![]()
![]()
![]()
B.A. Wilcox
Bruce A. Wilcox travaille à l'Institut du développement durable de Menlo Park, Californie (Etats-Unis).
Analyse des démarches utilisées actuellement pour évaluer les effets du déboisement sur la biodiversité, avec réexamen du concept de diversité biologique à la lumière des progrès récents de la théorie écologique, et description d'un cadre rationnel permettant d'appliquer le principe de l'analyse des risques à l'évaluation et à la réduction des effets négatifs du déboisement tropical sur la biodiversité. Il est suggéré d'adopter une perspective plus large qui mette l'accent sur le fonctionnement des écosystèmes en même temps que sur la biodiversité; sur la nécessité d'intégrer les valeurs écologiques, économiques et sociales dans l'évaluation des effets du déboisement; et sur les perspectives de conservation et d'utilisation durable.
Quand on parle de conservation de la biodiversité, on met surtout l'accent sur les espèces (souvent certaines espèces particulières, emblématiques, menacées d'extinction), en particulier lorsqu'on estime les pertes et qu'on prend des mesures de protection; toutefois, l'espèce n'est qu'un niveau de la diversité. La biodiversité vise aussi la variété et la variabilité, à des niveaux supérieurs et inférieurs de l'organisation biologique, entre écosystèmes ou habitats et à l'intérieur des espèces.
Pour l'heure, il est seulement possible de faire une caractérisation très générale des types et de la variété des écosystèmes et organismes qui existent dans les forêts (Centre mondial de surveillance de la conservation, 1992). Au niveau génétique, malgré une augmentation considérable des connaissances sur les ressources forestières (Namkoong, 1986), la caractérisation de la variabilité génétique au sein des espèces n'a été effectuée que sur quelques centaines parmi les centaines de milliers d'espèces qui existent. Les répartitions géographiques des types, des espèces et de la diversité génétique, des écosystèmes ou des habitats, ne sont connues que dans leurs très grandes lignes pour l'ensemble des tropiques, et l'on ne dispose de renseignements d'enquête détaillés sur l'un quelconque de ces niveaux que pour un nombre relativement limité de sites forestiers.
Les connaissances scientifiques sur les schémas et processus de la biodiversité des forêts tropicales ont beaucoup progressé ces dernières années. Les résultats publiés par le Projet d'évaluation des ressources forestières tropicales de 1990 (FAO, 1993), par exemple, ont beaucoup contribué à notre connaissance de l'état des forêts tropicales dans le monde. Grâce à l'expérience acquise au cours de l'évaluation mondiale précédente (FAO, 1982), l'évaluation de 1990 a utilisé des méthodologies plus fines, ce qui a permis de réviser les estimations de 1980, et de faire des estimations de l'état et du taux d'évolution du couvert forestier en 1990 pour 90 pays de la zone des forêts tropicales. L'évaluation a aussi utilisé les images-satellites de plus en plus courantes, et les nouveaux instruments d'analyse pour estimer la biomasse, la fragmentation et le risque pour la diversité biologique afin d'améliorer l'échelle de résolution et notre compréhension des conséquences de la dégradation et de la disparition du couvert forestier.
Toutefois, il subsiste d'importantes lacunes d'information concernant la répartition géographique des éléments, et leurs effets correspondants sur les forêts, ce qui ne permet pas de faire une estimation fiable de la perte de biodiversité. Même si on disposait de ces informations d'ordre spatial, les réponses extrêmement variables des espèces à la disparition et à la dégradation de l'habitat ne peuvent pas être calculées avec précision, même à une échelle relativement réduite, et encore moins au niveau mondial. L'emploi d'expressions mathématiques pour mettre en rapport le nombre d'espèces existantes et la superficie de l'habitat, tout en employant des modèles espèce-zone dont la généralité est avérée, ne permet pas de réduire cette incertitude (Lugo, Parrotta et Brown, 1993) Les estimations ainsi mises au point, qui sont de l'ordre de 3 à 10 pour cent des pertes totales d'espèces, reposent sur des estimations du déboisement des forêts tropicales d'ici l'an 2000, et sur de nombreuses hypothèses dont la validité est pour l'essentiel impossible à vérifier (Whitmore et Sayer, 1992). Elles ne tiennent pas compte non plus de la perte de biodiversité au niveau de l'habitat ou au niveau génétique, ce qui fait que l'on risque de sous-estimer les effets négatifs qu'entraîne la disparition de l'habitat forestier sur la diversité des éléments en général. En outre, les informations obtenues ne donnent guère d'orientation véritable pour des stratégies de conservation ou d'utilisation durable.
Il ne fait pas de doute que le déboisement dans les régions tropicales menace ou élimine de nombreux écosystèmes, espèces et populations exceptionnelles du point de vue génétique des forêts tropicales (Centre mondial de surveillance de la conservation, 1992), y compris des ressources génétiques de grande valeur des forêts tropicales (FAO, 1990). L'identification des priorités et l'accent des programmes de nombreux organismes internationaux et organisations non gouvernementales (ONG) s'appuient sur une liste, déjà longue et qui s'allonge sans cesse, d'espèces et d'habitats menacés sous les tropiques. Toutefois, le sens et la valeur de la biodiversité vont bien au-delà des éléments menacés et visent son rôle fonctionnel et sa valeur tels qu'ils sont perçus, quelle que soit l'échelle géographique et institutionnelle, et indépendamment du degré de rareté, du caractère exceptionnel ou de la nature de la menace actuelle.
Les écologistes forestiers reconnaissent depuis longtemps les interactions réciproques entre les structures géométriques et le fonctionnement des organismes et des habitats. Qualifiées aussi de «schémas et processus», ces relations envisagées à différentes échelles spatiales et temporelles sont considérées comme essentielles dans la détermination de l'importance des différents facteurs qui régissent l'évolution de la végétation (Solomon et Shugart, 1993). Une interaction à trois dimensions entre les composants (éléments), la structure et le fonctionnement est nécessaire pour élaborer des indicateurs permettant d'évaluer la biodiversité. Les caractères dominants et les facteurs importants peuvent être définis selon différentes échelles spatiales (paysages régionaux, communautés écologiques, sites locaux), et différents niveaux de diversité biologique (écosystème, espèce, population et génétique) (Noss, 1990). Différentes échelles institutionnelles ou de gestion (internationale, nationale, de district et de communauté) peuvent être ajoutées à ce schéma comme troisième dimension (FAO, 1994).
Une perspective centrée sur les éléments, en particulier si elle est exclusivement basée sur l'espèce, présente des limitations majeures de données et passe également à côté d'un ensemble important et croissant d'informations, en particulier pour ce qui est des conséquences indirectes sur la diversité des éléments, découlant des changements de la structure et de la fonction des écosystèmes provoqués par la disparition des forêts ou leur dégradation. En général, les travaux scientifiques récents suggèrent d'envisager la bio-diversité d'un point de vue plus large, la variété et la variabilité des éléments étant considérées dans leurs interactions avec les «schémas et processus» aux différentes échelles spatiales et temporelles. Il est devenu manifeste, par exemple, que la diversité extrêmement grande des arbres des forêts tropicales humides est la conséquence des bouleversements continus dans des zones non contiguës, ce qui entraîne une structure en mosaïque. De même, la variation de l'écotype, qui est à l'origine d'un grand nombre de ressources génétiques d'essences forestières, est assurée par et correspond à des schémas de variation de l'environnement dans toute une région ou paysage forestier. Les schémas de variation dans l'environnement et le paysage peuvent être de bonnes indications de la biodiversité et, en général, il est possible de les mesurer efficacement en utilisant des techniques de relevés au sol ou de télédétection.
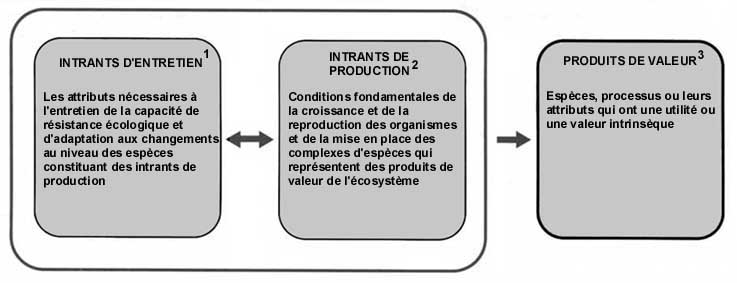
Les ressources forestières, comme toutes les ressources produites par des systèmes biologiques, sont renouvelables tant que leur «capital biologique» reste intact. Ce capital comprend les biens et services fournis par le matériel sur pied, tels que le bois ou la protection de bassins versants, et aussi la variété et la variabilité générales. Bien qu'on risque de réduire la complexité de l'interdépendance hommes-forêts à une perspective strictement économique ou de production, les conséquences du déboisement, et la dépendance des hommes vis-à-vis des écosystèmes forestiers, peuvent être évaluées et présentées clairement en utilisant les concepts de risque, de coût et de bénéfice. Grâce à une représentation visuelle du système de production des écosystèmes forestiers (fig. 1), les risques, les coûts et les bénéfices de la conversion ou de la conservation de la forêt peuvent être mis en relation sur la base des valeurs «d'usage» et de «non-usage» de la biodiversité. Les valeurs d'usage correspondent aux bénéfices productifs directs, tels que les produits ligneux et non ligneux, et aux bénéfices fonctionnels indirects. Ceux-ci comprennent les effets positifs sur la biosphère, l'entretien et la régulation des fonctions hydrologiques qui affectent le climat local, l'énergie hydroélectrique, les ressources en eau et les transports, ainsi que la protection des cultures, le cycle des éléments nutritifs, la conservation in situ des ressources génétiques, etc. Les valeurs de non-usage sont «intrinsèques» à une espèce ou à un habitat, et reposent sur des perspectives culturelles, esthétiques et spirituelles. La valeur monétaire de la plupart de ces bénéfices peut être estimée et, lorsque c'est le cas, leur contribution à l'économie locale et nationale s'avère en général beaucoup plus importante que d'autres utilisations des terres forestières (Pearce, Moran et Fripp, 1994).
Malgré ce qui vient d'être dit, la valeur réelle de la biodiversité échappe à la quantification en termes monétaires ou autres. Les calculs ne tiennent pas compte des difficultés de mesure, ou des bénéfices difficiles à identifier tels que les bénéfices indirects de production, ou des attributs et éléments de la biodiversité dont dépendent les éléments de la valeur productive et fonctionnelle pour leur croissance et leur reproduction. Dans les forêts tropicales, cela vise par exemple les oiseaux et les mammifères responsables de la pollinisation et de la dispersion des graines des nombreuses espèces de feuillus de grande valeur, ainsi que d'autres espèces végétales qui assurent leur alimentation et leur protection. Les calculs ne prennent pas non plus en compte l'évolution des besoins ou exigences des générations futures.
FIGURE 1 Représentation visuelle du système de production de la biodiversité
1Les attributs comprennent la complexité et la diversité écologiques (y compris la pléthore fonctionnelle) au niveau de l'espèce et au niveau génétique qui constituent la «variété et la variabilité» nécessaires pour entretenir les intrants de production et les produits de valeur.
2Par conditions fondamentales on entend les réseaux alimentaires, les relations trophiques, la fixation de l'énergie et des éléments nutritifs et les cycles biogéochimiques.
3Ces produits comprennent les «bénéfices de production directs» (biens), «les bénéfices fonctionnels indirects» (services), et les espèces de valeur intrinsèque.
Note: Presque toute la biodiversité d'un écosystème contribue de manière indirecte en protégeant le système du stress anthropique ou naturel (intrants d'entretien) ou en fournissant l'énergie immédiate et les matériaux nécessaires. Ces intrants d'entretien et de production conditionnent la fourniture des produits de valeur de manière durable.
Il est encore plus difficile de mesurer les attributs de la variété et de la variabilité qui contribuent à la capacité d'adaptation écologique et au potentiel d'évolution des écosystèmes forestiers, et donc à leur caractère durable. Ces «bénéfices complémentaires indirects» sont des attributs de la biodiversité qui renforcent ou sont responsables des processus opérant à chaque niveau (Wilcox, 1994). La mosaïque hétérogène du paysage forestier est une protection contre les effets des menaces naturelles que sont par exemple le mauvais temps et les épidémies de ravageurs. La concurrence entre les espèces et leur pléthore au sein des écosystèmes renforcent la dynamique des espèces et contribuent à assurer la résistance, leur capacité d'adaptation et leur survie génétique face aux stress de l'environnement d'origine humaine et naturelle.
Bien qu'on ne sache pas exactement quels degrés de variété et de variabilité sont nécessaires pour fournir des biens et services spécifiques, des méthodes existent pour estimer les conditions du maintien de la viabilité de populations végéta les et animales de valeur (Soulé, 1987), ainsi que de l'intégrité de l'écosystème naturel correspondant (Karr, 1994). Selon les conclusions d'enquêtes récentes, il existe beaucoup de combinaisons en surnombre dans les assemblages fonctionnels des écosystèmes riches en espèces (Schulze et Mooney, 1993). Toutefois, certains groupes fonctionnels ne sont constitués que d'une seule espèce, qui joue donc un rôle de «clé de voûte» soutenant souvent tout un réseau de nombreuses autres espèces au sein d'une communauté écologique.
Du point de vue de l'évaluation et de l'aménagement, il est évident qu'un petit nombre d'espèces et de processus «structurants» sont déterminants pour le maintien des valeurs d'usage direct et indirect et de non-usage de la biodiversité forestière. Ces facteurs structurants sont, par exemple, la «charpente» assurée par les arbres dominants faisant voûte, ou bien la «structure» au sens des réseaux alimentaires ou des interactions mutuelles entre animaux et arbres à fruits ou plantes à nectar. Parmi les facteurs structurants essentiels, on compte aussi des animaux tels les éléphants ou les castors, ou des agents physiques comme le feu qui bouleversent ou restructurent l'environnement (Bond, 1993).
La faune forestière tropicale a un rôle particulièrement important pour la diversité des essences. Les modifications de la composition ou de la diversité des espèces qui consomment des fruits et des graines peuvent influer sur la diversité des essences et même affecter la régénération des feuillus de grande valeur commerciale (Bawa et al 1989). Les effets d'une réduction de la diversité des mammifères sur les quantités de fruits tombés sont directement observables (Dirzo et Miranda, 1990), et la réduction des grands prédateurs, en général les plus vulnérables lorsqu'il y a des disparitions importantes d'habitat, influence de manière différentielle les densités de mammifères qui se nourrissent de fruits et de graines, et donc la régénération des arbres dominants à grosses graines (Terborgh, 1992). Par conséquent, le déclin général de la diversité de la faune des forêts tropicales aura probablement des effets considérables mais différés, dont les conséquences pour la biodiversité générale dans les forêts tropicales pourraient être dramatiques. On peut également faire valoir avec force qu'il est important de maintenir la diversité des insectes mangeurs de plantes, ainsi que des invertébrés en général, pour assurer l'évolution continue et la diversification des espèces végétales, y compris le maintien des ressources génétiques (Brown, 1987).
Comme on l'a déjà souligné, les risques visant la biodiversité liés au déboisement sont souvent considérés en termes de perte d'espèces. Il est également important d'étudier les effets du déboisement sur la fonction de l'écosystème, qui touchent à la fois la valeur biologique et la valeur d'usage qui s'appuient sur le maintien de l'intégrité écologique. Comme on l'a déjà indiqué, l'estimation des effets du déboisement sur la disparition d'espèces pose un problème pour diverses raisons. Par exemple, les modèles qui mettent en rapport l'extinction des espèces et la perte d'habitat montrent que le taux de disparition des espèces s'accélère seulement lorsqu'environ les deux tiers de l'habitat original ont été perdus (FAO, 1994; Tilman, May et Nowak, 1994). Toutefois, ces modèles ne permettent pas d'étudier les effets du déboisement sur la fonction de l'écosystème, qui peuvent intervenir de manière plus importante bien avant que les deux tiers du couvert forestier primitif d'un écosystème forestier aient disparu. En outre, une analyse du couvert forestier montre que la plupart des régions ont déjà perdu plus des deux tiers de leur couvert primitif (voir tableau). A cet égard, «l'appauvrissement biotique» général qui accompagne la dégradation de l'écosystème forestier est plus important que la perte d'espèces en elle-même (fig. 2).
TABLEAU. Comparaison régionale des pourcentages de forêts primitives subsistant en 1990
|
Région |
Forêt primitive1 |
Forêt en 19902 |
Forêt primitive encore en place en 1990 |
|
(milliers ha) |
(%) |
||
|
Asie |
|||
|
Asie du Sud |
311 722 |
61 150 |
20 |
|
Asie du Sud-Est continentale |
189 609 |
75 197 |
40 |
|
Asie du Sud-Est insulaire |
244 140 |
135 349 |
60 |
|
Amérique latine |
|||
|
Amérique centrale |
176 744 |
66 673 |
40 |
|
Amérique du Sud |
1 278371 |
802716 |
60 |
|
Afrique |
|||
|
Afrique Sahel Ouest |
145 667 |
40 750 |
30 |
|
Afrique Sahel Est |
251 973 |
64 631 |
30 |
|
Afrique de l'Ouest |
200 566 |
55 607 |
30 |
|
Afrique centrale |
394 837 |
204 112 |
50 |
|
Afrique tropicale |
457 249 |
143 297 |
30 |
|
Afrique insulaire |
116 508 |
15 782 |
10 |
1Olson, Watts et Allison (1985).
2Données FAO.
Note: Les estimations de la superficie de la «Forêt primitive» sont fondées sur la cane numérique d'Olson, Watts et Allison (1985). Elles ont été comparées à d'autres estimations fondées sur la combinaison de diverses méthodes telles que la cartographie bioclimatique, la télédétection et la vérification sur le terrain, et ont été jugées les plus exactes et les plus conservatrices, malgré l'insuffisance générale de données existant sur la végétation mondiale. Le couvert forestier primitif est une notion hypothétique fondée sur une reconstitution théorique des conditions préhistoriques, mais qui suppose que le climat et d'autres conditions physiques et biotiques n'ont pas changé depuis les temps préhistoriques. La qualité, la résolution et l'interprétation des données des cartes numériques en ce qui concerne la végétation et les frontières géopolitiques rendent les estimations numériques peu fiables (plus ou moins 20 pour cent d'erreur). Toutefois les valeurs concernant les forêts primitives subsistances, qui ont été arrondies aux 10 pour cent les plus proches, donnent une idée assez juste de la véritable étendue des forêts actuelle par rapport à ce qu'elle aurait été en l'absence de toute intervention humaine.
La biodivèrsité ainsi que les risques, les coûts et les bénéfices liés à l'état et au changement du couvert forestier sont évalués de manière beaucoup plus utile si l'on se place dans l'optique de la lutte contre l'appauvrissement biotique et du maintien de l'intégrité biotique ou de la santé de l'écosystème. Un écosystème forestier sain c'est-à-dire dont l'intégrité biotique est maintenue - peut se définir comme suit: son potentiel inhérent est réalisé, son état est stable, sa capacité d'autoréparation, lorsqu'il est perturbé (stressé), est préservée et il a besoin d'un minimum d'aménagements extérieurs. La biodiversité et l'intégrité biotique sont interdépendantes et elles sont toutes deux menacées ou diminuées par le déboisement (Rapport, Regier et Hutchison, 1985). Le stress et la réaction au stress peuvent avoir des effets négatifs sur l'intégrité biotique et sur la biodiversité qui provoquent un syndrome d'effets associés à la dégradation de l'écosystème, aboutissant à l'appauvrissement biotique (fig. 2). Le stress, la réaction, les effets et le degré d'appauvrissement biotique sont tous mesurables par des indicateurs appropriés. La réaction, ou changement de la structure ou de la composition des écosystèmes (composition des espèces) est souvent plus utile en tant qu'indicateur d'alerte rapide du stress que des changements fonctionnels (par exemple cycle des éléments nutritifs).
FIGURE 2 Modèle de dégradation d'un écosystème forestier
Le stress d'un écosystème forestier peut provenir de diverses sources:
· surexploitation des ressources renouvelables
· pollution
· restructuration physique
· espèces exotiques
· variations naturelles
Source: Rapport, Regier et Hutchison (1985).
Source du graphique: Modifiée d'après Jordan (1985).
Le processus général de dégradation des écosystèmes forestiers, également appelé rétrogression écologique par certains écologistes, s'accompagne de:
· la réduction de la diversité des essences
· la raréfaction des éléments nutritifs des plantes
· un changement structurel, la forêt cédant la place à des broussailles robustes et des prairies
· la simplification des relations entre les essences
· une baisse de la productivité primaire
Source: Woodwell (1967, 1970).
Au sens analytique strict, l'évaluation des risques suppose l'identification des risques ou des dangers et l'estimation de leur degré de probabilité, Ainsi, l'évaluation des risques concernant la biodiversité nécessite de définir ce qu'est une conséquence ou un résultat indésirable. Les résultats indésirables de ce point de vue sont par exemple la disparition d'éléments ou de produits de valeur de la biodiversité. Par conséquent, le risque pour la biodiversité peut se définir comme une situation qui pourrait avoir des conséquences indésirables impliquant une modification dans la variété ou la variabilité des écosystèmes, des espèces, des populations et des organismes.
On considère qu'un niveau particulier de dégradation forestière et d'appauvrissement biotique constitue un risque du point de vue de la société en fonction des valeurs et préférences des utilisateurs de la forêt, ou des bénéficiaires des biens et services qu'elle offre. Ainsi, l'évaluation des risques pour la biodiversité se fait effectivement à deux niveaux distincts. L'un est la détermination de l'effet anthropique sur l'intégrité biotique. L'autre est la détermination de l'effet de l'appauvrissement biotique sur la société, et la diminution à long terme des valeurs d'usage et de non-usage des écosystèmes.
L'examen des relations causales entre le déboisement, la fonction de l'écosystème et la biodiversité montre qu'un certain nombre de facteurs interviennent dans la dégradation de l'écosystème et l'appauvrissement biotique. Certains de ces facteurs dépendent autant des schémas de déboisement et des modes d'utilisation de la forêt que du montant total de la perte d'habitat forestier. Ainsi, on pourrait souvent limiter les effets les plus graves de la perte de zones forestières en mettant en place des zones tampons, des couloirs, de larges espaces de forêt naturelle, et en promouvant des actions de reboisement écologique. Il existe des mesures spécifiques de réduction des risques pour conserver le paysage local, ou les attributs structurels ou fonctionnels au niveau régional qui sont bouleversés par la disparition et la fragmentation des habitats forestiers.
Il y a toujours un risque de réductionnisme dans l'évaluation des risques. Accorder une attention excessive aux mécanismes détaillés et aux relations causales directes peut faire perdre de vue des évidences qui sont connues avec certitude. Au cours de ces 20 à 30 dernières années, le couvert forestier des pays tropicaux s'est réduit à un taux sans précédent dans l'histoire de l'humanité. Bien que des taux de déboisement comparables se soient produits localement et dans certaines régions dans le passé (Williams, 1990),l'extension géographique et la rapidité du phénomène sont sans précédent. En outre, l'augmentation des précipitations, du rayonnement solaire et de la dépendance directe des populations locales vis-à-vis des forêts pour leur alimentation renforce les conséquences écologiques, sociales et économiques du déboisement dans les pays tropicaux. Associé à des augmentations tout aussi extraordinaires des émissions de gaz carbonique dans les pays industrialisés, le déboisement tropical aggrave le risque de modification du climat mondial (à cause du dégagement de gaz carbonique dû à la combustion de la végétation forestière et à l'augmentation de l'albédo ou réfléchissement de l'énergie solaire). Ce dernier phénomène a pour effet complémentaire de produire des sécheresses et des réchauffements locaux et régionaux, ce qui rend problématique la régénération naturelle des forêts et même les efforts déployés par l'homme pour régénérer les terres forestières.
De nombreuses formes d'activité et de production humaines sont compatibles avec le maintien des forêts et reposent même sur l'entretien des forêts, y compris de leur biodiversité naturelle. L'exploitation durable des ressources forestières renouvelables et le remaniement des stocks, et des variétés et variabilités correspondantes, ne diminuent pas nécessairement leur capital biologique. En fait, accélérer le développement durable des ressources forestières dans les pays en développement, y compris convertir des forêts qui n'étaient pas encore perturbées en forêts productives aménagées, afin d'élever le niveau de vie et de rompre le cycle de la pauvreté et de la dégradation de l'environnement, est peut-être le moyen d'assurer au mieux la conservation de la biodiversité dans de nombreux pays en développement. La quantité totale de forêts naturelles nécessaire pour «capturer» les zones où se trouve la plus grande richesse d'essences indigènes, le plus d'espèces endémiques et de ressources génétiques forestières, est probablement assez faible tant que le couvert forestier total reste suffisant pour entretenir les propriétés fonctionnelles et structurelles d'un écosystème ou paysage forestier régional. Une évaluation de notre expérience actuelle tend à montrer que la conservation de 75 à 90 pour cent du couvert forestier actuel est probablement nécessaire pour assurer la stabilité des fonctions écologiques locales et globales (Woodwell, 1993). Cette condition pourrait être satisfaite avec des mosaïques de paysages forestiers bien aménagées comprenant des forêts naturelles, des plantations, des zones d'agroforesterie, des rideaux-abris et autres formations. Toutefois, même en conservant cette quantité de couvert forestier, on ne garantit pas la santé et la durabilité d'un écosystème forestier régional, y compris le maintien de sa capacité de résistance et son adaptabilité. Cela suppose de porter une attention particulière aux conditions requises pour le maintien des attributs de la biodiversité qui contribuent à la valeur de la forêt, qu'elle soit aménagée ou non.
Pour ce faire, il faut disposer d'un cadre rationnel concernant les politiques et la planification de l'utilisation des terres forestières, qui repose sur une comptabilité exhaustive des valeurs d'usage et de non-usage des forêts.
Bien que l'ensemble des connaissances sur le fonctionnement des écosystèmes et la biodiversité augmente rapidement et permette de mieux utiliser les ressources forestières de manière durable, ces mêmes connaissances prouvent que les tendances et schémas actuels de déboisement sont incontestablement négatifs. Les méthodes empiriques quantitatives qui lient le couvert forestier au fonctionnement des écosystèmes et à la biodiversité sont trop générales pour prévoir de manière fiable les niveaux précis de dégradation et d'appauvrissement à grande échelle (par exemple au niveau régional), mais elles sont adéquates pour montrer que la réduction ou la disparition de forêts dans presque tous les pays tropicaux est grave et même catastrophique dans certains cas.
Toutefois, il ne s'ensuit pas automatiquement qu'il y aura une perte désastreuse correspondante de la biodiversité des forêts tropicales, à un niveau qui mettrait en péril le rôle essentiel des forêts dans la société. La théorie écologique laisse aussi penser que l'érosion de la biodiversité a du retard sur la disparition et la fragmentation des forêts. Et on peut aussi inverser les conditions qui ont antérieurement abouti à une utilisation non durable des forêts. Ces tendances pourraient se développer suffisamment pour entraîner une stabilisation et même un accroissement net du couvert forestier dans certains pays tropicaux d'ici 10 ou 20 ans.
Les préoccupations que suscitent la biodiversité des forêts tropicales et leur évaluation, compte tenu des taux élevés de disparition de forêts tropicales que l'on constate actuellement, imposent d'adopter une démarche rationnelle large pour infléchir les priorités stratégiques en faveur de la conservation et du développement durable. Cette démarche devrait mettre l'accent sur ce qui est scientifiquement connu au sujet de la biodiversité, y compris le rôle important de ses attributs structurels et fonctionnels. Mais elle devrait aussi prendre en compte ce qui n'est pas connu, et qui pourrait prendre trop de temps et d'argent à définir. Malgré les importantes lacunes de l'information, une masse impressionnante de connaissances existe à partir de laquelle une telle démarche rationnelle peut se développer; une grande quantité de données été accumulée et synthétisée ces dernières années. Il s'agit de démarches conceptuelles et méthodologiques permettant d'évaluer: i) les valeurs et avantages relatifs des différents attributs et éléments de la biodiversité; ii) le rôle de la biodiversité dans le fonctionnement des écosystèmes; iii) les relations entre les éléments et les aspects structurels et fonctionnels critiques de la biodiversité; iv) les schémas et processus des écosystèmes à différentes échelles spatiales et temporelles qui influencent la biodiversité; et v) les modes et schémas d'utilisation des terres et des forêts compatibles avec la conservation et l'aménagement durable de la biodiversité des forêts tropicales. Prises ensemble, ces informations permettent de construire un cadre d'évaluation des risques potentiels liés au déboisement et d'indiquer les mesures à prendre pour les limiter.
Dans de nombreux pays, cependant, les pressions de la collecte de bois de feu et de l'agriculture qui s'exercent sur les forêts font de la stabilisation des superficies forestières un défi sans précédent pour les décideurs et les responsables de l'aménagement des forêts, avant que presque toutes les forêts primitives aient disparu et que ne disparaissent des quantités importantes de biodiversité indigène.
Bawa, K.S., Ashton, P.S., Primack, R.B., Terborgh, J., Salleh, M.N., Ng, F.S.P. et Hadley, M. 1989. Reproductive ecology of tropical forest plants: research insights and management implications. Paris, UISB.
Bond, WJ. 1993. Keystone Species. In E.-D. Schulze et H.A. Mooney (éds). Biodiversity and ecosystem function, p. 237-253. New York, Springer.
Brown Jr, K.S. 1987. O papel dos consumidores na conservação e no manejo de recursos genéticos florestais in situ. IPEF, 35: 61 -69.
Centre mondial de surveillance de la conservation. 1992. Global biodiversity: status of the earth's living resources. Londres, Chapman and Hall.
Dirzo, R. et Miranda, A. 1990. Contemporary neotropical defaunation and forest structure, fonction, and diversity - a sequel to John Terborgh. Conserv. Biol. 4(4): 444-447.
FAO. 1982. Les ressources forestières tropicales. Etude FAO: Forêts n° 30. Rome, FAO/PNUE.
FAO. 1990. Groupe FAO d'experts des ressources génétiques forestières. Septième session, 4-6 décembre 1989, Rome.
FAO. 1994. Evaluation des ressources forestières 1990: pays tropicaux. Etude FAO: Forêts n° 112. Rome.
FAO. 1994. Tropical deforestation and the loss of biological diversity: assessing the risks. Rome.
Franklin, J.F. 1992. Scientific basis for new perspectives in forests and streams. In R. J. Naiman (éd.). Watershed management: balancing sustainability and environmental change, p. 25-72. New York, Springer.
Jordan. C.F. 1985. Nutrient cycling in tropical forest ecosystems. Chichester, Royaume-Uni, Wiley.
Karr, J.R. 1992. Ecological integrity: protecting earth's life support systems. In R. Costanza, B.G. Norton et B.D. Haskell (éds). Ecosystem health: new goals for environmental management. Washington, Island Press.
Karr, J.R. 1994. Landscapes and management for ecological integrity. In K.C. Kim et R.D. Weaver (éds). Biodiversity and landscape: a paradox for humanity. New York, Cambridge University Press.
Lugo, A.E., Parrotta, J.A. et Brown, S. 1993. Loss in species caused by tropical deforestation and their recovery through management. Ambio 22(2-3): 106-109.
Namkoong, G. 1986. La génétique et les forêts de l'avenir. Unasylva 38(152): 2-18.
Nepstad, D.C. et Schwartzman, S., (éds). 1992. Non-timber products from tropical forests: evaluation of a conservation and development strategy. Advances in economic botany, Vol. 9. New York, New York Botanical Garden.
Nepstad, D.C., de Carvalho, C.R., Davidson, P., Jipp, H., Lefebvre, P.A., Hebreiros, G.H. et da Silva, E.D. The deep-soil link between water and carbon cycles of Amazonian forests and pastures. Woods Hole Research Center/EMBRAPA-CPATU, Université de Californie, Irvine, Etats-Unis. (Sous presse)
Noss, R.F. 1990. Indicators for monitoring biodiversity: a hierarchical approach. Conserv. Biol. 4(4): 355-364.
Olson, J., Watts, J.A. et Allison, LJ. 1985. Major world ecosystem complexes ranked by carbon in live vegetation: a database. NDP 017. Oak Ridge, Tennessee, Etats-Unis, Oak Ridge National Laboratory.
Pearce, D., Moran, D. et Fripp, E.1994. The economic value of biological and cultural diversity. Gland, Suisse, UICN.
Rapport, D.J., Regier, H.A. et Hutchison, T.C. 1985. Ecosystem behavior under stress. Am. Nat., 125(5): 429-433.
Schulze, E.D et Mooney, H.A., (éds). 1993. Biodiversity and ecosystem function. Ecological Studies. New York, Springer.
Singh, K.D. 1993. L'évaluation des ressources forestières tropicales en 1990. Unasylva 44(174): 10-19.
Solbrig, O.T. et Nicolis, G. (éds). 1991. Perspectives on biological complexity. Paris, UISB.
Solomon, A.M. et Shugart, H.H., (éds).1993. Vegetation dynamics and global change. New York, Chapman and Hall.
Soulé, M.E., (éd). 1987. Viable populations for conservation. Cambridge, Royaume-Uni, Cambridge University Press.
Terborgh, J T. 1992. Maintenance of diversity in tropical forests. Biotropica, 24(2b): 283-292.
Thomas, C.D. 1990. Fewer Species. Nature, 347:237.
Tilman, D., May, R.M. et Nowak, M.A. 1994. Habitat destruction and the extinction debt. Nature, 371: 65-66.
Whitmore, T.C. et Sayer, J.A. (éds). 1992. Tropical deforestation and species extinction. Londres. Chapman and Hall.
Wilcox, B.A. 1994. Ecosystem goods and services: an operational framework. Working Paper of the 2050 Project. Washington, Institut mondial pour les ressources.
Williams, M. 1990. Forests. In B.L. Turner II, W.C. Clark et R.W. Kates (éds). The earth as transformed by human action, p. 179-202. Cambridge, Royaume-Uni, Cambridge University Press.
Woodwell, G.M. 1967. Radiation and the paterns of nature. Science, 156: 461-470.
Woodwell, G.M.1970. Effects of pollution on the structure and physiology of ecosystems. Science, 168: 429-433. Woodwell, G.M. 1993. Forests: What in the world are they for? In K. Ramakrishna et G. M. Woodwell (éds). World forests for the future: their use and conservation p. 1-20. New Haven et Londres, Yale University Press.
![]()
![]()
![]()
{kind=link}
{kind=link}