly by tropical rain forest, tropical grassland, lakes and streams. Compared with other vegetation areas (see table) it must be regarded as being endowed with above average to outstandingly high natural productive power. This is demonstrated all the more in cartographic evaluation of net primary productivity in the world. Using a combination of direct and indirect productivity assessment methods, we have constructed a computer map of the world's net primary productivity patterns (Lieth, 1973). This map is known as the Miami Model (Figure 1).
A summary of productivity for tropical ecosystems was recently compiled by Murphy (1975). His collection is shown in Figure 2. These figures are based on actual measurements. They show the wide range of productivity levels possible in each tropical biome type. Although most of these figures have been acquired outside of South America, their levels can be assumed to be representative of similar vegetation types in the Amazon basin.
One can assess in this way the productivity of small areas reasonably accurately. Such measurements are needed in sufficient quantities to construct correlation models with environmental parameters, as used to construct the world map (Figure 1). However, the assessment of natural productivity patterns in such direct form is very difficult in most tropical forest areas. Methods of indirect assessment are therefore needed to obtain a reasonable picture of the productive capacity of the Amazon area in the time allowed for its use by planners. But even before we can do much indirect modelling of productive capacity, we need some basic data to work with. Unfortunately, for the Amazon basin such data are almost completely lacking and their gathering is delayed by a variety of circumstances. Therefore, when one elaborates on the productivity of this area and the possible effects of human interference, one must rely on deductions from measurements in other areas of the world.
When agricultural systems are developed in areas where natural ecosystems prevailed previously, one automatically assumes that the manmade system is more useful and at the same time more productive. According to our recent findings, however, there is reason to believe that this is not necessarily true.
PHILIPPINE FORESTER AT AN FAO COMPUTER where data is scarce, computers provide new dimensions
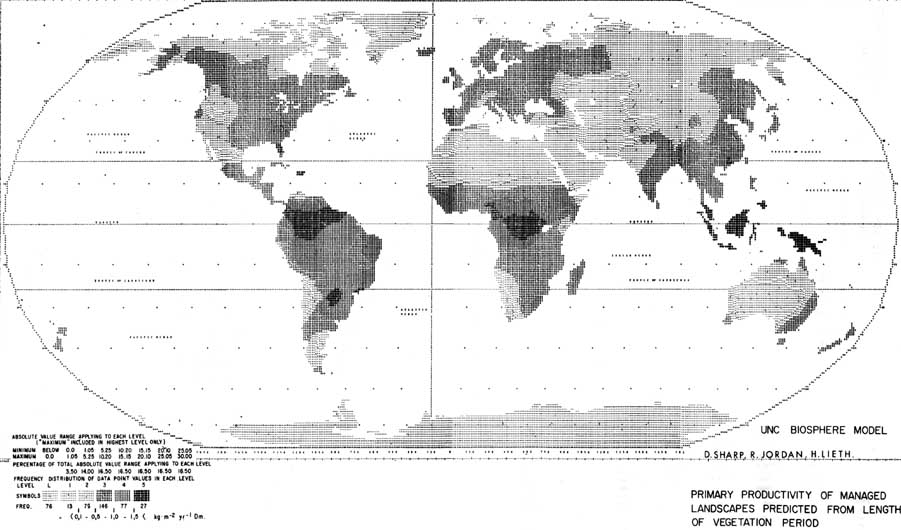
Together with a number of colleagues in the Eastern Deciduous Forest Biome of the U. S. International Biological Programme (US-IBP) we analysed the net primary productivity of individual states like Wisconsin, New York, Tennessee and North Carolina. The results of these studies were evaluated by Reader (1973) to see how they correlate to the length of the growing period (Figure 3). The outcome of this study revealed to our surprise that managed landscapes seem to become less productive in comparison to natural landscapes the further south we are.
Comparisons of statistical assessments of the productivity of total areas vs. careful measurements of natural and seminatural plant communities remaining in that same area serve as a basis for this statement (Sharp et al., 1975). For most parts of the world, such comparisons are impossible for lack of basic data. But using computer models we can make such comparisons in order to present our concern for the need of careful studies in tropical areas.
Figure 3 shows the comparison of two models predicting net primary production from the length of the vegetation period. We compare two linear regressions, the one based upon the production assessment of managed landscapes in the United States and the other based on data from the entire world, mostly taken in natural ecosystems. Comparison of the two lines allows the following conclusion: if the trend indicated by the North American studies is valid for the entire world, global productive capacity will drop more than 30 percent, with the greatest loss occurring in tropical humid regions. In order to demonstrate the changes in the world pattern we have prepared special computer maps similar to the one shown in Figure 1.
Figure 4 shows the map constructed from nearly a thousand points of known length of vegetation period.
The map has been based on the regression calculated by Reader (1973) for managed landscapes in eastern North America and extrapolated for the whole world. This map is known as the Hague Model.
Figure 5 shows the computer map based on the regression line for natural vegetation, using the same thousand points of known vegetation period. We call this map the Manaus Model.
It is easy to see that the Manaus Model yields productivity levels similar to those of the Miami Model (Figure 1), whereas the Hague Model yields far lower values, especially in tropical regions.
The conclusions one can draw from such preliminary research is that great care should be taken in converting the present medium to very high productive systems in the Amazon areas into agricultural systems, because there is danger of losing much of the productive potential.
The productivity of an agricultural system depends in large measure on adequate fertilizer input. In contrast to natural systems, where the essential elements circulate efficiently, modern agricultural crops depend almost entirely on fertilizer subsidies. This is the reason why many developing countries produce on the average far lower crop yields per ha than can be expected from the photosynthetic capacity inherent in the plants.
Present knowledge of productive patterns is best in temperate zones and poorest in the tropics
NET PRIMARY PRODUCTIVITY OF MAJOR VEGETATION UNITS OF THE WORLD
|
Vegetation unit |
Area 106 km² |
Net primary productivity |
Total for area 109 metric tons |
||
|
Range kg m-2 yr-1 |
Aproximate mean kg m-2 yr-1 |
||||
|
1 |
2 |
3 |
4 |
5 |
6 |
|
Forests |
50 |
|
|
|
81.6 |
|
Tropical rain forest |
17.0 |
T |
1 - 3.5 |
2.8 |
47.4 |
|
Raingreen forest |
7.5 |
T |
1.6 - 2.5 |
1.75 |
13.2 |
|
Summergreen forest |
7.0 |
t |
0.4 - 2.5 |
1.0 |
7.0 |
|
Mediterranean sclerophyll |
|
|
|
|
|
|
forest (chaparral) |
1.5 |
t |
0.25 - 1.5 |
0.8 |
1.2 |
|
Warm temperature mixed |
|
|
|
|
|
|
forest |
5.0 |
t |
0.6 - 2.5 |
1.0 |
5.0 |
|
Boreal forest |
12.0 |
t |
0.3 - 1.2 |
0.65 |
7.8 |
|
Woodland |
7 |
|
0.2 - 1.0 |
0.6 |
4.2 |
|
Dwarf and open scrub |
26 |
|
|
|
2.6 |
|
Tundra |
8.0 |
t |
0.06 - 1.3 |
0.16 |
1.3 |
|
Desert scrub |
18.0 |
Tt |
0.01 - 0.25 |
0.07 |
1.3 |
|
Grassland |
24 |
|
|
|
19.2 |
|
Tropical grassland (including grass-dominated savanna) |
15.0 |
T |
0.2 - 2.9 |
0.8 |
12.0 |
|
Temperate grassland |
9.0 |
t |
0.07 - 1.3 |
0.8 |
7.2 |
|
Desert (extreme) |
24 |
|
|
|
19.2 |
|
Cry desert |
8.5 |
T |
0 - 0.01 |
0.003 |
- |
|
Ice desert |
15.5 |
t |
0 - 0.001 |
- |
- |
|
Cultivated land |
14 |
|
0.1 - 4.0 |
0.65 |
9.1 |
|
Fresh water |
4 |
|
|
|
5.0 |
|
Swamp and marsh |
2.0 |
Tt |
0.8 - 4.0 |
2.0 |
4.0 |
|
Lake and stream |
2.0 |
Tt |
0.1 - 1.5 |
0.5 |
1.0 |
|
Total for continents |
149 |
|
|
|
121.7 |
T = tropical; t = temperare.
To demonstrate this fact we show in Figure 6 the assessment of yield and productivity figures for several areas. The work was carried out by Terjung et al. (in press). They used the energy budget to predict the productive potential of particular crops and compared these with yield data from FAO statistics. The example shown in Figure 6 is wheat, for which 30 percent of total productivity is assumed to be yield. The prediction is plotted in Figure 6A against the observed average yield. As one can see, predicted yield is greater than observed yield in most countries. According to Mitscherlich's yield law, crop yield may be expected to depend on the amount of fertilizer added to the system. Using the FAO statistics again, we can calculate the yield to be expected in each country according to the amount of fertilizer its agriculture is accustomed to apply.
Using such countries, where in Figure 6A the predicted and observed yields are almost equal as a base for the determination of maximum yield levels, one can derive a curvilinear regression, as presented in Figure 6B. The curve depicts the well-known economic law of diminishing return. This may be applied as a correction to the prediction presented in Figure 6A. The result is shown in Figure 6C. The linear regression between prediction and observation is now very good. The conclusion is that predictable high yields in tropical areas can only be attained by high nutrient inputs.
This fundamental need will probably cause a variety of environmental problems for the Amazon area. Together with the increased loss of soil from erosion, it should result in great changes in the ecology of the rivers. Black-water areas may change into white-water streams. Increased fertilizer leakage from fields will result in eutrophication. Such modifications may be expected to result in big changes in fish and invertebrate populations in the affected rivers Distribution patterns of disease vectors may also be expected to change and cause shifts of epidemic diseases, also affecting areas hitherto considered safe.
While we realize that certain areas of the Amazon forests need to be converted to agricultural use, one can only warn planners and developers to, first, take all necessary precautions to destroy as little as possible of the natural areas until their management for sustained yield is sufficiently understood, and second, to start development in areas that promise the best results. As an example we want to discuss the problem of phosphate fertilization.
It is common knowledge that soils vary in their suitability for agriculture, the major factors being natural fertility, response to fertilizer, and readiness for low-cost mechanization. Other important factors, such as distance to consumer centres, are also likely to play a role in the near future because climatic factors are against the development of a dense urban network in the Amazon area.
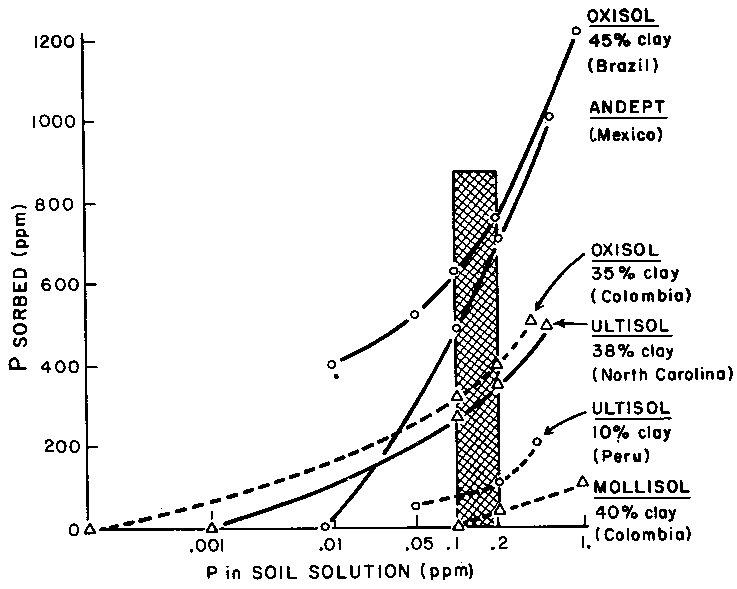
With regard to fertilizers, phosphate is probably the most difficult factor. World phosphate supply is limited. The need for phosphate varies for the same crop in different soils.
Figure 7 shows a diagram (provided by P. Sanchez from N.C. State University) of phosphate availability in different soils (see also Sanchez and Buol, 1975). From the phosphate added (ordinate) to a unit volume of soil, only a certain percentage can be retrieved by the plant in the soil solution. Since the average crop plant requires everywhere sufficient phosphate for its best growth (0.1-0.2 ppm on the abscissa), it is obvious that. different soils must show differential success for crops of high demands on fertilizer. The graph shows the sequence of theoretical suitability for cropping with regard to minimum phosphate supply needed.
FIGURE 6. - Analysis of global and potential wheat yield by countries
Different crops make different demands on environmental sources. Most species developed in temperate-zone agriculture have high fertilizer demands. It would probably be very advantageous if regional governments in the Amazon area directed their research stations to look for native or new crop plants with minimum demand for fertilizer for a given starch, oil or sugar yield. Future conditions may well demand more ingenuity from food technologists to supply taste, texture, and final chemical composition of marketed products, the kind of crop properties which are now requested from the farmer.
FIGURE 7. - Phosphate availability in different soils
To sum up, we may list the following points:
1. The natural productivity of the humid tropics is among the highest in the world.2. Conversion of natural vegetation in these areas into crop land may reduce the productivity level substantially if present agricultural practices are used.
3. Demand for fertilizer will increase enormously, probably causing a variety of environmental problems in some of the major rivers.
4. Fertilizer requirements and suitability of soils and landscapes in the Amazon area vary greatly. Government planning boards should work out priorities for development, choosing low-risk areas for development and imposing moratoriums on others.
5. Native species and varieties best suited for production of staple food in the tropics should be developed, rather than forcing temperate-zone plants into these areas.
LIETH, H. 1973 Primary production: terrestrial ecosystems. Human Ecology, 1: 303-332.
LIETH, H. 1975 The primary productivity in ecosystems. Comparative analysis of global patterns. In van Dobben, W. H. and Lowe-McConnel, R.H., eds. Unifying concepts in ecology The Hague, Junk.
LIETH, H. & WHITTAKER, R.H., eds. 1975 The primary productivity of the biosphere. New York, Springer Verlag.
MURPHY, P.G. 1975 Net primary productivity in tropical terrestrial ecosystems. In Lieth, H. and Whittaker, R.H., eds. The primary productivity of the biosphere. New York, Springer Verlag.
READER, J.R. 1973 Leaf emergence, leaf coloration, and photosynthetic period. Productivity models for the eastern deciduous forest biome. Chapel Hill, University of North Carolina, Department of Botany. 182 p. (Ph.D. thesis)
SANCHEZ, P.A. Characteristics and management of soils in the tropics. Raleigh, North Carolina State University Soil Science Department. (Manuscript)
SANCHEZ, P.A. & BUOL, S.W. 1975 Soils of the tropics and the world food crisis. Science, 188: 598-603.
SHARP, D.D., LIETH, H. & WHIGHAM, D. 1975 Assessment of regional productivity in North Carolina. In Lieth, H. and Whittaker, R.H., eds. The primary productivity of the biosphere. New York, Springer Verlag.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}