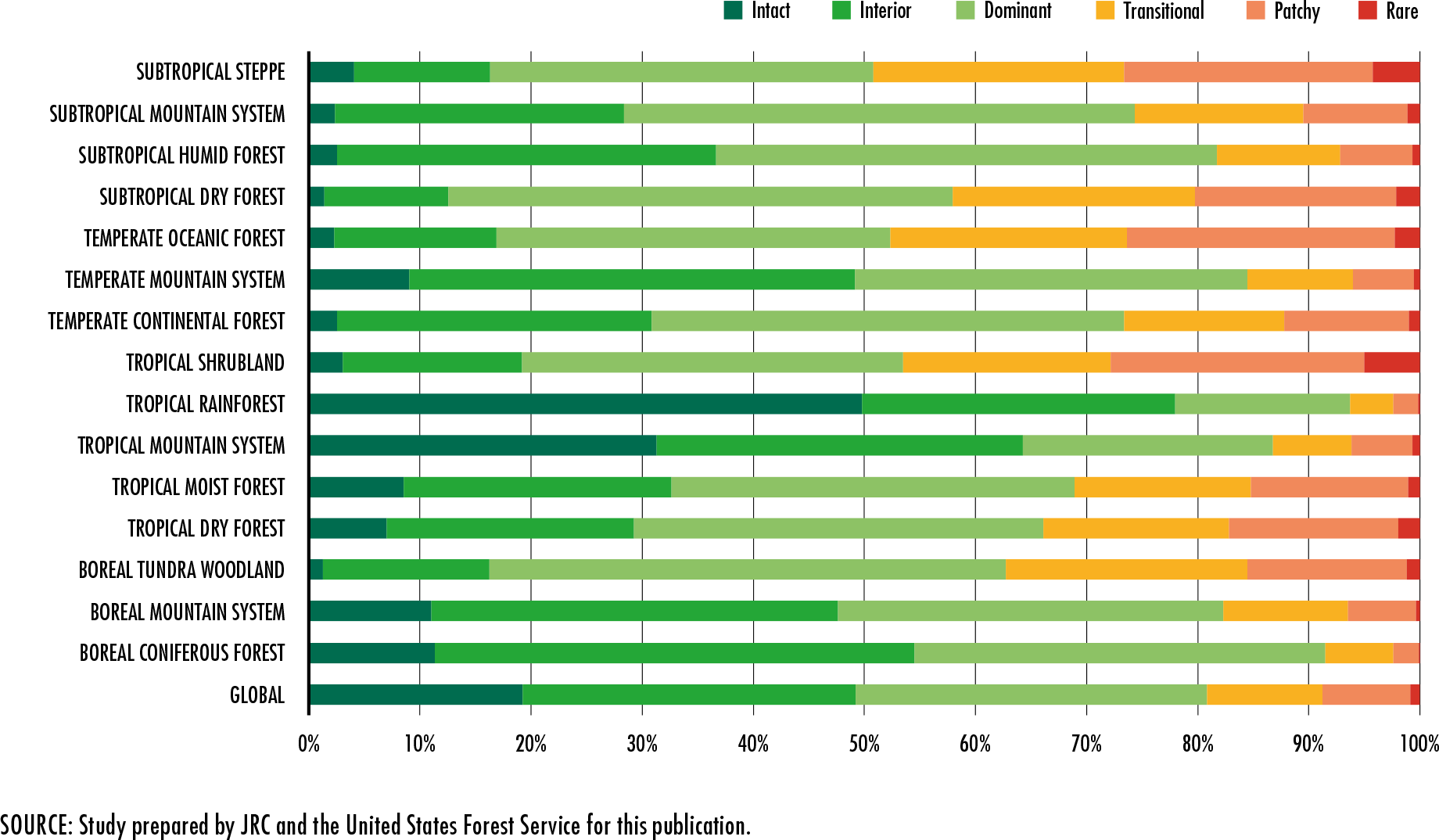
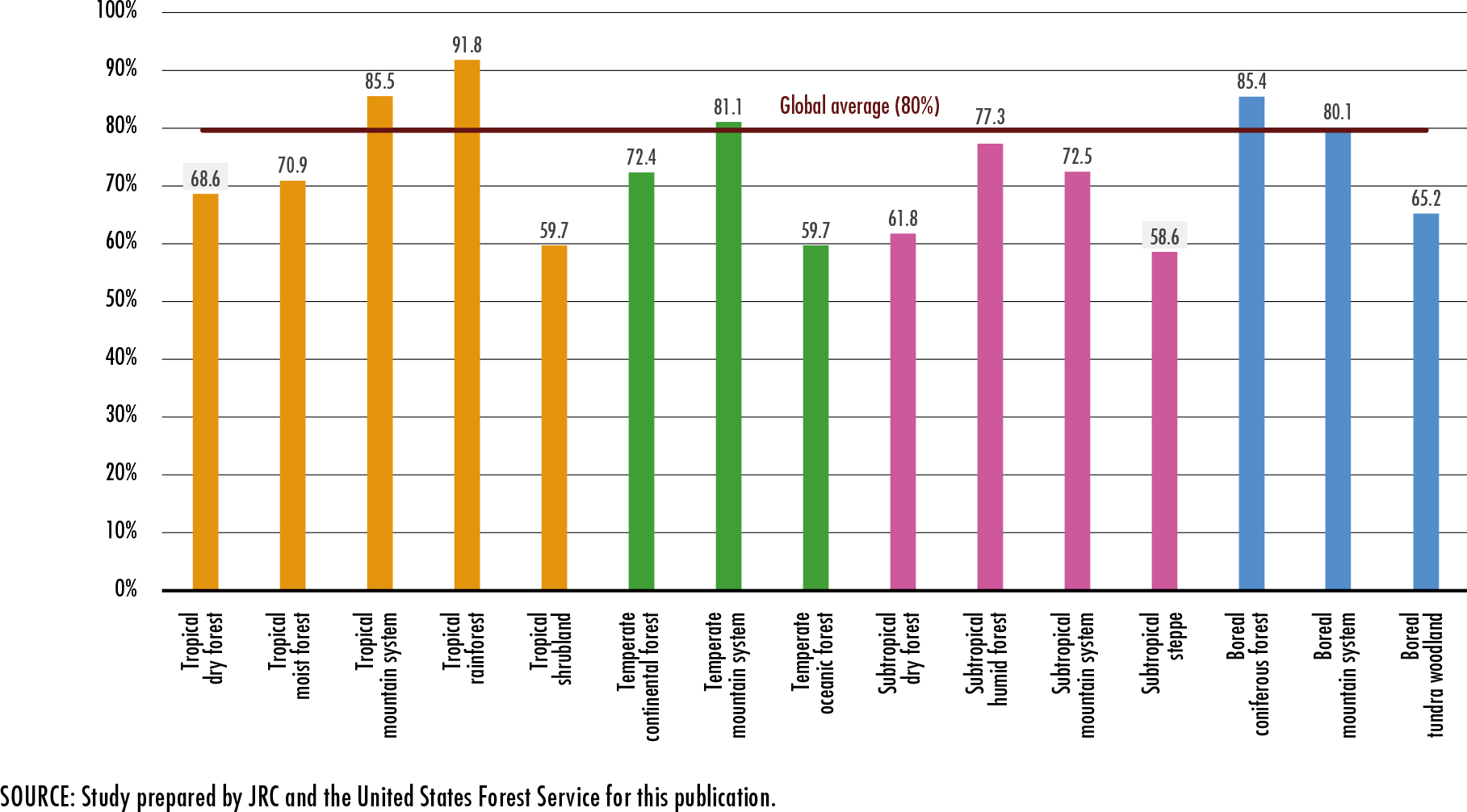
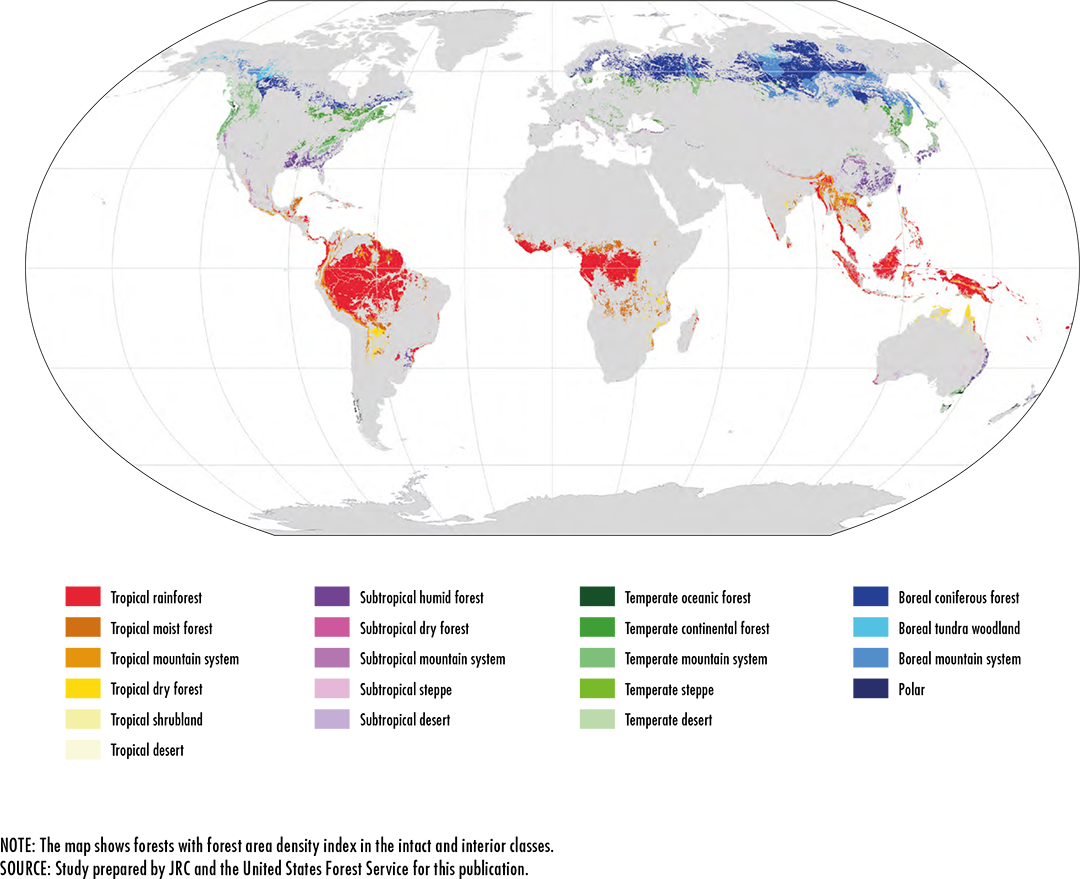
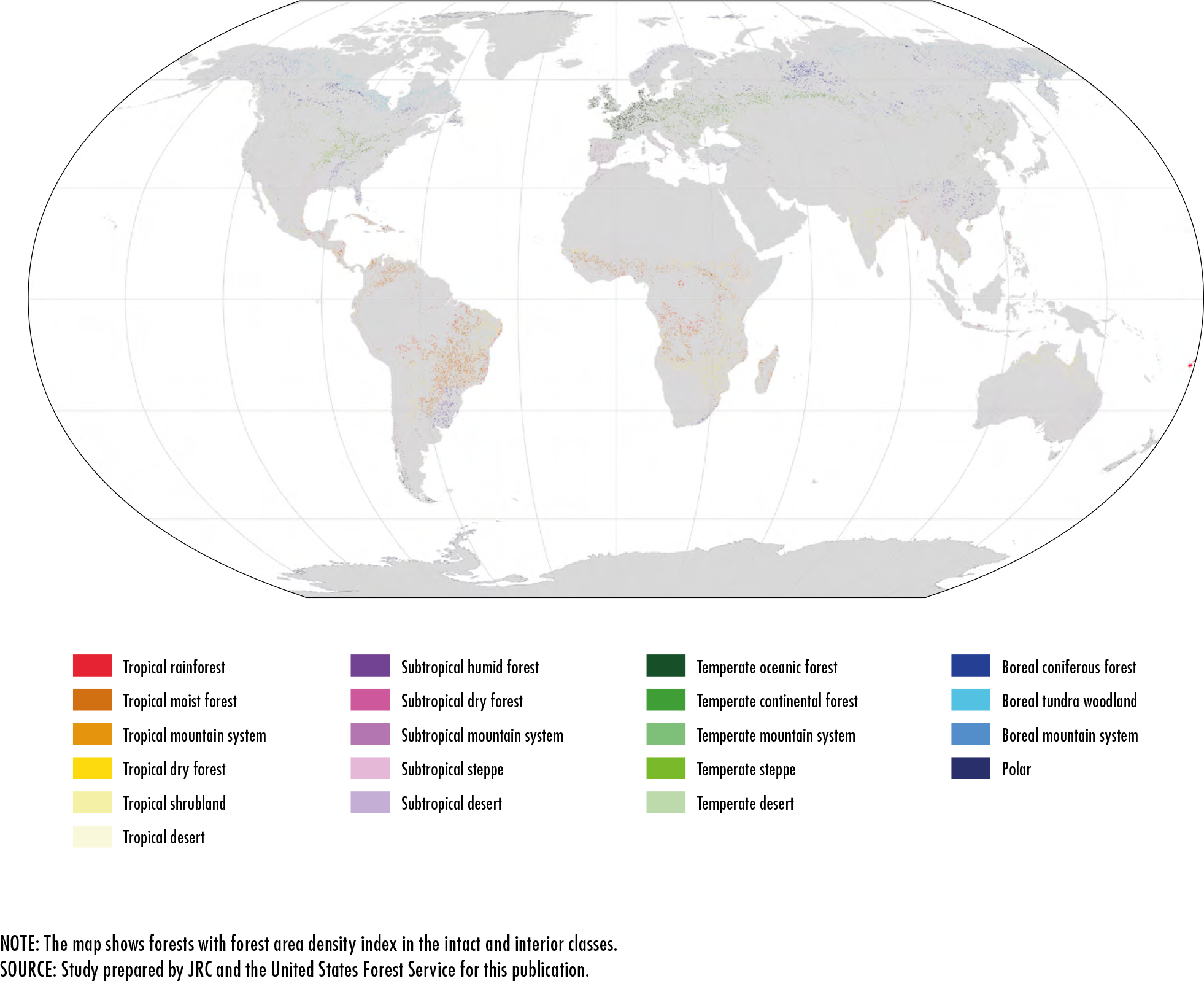
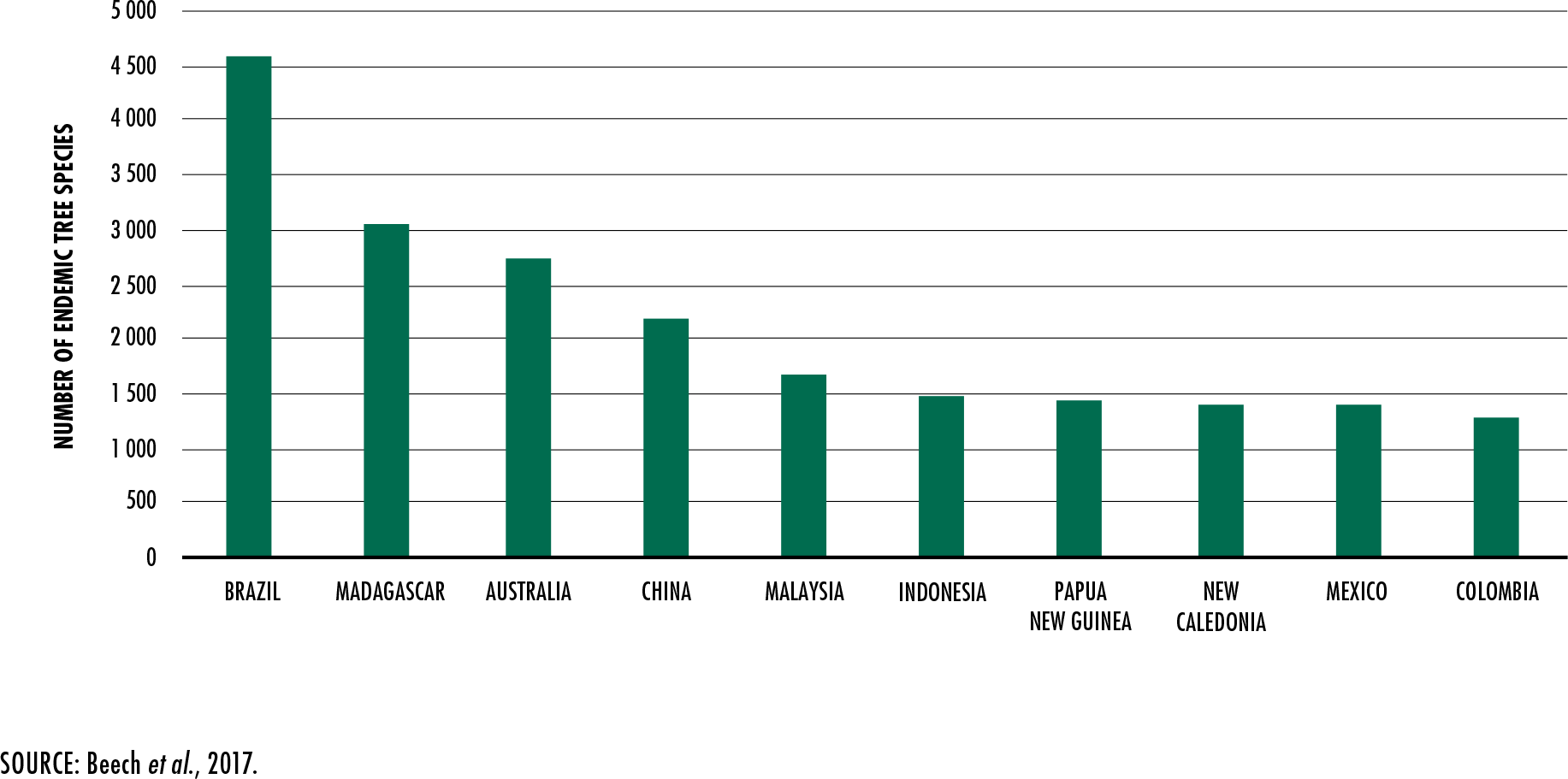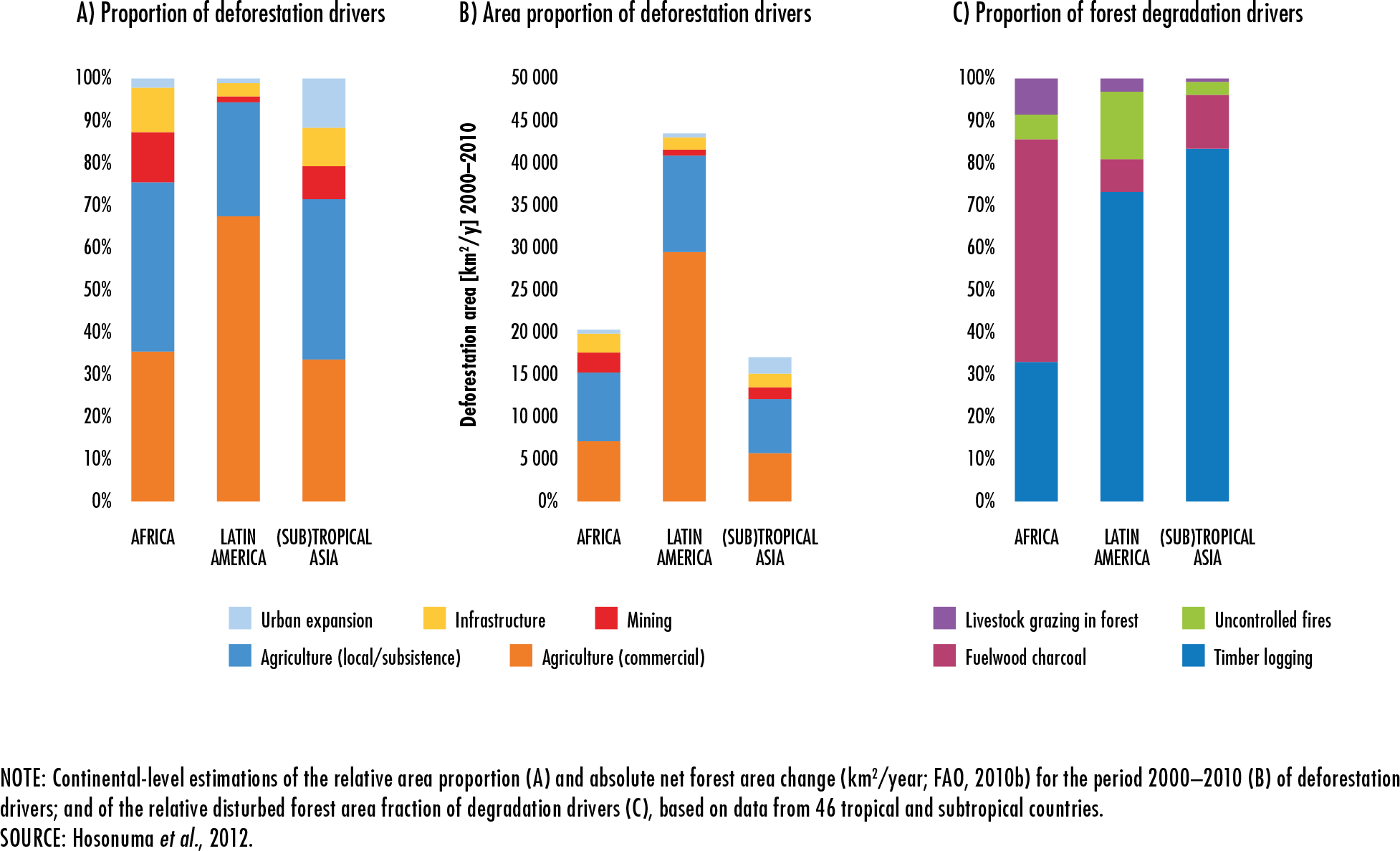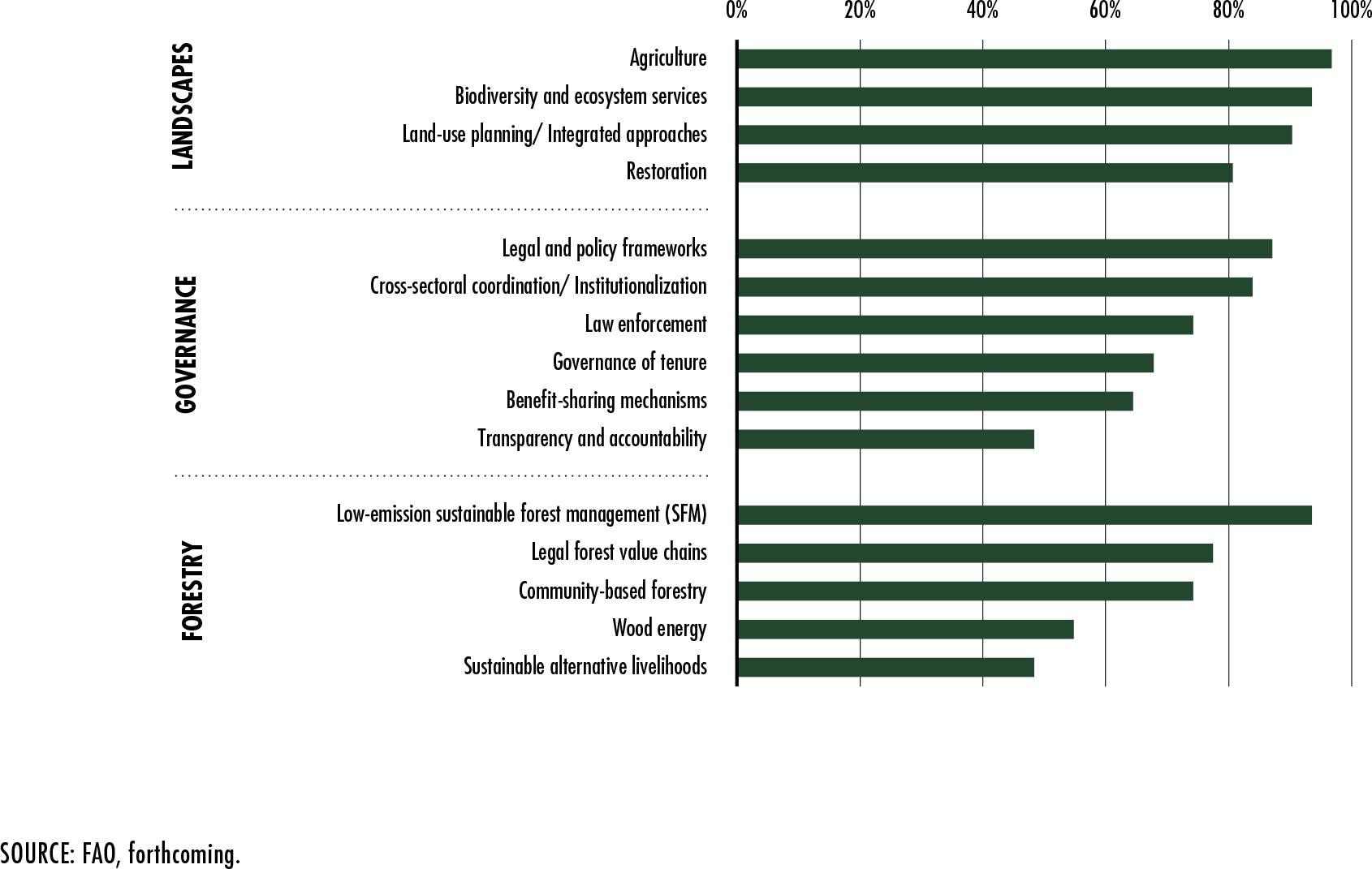References
ADB. 2016. Illicit trade in natural
resources in Africa –– A forthcoming report from the African Natural Resources
Center. Abidjan. [also available at
https://www.afdb.org/fileadmin/uploads/afdb/Documents/Events/IFF/Documents_IFF/ANRC_ILLICIT_TRADE_IN_NATURAL_RESOURCES.pdf].
AFR100. n.d. Home [online]. Midrand, South Africa. [Cited
18 December 2019].
https://afr100.org/
African Union. n.d. Agenda 2063: The Africa we want. In:
African Union [online]. Addis Ababa. [Cited 13 January 2020].
https://au.int/en/agenda2063/overview
Agrawal, A., Chhatre, A., & Hardin, R.
2008. Changing governance of the world’s forests.
Science, 320(5882): 1460–1462.
Aguilar, R., Quesada, M., Ashworth, L., Herrerias-Diego, Y. & Lobo,
J.
2008. Genetic consequences of habitat fragmentation in plant populations:
susceptible signals in plant traits and methodological approaches. Molecular
Ecology, 17: 5177–5188.
Ahenkan, A.
& Boon, E. 2011. Improving nutrition and health through non-timber
forest products in Ghana. Journal of Health, Population and Nutrition,
29(2): 141–148.
Alix-Garcia, J., Sims, K.R. & Yañez-Pagans, P.
2015. Only one tree from each seed? Environmental effectiveness and poverty
alleviation in Mexico’s payments for Ecosystem Services Program.
American Economic Journal: Economic Policy, 7(4):1–40.
Alix-Garcia, J., McIntosh, C., Sims, K., & Welch, J.
2013. The ecological footprint of poverty alleviation: Evidence from Mexico’s
Oportunidades Program.
The Review of Economics and Statistics, 95(2): 417–435.
Alkire, S.
& Santos, M.E. 2014. Measuring acute poverty in the developing
world: robustness and scope of the multidimensional poverty index. World
Development, 59: 251–274.
Andam, K.S., Ferraro, P.J., Pfaff, A., Sanchez-Azofeifa, G.A. &
Robalino, J.A.
2008. Measuring the effectiveness of protected area networks in reducing
deforestation. PNAS, 105(42): 16089–16094.
Angelsen, A., Jagger, P., Babigumira, R., Belcher, B., Hogarth, N.J., Bauch,
S., Börner, J., Smith-Hall, C. & Wunder, S.
2014. Environmental income and rural livelihoods: a global-comparative analysis.
World Development, 64: S12–S28. [online]. [Cited 3 January 2020].
https://doi.org/10.1016/j.worlddev.2014.03.006
Anup, K.C. 2017. Community forestry management and its role in
biodiversity conservation in Nepal. In G.A. Lameed, ed. Global
exposition of wildlife management [online]. [Cited 3 January 2020].
https://www.intechopen.com/books/global-exposition-of-wildlife-management/community-forestry-management-and-its-role-in-biodiversity-conservation-in-nepal
Azevedo, A.A., Rajão, R., Costa, M.A., Stabile, M.C.C., Macedo, M.N., Dos
Reis, T.N.P., Alencar, A., Soares-Filho, B.S. & Pacheco, R.
2017. Limits of Brazil’s Forest Code as a means to end illegal deforestation.
PNAS, 114(29): 7653–7658.
Balmford, A., Green, J.M., Anderson, M., Beresford, J., Huang, C., Naidoo,
R., Walpole, M. & Manica, A.
2015. Walk on the wild side: estimating the global magnitude of visits to protected
areas. PLOS Biology, 13(2): p.e1002074 [online]. [Cited 3 January 2020].
https://doi.org/10.1371/journal.pbio.1002074
Banerjee, O., Cicowiez, M., Horridge, M., & Vargas, R.
2016. A Conceptual Framework for Integrated Economic–Environmental Modeling.
Journal of Environment and Development, 25(3): 276–305. [also available at
doi: 10.1177/1070496516658753]
Barlow, J., Gardner, T.A., Araujo, I.S., Ávila-Pires, T.C., Bonaldo, A.B.,
Costa, J.E., Esposito, M.C.
et al. 2007. Quantifying the biodiversity value of tropical primary,
secondary, and plantation forests. PNAS, 104: 18555–18560.
Barros, F.M., Peres, C.A., Pizo, M.A. & Ribeiro, M.C.
2019. Divergent flows of avian-mediated ecosystem services across forest-matrix
interfaces in human-modified landscapes. Landscape Ecology, 35(4): 879
[online]. [Cited 3 January 2020].
https://doi.org/10.1007/s10980-019-00812-z
Bastin, J.-F., Finegold, Y., Garcia, C., Mollicone, D., Rezende, M., Routh,
D., Zohner, C.M. & Crowther, T.W.
2019. The global tree restoration potential.
Science, 365(6448): 76–79.
Baynham-Herd, Z., Amano, T., Sutherland, W.J. & Donald, P.F.
2018. Governance explains variation in national responses to the biodiversity
crisis.
Environmental Conservation, 45(4): 407–418.
Beatty, C.R., Cox, N.A. & Kuzee,
M.E.
2018.
Biodiversity guidelines for forest landscape restoration opportunities
assessments. 1st edition. Gland, Switzerland, IUCN.
Beck,
H. 2008. Tropical ecology. In Jørgensen, S.E. & Fath, B.D.
eds. General ecology: Encyclopedia of ecology, pp. 3616–3624. Elsevier,
Oxford, UK.
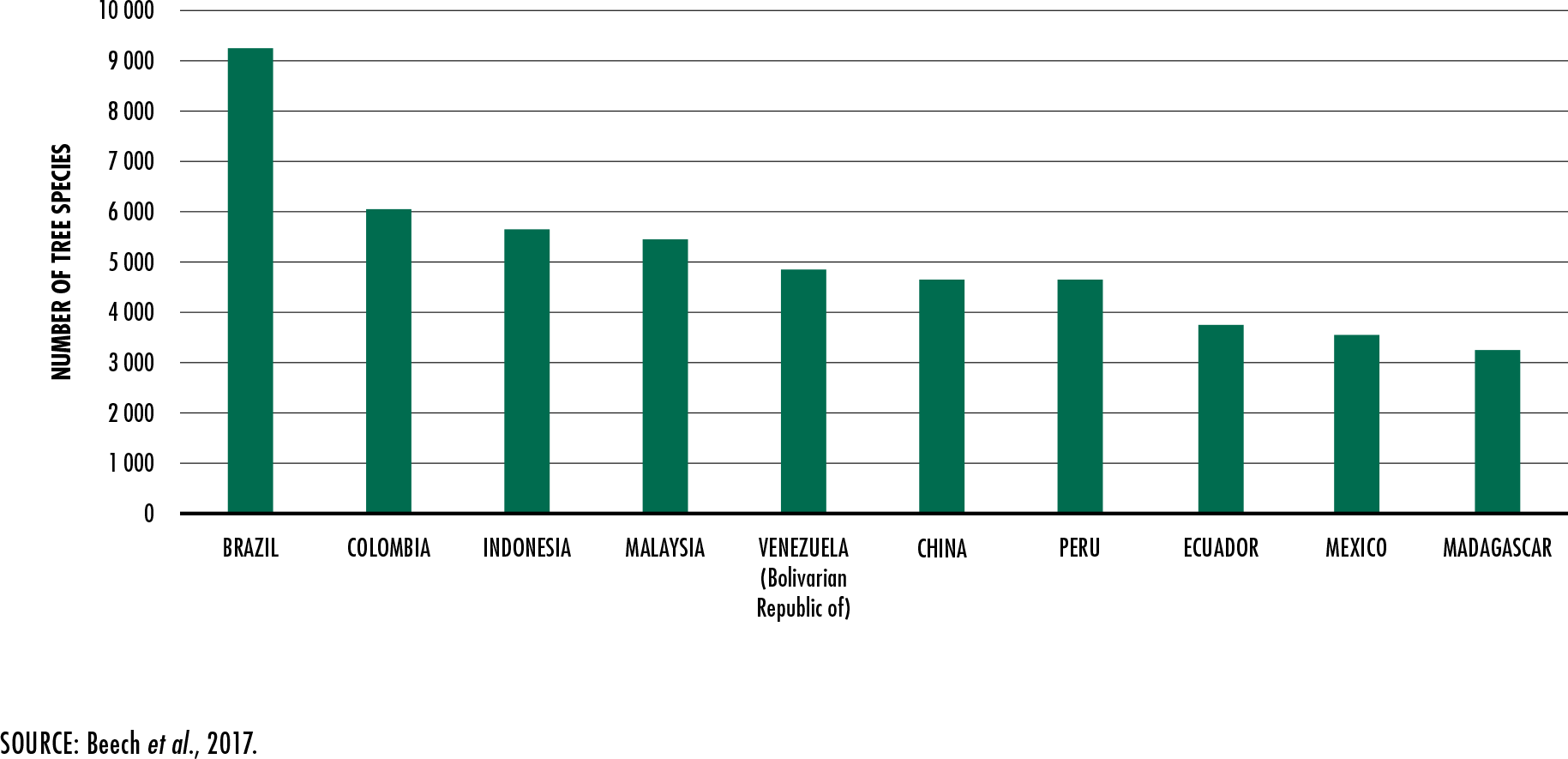
Beech, E., Rivers, M., Oldfield, S. & Smith, P.
2017. GlobalTreeSearch: the first complete global database of tree species and
country distributions.
Journal of Sustainable Forestry, 36(5): 454–489.
Bello, C., Galetti, M., Pizo, M.A., Magnago, L.F.S., Rocha, M.F., Lima,
R.A.F., Peres, C.A., Ovaskainen, O. & Jordano, P.
2015. Defaunation affects carbon storage in tropical forests. Science
Advances, 1(11): e1501105 [online]. [Cited 3 January 2020].
https://doi.org/10.1126/sciadv.1501105
Belluco, S., Halloran, A. & Ricci, A.
2017. New protein sources and food legislation: the case of edible insects and EU
law. Food Security, 9(4): 803–814.
Bengston, D.N., Butler, B.J. & Asah, S.T.
2008. Values and motivations of private forest owners in the United States: a
framework based on open-ended responses in the national woodland owner survey.
In D.B. Klenosky & C.L. Fisher, eds.
Proceedings of the 2008 Northeastern Recreation Research Symposium, pp.
60–66. General Technical Report NRS-P-42. Newtown Square, Pennsylvania, USA, USDA
Forest Service, Northern Research Station. [also available at
https://www.nrs.fs.fed.us/pubs/gtr/gtr-p-42papers/09bengston-p-42.pdf].
Benítez-López, A., Alkemade, J.R.M., Schipper, A.M., Ingram, D.J., Verweij,
P.A., Eikelboom, J. & Huijbregts, M.
2017. The impact of hunting on tropical mammal and bird populations.
Science, 356(6334): 180–183.
Bennett,
G. 2004. Integrating biodiversity conservation and sustainable use:
lessons learned from ecological networks. Gland, Switzerland, IUCN.
Bennett, G. & Mulongoy, K.J. 2006.
Review of Experience with Ecological Networks, Corridors and Buffer Zones.
Technical Series No. 23. Secretariat of the Convention on Biological Diversity,
Montreal, Canada.
Bentz, B.J., Régnière, J., Fettig, C.J., Hansen, E.M., Hayes, J.L., Hicke,
J.A., Kelsey, R.G., Negrón, J.F. & Seybold, S.J.
2010. Climate change and bark beetles of the Western United States and Canada:
Direct and indirect effects,
BioScience, 60(8): 602–613.
Berman, M., Jonides, J. & Kaplan, S.
2008. The cognitive benefits of interacting with nature.
Psychological Science, 19(12): 1207–1212.
Bernier, P.Y., Paré, D., Stinson, G., Bridge, S.R.J., Kishchuk, B.E.,
Lemprière, T.C., Thiffault, E., Titus, B.D. & Vasbinder, W.
2017. Moving beyond the concept of “primary forest” as a metric of forest
environment quality.
Ecological Applications, 27: 349–354.
BESNet. 2019. Thematic area: Biodiversity finance. In:
Biodiversity and Ecosystem Services Network
[online]. Nairobi. [Cited 3 January 2020].
https://www.besnet.world/biodiversity-finance-solutions
BGCI. 2019. GlobalTreeSearch. Botanic Gardens Conservation
International. Richmond, UK. [Cited 31 December 2019].
www.bgci.org/globaltree_search.php
Bharucha, Z. & Pretty, J. 2010. The roles and values of wild
foods in agricultural systems. Philosophical Transactions of the Royal Society
B: Biological Sciences, 365(1554): 2913–2926.
Bickford, D., Posa, M.R.C., Qie, L, Campos-Arceiz, A. & Kudavidanage,
E.P.
2012 Science communication for biodiversity conservation.
Biological Conservation, 151(1): 74–76.
Billings, R.F., Clarke, S.R., Mendoza, V.E., Cabrera, P.C., Figueroa, B.M.,
Campos, J.R. & Baeza, G.
2004. Bark beetle outbreaks and fire: A devastating combination for Central
America’s pine forests.
Unasylva, 55: 10–15.
Biodiversity Indicators Partnership. 2018. Living Planet Index
(forest specialists). In: Biodiversity Indicators Partnership [online].
Cambridge, UK. [Cited 3 January 2020].
https://www.bipindicators.net/indicators/living-planet-index/living-planet-index-forest-specialists
Biosecurity New Zealand. 2018. Biosecurity 2025 Implementation
Plan. Strengthening the biosecurity system together. Ko Tātou This
Is Us. Biosecurity New Zealand 2025. Ministry for Primary Industries, Government of
New Zealand. [also available
at: https://www.thisisus.nz/assets/Resources/163e2a594e/Biosecurity_2025_implementation_plan_full_version.pdf]
BirdLife International. 2019. World
Database on Key Biodiversity Areas [online]. [Cited 3 January 2020].
http://www.keybiodiversityareas.org/home
Blackman,
A. 2015. Strict versus mixed-use protected areas: Guatemala’s Maya
Biosphere Reserve. Ecological Economics, 112: 14–24.
Blackman, A.
& Veit, P. 2018. Titled Amazon indigenous communities cut forest
carbon emissions. Ecological Economics, 153: 56–67.
Blackman, A., Corral, L., Lima, E.S. & Asner, G.P.
2017. Titling indigenous communities protects forests in the Peruvian Amazon.
PNAS, 114(16): 4123–4128.
Blackwell, S. 2015. Resilience, wellbeing and confidence
development at forest schools. In: Get children outdoors [online]. [Cited 3
January 2020].
http://getchildrenoutdoors.com/resilience-wellbeing-and-confidence-development-at-forest-schools
Blomley, T. 2013. Lessons learned from community forestry in
Africa and their relevance for REDD+. Washington, DC, USAID-supported
Forest Carbon, Markets and Communities Program. [also available at
https://rmportal.net/library/content/fcmc/publications/CF_Africa.pdf].
Blomley, T., Pfliegner, K., Isango, J., Zahabu, E., Ahrends, A. &
Burgess, N.D.
2008. Seeing the wood for the trees: an assessment of the impact of participatory
forest management on forest condition in Tanzania. Oryx, 42(3): 380–391.
Bocci, C., Fortmann, L., Sohngen, B. & Milian, B.
2018. The impact of community forest concessions on income: an analysis of
communities in the Maya Biosphere Reserve.
World Development, 107: 10–21.
Bolognesi, M., Vrieling, A., Rembold, F., & Gadain, H.
2015. Rapid mapping and impact estimation of illegal charcoal production in southern
Somalia based on WorldView-1 imagery. Energy for Sustainable Development,
25: 40–49.
Bontemps, S., Defourny, P., Radoux, J., Van Bogaert, E., Lamarche, C.,
Achard, F., Mayaux, P.
et al. 2013. Consistent global land cover maps for climate modelling
communities: current achievements of the ESA’s land cover CCI. In Proceedings of
the ESA Living Planet Symposium, Edinburgh, UK, 9–13 September 2013, pp.
9–13. Paris, European Space Agency.
[also available at https://ftp.space.dtu.dk/pub/Ioana/papers/s274_2bont.pdf].
Borrini-Feyerabend, G., Dudley, N., Jaeger, T., Lassen, B., Pathak Broome,
N., Phillips, A. & Sandwith, T.
2013.
Governance of protected areas: from understanding to action. Best Practice
Protected Area Guidelines Series No. 20, Gland, Switzerland, IUCN.
Bowler, D.E., Buyung-Ali, L.M., Knight, T.M. & Pullin, A.S.
2010. A systematic review of evidence for the added benefits to health of exposure
to natural environments.
BMC Public Health, 10: Article number 456 [online]. [Cited 3 January 2020].
https://doi.org/10.1186/1471-2458-10-456
Boyce, M.S. 2018. Wolves for Yellowstone: dynamics in time and
space, Journal of Mammalogy, 99(5): 1021–1031.
https://doi.org/10.1093/jmammal/gyy115
Breed, M.F., Ottewell, K.M., Gardner, M.G., Marklund, M.H.K., Dormontt, E.E.
& Lowe, A.J.
2015. Mating patterns and pollinator mobility are critical traits in forest
fragmentation genetics. Heredity, 115(2): 108–114.
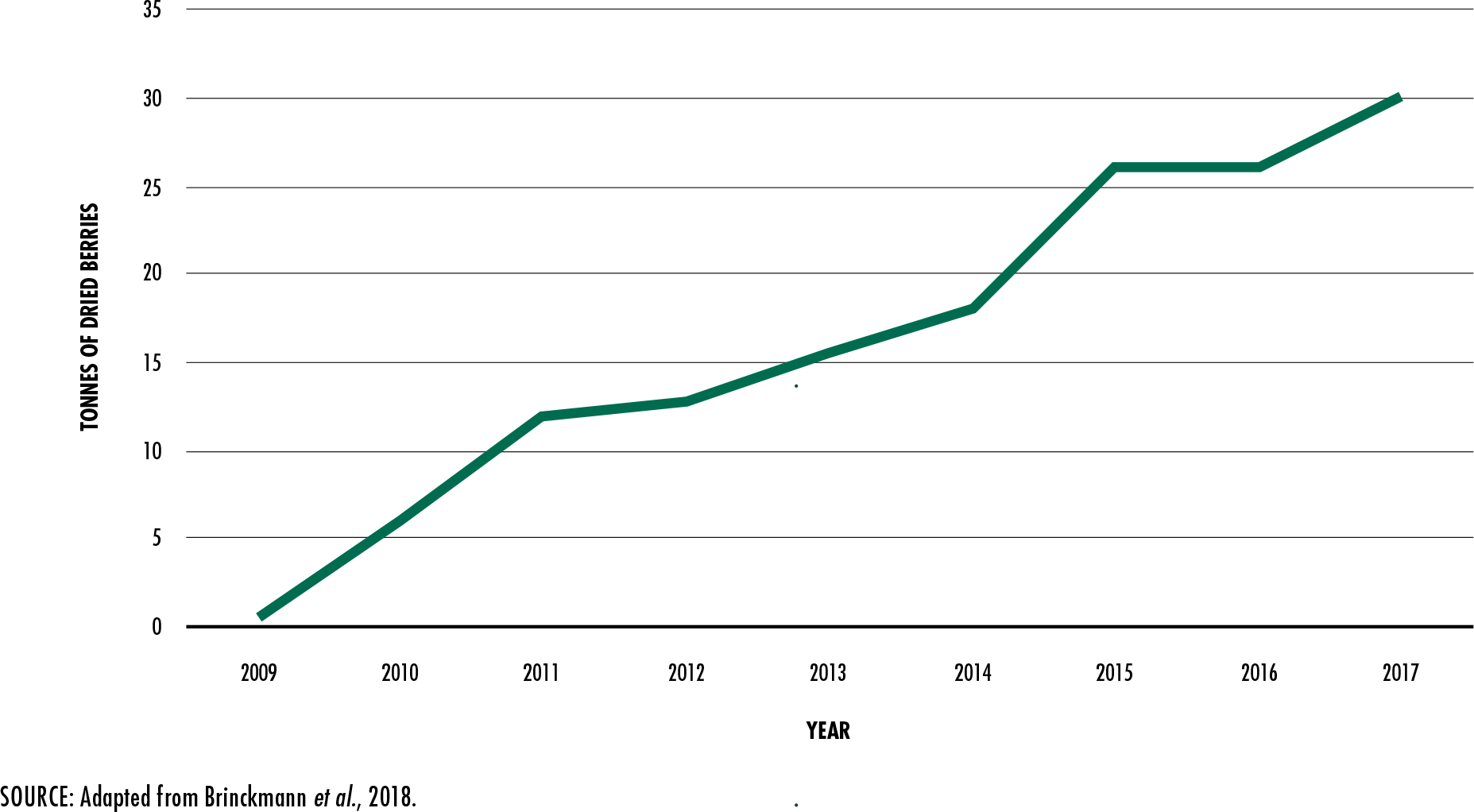
Brinckmann, J.A., Luo, W., Xu, Q., He, X., Wu, J., & Cunningham,
A.B.
2018. Sustainable harvest, people and pandas: Assessing a decade of managed wild
harvest and trade in
Schisandra sphenanthera. Journal of Ethnopharmacology, 224:
522–534.
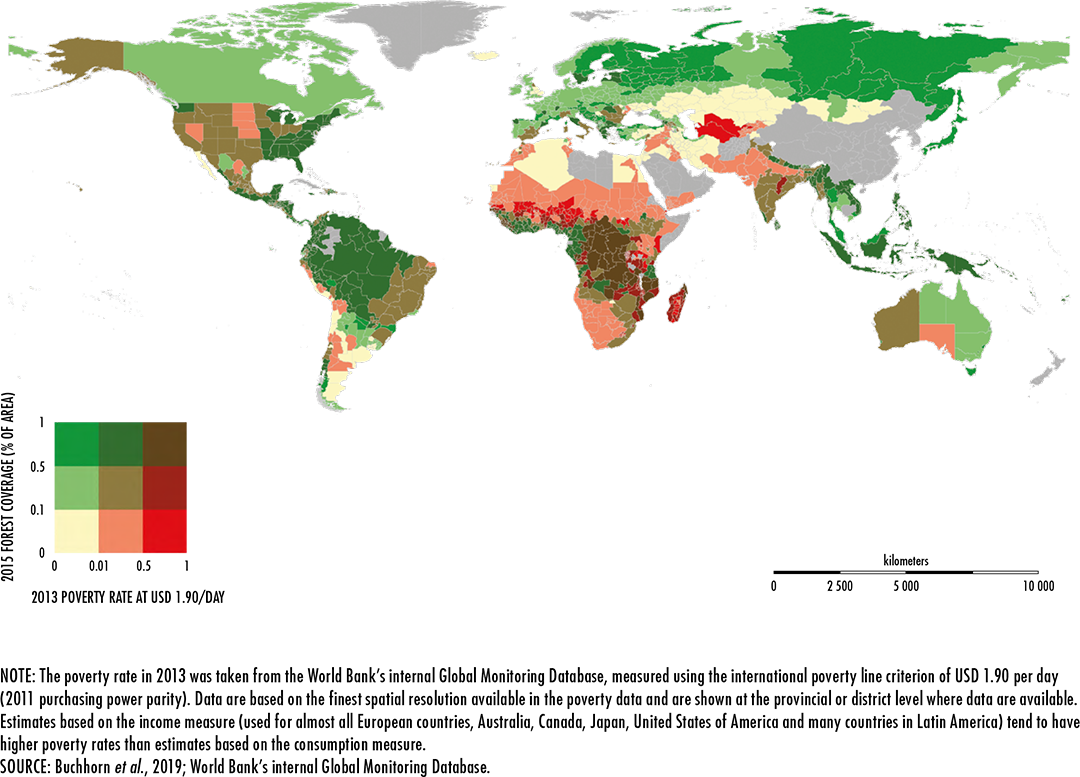
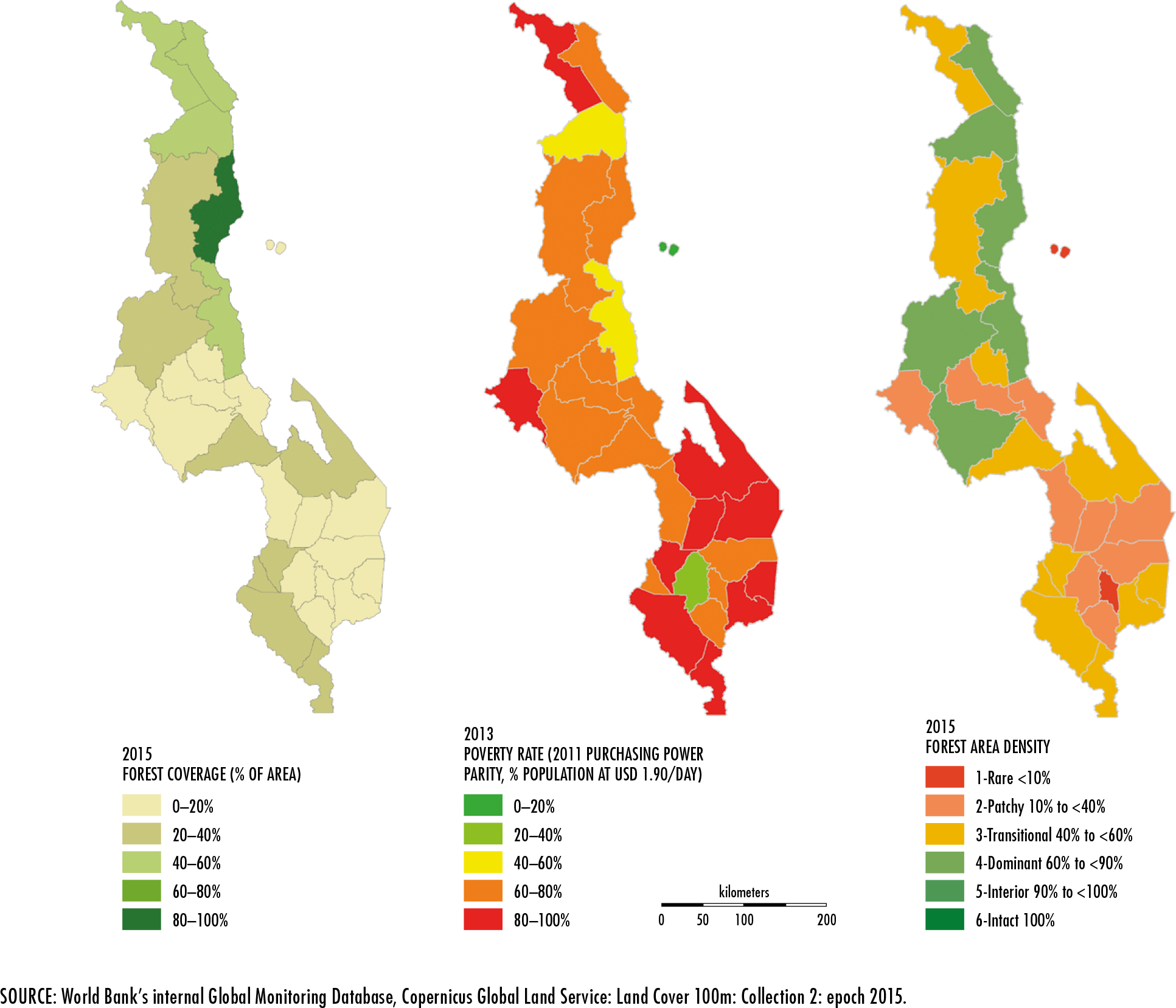
Buchhorn, M., Smets, B., Bertels, L., Lesiv, M., Tsendbazar, N.-E., Herold,
M. & Fritz, S.
2019. Copernicus Global Land Service: Land Cover 100m: epoch 2015: Globe. In:
Zenodo [online]. Geneva, Switzerland. [Cited 3 January 2020].
https://zenodo.org/record/3243509
Burgess, D., Bahane, B., Clairs, T., Danielsen, F., Dalsgaard, S., Funder,
M., Hagelberg, N.
et al. 2010. Getting ready for REDD+ in Tanzania: a case study of progress
and challenges. Oryx, 44(3): 339–351.
Burley,
J. 2002. Forest biological diversity: an overview. Unasylva,
209: 3–9.
Burlingame,
B. 2000. Editorial: Wild nutrition. Journal of Food Composition and
Analysis, 13: 99–100.
Busch, J.
& Ferretti-Gallon, K. 2017. What drives deforestation and what
stops it? A meta-analysis. Review of Environmental Economics and Policy,
11(1): 3–23.
Camara-Leret, R. & Denney, Z. 2019. Indigenous knowledge of New
Guinea’s useful plants: A review. Economic Botany, 73(3): 405–415.
Camara-Leret, R., Fortuna, M.A. & Bascompte, J.
2019. Indigenous knowledge networks in the face of global change. PNAS,
116(20): 9913–9918.
Campese, J., Sunderland, T., Greiber, T. and Oviedo, G.
(eds.) 2009.
Rights-based approaches: Exploring issues and opportunities for
conservation.
CIFOR and IUCN. Bogor, Indonesia.
Canuto, M.A., Estrada-Belli, F., Garrison, T.G., Houston, S.D., Acuña, M.J.,
Kováč, M., Marken, D.
et al. 2018. Ancient lowland Maya complexity as revealed by airborne laser
scanning of northern Guatemala. Science, 361(6409): p.eaau0137 [online].
[Cited 3 January 2020]. DOI: 10.1126/science.aau0137
Cariñanos, P., Grilo, F., Pinho, P., Casares-Porcel, M., Branquinho, C.,
Acil, N., Andreucci, M.B.
et al. 2019. Estimation of the allergenic potential of urban trees and
urban parks: towards the healthy design of urban green spaces of the future.
International Journal of Environmental Research and Public Health, 16(8):
1357 [online]. [Cited 3 January 2020].
https://doi.org/10.3390/ijerph16081357
Carnus, J.-M., Parrotta, J., Brockerhoff, E., Arbez, M., Jactel, H., Kremer,
A., Lamb, D., O’Hara, K. & Walters, B.
2006. Planted forests and biodiversity.
Journal of Forestry, 104(2): 65–77.
Carodenuto,
S. 2019. Governance of zero deforestation cocoa in West Africa: New
forms of public–private interaction. Environmental Policy and Governance,
29(1): 55–66.
Carr, D.L., Suter, L., & Barbier, A.
2005. Population dynamics and tropical deforestation: State of the debate and
conceptual challenges.
Population and Environment, 27(1): 89–113.
Castellanos, E., Regalado, O., Pérez, G., Montenegro, R., Ramos, V., &
Incer, D.
2011.
Mapa de cobertura forestal de Guatemala 2006 y dinámica de la cobertura forestal
2001–2006.
Guatemala, Universidad del Valle de Guatemala, Instituto Nacional de Bosques,
Consejo Nacional de Áreas Protegidas, Universidad Rafael Landívar.
Castello, L., Hess, L.L., Thapa, R., McGrath, D.G., Arantes, C.C., Renó,
V.F. & Isaac, V.J.
2018. Fishery yields vary with land cover on the Amazon River floodplain. Fish
and Fisheries, 19(3): 431–440.
CBD.
n.d.a. COP decisions – COP2 Decision II/9: Forests and biological diversity. In:
Convention on Biological Diversity [online]. Montreal, Canada. [Cited 19
December 2019].
https://www.cbd.int/decision/cop/?id=7082
CBD. n.d.b. What is forest biological diversity? In: Convention
on Biological Diversity [online]. Montreal, Canada. [Cited 13 December
2019].
http://www.cbd.int/forest/what.shtml
CBD. 2006. Definitions. In: Convention on Biological
Diversity [online]. Montreal, Canada. [Cited 13 January 2020].
https://www.cbd.int/forest/definitions.shtml
CBD. 2009. Invasive alien species. A threat to
biodiversity. Montreal, Canada, Secretariat of the Convention on Biological
Diversity. [also available at
https://www.cbd.int/doc/bioday/2009/idb-2009-booklet-en.pdf].
CBD. 2010a. Decision adopted by the Conference of the Parties
to the Convention on Biological Diversity at its tenth meeting. X/2. The
Strategic Plan for Biodiversity 2011–2020 and the Aichi Biodiversity
Targets. Tenth Meeting of the Conference of the
Parties to the Convention on Biological Diversity, Nagoya, Japan, 18–29 October
2010. UNEP/CBD/COP/DEC/X/2. Montreal, Canada, Secretariat of the Convention on
Biological Diversity. [also available at
https://www.cbd.int/doc/decisions/cop-10/cop-10-dec-02-en.pdf].
CBD. 2010b. Linking Biodiversity Conservation and Poverty
Alleviation: A State of Knowledge Review. CBD Technical Series No: 55.
Montreal, Canada, Secretariat of the Convention on Biological Diversity. [also
available at
https://www.cbd.int/doc/publications/cbd-ts-55-en.pdf].
CBD. 2011. Nagoya Protocol on Access to Genetic Resources and
the Fair and Equitable Sharing of Benefits Arising from their Utilization to the
Convention on Biological Diversity - Text and Annex. Montreal, Canada,
Secretariat of the Convention on
Biological Diversity.
CBD. 2012a.
Resourcing the biodiversity targets: A first assessment of the resources
required for implementing the strategic plan for biodiversity 2011–2020.
Montreal, Canada, Secretariat of the Convention on Biological Diversity. [also
available at
https://www.cbd.int/doc/meetings/fin/hlpgar-sp-01/official/hlpgar-sp-01-01-report-en.pdf].
CBD. 2012b. Cities and biodiversity outlook. Montreal,
Canada, Secretariat of the Convention on Biological Diversity.
CBD. 2014. Global Biodiversity Outlook 4. Montreal,
Canada, Secretariat of the Convention on Biological Diversity. [also available at
https://www.cbd.int/gbo/gbo4/publication/gbo4-en-hr.pdf].
CBD. 2016a.
Ecosystem restoration: short-term action plan.
Decision XIII/5 of the Conference of the Parties to the Convention on Biological
Diversity. Montreal, Canada, Secretariat of the Convention on Biological Diversity.
[also available at
https://www.cbd.int/doc/decisions/cop-13/cop-13-dec-05-en.pdf].
CBD. 2016b. Updated assessment of progress towards Aichi
Biodiversity Targets 5 and 15. Thirteenth Meeting of the Conference of the
Parties to the Convention on Biological Diversity, Cancun, Mexico, 4–17 December
2016. UNEP/CBD/COP/13/INF/12. Montreal,
Canada, Secretariat of the Convention on Biological Diversity.
CBD. 2017. The ABS Clearing-House. In: Convention on Biological
Diversity [online]. Montreal, Canada. [Cited 26 December 2019].
https://www.cbd.int/abs/theabsch.shtml
CBD. 2018a. Decision adopted by the Conference of the Parties
to the Convention on Biological Diversity. 14/8. Protected areas and other
effective area-based conservation measures. 14th meeting of the Conference
of the Parties to the CBD, Sharm
El-Sheikh, Egypt, 17–29 November 2018. CBD/COP/DEC/14/8. Montreal, Canada,
Secretariat of the Convention on Biological Diversity. [also available at
https://www.cbd.int/doc/decisions/cop-14/cop-14-dec-08-en.pdf].
CBD. 2018b.
Decision adopted by the Conference of the Parties to the Convention on
Biological Diversity. 14/7. Sustainable wildlife management.
14th meeting of the Conference of the Parties, Sharm el-Sheikh, Egypt, 17–29
November 2018. CBD/COP/DEC/14/7. Montreal, Canada, Secretariat of the Convention on
Biological Diversity. [also available at
https://www.cbd.int/doc/decisions/cop-14/cop-14-dec-07-en.pdf].
CBD. 2018c. Progress of the application of the Singapore Index
on Cities’ Biodiversity. Note by the Executive Secretary. 14th meeting of
the Conference of the Parties, Sharm el-Sheikh, Egypt, 17–29 November.
CBD/COP/14/INF/34. Montreal, Canada,
Secretariat of the Convention on Biological Diversity.
CBD. 2019. The Nagoya Protocol on Access and Benefit-sharing. In:
Convention on Biological Diversity [online]. Montreal, Canada. [Cited 19
December 2019].
https://www.cbd.int/abs/
CBD. 2020a. Parties to the Nagoya Protocol. In: Convention on
Biological Diversity [online]. Montreal, Canada. [Cited 13 January 2020].
https://www.cbd.int/abs/nagoya-protocol/signatories/
CBD. 2020b.
The Access and Benefit-Sharing Clearing-House
[online]. Montreal, Canada. [Cited 13 January 2020].
https://absch.cbd.int/
CEPF. 2020. Biodiversity hotspots defined. In: Critical
Ecosystem Partnership Fund [online]. Arlington, VA, USA. [Cited 13 January
2020].
https://www.cepf.net/our-work/biodiversity-hotspots/hotspots-defined
Ceres. 2019. Out on a limb: The state of corporate
no-deforestation commitments and reporting indicators that count. Boston,
MA, USA. [also available at
www.ceres.org/sites/default/files/reports/2019-06/OutOnaLimb.pdf].
CFS. 2014. Principles for
responsible investment in agriculture and food systems. Rome. [also
available at
http://www.fao.org/3/a-au866e.pdf].
CGRFA. 2019. First report on the implementation of the Global
Plan of Action for the Conservation, Sustainable Use and Development of Forest
Genetic Resources. 17th regular session, Rome, 18–22 February 2019.
CGRFA-17/19/10.2/Inf.1. Rome. [also available
at http://www.fao.org/3/my877en/my877en.pdf].
Chan, K.M.A., Pringle, R.M., Ranganathan, J., Boggs, C.L., Chan, Y.L.,
Ehrlich, P.R., Haff, P.K., Heller, N.E, Al-Khafaji, K. & Macmynowski,
D.P.
2007. When agendas collide: human welfare and biological conservation.
Conservation Biology, 21(1): 59–68.
Chan, L., Hillel, O., Elmqvist, T., Werner, P., Holman, N., Mader, A. &
Calcaterra, E.
2014.
User’s manual on the Singapore Index on Cities’ Biodiversity (also known as the
City Biodiversity Index). Singapore, National Parks Board, Singapore.
Chao, S. 2012.
Forest peoples: numbers across the world.
Moreton-in-Marsh, UK, Forest Peoples Programme.
Chazdon, R.L., Bodin, B., Guariguata, M., Lamb, D., Walder, B.,
Chokkalingam, U. & Shono, K.
2017.
Partnering with nature: The case for natural regeneration in forest and
landscape restoration. FERI Policy Brief. Montreal, Canada, FERI.
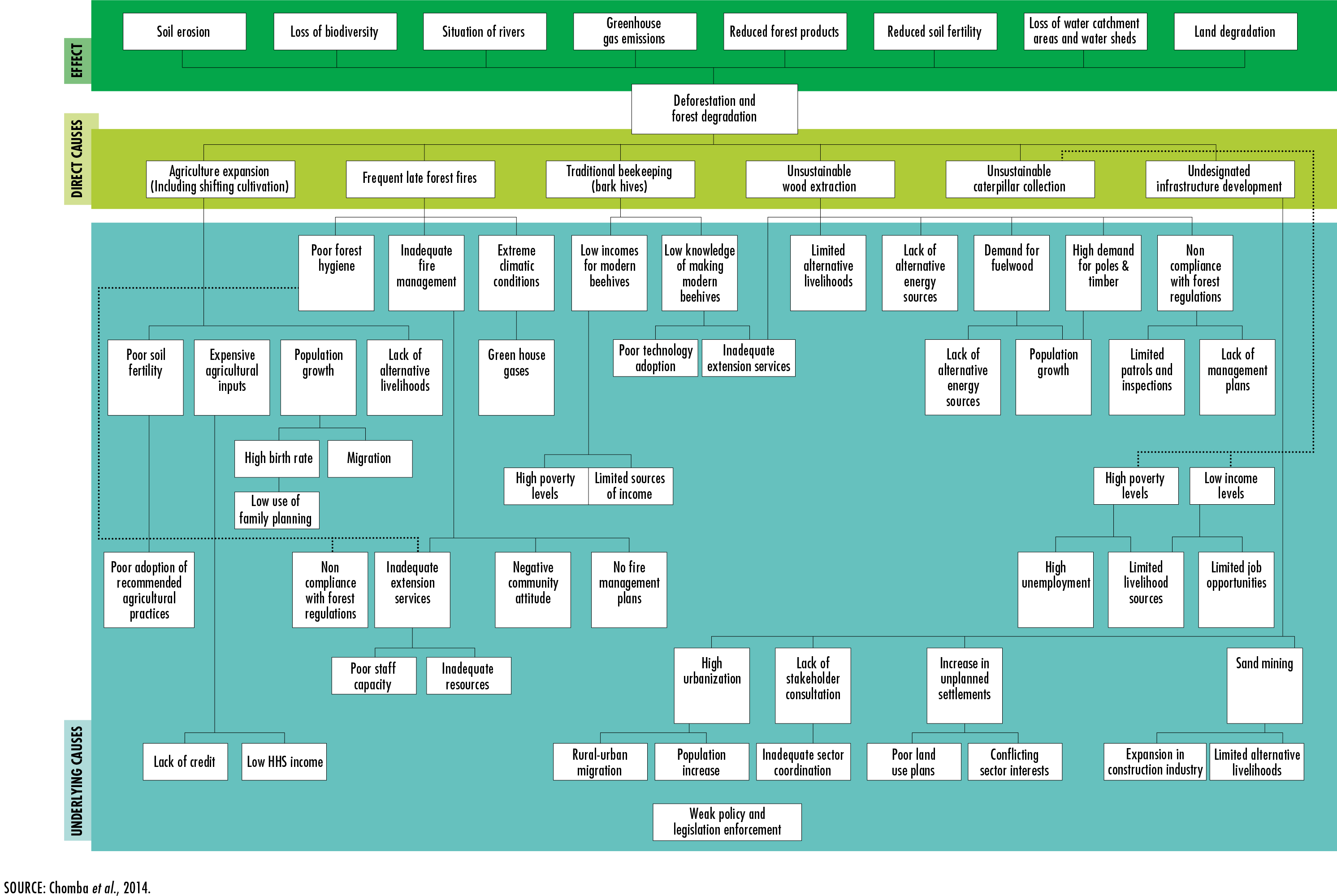
Chomba, B.M., Tembo, O., Mutandi, K., Mtongo, C.S. & Makano, A.
2014.
Drivers of deforestation, identification of threatened forests and forest
co-benefits other than carbon from REDD+ implementation in Zambia. A
consultancy report prepared for the Forestry Department and the Food and Agriculture
Organization of the United Nations under
the national UN-REDD Programme. Lusaka, Ministry of Lands, Natural Resources and
Environmental Protection. [also available at
http://landforlions.org/data/documents/drivers-deforestation-Zambia-WEB_final.pdf].
CITES. 1983.
Convention on International Trade in Endangered Species of Wild Flora and
Fauna
[online]. [Cited 19 December 2019].
https://www.cites.org/sites/default/files/eng/disc/CITES-Convention-EN.pdf
CITES. 2019. Projects and initiatives – Supporting sustainable
management of endangered tree species. In:
Convention on International Trade in Endangered Species of Wild Flora and
Fauna
[online]. Geneva, Switzerland. [Cited 4 January 2020].
https://www.cites.org/eng/prog/flora/trees/trees_project
Clean Cooking Alliance. 2015. Five years of impact 2010–2015. In:
Clean Cooking Alliance [online]. New York, USA, United Nations Foundation.
[Cited 4 January 2020].
https://www.cleancookingalliance.org/resources/reports/fiveyears.html
Coad, L., Fa, J., Abernathy, K., Van Vliet, N., Santamaria, C., Wilkie,
D.S., El Biziri, H.R., Ingram, D.J., Cawthorn, D. & Nasi, R.
2019.
Towards a sustainable, participatory and inclusive wildmeat sector. Bogor,
Indonesia, CIFOR.
Coady, D., Parry, I., Le, N.-P. & Shang, B.
2019.
Global fossil fuel subsidies remain large: an update based on country-level
estimates. IMF Working Paper. Washington, DC, IMF.
COMIFAC. 2020. Commission des Forêts d’Afrique Centrale
[online]. Yaoundé. [Cited 2 January 2020].
https://comifac.org/
CONAP & WCS. 2018. Monitoreo de la Gobernabilidad en la
Reserva de la Biosfera Maya: Actualización al año 2017. Con el apoyo de
USAID y el USDOI/ITAP. 56 pp. San Benito, Petén, Guatemala. [also available at:
https://conap.gob.gt/wp-content/uploads/2019/10/MONITOREO-DE-LA-GOBERNABILIDAD-EN-LA-RBM.pdf]
Cook, B., Anchukaitis, K., Kaplan, J., Puma, M., Kelley, M. & Gueyffier,
D.
2012. Pre-Columbian deforestation as an amplifier of drought in Mesoamerica.
Geophysical Research Letters, 39(16): L16706 [online]. [Cited 4 January
2020].
https://doi.org/10.1029/2012GL052565
CPW. 2016. Sustainable wildlife management and human–wildlife
conflict. CPW Fact Sheet 4. Rome. [also available at http://www.fao.org/3/a-i4893e.pdf].
CRITFC. 2020. The Plan: Wy-Kan-Ush-Mi Wa-Kish-Wit. In:
Colombia River Inter-Tribal Fish Commission
[online]. Portland, OR, USA. [Cited 1 January 2020].
https://www.critfc.org/fish-and-watersheds/fish-and-habitat-restoration/the-plan-wy-kan-ush-mi-wa-kish-wit/
Dargie, G.C., Lewis, S.L., Lawson, I.T., Mitchard, E.T.A., Page, S.E.,
Bocko, Y.E. & Ifo, S.A.
2017. Age, extent and carbon storage of the central Congo Basin peatland complex.
Nature, 542(7639): 86–90.
Dave, R., Saint-Laurent, C., Murray, L., Antunes Daldegan, G., Brouwer, R.,
de Mattos Scaramuzza, C.A., Raes, L.
et al. 2019. Second Bonn Challenge progress report – application of the
barometer in 2018. Gland, Switzerland, IUCN.
Davies, J.D., Hill, R., Walsh, F., Sandford, M., Smyth, D. & Holmes,
M.C.
2013. Innovation in management plans for community conserved areas: Experiences from
Australian indigenous protected areas. Ecology and Society, 18(2): 14
[online]. [Cited 4 January 2020].
http://dx.doi.org/10.5751/ES-05404-180214
Dawson, I.K., Leakey, R., Clement, C.R., Weber, J.C., Cornelius, J.P.,
Roshetko, J.M., Vinceti, B.
et al. 2014. The management of tree genetic resources and the livelihoods
of rural communities in the tropics: Non-timber forest products, smallholder
agroforestry practices and tree commodity crops. Global Forest Genetic
Resources, 333: 9–21.
Deacon,
R.T. 1995. Assessing the relationship between government policy and
deforestation. Journal of Environmental Economics and Management,
28(1):1–18.
Delelegn, A., Sahile, S. & Husen, A.
2018. Water purification and antibacterial efficacy of
Moringa oleifera Lam. Agriculture and Food Security, 7: Article 25
[online]. [Cited 4 January 2020].
https://doi.org/10.1186/s40066-018-0177-1
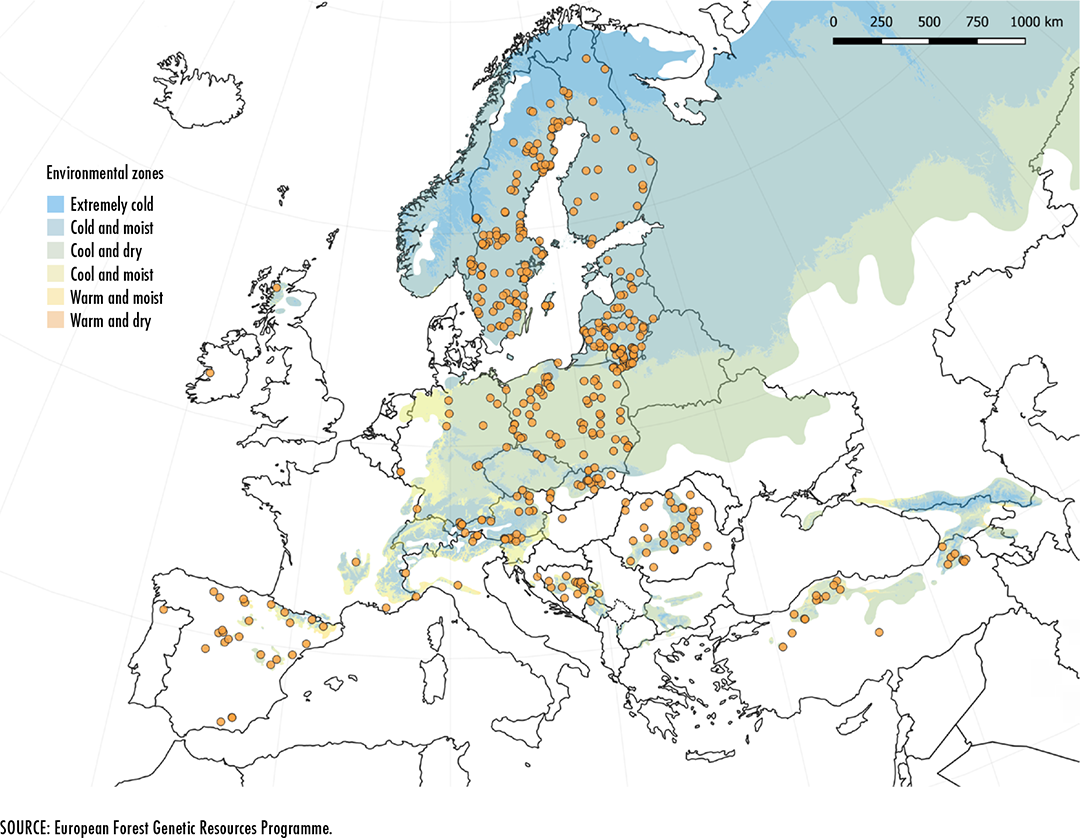
de Vries, S.M.G., Alan, M., Bozzano, M., Burianek, V., Collin, E., Cottrell,
J., Ivankovic, M.
et al. 2015. Pan-European strategy for genetic conservation of forest
trees and establishment of a core network of dynamic conservation units.
European Forest Genetic Resources Programme (EUFORGEN). Rome, Bioversity
International.
Ding, H., Veit, P.G., Blackman, A., Gray, E.,
Reytar, K., Altamirano, J.C. & Hodgdon, B.
2016.
Climate benefits, tenure costs: the economic case for securing indigenous land
rights in the Amazon. Washington, DC, WRI.
Dirzo, R.
& Raven, P.H. 2003. Global state of biodiversity and loss.
Annual Review of Environment and Resources, 28: 137–167.
Dounias, E.
& Ichikawa, M. 2017. Seasonal bushmeat hunger in the Congo Basin.
EcoHealth, 14: 575–590.
Dourojeanni, M. 2017. [Opinión] ¿Las sociedades prehispánicas
cuidaron mejor la Amazonía? In: SPDA Actualidad Ambiental [online]. Lima.
[Cited 4 January 2020].
www.actualidadambiental.pe/opinion-las-sociedades-preshispanicas-cuidaron-mejor-la-amazonia/
Drescher, M. & Brenner, J.C. 2018. The practice and promise of
private land conservation. Ecology and Society 23(2) [online]. [Cited 4
January 2020].
www.jstor.org/stable/26799076
Dudley, N., Jonas, H., Nelson, F., Parrish, J., Pyhälä, A., Stolton, S.
& Watson, J.
2018. The essential role of other effective area-based conservation measures in
achieving big bold conservation targets. Global Ecology and Conservation,
15: e00424 [online]. [Cited 4 January 2020].
https://doi.org/10.1016/j.gecco.2018.e00424
Duffy, J., Godwin, C. & Cardinale, B.
2017. Biodiversity effects in the wild are common and as strong as key drivers of
productivity. Nature, 549: 261–264.
EC. 2019a. Communication from the Commission to the European
Parliament, the Council, the European Economic and Social Committee and the
Committee of the Regions: Stepping up EU action to protect and restore the
world’s forests. COM(2019) 352 final.
Brussels. (also available at https://ec.europa.eu/info/sites/info/files/communication-eu-action-protect-restore-forests_en.pdf).
EC. 2019b. Nature and biodiversity – Natura 2000. In: European
Commission, Environment [online]. Brussels. [Cited 4 January 2020].
https://ec.europa.eu/environment/nature/natura2000/index_en.htm
Ege, M.J., Mayer, M., Normand, A.C., Genuneit, J., Cookson, W.O.,
Braun-Fahrländer, C., Heederik, D., Piarroux, R. & von Mutius, E.
2011. Exposure to environmental microorganisms and childhood asthma. The New
England Journal of Medicine, 364: 701–709.
Eilers, E.J., Kremen, C., Smith Greenleaf, S., Garber, A.K. & Klein,
A.-M.
2011. Contribution of pollinator-mediated crops to nutrients in the human food
supply. PLOS ONE, 6(6): e21363 [online]. [Cited 13 January 2020].
https://doi.org/10.1371/journal.pone.0021363
Eriksson, M., Samuelson, L., Jägrud, L., Mattsson, E., Celander, T., Malmer,
A., Bengtsson, K.
et al. 2018. Water, forests, people: The Swedish Experience in building
resilient landscapes. Environmental Management, 62(1): 45–57.
Erwin, T.L. 1982. Tropical forests: their richness in Coleopteran
and other arthropod species. The Coleopterists’ Bulletin, 36: 74–75., cited
by Dirzo, R. & Raven, P. H. 2003. Global state of biodiversity and loss.
Annual Review of Environment and Resources, 28: 137–167.
ESA CCI. 2017. Global Land Cover Maps for 2015. In:
Land Cover CCI Climate Research Data Package
[online]. ESA Climate Change Initiative – Land Cover led by UCLouvain.
https://www.esa-landcover-cci.org/?q=node/164
EU. 2011. Voluntary Partnership Agreement between the European
Union and the Republic of Cameroon on forest law enforcement, governance and trade
in timber and derived products to the European Union (FLEGT). 6 April.
Official Journal of the European Union, 92: 4–125.
EU FLEGT Facility. n.d.
FLEGT licensed timber – Essential information
[online]. Brussels. [Cited 4 January 2020].
www.flegtlicence.org/home
Evans, N.P., Bauska, T.K., Gázquez-Sánchez, F., Brenner, M., Curtis, J.H.
& Hodell, D.A.
2018. Quantification of drought during the collapse of the classic Maya
civilization. Science, 361(6401): 498–501.
Fa, J.E., Currie, D. & Meeuwig, J.
2003. Bushmeat and food security in the Congo Basin: linkages between wildlife and
people’s future.
Environmental Conservation, 30: 71–78.
Fabricant, D.S. & Fransworth, N.R.
2001. The value of plants used in traditional medicine for drug discovery.
Environmental Health Perspectives, 109(1): 69–75.
FairWild Foundation. 2019. The FairWild standard [online].
Cambridge, UK. [Cited 18 December 2019].
https://www.fairwild.org/the-fairwild-standard
FAO. 1989. Forestry and food security. FAO Forestry Paper
No. 90. Rome. [also available at http://www.fao.org/3/T0178E/T0178E00.htm].
FAO. 1997. The State of the World’s Plant Genetic Resources for
Food and Agriculture. Rome. [also available at http://www.fao.org/tempref/docrep/fao/meeting/015/w7324e.pdf].
FAO. 2006.
Fire management: voluntary guidelines. Principles and strategic actions.
Fire Management Working Paper 17. Rome. [also available at
http://www.fao.org/3/j9255e/j9255e00.htm].
FAO. 2007. The State of the World’s Animal Genetic Resources
for Food and Agriculture. Rome. [also available at http://www.fao.org/3/a-a1250e.pdf].
FAO. 2009.
Declaration of the World Food Summit on Food Security
[online]. Rome. [Cited 4 January 2020].
http://www.fao.org/3/w3613e/w3613e00.htm
FAO. 2010a. The Second Report on the State of the World’s Plant
Genetic Resources for Food and Agriculture. Rome. [also available at http://www.fao.org/3/i1500e/i1500e.pdf].
FAO. 2010b. Global Forest Resources Assessment 2010 – Main
report. FAO Forestry Paper No. 163. Rome. [also available at http://www.fao.org/3/i1757e/i1757e00.htm].
FAO. 2011a. International Plant Protection Convention.
Rome, Secretariat of the International Plant Protection Convention. [also available
at
https://www.ippc.int/static/media/files/publication/en/2019/02/1329129099_ippc_2011-12-01_reformatted.pdf].
FAO. 2011b. State of the World’s
Forests 2011. Rome. [also available at http://www.fao.org/3/i2000e/i2000e00.htm].
FAO. 2012a. Global ecological zones for FAO forest reporting:
2010 Update. Forest Resources Assessment Working Paper 179. Rome. [also
available at http://www.fao.org/3/ap861e/ap861e00.pdf].
FAO. 2012b. Voluntary guidelines on the responsible governance
of tenure of land, fisheries and forests in the context of national food
security. Rome. [also available at http://www.fao.org/3/a-i2801e.pdf].
FAO. 2013a. Forests and trees outside forests are essential for
global food security and nutrition. Summary of the International Conference
on Forests for Food Security and Nutrition, Rome, 13–15 May 2013. Rome. [also
available at
http://www.fao.org/3/aq110e/aq110e.pdf].
FAO. 2013b. Edible insects – future prospects for food and feed
security. FAO Forestry Paper No. 171. Rome. [also available at http://www.fao.org/3/i3253e/i3253e.pdf].
FAO. 2013c. Six-legged livestock: edible insect farming,
collection and marketing in Thailand. RAP Publication No. 2013/03. Bangkok,
Thailand, Food and Agriculture Organization of the United Nations, Regional Office
for Asia and the Pacific. [also
available at http://www.fao.org/3/a-i3246e.pdf].
FAO. 2014a.
The State of the World’s Forest Genetic Resources.
Rome. [also available at
http://www.fao.org/forestry/fgr/64582/en/].
FAO. 2014b. Global plan of action for forest genetic
resources. Rome. (available at http://www.fao.org/3/a-i3849e.pdf).
FAO. 2014c. State of the World’s Forests 2014. Rome. [also
available at http://www.fao.org/3/a-i3710e.pdf].
FAO. 2014d. Women in forestry: Challenges and
opportunities. Rome. [also available at http://www.fao.org/3/a-i3924e.pdf].
FAO. 2015a. The Second Report on the State of the World’s
Animal Genetic Resources for Food and Agriculture. Rome. [also available at
http://www.fao.org/3/a-i4787e.pdf].
FAO. 2015b. Global guidelines for the restoration of degraded
forests and landscapes in drylands: building resilience and benefiting
livelihoods. Forestry Paper No. 175. Rome. [also available at
http://www.fao.org/3/a-i5036e.pdf].
FAO. 2015c.
Global Forest Resources Assessment 2015. How are the world’s forest
changing?
Rome. 2nd edition. Rome. [also available at
http://www.fao.org/3/a-i4793e.pdf].
FAO. 2016a. Follow-up to the second International Conference on
Nutrition. 23rd session of the Committee on Forestry, Rome, 18–22 July
2016. COFO/2016/7.4. Rome. [also available at
www.fao.org/3/a-mq485e.pdf].
FAO. 2016b. Payments for forest environmental services in
sub-Saharan Africa: a practical guide. Accra, FAO. [also available at http://www.fao.org/3/a-i5578e.pdf].
FAO. 2017a.
Sustainable woodfuel for food security. A smart choice: green, renewable and
affordable.
Working paper. Rome. [also available at
http://www.fao.org/3/a-i7917e.pdf].
FAO. 2017b. Strengthening sector policies for better food
security and nutrition results – Forestry. Policy Guidance Note 3. Rome.
[also available at http://www.fao.org/3/a-i7215e.pdf].
FAO. 2017c. Non-wood forest products in international
statistical systems. Rome. [also available at http://www.fao.org/3/a-i6731e.pdf].
FAO. 2017d. The Agadir commitment towards a Mediterranean
regional initiative on forest and landscape restoration. AFWC/EFC/NEFC
Committee on Mediterranean Forestry Questions – Silva Mediterranea, 22nd
session, Agadir, Morocco, 22 March 2017.
[also available at www.fao.org/forestry/45685-0ad87e3a1d4ccc359b37c38ffcbb5b1fc.pdf].
FAO. 2017e. The future of food and agriculture – Trends and
challenges. Rome. [also available at http://www.fao.org/3/a-i6583e.pdf].
FAO. 2018a. Terms and definitions: FRA 2020. Forest
Resources Assessment Working Paper 188. Rome. [also available at http://www.fao.org/3/I8661EN/i8661en.pdf].
FAO. 2018b. State of the World’s Forests 2018. Rome. [also
available at http://www.fao.org/3/I9535EN/i9535en.pdf].
FAO. 2018c. REDD+ finance and investments. Rome. [also
available at http://www.fao.org/3/CA0907EN/ca0907en.pdf].
FAO. 2019a. The State of the World’s Biodiversity for Food and
Agriculture. Rome, FAO and Commission on Genetic Resources for Food and
Agriculture. [also available at
http://www.fao.org/3/CA3129EN/CA3129EN.pdf].
FAO. 2019b. The State of the World’s Aquatic Genetic Resources
for Food and Agriculture. Rome. [also available at http://www.fao.org/3/CA5256EN/CA5256EN.pdf].
FAO. 2019c. Trees, forests and land use in drylands: the first
global assessment – Full report. FAO Forestry Paper No. 184. Rome. [also
available at http://www.fao.org/3/ca7148en/ca7148en.pdf].
FAO. 2019d.
International Treaty on Plant Genetic Resources for Food and Agriculture
[online]. [Cited 13 January 2020].
http://www.fao.org/fileadmin/user_upload/legal/docs/033s-e.pdf
FAO. 2019e. FAOSTAT. In:
Food and Agriculture Organization of the United Nations
[online]. Rome. [Cited 4 January 2020].
www.fao.org/faostat
FAO. 2019f. Collaborative Partnership on Sustainable Wildlife
Management. In:
Food and Agriculture Organization of the United Nations
[online]. Rome. [Cited 18 December 2019].
http://www.fao.org/forestry/wildlife-partnership/en/
FAO. 2019g. Restoring forest landscapes through assisted
natural regeneration (ANR) – A practical manual. Bangkok. [also available
at http://www.fao.org/3/ca4191en/CA4191EN.pdf]
FAO. 2019h. Action Against
Desertification. In:
Food and Agriculture Organization of the United Nations
[online]. Rome. [Cited 4 January 2020].
http://www.fao.org/in-action/action-against-desertification
FAO. 2019i. Championing sustainable agriculture in the
Caribbean Region of Colombia: a case study. Rome. [also available at www.fao.org/3/ca6753en/CA6753EN.pdf].
FAO. 2019j.
Sustainable Food and Agriculture – An Integrated Approach,
by Campanhola, C. and Pandey, S. (eds). FAO and Elsevier.
FAO. 2020. Global Forest Resources Assessment 2020 – Main
report. Rome.
FAO. forthcoming. Analysis of 32 REDD+ Strategies. Rome.
FAO & CPF. 2018. A joint initiative of the Collaborative
Partnership on Forests (CPF). Co-chairs summary report. Presented to the
international conference on Working across Sectors to Halt Deforestation and
Increase Forest Area – From Aspiration to
Action, FAO headquarters, Rome, 20–22 February 2018.
FAO & Global Mechanism of UNCCD. 2015. Sustainable
financing for forest and landscape restoration: Opportunities, challenges and
the way forward. Rome, FAO. [also available at
http://www.fao.org/3/a-i5174e.pdf].
FAO & Plan Bleu. 2018. State of Mediterranean forests
2018. Rome, FAO, and Marseille, France, Plan Bleu. [also available at http://www.fao.org/3/CA2081EN/ca2081en.PDF].
FAO & WRI. 2019. The road to restoration: a guide to
identifying priorities and indicators for monitoring forest and landscape
restoration, by Kathleen Buckingham, Sabin Ray, Carolina Gallo Granizo,
Lucas Toh, Fred Stolle, Faustine Zoveda, Katie
Reytar, Rene Zamora, Peter Ndunda, Florence Landsberg, Marcelo Matsumoto & John
Brandt. Washington, DC, USA.
FAO, DFSC & IPGRI. 2001.
Forest genetic resources conservation and management. Vol. 2: In managed natural
forests and protected areas
(in situ). Rome, IPGRI.
FAO, FLD & IPGRI. 2004. Forest genetic resources
conservation and management. Vol. 3: In plantations and genebanks (ex
situ). Rome, International Plant Genetic Resources Institute.
Fedigan, L.M. & Jack, K.M. 2012. Tracking neotropical Monkeys
in Santa Rosa: Lessons from a regenerating Costa Rican dry forest. In P.M.
Kappeler & D.P. Watts, eds. Long-term field studies of primates, pp.
165–184. Berlin, Springer.
Ferraro, P., Sanchirico, J., & Smith, M.
2019. Causal inference in coupled human and natural systems,
PNAS, 116(12): 5311–5318.
Field, C.D.,
ed. 1996. Restoration of mangrove ecosystems. Okinawa, Japan,
International Society for Mangrove Ecosystems.
Fisher, B.
& Christopher, T. 2007. Poverty and biodiversity: Measuring the
overlap of human poverty and the biodiversity hotspots. Ecological
Economics, 62: 93–101.
Fluet-Chouinard, E., Funge-Smith, S. & McIntyre, P.B.
2018. Global hidden harvest of freshwater fish revealed by household surveys.
PNAS, 115(29): 7623–7628.
FONAFIFO, CONAFOR and Ministry of Environment.
2012.
Lessons learned for REDD+ from PES and conservation incentive programs. Examples
from Costa Rica, Mexico, and Ecuador. Washington, DC, The International
Bank for Reconstruction and Development/The World Bank.
Food and Land Use Coalition. 2019.
Ten Critical Transitions to Transform Food and Land Use.
[also available at
https://www.foodandlandusecoalition.org/wp-content/uploads/2019/09/FOLU-GrowingBetter-GlobalReport.pdf]
Forest Europe. n.d. Home page [online]. Zvolen, Slovakia.
[Cited 26 December 2019].
https://foresteurope.org/
Forest Europe. 2019. Human health and sustainable forest
management, edited by Ľ. Marušáková & M. Sallmannshoferet. Forest
Europe Study. Zvolen, Slovak Republic. [also available at
https://foresteurope.org/wp-content/uploads/2017/08/Forest_book_final_WEBpdf.pdf]
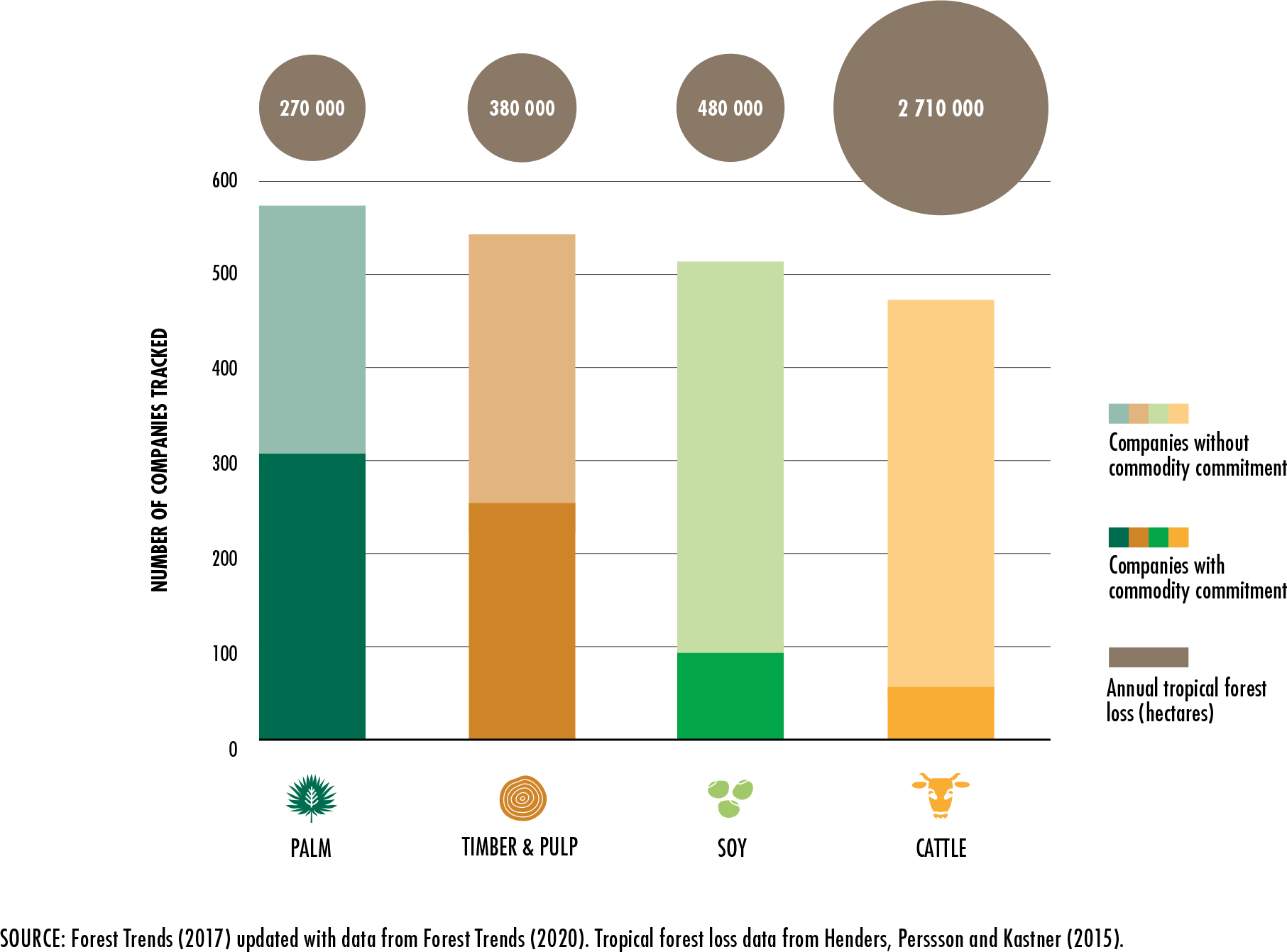
Forest
Trends. 2017. Supply change: Tracking corporate commitments to
deforestation-free supply chains, 2017. Washington, DC.
Forest Trends. 2020. Forest Trends Supply Change
Initiative [online] [Cited 17 March, 2020].
http://supply-change.org/
Fritz-Vietta, N.V.M. 2016. What can forest values tell us about
human well-being? Insights from two biosphere reserves in Madagascar. Landscape
and Planning 147: 28–37.
Fung, E., Imbach, P., Corrales, L., Vilchez, S. Zamora, N., Argotty, F.,
Hannah, L. & Ramos, Z.
2017. Mapping conservation priorities and connectivity pathways under climate change
for tropical ecosystems.
Climatic Change 141: 77–92.
Gaisberger, H., Kindt, R., Loo, J., Schmidt, M., Bognounou, F., Da, S.S.,
Diallo, O.B.
et al. 2017. Spatially explicit multi-threat assessment of food tree
species in Burkina Faso: A fine-scale approach. PLOS ONE, 12(9): e0184457
[online]. [Cited 4 January 2020].
https://doi.org/10.1371/journal.pone.0184457
Galetti, M.
& Costa-Pereira, R. 2017. Scientists need social media influencers.
Science, 357(6354): 880–881.
Galway, L.P., Acharya, Y. & Jones, A.D.
2018. Deforestation and child diet diversity: A geospatial analysis of 15
sub-Saharan African countries.
Health & Place, 51: 78–88.
Gardner, C.J., Bicknell, J.E., Struebig, M.J., & Davies, Z.G.
2017.
Vertebrate populations, forest regeneration and carbon: a rapid evidence
assessment. Canterbury, UK, University of Kent, Durrell Institute of
Conservation and Ecology.
Garnett,
S.T. & Lindenmayer, D.B. 2011. Conservation science must engender
hope to succeed. Trends in Ecology and Evolution, 26(2): 59–60.
Garnett, S.T., Burgess, N.D., Fa, J.E.,
Fernández-Llamazares, Á., Molnár, Z., Robinson, C.J., Watson, J.E.
et al. 2018. A spatial overview of the global importance of indigenous
lands for conservation. Nature Sustainability, 1(7): 369–374.
Gayi, S. & Tsowou, K. 2016. Cocoa industry: Integrating
small farmers into the global value chain. Geneva, Switzerland, UNCTAD.
[also available at
https://unctad.org/en/PublicationsLibrary/suc2015d4_en.pdf].
Gentry, A.H. & Dodson, C.H. 1987. Contribution of nontrees to
species richness of a tropical rain forest. Biotropica, 19:149– 56, cited
by Dirzo, R. & Raven, P. H. 2003. Global state of biodiversity and loss.
Annual Review of Environment and Resources, 28: 137–167.
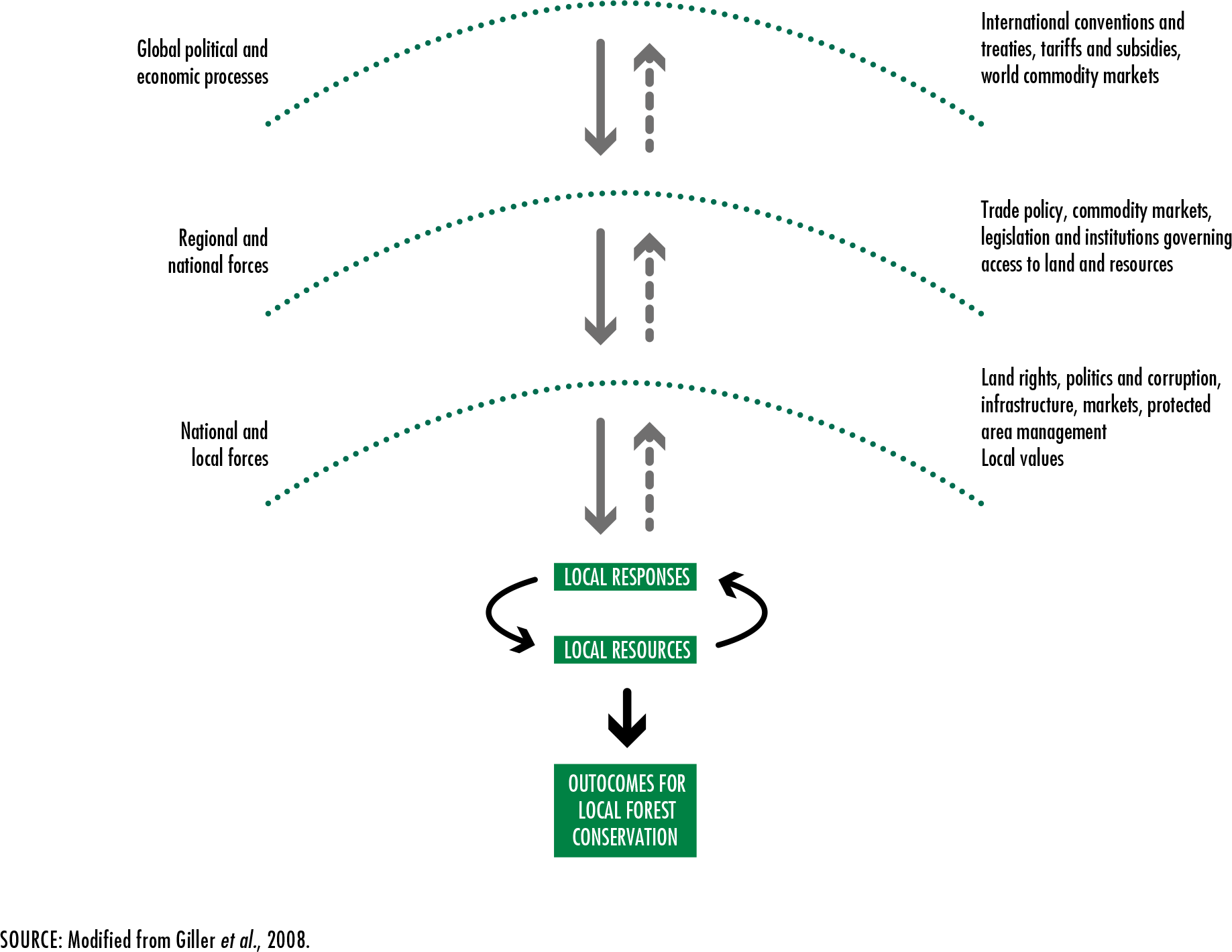
Giller, K.E., Leeuwis, C., Andersson, J.A., Andriesse, W., Brouwer, A.,
Frost, P., Hebinck, P.
et al. 2008. Competing claims on natural resources: what role for science?
Ecology and Society, 13(2): 34 [online]. [Cited 4 January 2020].
http://www.ecologyandsociety.org/vol13/iss2/art34/
Global Trees Campaign. 2020. Red lists. In: Global Trees
Campaign [online]. [Cited 4 January 2020].
https://globaltrees.org/threatened-trees/red-list/
Golden, C.D., Fernald, L.C.H., Brashares, J.S., Rasolofoniaina, B.J.R. &
Kremen, C.
2011. Benefits of wildlife consumption to child nutrition in a biodiversity hotspot.
PNAS, 108: 19653–19656.
González-Oreja, J.A., Bonache-Regidor, C. & de la Fuente-Díaz-Ordaz,
A.A.
2010. Far from the noisy world? Modelling the relationships between park size, tree
cover and noise levels in urban green spaces of the city of Puebla, Mexico.
Interciencia, 35(7): 486–492.
Gosnell, H.
& Abrams, J. 2011. Amenity migration: diverse conceptualizations of
drivers, socioeconomic dimensions, and emerging challenges. GeoJournal, 76,
303–322.
Government of Bhutan. 1997. Biodiversity Action Plan for
Bhutan. Thimpu, Bhutan. [also available at www.cbd.int/doc/world/bt/bt-nr-01-en.pdf].
Government of the United States of America.
1973. Endangered Species Act of 1973. Washington, DC. [also available at https://www.fws.gov/international/pdf/esa.pdf].
GPFLR. n.d. What is forest and landscape restoration (FLR)? In:
Global Partnership on Forest and Landscape Restoration
[online]. [Cited 4 January 2020].
www.forestlandscaperestoration.org/what-forest-and-landscape-restoration-flr
Great Green Wall. 2019a. The great green wall. In: Great Green
Wall [online]. Bonn, Germany. [Cited 31 December 2019].
https://www.greatgreenwall.org/about-great-green-wall
Great Green Wall. 2019b. Results. In: Great Green Wall
[online]. Bonn, Germany. [Cited 4 January 2020].
www.greatgreenwall.org/results
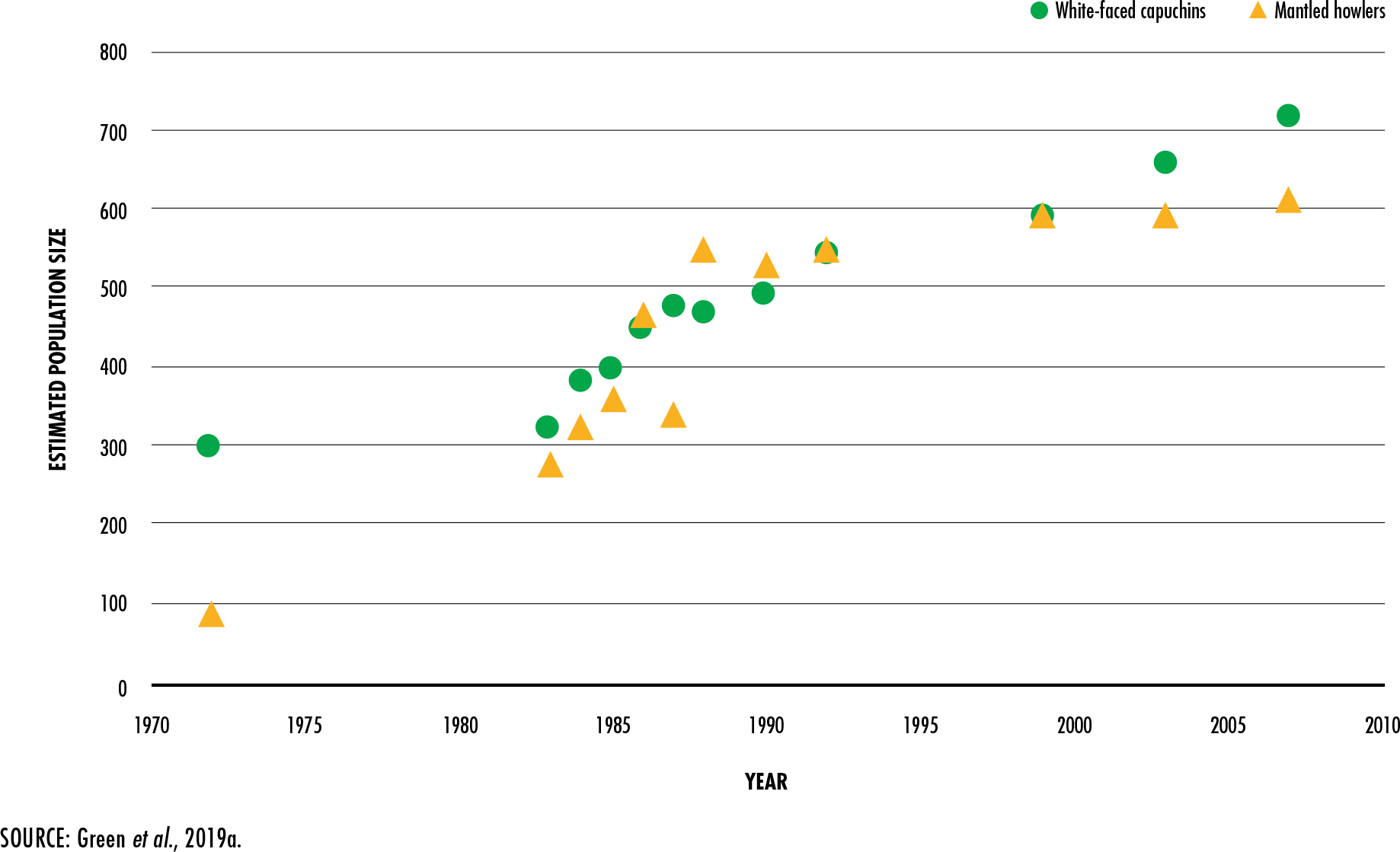
Green, E., McRae, L., Harfoot, M., Hill, S., Simonson, W. &
Baldwin-Cantello, W.
2019a.
Below the canopy: plotting global trends in forest wildlife populations.
Woking, UK, WWF-UK.
Green, E., McRae, L., Harfoot, M., Hill, S., & Baldwin-Cantello, W.,
Simonson, W.
2019b. Below the canopy: global trends in forest vertebrate populations and their
drivers. PeerJ Preprints, 7: e27882v1 [online]. [Cited 4 January 2020].
https://doi.org/10.7287/peerj.preprints.27882v1
Gretzinger, S. 2016. Latin American experiences in natural
forest management concessions. Forestry Policy and Institutions Working
Paper 35. Rome, FAO. [also available at
http://www.fao.org/forestry/45023-0707f17f1cce86c7e4f4e870bf4edd2f0.pdf].
Groenewegen, P.P., Van den Berg, A.E., De Vries, S. & Verheij,
R.A.
2006. Vitamin G: effects of green space on health, well-being, and social safety.
BMC public health, 6(1), 149 [online]. [Cited 4 January 2020].
https://doi.org/10.1186/1471-2458-6-149
Grogan, J., Free, C., Pinelo, G., Johnson, A. & Alegria, R.
2016.
Conservation status of five timber species populations in the forestry
concessions of the Maya Biosphere Reserve, Guatemala. Turrialba, Costa
Rica, CATIE.
Guariguata, M., Cronkleton, P., Duchelle, A. & Zuidema, P.
2017. Revisiting the ‘cornerstone of Amazonian conservation’: a socioecological
assessment of Brazil nut exploitation. Biodiversity and Conservation, 26:
2007–2027.
Gurnell, A.M., England, J., Shuker, L. & Wharton, G.
2019. The contribution of citizen science volunteers to river monitoring and
management: International and national perspectives and the example of the MoRPh
survey.
River Research and Applications, 35(8): 1359–1373.
Gurung,
J.D. 2002. Getting at the heart of the issue: Challenging male bias in
Nepal’s Department of Forests. Mountain Research and Development, 22(3):
212–216.
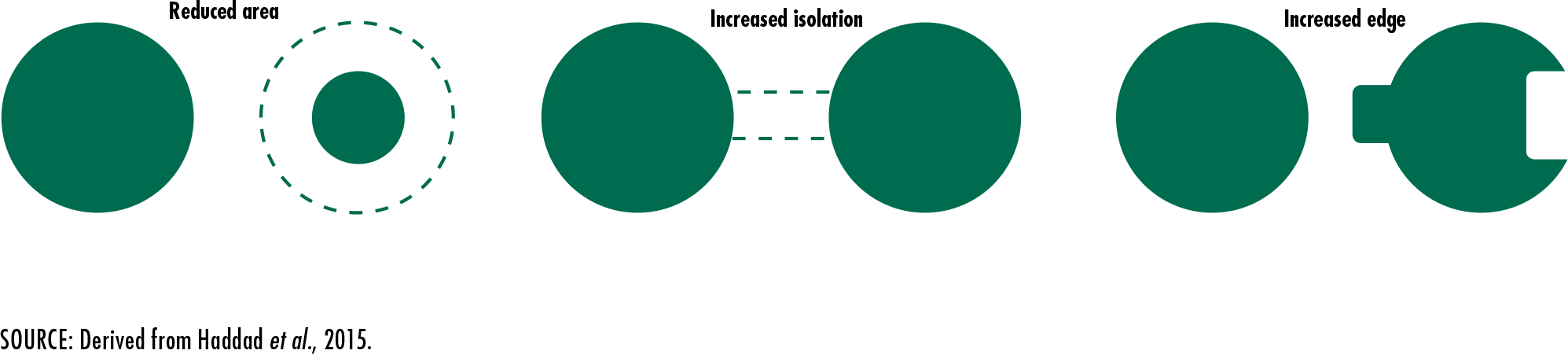
Haddad, N.M., Brudvig, L.A., Clobert, J., Davies, K.F., Gonzalez, A., Holt,
R.D., Lovejoy, T.E.
et al. 2015. Habitat fragmentation and its lasting impact on Earth’s
ecosystems. Science Advances, 1: e1500052 [online]. [Cited 4 January 2020].
DOI: 10.1126/sciadv.1500052
Hansen, M.M., Jones, R., & Tocchini, K.
2017. Shinrin-yoku (forest bathing) and nature therapy: A state-of-the-art review.
International Journal of Environmental Research and Public Health, 14(8):
851.
Hansen, M.C., Potapov, P.V., Moore, R., Hancher, M., Turubanova, S.A.,
Tyukavina, A., Thau, D.
et al. 2013. High-resolution global maps of 21st-century forest cover
change. Science, 342(6160): 850–853.
Hanski, I., von Hertzen, L., Fyhrquist, N., Koskinen, K., Torppa, K.,
Laatikainen, T., Karisola, P.
et al. 2012. Environmental biodiversity, human microbiota, and allergy are
interrelated. PNAS, 109(21): 8334–8339.
Hart,
D. 2018. Man the hunted: primates, predators, and human
evolution. New York, USA, Routledge.
Hartig, T., Mang, M., & Evans, G.W.
1991. Restorative effects of natural environment experiences. Environment and
Behavior, 23(1): 3–26.
Health Council of the Netherlands. 2004. Nature and Health. The
influence of nature on social, psychological and physical well-being. The
Hague, Health Council of the Netherlands and the Advisory Council for Research on
Spatial Planning, Nature and the
Environment in the Netherlands.
Hegetschweiler, K.T., Plum, C., Fischer, C.,
Brändli, U.B., Ginzler, C. & Hunziker, M. 2017. Towards a
comprehensive social and natural scientific forest-recreation monitoring
instrument – A prototypical approach. Landscape and Urban Planning, 167:
84–97.
Henders, S., Persson, U.M. & Kastner, T.
2015. Trading forests: land-use change and carbon emissions embodied in production
and exports of forest-risk commodities. Environmental Research Letters 10,
no. 12, doi:10.1088/1748-9326/10/12/125012.
Henriksen, L. 2018. Blue Targeting – manual. How to do Blue
Targeting for best management practice (BMP) for forestry along small
streams. Swedish Forest Agency, EU Interreg project Water Management in
Baltic Forests. [also available at
https://www.skogsstyrelsen.se/globalassets/projektwebbplatser/wambaf/blue-targeting/blue-targeting-manual.pdf].
Hermosilla, T., Wulder, M.A., White, J.C., Coops, N.C., Pickell, P.D. &
Bolton, D.K.
2019. Impact of time on interpretations of forest fragmentation: three-decades of
fragmentation dynamics over Canada. Remote Sensing of Environment, 222:
65–77.
Heß, S., Jaimovich, D., & Schündeln, M.
2019.
Environmental effects of development programs: Experimental evidence from West
African dryland forests
[online]. [Cited 13 January 2020].
http://hesss.org/Gambia%20Forest.pdf
Hilderbrand, G.V., Schwartz, C.C., Robbins, C.T., Jacoby, M.E., Hanley,
T.A., Arthur, S.M. & Servheen, C.
1999. The importance of meat, particularly salmon, to body size, population
productivity, and conservation of North American brown bears. Canadian Journal
of Zoology, 77: 132–138.
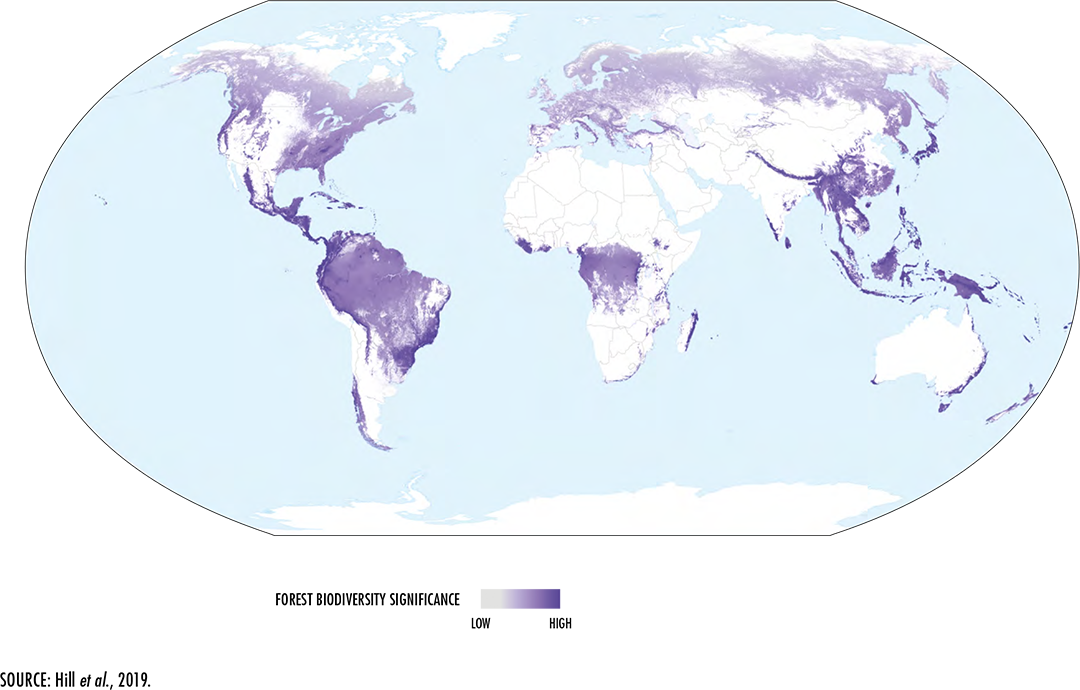
Hill, S.L.L., Arnell, A., Maney, C., Butchart, S.H.M., Hilton-Taylor, C.,
Ciciarelli, C., Davis, C., Dinerstein, E., Purvis, A. & Burgess,
N.D.
2019. Measuring forest biodiversity status and changes globally. Frontiers in
Forest and Global Change, 2: 70 [online]. [Cited 4 January 2020].
https://doi.org/10.3389/ffgc.2019.00070
Hlásny, T., Krokene, P., Liebhold, A., Montagné-Huck, C., Müller, J., Qin,
H., Raffa, K.
et al. 2019. Living with bark beetles: impacts, outlook and management
options. From Science to Policy 8. Barcelona, Spain, European Forest
Institute.
HLPE. 2017. Sustainable forestry for food security and
nutrition. A report by the High Level Panel of Experts on Food Security
and Nutrition of the Committee on World Food Security. Rome. [also
available at
www.fao.org/3/a-i7395e.pdf].
Hoare, A. 2015.
Tackling illegal logging and the related trade: what progress and where
next?
Chatham House Report. London, Chatham House, The Royal Institute of International
Affairs.
Hodgdon, B.D., Hughell, D., Ramos, V.H. & McNab, R.B.
2015.
Deforestation trends in the Maya Biosphere Reserve, Guatemala 2000–2013.
New York, USA, Rainforest Alliance.
Hoffmann, B., Roeger, S., Wise, P., Dermer, J., Yunupingu, B., Lacey, D.,
Yunupingu, D., Marika, B., Marika, M. & Panton, B.
2012. Achieving highly successful multiple agency collaborations in a cross-cultural
environment: experiences and lessons from Dhimurru Aboriginal Corporation and
partners. Ecological Management and Restoration, 13(1): 42–50.
Hosonuma, N., Herold, M., De Sy, V., De Fries, R.S., Brockhaus, M., Verchot,
L., Angelsen, A. & Romijn, E.
2012. An assessment of deforestation and forest degradation drivers in developing
countries.
Environmental Research Letters, 7(4): 044009 [online]. [Cited 4 January
2020].
https://doi.org/10.1088/1748-9326/7/4/044009
Hudson, L.N., Newbold, T., Contu, S., Hill, S.L., Lysenko, I., De Palma, A.,
Phillips, H.R.
et al. 2017. The database of the PREDICTS project. Ecology and
Evolution, 7(1): 145–188.
Hughes, T.W. & Lee, K. 2015. The role of recreational hunting
in the recovery and conservation of the wild turkey (Meleagris gallopavo
spp.) in North America. International Journal of Environmental Studies,
72(5): 797–809.
Huntley,
B.J. & Redford, K.H. 2014. Mainstreaming biodiversity in
practice: a STAP advisory document. Washington, DC, USA, GEF.
Ickowitz, A., Powell, B., Salim, M.A. & Sunderland, T.
2014. Dietary quality and tree cover in Africa.
Global Environmental Change, 24: 287–294.
IDH. 2019. Green Cocoa Cameroon. In: IDH, The Sustainable Trade
Initiative [online]. Utrecht, The Netherlands. [Cited 4 January 2020].
https://www.idhsustainabletrade.com/contact-directions/
IFAD &
UNEP. 2013. Smallholders, food security, and the environment.
Rome, IFAD.
IIED. 2019.
Darwin Initiative Main and Post Project Annual Report: Livelihoods Insurance
from Elephants (LIFE) in Kenya and Sri Lanka
[online]. London. [Cited 4 January 2020].
https://pubs.iied.org/pdfs/G04412.pdf
ILO.
2017. NORMLEX – C169 – Indigenous and Tribal Peoples Convention, 1989 (No. 169). In:
International Labour Organization [online]. Geneva, Switzerland. [Cited 2
January 2020].
https://www.ilo.org/dyn/normlex/en/f?p=NORMLEXPUB:12100:0::NO::P12100_ILO_CODE:C169
INAB. 2019. Cobertura forestal. In:
SIFGUA – Sistema de Información Forestal de Guatemala
[online]. Guatemala. [Cited 4 January 2020].
http://www.sifgua.org.gt/Cobertura.aspx
Ingwall-King, L. & Gangur, A.
forthcoming. Integrating traditional knowledge into conservation policy and
practice: a good practice review. Cambridge, UK, UNEP-WCMC.
Initiative 20x20. n.d. Healthy lands for food, water and
nature [online]. Washington, DC. [Cited 18 December 2019].
https://initiative20x20.org/
Institute of Medicine. 2001.
Dietary reference intakes for vitamin A, vitamin K, arsenic, boron, chromium,
copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and
zinc.
Washington, DC, National Academies Press.
Instituto Socioambiental. 2015. Advances and setbacks in
territorial rights in Brazil. Brasilia. Cited in RRI. 2015. Protected
areas and the land rights of indigenous peoples and local communities: current
issues and future agenda. Washington,
DC, RRI.
IPBES. 2016.
The assessment report on pollinators, pollination and food production-policy
platform on biodiversity and ecosystem services on pollinators, pollination and
food production. Bonn, Germany.
IPBES. 2019a. Summary for policymakers of the global assessment
report on biodiversity and ecosystem services of the Intergovernmental
Science-Policy Platform on Biodiversity and Ecosystem Services. Bonn,
Germany.
IPBES. 2019b.
Chapter 2.2 Status and Trends – Nature. Unedited draft chapter for IPBES Global
Assessment on Biodiversity and Ecosystem Services
[online]. Bonn, Germany. [Cited 13 January 2020].
https://ipbes.net/sites/default/files/ipbes_global_assessment_chapter_2_2_nature_unedited_31may.pdf].
IPCC. 2019.
Climate Change and Land: an IPCC special report on climate change,
desertification, land degradation, sustainable land management, food security,
and greenhouse gas fluxes in terrestrial ecosystems
[P.R. Shukla, J. Skea, E. Calvo Buendia, V. Masson-Delmotte, H.-O. Pörtner, D. C.
Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S.
Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K.
Kissick, M. Belkacemi, J. Malley
(eds.)]. [also available at:
https://www.ipcc.ch/srccl/]
Irvine, K.N., Devine-Wright, P., Payne, S.R., Fuller, R.A., Painter, B.
& Gaston, K.J.
2009. Green space, soundscape and urban sustainability: an interdisciplinary,
empirical study.
Local Environment, 14(2): 155–172.
Isted, A. 2013. An investigation into the benefits of forest
school intervention for young people with ADHD in the education system
(Examination paper). London, University of Greenwich.
ITC. 2016. Sustainable sourcing: Markets for certified Chinese
medicinal and aromatic plants. Geneva, Switzerland.
ITTO. 2002. ITTO guidelines for the restoration, management and
rehabilitation of degraded and secondary tropical forests. ITTO, CIFOR,
FAO, IUCN and WWF International.
ITTO & IUCN. 2009.
ITTO/IUCN Guidelines for the conservation and sustainable use of biodiversity in
tropical timber production forests.
ITTO Policy Development Series No. 17. Yokohama, Japan, ITTO.
IUCN. 2013. Mitigating human-wildlife conflict. In: IUCN
[online]. Gland, Switzerland. [Cited 4 January 2020].
https://www.iucn.org/content/mitigating-human-wildlife-conflict
IUCN. 2016. A global standard for the identification of key
biodiversity areas. Version 1.0. First edition. Gland, Switzerland.
IUCN. 2017. The IUCN red list of threatened species.
Version 2017.3. http://www.iucnredlist.org.
IUCN. 2018. The Bonn Challenge barometer. In: InfoFLR
[online]. Gland, Switzerland. [Cited 13 January 2020].
https://infoflr.org/bonn-challenge-barometer
IUCN. 2019a. The IUCN red list of threatened species.
Version 2019-2. http://www.iucnredlist.org.
Downloaded on 4 October 2019.
IUCN. 2019b. Over half of Europe’s endemic trees face extinction.
In: IUCN [online]. Gland, Switzerland. [Cited 4 January 2020].
https://www.iucn.org/news/species/201909/over-half-europes-endemic-trees-face-extinction
IUCN
WCPA. 2018. PARKS. The International Journal of Protected Areas and
Conservation. Volume 24 Special Issue. Gland, Switzerland, IUCN.
Jalonen, R., Valette, M., Boshier, D., Duminil, J. & Thomas, E.
2017. Forest and landscape restoration severely constrained by a lack of attention
to the quantity and quality of tree seed: Insights from a global survey.
Conservation Letters, 11(4): e12424 [online]. [Cited 4 January 2020].
https://doi.org/10.1111/conl.12424
Jamnadass, R., McMullin, S., Iiyama, M., Dawson, I.K., Powell, B., Termote,
C., Ickowitz, A.
et al. 2015. Understanding the roles of forests and tree-based systems in
food provision. In B. Vira, C. Wildburger & S. Mansourian, eds.
Forests and food: Addressing hunger and nutrition across sustainable
landscapes. Cambridge, UK, Open Book
Publishers.
http://dx.doi.org/10.11647/OBP.0085
Jayachandran S., de Laat, J., Lambin, E.F., Stanton, C.Y., Audy, R. &
Thomas, N.E.
2017. Cash for carbon: A randomized trial of payments for ecosystem services to
reduce deforestation.
Science, 357(6348): 267–273.
Jenkins, M., Timoshyna, A. & Cornthwaite, M.
2018.
Wild at home: exploring the global harvest, trade and use of wild plant
ingredients.
Cambridge, UK, TRAFFIC International.
Jonas, H.D., MacKinnon K., Dudley N., Hockings M., Jessen S., Laffoley D.,
MacKinnon D.
et al. 2018. Editorial essay: Other effective area-based conservation
measures: From Aichi Target 11 to the Post-2020 biodiversity framework. PARKS,
The International Journal of Protected Areas and Conservation, 24 (Special
issue on OECMs): 9–16.
Jorgensen, A., Hitchmough, J. & Dunnet, N.
2006. Woodland as a setting for housing-appreciation and fear and the contribution
of residential satisfaction and place identity in Warrington New Town, UK.
Landscape and Urban Planning, 79(3–4): 273–287.
Kaimowitz,
D., & Sheil, D. 2007. Conserving what and for whom? Why
conservation should help meet basic human needs in the tropics. Biotropica,
39(5): 567–574.
Kaplan, R.
& Kaplan, S. 1989. The experience of nature – a psychological
perspective. Cambridge, UK, Cambridge University Press.
Kapos, V., Lysenko, I. & Lesslie, R.
2002.
Assessing forest integrity and naturalness in relation to biodiversity. FAO
Forest Resources Assessment Programme Working Paper 54. Rome. [also available at http://www.fao.org/3/ad654e/ad654e00.htm].
Kareiva, P., Watts, S., McDonald, R. & Boucher, T.
2007. Domesticated nature: Shaping landscapes and ecosystems for human welfare.
Science, 316(5833): 1866–1869.
Katila, P., Pierce Colfer, C., De Jong, W., Galloway, G., Pacheco, P., &
Winkel, G., eds.
2019.
Sustainable Development Goals: their impacts on forests and people.
Cambridge, UK, Cambridge University Press.
Kawarazuka, N. & Béné, C. 2011. The
potential role of small fish species in improving micronutrient deficiencies in
developing countries: building evidence.
Public Health Nutrition, 14(11): 1927–1938.
Kay, C.E. 2018. The Condition and Trend of Aspen, Willows, and
Associated Species on the Northern Yellowstone Range. Rangelands, 40(6):
202–211. [also available at
https://www.sciencedirect.com/science/article/pii/S0190052818300774?via%3Dihub]
Keenan, R.J., Reams, G.A., Achard, F., de Freitas, J.V., Grainger, A. &
Lindquist, E.
2015. Dynamics of global forest area: Results from the FAO Global Forest Resources
Assessment 2015.
Forest Ecology and Management, 352: 9–20.
Kelleher, C.T., de Vries, S.M.G., Baliuckas, V., Bozzano, M., Frydl, J.,
Gonzalez Goicoechea, P., Ivankovic, M.
et al. 2015. Approaches to the conservation of forest genetic resources
in Europe in the context of climate change. European Forest Genetic
Resources Programme (EUFORGEN). Rome, Bioversity International.
Kerr, J., Pender, J. & Suyanto, B.L.
2008. Property rights, environmental services and poverty alleviation in Indonesia.
BASIS Brief 2008-03. Madison, WI, University of Wisconsin.
King, L., Lala, F., Nzumu, H., Mwambingu, E. & Douglas-Hamilton,
I.
2017. Beehive fences as a multidimensional conflict-mitigation tool for farmers
coexisting with elephants. Conservation Biology, 31(4): 743–752.
Klein, A.M., Vaissiere, B.E., Cane, J.H., Steffan-Dewenter, I., Cunningham,
S.A., Kremen, C. & Tscharntke, T.
2007. Importance of pollinators in changing landscapes for world crops.
Proceedings of the Royal Society B – Biological Sciences, 274: 303–313.
Koskela, J., Lefèvre, F., Schueler, S., Kraigher, H., Olrik, D.C., Hubert,
J., Longauer, R.
et al. 2013. Translating conservation genetics into management:
Pan-European minimum requirements for dynamic conservation units of forest tree
genetic diversity. Biological Conservation, 157: 39–49.
Koskela, J., Vinceti, B., Dvorak, W., Bush, D., Dawson, I., Loo, J., Kjær,
E.D.
et al. 2014. Use and transfer of forest genetic resources: A global review.
Forest Ecology and Management, 333: 22–34.
Krishnan, S., Wiederkehr Guerra, G., Bertrand, D., Wertz-Kanounnikoff, S.
& Kettle, C.
forthcoming.
Enhancing the cross-sectoral benefits from forests for pollination services at
landscape scales: a review of management interventions. [tentative title].
FAO working paper. Rome, FAO and Bioversity International.
Lambin,
E.F., & Meyfroidt, P. 2011. Global land use change, economic
globalization, and the looming land scarcity. PNAS, 108(9): 3465–3472.
Lambin, E.F., Turner, B.L., Geist, H.J., Agbola, S.B., Angelsen, A., Bruce,
J.W., Coomes, O.T.
et al. 2001. The causes of land-use and land-cover change: moving beyond
the myths. Global Environmental Change, 11(4): 261–269.
Laurance, W.F., Nascimento, H.E.M., Laurance, S.G., Andrade, A.C.,
Fearnside, P.M., Ribeiro, J.E.L. & Capretz, R.L.
2006. Rain forest fragmentation and the proliferation of successional trees.
Ecology, 87(2): 469–482.
Le Bel, S., Mapuivre, G. & Czudek, R.
2010. Human–wildlife conflict toolkit: comprehensive solutions for farmers and
communities. Unasylva, 236: 12–13.
Lefèvre, F., Koskela, J., Hubert, J., Kraigher, H., Longauer, R., Olrik,
D.C., Schüler, S.
et al. 2013. Dynamic conservation of forest genetic resources in 33
European countries. Conservation Biology, 27(2): 373–384.
Lele, S., Wilshusen, P., Brockington, D., Seidler, R. & Bawa,
K.
2010. Beyond exclusion: alternative approaches to biodiversity conservation in the
developing tropics.
Current Opinion in Environmental Sustainability, 2(1): 94–100.
Leticia Pact. 2019. Leticia Pact for the Amazon. [Cited 2
January 2020].
https://id.presidencia.gov.co/Documents/190906-Pacto-Leticia-Amazonia-Ingles.pdf
Leverington, F., Lemos Costa, K., Pavese, H., Lisle, A. & Hockings,
M.
2010. A global analysis of protected area management effectiveness.
Environmental Management, 46(5): 685–698.
Levis, C., Costa, F.R., Bongers, F., Peña-Claros, M., Clement, C.R.,
Junqueira, A.B., Neves, E.G.
et al. 2017. Persistent effects of pre-Columbian plant domestication on
Amazonian forest composition. Science, 355(6328): 925–931.
Lham, D., Wangchuk, S., Stolton, S. & Dudley, N.
2019. Assessing the effectiveness of a protected area network: a case study of
Bhutan. Oryx, 53(1): 63–70.
Li, Q., Morimoto, K., Kobayashi, M., Inagaki, H., Katsumata, M., Hirata, Y.,
Hirata, K.
et al. 2008. Visiting a forest, but not a city, increases human natural
killer activity and expression of anti-cancer proteins. International Journal of
Immunopathology and Pharmacology, 21: 117–128.
Lindenmayer,
D.B. & Fischer, J. 2006. Habitat fragmentation and landscape
change: An ecological and conservation synthesis. Washington, DC, Island
Press.
Linnell, J.D. & Alleau, J. 2016. Predators that kill humans:
myth, reality, context and the politics of wolf attacks on people. In F.M.
Angelici, ed. Problematic wildlife: A cross-disciplinary approach, pp.
357–371. Cham, Switzerland,
Springer.
Liu, X., Li, Y., Guasch-Ferré, M., Willett, W.C., Drouin-Chartier, J.-P.,
Bhupathiraju, S.N. & Tobias, D.K.
2019. Changes in nut consumption influence long-term weight change in US men and
women.
BMJ Nutrition, Prevention & Health, 2(2) [online] [Cited 13 January
2020].
http://dx.doi.org/10.1136/bmjnph-2019-000034
Lo, M., Narulita, S. & Ickowitz, A.
2019. The relationship between forests and freshwater fish consumption in rural
Nigeria. PLOS ONE, 14(6): e0218038 [online]. [Cited 4 January 2020].
https://doi.org/10.1371/journal.pone.0218038
Lobón-Cerviá, J., Hess, L.L., Melack, J.M. & Araujo-Lima, C.A.
2015. The importance of forest cover for fish richness and abundance on the Amazon
floodplain. Hydrobiologia, 750(1): 245–255.
Lompo, D., Vinceti, B., Gaisberger, H., Konrad,
H., Duminil, J., Quedraogo, M., Sina, S. & Geburek, T.
2017. Genetic conservation in
Parkia biglobosa (Fabaceae: Mimosoideae) – what do we know? Silvae
Genetica, 66(1): 1–8.
Lompo, D., Vinceti, B., Konrad, H., Gaisberger, H. & Geburek,
T.
2018. Phylogeography of African locust bean (Parkia biglobosa) reveals
genetic divergence and spatially structured populations in West and Central Africa.
Journal of Heredity, 109(7): 811–824.
Luke (Natural Resources Institute Finland).
2018. 5+1 Steps towards a functioning insect economy. In:
Luke, Natural Resources Institute Finland [online]. Helsinki. [Cited 4
January 2020].
https://www.luke.fi/en/51-steps-towards-functioning-insect-economy/
Lung, T. & Schaab, G. 2010. A comparative assessment of land
cover dynamics of three protected forest areas in tropical eastern Africa.
Environmental Monitoring and Assessment, 161(1): 531–548.
Lupala, Z.J., Lusambo, L.P., Ngaga, Y.M. & Makatta, A.A.
2015. The land use and cover change in Miombo woodlands under community based forest
management and its implication to climate change mitigation: a case of southern
highlands of Tanzania.
International Journal of Forestry Research, Volume 2015: Article ID 459102
[online]. [Cited 4 January 4, 2020].
http://dx.doi.org/10.1155/2015/459102
Maas, J., Verheij, R.A., Groenewegen, P.P., de Vries, S. &
Spreeuwenberg, P.
2006. Green space, urbanity, and health: how strong is the relation?
Journal of Epidemiology and Community Health, 60(7): 587–592.
Mace,
G.M. 2014. Whose conservation? Science, 345(6204): 1558–1560.
Mahoney,
S.P. & Geist, V., eds. 2019. The North American model of
wildlife conservation. Baltimore, MD, USA, Johns Hopkins University Press.
Maisels, F., Strindberg, S., Blake, S., Wittemyer, G., Hart, J., Williamson,
E.A., Aba’a, R.
et al. 2013. Devastating decline of forest elephants in central Africa.
PLOS ONE, 8(3): e59469 [online]. [Cited 4 January 2020].
https://doi.org/10.1371/journal.pone.0059469
Maxwell, S.L., Fuller, R.A., Brooks, T.M. & Watson, J.E.M.
2016. The ravages of guns, nets and bulldozers.
Nature, 536(7615): 143–145.
May,
R. 2010. Tropical arthropod species, more or less? Science,
329(5987): 41–42.
Mbora A., Jamnadass R. & Lillesø J.-P.B.
2008.
Growing high priority fruits and nuts in Kenya: Uses and management.
Nairobi, The World Agroforestry Centre.
McDonell, E. 2019. Creating the culinary frontier. A critical
examination of Peruvian chefs’ narratives of lost/discovered foods. Anthropology
of Food, 14 [online]. [Cited 4 January 2020].
http://journals.openedition.org/aof/10183
McFarlane, R.A., Barry, J., Cissé, G., Gislason, M., Gruca, M., Higgs, K.,
Horwitz, P.
et al. 2019. SDG 3: Good health and well-being – framing targets to
maximise co-benefits for forests and people. In P. Katila, C.J. Pierce
Colfer, W. de Jong, G. Gallowa, P. Pacheco & G. Winkel, eds.
Sustainable Development Goals: their impacts on forests and people, pp.
72–107. Cambridge, UK, Cambridge University Press.
McKeown, R. 2002. Education for sustainable development
toolkit. Version 2. [Cited 4 January 2020].
http://esdtoolkit.org/esd_toolkit_v2.pdf
McShane, T.O., Hirsch, P.D., Trung, T.C., Songorwa, A.N., Kinzig, A.,
Monteferri, B., Mutekanga, D.
et al. 2011. Hard choices: Making trade-offs between biodiversity
conservation and human well-being. Biological Conservation, 144: 966–972.
MEA. 2005. Ecosystems and human well-being: current state and
trends. Washington, DC, Island Press.
Medaglia, J.C., Phillips, L.-K. & Perron-Welch, F.
2014.
Biodiversity legislation study: a review of biodiversity legislation in 8
countries. London, The Global Legislators’ Organisation, Hamburg, Germany,
the World Future Council, and Montreal, Canada, the Centre for International
Sustainable Development Law. [also available
at http://www.cisdl.org/wp-content/uploads/2018/04/Biodiversity-Legislation-Study.pdf]
MEF (Ministry of Environment and Forestry).
2018. The state of Indonesia’s forests 2018. Jakarta.
MERECP. 2007. Mount Elgon Regional Ecosystem Conservation
Programme (MERECP), Work Plan (version March 2007). Nairobi, Kenya.
Min, Q. 2017. Learning from the past for the future: experiences of
Hani Rice Terraces in coping with extreme drought. Presentation at a side event on
Globally Important Agricultural Heritage Systems and Climate Change, 23rd session of
the Conference of the
Parties to UNFCCC, Bonn, Germany, 10 November.
MINEF. 1998. Décision No. 0108/D/MINEF/CAB du 9 février 1998:
“Portant application des normes d’intervention en milieu forestier en République du
Cameroun.” Chapitre VI, Articles 28, 29 et 30 – “Protection de la faune.” Yaoundé.
MINEF. 2001. Order No. 0222/A/MINEF of May 25, 2002 on “procedures
for developing, approval, monitoring and control of the implementation of forest
management plans for the production forests in the permanent forest estate.” Article
11(1) and (3). Yaoundé.
MINEPDED. 2013. “Readiness Preparation Proposal (R-PP) submitted to
the World Bank’s Forest Carbon Partnership Facility (FCPF)” (unpublished).
MIPAAF. 2017. Comunicati stampa – Creato primo elenco alberi
monumentali d’Italia [Press release – First list of monumental trees of Italy
created]. In:
Ministero delle politiche agricole alimentari e forestali
[online]. Rome. [Cited 4 January 2020].https://www.politicheagricole.it/flex/cm/pages/ServeBLOB.php/L/IT/IDPagina/12052
MIPAAF. 2019. Elenco degli alberi monumentali d’Italia ai sensi
della Legge n. 10/2013 e del Decreto 23 ottobre 2014 [List of the monumental trees
of Italy under Law No. 10/2013 and the Decree of 23 October 2014]. In:
Ministero delle politiche agricole alimentari e forestali
[online]. Rome. [Cited 4 January 2020].
www.politicheagricole.it/flex/cm/pages/ServeBLOB.php/L/IT/IDPagina/11260
Mitchell, R. & Popham, F. 2008.
Effect of exposure to natural environment on health inequalities: an observational
population study. Lancet, 372(9650): 1655–1660.
Mittermeier, R.A., Myers, N., Thomsen, J.B., da Fonseca, G.A.B. &
Olivieri, S.
1998. Biodiversity hotspots and major tropical wilderness areas: approaches to
setting conservation priorities.
Conservation Biology, 12(3): 516–520.
Mittermeier, R.A., Gil, P.R., Hoffman, M., Pilgrim, J., Brooks, T.,
Mittermeier, C.G., Lamoreux, J. & da Fonseca, G.A.B.
2004.
Hotspots revisited: Earth’s biologically richest and most endangered terrestrial
ecoregions. Colonia Centro, Monterrey, Mexico, Cemex.
Mittermeier, R.A., Turner, W.R., Larsen, F.W., Brooks, T.M. & Gascon,
C.
2011. Global biodiversity conservation: The critical role of hotspots. In
F.E. Zachos & J.C. Habel, eds. Biodiversity hotspots: Distribution and
protection of conservation priority areas, pp. 3–22. Berlin, Springer, cited by
IPBES. 2019b.
Chapter 2.2 Status and Trends – Nature. Unedited draft chapter for IPBES Global
Assessment on Biodiversity and Ecosystem Services
[online]. Bonn, Germany. [Cited 13 January 2020].
https://ipbes.net/sites/default/files/ipbes_global_assessment_chapter_2_2_nature_unedited_31may.pdf
MNRT. 2015. National Forest Resources Monitoring and Assessment
of Tanzania mainland (NAFORMA). Main results. Dar es Salaam, MNRT.
Molinario, G., Hansen, M., Potapov, P., Tyukavina, A. & Stehman,
S.
2020. Contextualizing Landscape-Scale Forest Cover Loss in the Democratic Republic
of Congo (DRC) between 2000 and 2015. Land 9(1), 23. [also available at https://doi.org/10.3390/land9010023]
Monbiot,
G. 2013. Feral: Rewilding the Land, Sea and Human Life.
Penguin.
Mongbo, R., Floquet, A., Choden, S. & Moreno Diaz, M.L.
2011.
Protected areas – Not just for biodiversity conservation. The contributions of
protected areas to the economic and social development in Bhutan, Costa Rica and
Benin. Costa Rica, Universidad Nacional.
MoP (Ministry of Planning and International Cooperation) & MoE (Ministry
of Environment Jordan).
2008.
Integrated financing strategy for sustainable land management in Jordan. Final
report. Amman. [also available at http://extwprlegs1.fao.org/docs/pdf/jor169877.pdf].
Mora, C., Tittensor, D.P., Adl, S., Simpson, A.G.B. & Worm, B.
2011. How many species are there on Earth and in the ocean?
PLOS Biology, 9(8): e1001127 [online]. [Cited 5 January 2020].
https://doi.org/10.1371/journal.pbio.1001127
Mulenga, B.P., Tembo, S.T. & Richardson, R.B.
2019. Electricity access and charcoal consumption among urban households in Zambia.
Development Southern Africa, 36(5): 585–599.
Myers,
N. 1990. The biodiversity challenge: Expanded hot-spots analysis.
Environmentalist, 10(4): 243–256.
Myers, N., Mittermeier, R.A., Mittermeier, C.G., da Fonseca, G.A.B. &
Kent, J.
2000. Biodiversity hotspots for conservation priorities.
Nature, 403: 853–858.
NACSO. 2017a. Human wildlife conflict – the hot potato. In:
Namibian Association of CBNRM Support Organizations
[online]. Windhoek. [Cited 5 March 2019].
http://www.nacso.org.na/news/2017/03/human-wildlife-conflict-%E2%80%93-the-hot-potato
NACSO. 2017b. Resources & publications: State of Community
Conservation figures and tables. In:
Namibian Association of CBNRM Support Organizations
[online]. [Cited 18 December 2019].
http://www.nacso.org.na/resources/state-of-community-conservation-figures-and-tables
Nadkarni,
N. 2004. Not preaching to the choir: Communicating the importance of
forest conservation to nontraditional audiences. Conservation Biology,
18(3): 602–606.
Nasi, R., Taber, A. & Van Vliet, N.
2011. Empty forests, empty stomachs? Bushmeat and livelihoods in the Congo and
Amazon Basins.
International Forestry Review, 13(3): 355–368.
Nasi, R., Brown, D., Wilkie, D., Bennett, E., Tutin, C., van Tol, G. &
Christophersen, T.
2008. Conservation and use of wildlife-based resources: the bushmeat crisis.
Technical Series No. 33. Montreal, Canada, Secretariat of the Convention on
Biodiversity, and Bogor, Indonesia, CIFOR.
Nature4Climate. 2019. Nature-based solutions: a summary of
announcements and developments during the UN Climate Action Summit and Climate Week.
In: Nature4Climate [online]. [Cited 5 January 2020].
https://nature4climate.org/news/nature-based-solutions-a-summary-of-announcements-and-developments-during-the-un-climate-action-summit-and-climate-week
NCED. 2019. What is a conservation easement? In:
NCED, National Conservation Easement Database
[online]. Greenville, SC, USA. [Cited 5 January 2020].
www.conservationeasement.us/what-is-a-conservation-easement
Nel, A. & Hill, D. 2013. Constructing walls of carbon–the
complexities of community, carbon sequestration and protected areas in Uganda.
Journal of Contemporary African Studies, 31(3): 421–440.
Nellemann, C., Henriksen, R., Kreilhuber, A., Stewart, D., Kotsovou, M.,
Raxter, P., Mrema, E. & Barrat, S., eds.
2016.
The rise of environmental crime: A growing threat to natural resources peace,
development and security. Nairobi, UNEP, and Oslo, Norwegian Center for
Global Analyses (RHIPTO).
Nelson F. & Sinandei, M. 2018. Building stronger grassroots
organizations that can take community land rights to scale. In: Land portal
[online]. Amersfoort, The Netherlands. [Cited 5 January 2020].
https://landportal.org/blog-post/2018/03/building-stronger-grassroots-organizations-can-take-community-land-rights-scale
New Generation Plantations. 2018.
Rainforest restoration in Brazil’s Atlantic Forest
[online]. [Cited 13 December 2019].
https://newgenerationplantations.exposure.co/rainforest-restoration-in-brazils-atlantic-forest
Newton, P., Miller, D.C., Byenkya, M.A.A. & Agrawal, A.
2016. Who are forest-dependent people? A taxonomy to aid livelihood and land use
decision-making in forested regions.
Land Use Policy, 57: 388–395.
Nguinguiri, J.C., Czudek, R., Larrubia, C.J.,
Ilama, L., Le Bel, S., Angoran, E.J., Trebuchon, J.F. & Cornelis,
D. 2017. Managing human–wildlife conflicts in central and southern
Africa. Unasylva, 249: 39–44.
Nielsen, M.R., Meilby, H., Smith-Hall, C., Pouliot, M. & Treue,
T.
2018. The importance of wild meat in the global south.
Ecological Economics, 146: 696–705.
Nilsson M., Griggs D. & Visbeck M.
2016. Policy: Map the interactions between Sustainable Development Goals.
Nature, 534: 320–322.
Nilsson, K., Sangster, M., Gallis, C., Hartig, T., De Vries, S., Seeland, K.
& Schipperijn, J., eds.
2010. Forests, trees and human health. New York, USA, Springer Science +
Business Media.
Nirmal, S.A., Pal, S.C., Otimenyin, S.O., Aye, T., Elachouri, M., Kundu,
S.K., Thandavarayan, R.A. & Mandal, S.C.
2013. Contribution of herbal products in the global market.
The Pharma Review, November–December 2013: 95–104.
Norgrove, L.
& Hulme, D. 2006. Confronting conservation at Mount Elgon, Uganda.
Development and Change, 37(5): 1093–1116.
Nowak, D.J., Crane, D.E. & Stevens, J.C.
2006. Air pollution removal by urban trees and shrubs in the United States.
Urban Forestry & Urban Greening, 4(3–4): 115–123.
Nwaokoro, N. & Kwon-Ndung, E. 2010. Exploiting the
potentials of Parkia biglobosa in Nigeria. Paper presented at
Plant Biology 2010, Joint Annual Meeting of the American Society of Plant Biologists
and the Canadian Society of Plant
Physiologists– La Société Canadienne de Physiologie Végétale, Montreal, Canada, 31
July–4 August 2010.
NYDF. 2019. Protecting and restoring forests: A story of large
commitments yet limited progress. New York Declaration on Forests Five-year
assessment report. Amsterdam, Climate Focus.
Nyhus,
P.J. 2016. Human–wildlife conflict and coexistence. Annual Review
of Environment and Resources, 41: 143–171.
O’Brien,
L. 2009. Learning outdoors: The Forest School approach. Education
3–13, 37(1): 45–60.
O’Brien, L.
& Murray, R. 2007. Forest school and its impacts on young children:
case studies in Britain. Urban Forestry & Urban Greening, 6(4):
249–265.
Ødegaard,
F. 2000. How many species of arthropods? Erwin’s estimate revised.
Biological Journal of the Linnean Society, 71(4): 583–597.
Odetokun, S.M. 1996. The nutritive value of baobab fruit
(Adansonia digitata). Rivista Italiana delle Sostanze Grasse, 73: 371–373,
cited by Manfredini, S., Vertuani, S. & Buzzoni, V. 2002. Adansonia
digitata. Il baobab farmacista.
L’integratore nutrizionale, 5: 25–29.
OECD. 2019a. Agricultural policy monitoring and evaluation
2019. Paris.
OECD. 2019b. Biodiversity: Finance and the economic and
business case for action. Paris.
Olival, K.J., Hosseini, P.R., Zambrana-Torrelio, C., Ross, N., Bogich, T.L.
& Daszak, P.
2017. Host and viral traits predict zoonotic spillover from mammals.
Nature, 546: 646–650.
Ollerton, J., Winfree, R. & Tarrant, S.
2011. How many flowering plants are pollinated by animals?
Oikos, 120(3): 321–326.
Olson, D.M., Dinerstein, E., Wikramanayake, E.D., Burgess, N.D., Powell,
G.V.N., Underwood, E.C., D’Amico, J.A.
et al. 2015. Terrestrial ecoregions of the world: A new map of life on
Earth. BioScience, 51(11): 933–938.
Onana, J.-M., Cheek, M. & Pollard, B.
2011.
Red Data Book of the Flowering Plants of Cameroon: IUCN global assessments.
Richmond, Surrey, UK, Kew Publishing.
Ong, S. & Carver, E. 2019. The rosewood trade: An illicit trail
from forest to furniture. In: YaleEnvironment360 [online]. New Haven, CT,
USA. [Cited 5 January 2020].
https://e360.yale.edu/features/the-rosewood-trade-the-illicit-trail-from-forest-to-furniture
Oregon Fish and Wildlife Office. n.d. Northern spotted owl. In:
U.S. Fish& Wildlife Service, Oregon Fish and Wildlife Office
[online]. Washington, DC. [Cited 5 January 2020].
www.fws.gov/oregonfwo/articles.cfm?id=149489595
Orgiazzi, A., Bardgett, R., Barrios, E., Behan Pelletier, V., Briones,
M.J.I., Chotte, J-L., De Deyn, G.
et al. eds. 2016. Global Soil Biodiversity Atlas. European
Commission, Publications Office of the European Union, Luxembourg.
Ostrom, E.
& Nagendra, H. 2006. Insights on linking forests, trees, and people
from the air, on the ground, and in the laboratory. PNAS, 103(51):
19224–19231.
Osuri, A.M., Ratnam, J., Varma, V., Alvarez-Loayza, P., Hurtado Astaiza, J.,
Bradford, M., Fletcher, C.
et al. 2016. Contrasting effects of defaunation on aboveground carbon
storage across the global tropics. Nature Communications, 7: 11351
[online]. [Cited 5 January 2020].
https://doi.org/10.1038/ncomms11351
Park, B.J., Tsunetsugu, Y., Kasetani, T., Kagawa, T. & Miyazaki,
Y.
2010. The physiological effects of
Shinrin-yoku (taking in the forest atmosphere or forest bathing): evidence
from field experiments in 24 forests across Japan. Environmental Health and
Preventive Medicine, 15: 18 [online]. [Cited 5 January 2020].
https://doi.org/10.1007/s12199-009-0086-9
Patenaude, G. & Lewis, K. 2014. The impacts of Tanzania’s
natural resource management programmes for ecosystem services and poverty
alleviation. International Forestry Review, 16(4): 459−473.
Paumgarten, F., Locatelli, B. & Witkowski, E.T.F.
2018. Wild foods: safety net or poverty trap? A South African case study. Human
Ecology, 46(2): 183–195.
Payn, T., Carnus, J.M., Freer-Smith, P., Kimberley, M., Kollert, W., Liu,
S., Orazio, C., Rodriguez, L., Neves Silva, L. & Wingfield, M.
2015. Changes in planted forests and future global implications. Forest Ecology
and Management, 352: 57–67.
Pereira, H.M., Leadley, P.W., Proenca, V.,
Alkemada, R., Scharlemann, J.P.W., Fernandez-Manjarres, J.F., Araujo.
M.B. et al. 2010. Scenarios for global biodiversity in the
21st
century. Science, 330(6010): 1496–1501.
Peres, C.A., Thaise, E., Schietti, J., Desmoulieres, S.J.M. & Levi,
T.
2016. Dispersal limitation induces long-term biomass collapse in overhunted
Amazonian forests. PNAS, 113: 892–897.
Persha, L., Agrawal, A. & Chhatre, A.
2011. Social and ecological synergy: Local rulemaking, forest livelihoods, and
biodiversity conservation.
Science, 331(6024): 1606–1608.
Peters, C.M. 2000. Pre-Columbian silviculture and indigenous
management of neotropical forests. In D.L. Lentz, ed. Imperfect
balance: landscape transformations in the Pre-Columbian Americas, pp.
203–223. New York, USA, Columbia University Press.
Phalan, B., Onial, M., Balmford, A. & Green, R.
2011. Reconciling food production and biodiversity conservation: land sharing and
land sparing compared.
Science, 333(6047): 1289–1291.
Plumptre, A.J., Kayitare, A., Rainer, H., Gray, M., Munanura, I.,
Barakabuye, N., Asuma, S., Sivha, M. & Namara, A.
2004.
The socio-economic status of people living near protected areas in the Central
Albertine Rift. Albertine Rift Technical Reports, 4. Kampala, Albertine
Rift Programme.
Polisar, J., de Thoisy, B., Rumiz, D., Dıaz Santos, F., Balas McNab, R.,
Garcia-Anleu, R., Ponce-Santizo, G., Arispe, R. & Venega, C.
2016. Using certified timber extraction to benefit jaguar and ecosystem
conservation. Ambio, 46: 588–603.
Porter-Bolland, L., Ellis, E.A., Guariguata, M.R., Ruiz-Mallén, I.,
Negrete-Yankelevich, S. & Reyes-García, V.
2012. Community managed forests and forest protected areas: An assessment of their
conservation effectiveness across the tropics. Forest Ecology and
Management, 268: 6–17.
Potapov, P., Hansen, M.C., Laestadius, L, Turubanova, S., Yaroshenko, A.,
Thies, C., Smith, W.
et al. 2017. The last frontiers of wilderness: Tracking loss of intact
forest landscapes from 2000 to 2013. Science Advances, 3(1): e1600821
[online]. [Cited 5 January 2020]. DOI: 10.1126/sciadv.1600821
Poudel, J.,
Zhang, D. & Simon, B. 2019. Habitat conservation banking trends in
the United States. Biodiversity and Conservation, 28(6): 1629–1646.
Poulsen, J.R., Clark, C.J. & Palmer, T.M.
2013. Ecological erosion of an Afrotropical forest and potential consequences for
tree recruitment and forest biomass. Biological Conservation, 163: 122–130.
Powell, B.,
Hall, J. & Johns, T. 2011. Forest cover, use and dietary intake in
the East Usambara Mountains, Tanzania. International Forestry Review,
13(3): 305–317.
Premauer J. & Berkes F. 2012. Makuira, Colombia: the
cosmological centre of origin for the Wayúu people. In N. Dudley & S.
Stolton, eds. Protected landscapes and wild biodiversity, p. 53–60. Gland,
Switzerland, IUCN.
Premauer, J.
& Berkes, F. 2015. A Pluralistic approach to protected area
governance: Indigenous peoples and Makuira National Park. Ethnobiology and
Conservation 4: 1–16.
Pretty, J.
& Smith, D. 2004. Social capital in biodiversity conservation and
management. Conservation Biology, 18(3): 631–638.
Price,
R. 2017. Economic drivers and effects of the illegal wildlife trade
in sub-Saharan Africa. K4D Helpdesk Report. Brighton, UK, IDS.
Rasolofoson, R.A., Hanauer, M.M., Pappinen, A., Fisher, B. & Ricketts,
T.H.
2018. Impacts of forests on children’s diet in rural areas across 27 developing
countries. Science Advances, 4(8): eaat2853 [online]. [Cited 5 January
2020]. DOI: 10.1126/sciadv.aat2853
Ratnam, W., Rajora, O.P., Finkeldey, R., Aravanopoulos, F., Bouvet, J.-M.,
Vaillancourt, R.E., Kanashiro, M., Fady, B., Tomita, M. & Vinson,
C.
2014. Genetic effects of forest management practices: Global synthesis and
perspectives.
Forest Ecology and Management, 333: 52–65.
Redford,
K.H. 1992. The empty forest. BioScience, 42: 412–422.
Redmond, I., Aldred, T., Jedamzik, K. & Westwood, M.
2006.
Recipes for survival: controlling the bushmeat trade. London, Ape Alliance
& World Society for the Protection of Animals.
Reed, J, van Vianen, J., Foli, S., Clendenning, J., Yang, K., MacDonald, M.,
Petrokofsky, G., Padoch, Ch., Sunderland, T.
2017. Trees for life: The ecosystem service contribution of trees to food production
and livelihoods in the tropics.
Forest Policy and Economics, 84: 62–71.
Reid, H. & Huq, S. 2005. Climate change-biodiversity and
livelihood impacts. In C. Robledo, M. Kanninen & L. Pedroni, eds.
Tropical forests and adaptation to climate change, pp. 57–70. Bogor,
Indonesia, CIFOR.
Reij, C.,
Tappan, G. & Smale, M. 2009. Agroenvironmental transformation
in the Sahel. Another kind of “Green Revolution”. IFPRI Discussion Paper
00914. Washington, DC, IFPRI.
Reimchen T.E. & Arbellay, E. 2019. Influence of spawning salmon
on tree-ring width, isotopic nitrogen, and total nitrogen in old-growth Sitka spruce
from coastal British Columbia. Canadian Journal of Forest Research, 49:
1078–1086.
Reimoser, F. 2000. Income from hunting in mountain forests of the
Alps. In M.F. Price & N. Butt, eds. Forests in sustainable mountain
development: a state of knowledge report for 2000, pp. 346–353. IUFRO
Research Series No. 5. New York, CABI
Publishing.
Repetto,
R. 1992. Accounting for environmental assets. Scientific
American, 266(6): 94–101.
Reyes-Garcia, V., Guèze, M., Luz, A.C., Paneque-Gálvez, J., Macía, M.J.,
Orta-Martínez, M., Pino, J. & Rubio-Campillo, X.
2013. Evidence of traditional knowledge loss among a contemporary indigenous
society.
Evolution and Human Behavior, 34(4): 249–257.
Ribeiro, M.C., Metzger, J.P., Martensen, A.C., Ponzoni, F.J. & Hirota,
M.M.
2009. The Brazilian Atlantic Forest: How much is left, and how is the remaining
forest distributed? Implications for conservation. Biological Conservation,
142(6): 1141–1153.
Ribot, J.C. 2002. Democratic
decentralization of natural resources: institutionalizing popular
participation. Washington, DC, WRI.
Ripple, W.J., Newsome, T.M., Wolf, C., Dirzo, R., Everatt, K.T., Galetti,
M., Hayward, M.W.
et al. 2015. Collapse of the world’s largest herbivores. Science
Advances, 1: e1400103 [online]. [Cited 5 January 2020].
https://doi.org/10.1126/sciadv.1400103
Ripple, W.J., Abernethy, K., Betts, M.G., Chapron, G., Dirzo, R., Galetti,
M., Levi, T.
et al. 2016. Bushmeat hunting and extinction risk to the world’s mammals.
Royal Society Open Science, 3: 160498 [online]. [Cited 5 January 2020].
https://doi.org/10.1098/rsos.160498
Ritchie, H., Roser, M., Mispy, J. & Ortiz-Ospina, E.
2018. SDG Tracker: Indicator 15.1.2. In:
SDG Tracker [online]. Oxford, UK. [Cited 19 December 2019].
https://sdg-tracker.org/biodiversity#15.1.2
Rivers, M.C., Beech, E., Bazos, I., Bogunić, F., Buira, A., Caković, D.,
Carapeto, A.
et al. 2019. European red list of trees. Cambridge, UK, IUCN.
RNZ.
2019. Calls to train a million UK volunteers to tackle invasive species. In:
RNZ [online]. Wellington, New Zealand. [Cited 5 January 2020].
www.rnz.co.nz/news/world/401840/calls-to-train-a-million-uk-volunteers-to-tackle-invasive-species
Roberts,
P. 2019. Tropical forests in prehistory, history, and
modernity. Oxford, UK, Oxford University Press.
Rodas, A. & Stoian, D. 2015. Determinación de los
beneficios socioeconómicos del aprovechamiento forestal percibidos por tres
comunidades con concesiones comunitarias en el Petén, Guatemala. Report of
the ADA Community Forestry Project in
Mesoamerica. Petén, Guatemala, Bioversity International.
Rohr, J.R., Civitello, D.J., Halliday, F.W., Hudson, P.J., Lafferty, K.D.,
Wood, C.L. & Mordecai, E.A.
2019. Towards common ground in the biodiversity–disease debate. Nature Ecology
& Evolution, 4: 24–33.
Rook,
G.A. 2013. Regulation of the immune system by biodiversity from the
natural environment: an ecosystem service essential to health. PNAS,
110(46): 18360–18367.
Roosevelt, A.C., Lima da Costa, M., Lopes Machado, C., Michab, M., Mercier,
N., Valladas, H., Feathers, J.
et al. 1996. Paleoindian cave dwellers in the Amazon: the peopling of the
Americas. Science, 272(5260): 373–384.
Roper, B.B., Saunders, W.C. & Ojala, J.V.
2019. Did changes in western federal land management policies improve salmonid
habitat in streams on public lands within the Interior Columbia River Basin?
Environmental Monitoring and Assessment, 191:574 [online]. [Cited 5 January
2020].
https://doi.org/10.1007/s10661-019-7716-5
Rowland, D., Blackie, R.R., Powell, B., Djoudi, H., Vergles, E., Vinceti, B.
& Ickowitz, A.
2015. Direct contributions of dry forests to nutrition: a review. International
Forestry Review, 17(S2): 45–53.
Rowland, D., Ickowitz, A., Powell, B., Nasi, R. & Sunderland,
T.
2017. Forest foods and healthy diets: quantifying the contributions.
Environmental Conservation, 44(2): 102–114.
RRI. 2015. Protected areas and the land rights of indigenous
peoples and local communities: current issues and future agenda.
Washington, DC, RRI.
RSCN. 2018.
Report on the benefits generated by local communities from DBR
[in Arabic]. Amman, RSCN.
RSCN and
Wild Jordan. 2017. Explore Dana: Jordan’s rift valley
spectacular. Brochure. Amman, RSCN.
Ruf, F. & Zadi, H. 1998. Cocoa: from deforestation to
reforestation. In:
Smithsonian’s National Zoo & Biology Institute
[online]. Washington, DC. [Cited 5 January 2020].
https://nationalzoo.si.edu/scbi/migratorybirds/research/cacao/ruf.cfm
Ruijs, A. & Vardon, M. 2019. Natural capital accounting for
mainstreaming biodiversity in public policy making. In Vardon, M., Bass,
S., and Ahlroth, S. eds.
Natural Capital Accounting for Better Policy Decisions: Climate change and
Biodiversity. Proceedings and Highlights of the 3rd Forum on Natural Capital
Accounting for Better Policy Decisions, pp. 73–100. World Bank WAVES,
Washington D.C.
Ruokolainen, L., Von Hertzen, L., Fyhrquist, N., Laatikainen, T., Lehtomäki,
J., Auvinen, P. & Knip, M.
2015. Green areas around homes reduce atopic sensitization in children.
Allergy, 70(2): 195–202.
Sabogal, C., Besacier, C. & McGuire, D.
2015. Forest and landscape restoration: concepts, approaches and challenges for
implementation. Unasylva, 245: 3–10.
Sacande, M., Jøker, D., Dulloo, M.E. & Thomsen, K.A.
eds. 2004.
Comparative storage biology of tropical tree seeds. Rome, IPGRI.
Sachedina, H. & Nelson, F. 2012. The development of payments
for ecosystem services as a community-based conservation strategy in East Africa.
In J. Ingram, F. DeClerck & C. Rumbaitis del Rio, eds.
Integrating ecology and poverty reduction: the application of ecology in
development solutions, pp. 149–171. New York, USA, Springer.
Sarkar, D., Walker-Swaney, J. & Shetty, K.
2019. Food diversity and indigenous food systems to combat diet-linked chronic
diseases.
Current Developments in Nutrition, nzz099 [online]. [Cited 5 January 2020].
https://doi.org/10.1093/cdn/nzz099
Sassen,
M. 2014. Conservation in a crowded place: forest and people on
Mount Elgon, Uganda. Wageningen University. (PhD thesis)
Sassen, M., Arnell, A.P. & van Soesbergen, A.
forthcoming.
Mapping risks to biodiversity and ecosystem services from cocoa-driven
deforestation in West Africa.
Sassen, M., Sheil, D., Giller, K.E. & ter Braak, C.J.
2013. Complex contexts and dynamic drivers: understanding four decades of forest
loss and recovery in an East African protected area. Biological
Conservation, 159: 257–268.
Saunders, C.D., Brook, A.T. & Meyers, O.E.
2006. Using psychology to save biodiversity and human well-being. Conservation
Biology, 20: 702–705.
Save the Elephants. 2019. Welcome to The Elephants and Bees
Project [online]. Nairobi. [Cited 5 January 2020].
https://elephantsandbees.com
Sayer, J.A., Campbell, B., Petheram, L., Aldrich,
M., Perez, M., Endamana, D., Nzooh Dongmo, Z.-L. et al. 2007.
Assessing environment and development outcomes in conservation
landscapes. Biodiversity and Conservation, 16(9), 2677–2694.
Sayer, J.A., Margules, C., Boedhihartono, A.K., Sunderland, T., Langston,
J.D., Reed, J., Riggs, R.
et al. 2017. Measuring the effectiveness of landscape approaches to
conservation and development. Sustainability Science, 12: 465–476.
Schelley, C., Cross, J.E., Franzen, W.S., Hall, P. & Reeve, S.
2012. How to go green: creating a conservation culture in a public high school
through education, modelling, and communication. Journal of Environmental
Education, 43(3): 143–161.
Schroth, G., Harvey, C.A., da Fonseca, G.A., Vasconcelos, H.L., Gascon, C.
& Izac, A.M.N.
eds. 2004.
Agroforestry and biodiversity conservation in tropical landscapes.
Washington, DC, Island Press.
Schroth, G., Läderach, P., Martinez-Valle, A.I., Bunn, C. & Jassogne,
L.
2016. Vulnerability to climate change of cocoa in West Africa: Patterns,
opportunities and limits to adaptation.
Science of the Total Environment, 556: 231–241.
Schueler, S., Falk, W., Koskela, J., Lefèvre, F., Bozzano, M., Hubert, J.,
Kraigher, H., Longauer, R. & Olrik, D.C.
2014. Vulnerability of dynamic genetic conservation units of forest trees in Europe
to climate change.
Global Change Biology, 20: 1498–1511.
Schulp, C.J., Thuiller, W. & Verburg, P.H.
2014. Wild food in Europe: A synthesis of knowledge and data of terrestrial wild
food as an ecosystem service.
Ecological Economics, 105: 292–305.
Schuster, R., Germain, R.R., Bennett, J.R., Reo, N.J. & Arcese,
P.
2019. Vertebrate biodiversity on indigenous-managed lands in Australia, Brazil, and
Canada equals that in protected areas. Environmental Science and Policy,
101: 1–6.
Schweik,
C.M. 2000. Optimal foraging, institutions and forest change: A case
from Nepal. Environmental Monitoring and Assessment, 62: 231–260.
SEGeF. 2018.
Suivi de la gestion de la faune dans les forêts de production
[online]. Yaoundé. [Cited 13 January 2020].
http://151.236.37.239/segef/public/
Shackleton, S., Paumgarten, F., Kassa, H., Husselman, M. & Zida,
M.
2011. Opportunities for enhancing poor women’s socioeconomic empowerment in the
value chains of three African non-timber forest products (NTFPs).
International Forestry Review, 13(2): 136–151.
Shaffer, L.J., Khadka, K.K., Van Den Hoek, J. & Naithani, K.J.
2019. Human-elephant conflict: a review of current management strategies and future
directions.
Frontiers in Ecology and Evolution, 6: 235 [online]. [Cited 5 January
2020].
https://doi.org/10.3389/fevo.2018.00235
Shanahan, D.F., Lin, B.B., Bush, R., Gaston, K.J., Dean, J.H., Barber, E.
& Fuller, R.A.
2015. Towards improved public health outcomes from urban nature. American
Journal of Public Health, 105: 470–477.
Sharpe,
B. 1998. First the forest: conservation, community and participation in
south-west Cameroon. Africa, 68(1): 25–45.
Shisegar,
N. 2014. The impact of green areas on mitigating urban heat island
effect: a review. International Journal of Environmental Sustainability, 9:
119–130.
Sichuan Forestry Department. 2015. The Pandas of Sichuan: The
4th Survey Report on Giant Panda in Sichuan Province. Chengdu, China,
Sichuan Science and Technology Press. Cited by Brinckmann, J.A., Luo W., Xu Q., He
X., Wu J., & Cunningham A.B. 2018.
Sustainable harvest, people and pandas: Assessing a decade of managed wild harvest
and trade in Schisandra sphenanthera. Journal of
Ethnopharmacology, 224: 522–534.
Silva, L.N., Freer-Smith, P. & Madsen, P.
2019. Production, restoration, mitigation: a new generation of plantations. New
Forests, 50(2): 153–168.
Sinovas, P., Price, B., King, E., Hinsley, A. & Pavitt, A.
2017.
Wildlife trade in the Amazon countries: an analysis of trade in CITES listed
species. Technical report prepared for the Amazon Regional Program
(BMZ/DGIS/GIZ). Cambridge, UK, UNEP-WCMC.
Sirén, A.
& Machoa, J. 2008. Fish, wildlife, and human nutrition in tropical
forests: a fat gap? Interciencia, 33: 186–193.
Skole, D.
& Tucker, C.J. 1993. Tropical deforestation and habitat
fragmentation in the Amazon: satellite data from 1978 to 1988. Science,
260(5116): 1905–1910.
Soares-Filho, B., Moutinho, P., Nepstad, D., Anderson, A., Rodrigues, H.,
Garcia, R., Dietzsch,L.
et al. 2010. Role of Brazilian Amazon protected areas in climate change
mitigation. PNAS, 107(24): 10821–10826.
Solecki,
R. 1975. Shanidar IV, a Neanderthal flower burial in northern Iraq.
Science, 190(4217): 880–881.
Song, X.P., Hansen, M.C., Stehman, S.V., Potapov, P.V., Tyukavina, A.,
Vermote, E.F. & Townshend, J.R.
2018. Global land change from 1982 to 2016. Nature, 560: 639–643.
Southworth, J., Nagendra, H. & Munroe, D.K.
2006. Introduction to the Special Issue: Are parks working? Exploring
human-environment tradeoffs in protected area conservation. Applied
Geography, 26(2): 87–95.
Spies, T.A., Stine, P.A., Gravenmier, R., Long, J.W., Reilly, M.J., tech.
coords.
2018.
Synthesis of science to inform land management within the Northwest Forest Plan
area. 3 volumes. General Technical Report PNW-GTR-966. Portland, OR, USA,
US Department of Agriculture, Forest Service, Pacific Northwest Research Station.
Stanturf, J., Mansourian, S. & Kleine, M., eds.
2017.
Implementing forest landscape restoration: A practitioner’s guide. Vienna,
IUFRO-SPDC.
Stanturf, J.A., Palik, B.J. & Dumroese, R.K.
2014. Contemporary forest restoration: a review emphasizing function. Forest
Ecology and Management, 331: 292–323.
St. John, F.A.V, Edwards-Jones, G. & Jones, J.P.G.
2010. Conservation and human behaviour: lessons from social psychology. Wildlife
Research, 37: 658–667.
Stattersfield, A.J., Crosby, M.J., Long, A.J.,
and Wege, D.C.
1998.
Endemic bird areas of the world: priorities for biodiversity conservation.
Cambridge, UK, BirdLife International.
Stavert, J.R., Pattemore, D.E., Gaskett, A.C., Beggs, J.R. & Bartomeus,
I.
2007. Exotic species enhance response diversity to land-use change but modify
functional composition.
Proceedings of the Royal Society B – Biological Sciences, 284(1860):
20170788 [online]. [Cited 13 January 2020].
https://doi.org/10.1098/rspb.2017.0788
Steffen, W., Richardson, K., Rockström, J., Cornell, S.E., Fetzer, I.,
Bennett, E.M., Biggs, R.
et al. 2015. Planetary boundaries: Guiding human development on a changing
planet. Science, 347(6223): 1259855 [online]. [Cited 5 January 2020].
https://doi.org/10.1126/science.1259855
Stevens, C., Winterbottom, R., Springer, J. & Reytar, K.
2014.
Securing rights, combating climate change: How strengthening community forest
rights mitigates climate change. Washington, DC, WRI.
Stoian, D. & Rodas, A. 2018. Successful community
stewardship of tropical forests: evidence from community forest concessions in
Petén, Guatemala. Paper presented at the 19th Annual Conference on Land and
Poverty held by the World Bank in Washington
DC on March 19–23, 2018 [online]. [Cited 5 January 2020].
https://cgspace.cgiar.org/bitstream/handle/10568/93439/Successful_Stoian_2018.pdf?sequence=1
Stoian, D., Rodas, A., Butler, M., Monterroso, I. & Hodgdon, B.
2018.
Forest concessions in Petén, Guatemala: A systematic analysis of the
socioeconomic performance of community enterprises in the Maya Biosphere
Reserve. Bogor, Indonesia, CIFOR.
Stolton, S., Redford, K.H., Dudley, N., Bill, W., Corcuera, E. &
Mitchel, B.A.
2014. The futures of privately protected areas. Gland, Switzerland, IUCN.
Strassburg, B., Beyer, H.L., Crouzeilles, R. Iribarrem, A., Barros, F.,
Siqueira, M., Sánchez-Tapia, A.
et al. 2019. Strategic approaches to restoring ecosystems can triple
conservation gains and halve costs. Nature Ecology & Evolution, 3:
62–70.
Sunderland, T., Sunderland-Groves, J., Shanley, P. & Campbell,
B.
2009. Bridging the gap: how can information access and exchange between conservation
biologists and field practitioners be improved for better conservation outcomes?
Biotropica, 41(5): 549–554.
Sunderlin, W.D., Angelsen, A., Belcher, B., Burgers, P., Nasi, R., Santoso,
L. & Wunder, S.
2005. Livelihoods, forest, and conservation in developing countries: an overview.
World Development, 33(9): 1383–1402.
Tamosiunas, A., Gražulevičienė, R., Luksiene, D., Dedele, A., Reklaitiene,
R., Baceviciene, M., & Milinaviciene, E.
2014. Accessibility and use of urban green spaces, and cardiovascular health:
findings from a Kaunas cohort study.
Environmental Health, 13: 20 [online]. [Cited 5 January 2020].
https://doi.org/10.1186/1476-069X-13-20
Taniwaki, R.H., Leal, C.G., de Barros Ferraz, S.F., Henrikson, L., Jägrud,
L. & de Paula, F.R.
2018.
Blue Targeting Tool: a simple forestry planning for riparian buffer zones
adapted to Brazilian streams. Poster presented at the Joint Conference on
Forests and Water, 2018, Valdivia, Chile. [also available at
https://www.researchgate.net/publication/329102135_Blue_Targeting_Tool_a_simple_forestry_planning_for_riparian_buffer_zones_adapted_to_Brazilian_streams].
Tauli-Corpuz, V., Alcorn, J. & Molnar, A.
2018.
Cornered by protected areas: replacing ‘fortress’ conservation with rights-based
approaches helps bring justice for indigenous peoples and local communities,
reduces conflict, and enables cost-effective conservation and climate
change. Washington, DC, RRI.
The Guardian. 2020. A rewilding triumph: wolves help to reverse
Yellowstone degradation. In: The Guardian [online]. [Cited 15 January
2020].
https://www.theguardian.com/environment/2020/jan/25/yellowstone-wolf-project-25th-anniversary
Tibesigwa, B., Siikamäki, J., Lokina R. & Alvsilver J.
2019. Naturally available wild pollination services have economic value for nature
dependent smallholder crop farms in Tanzania. Scientific Reports, 9: 3434
[online]. [Cited 5 January 2020].
https://doi.org/10.1038/s41598-019-39745-7
TNC. 2019. Tropical Forest Conservation Act. Benefits for Natural
Resources and the American People. In: The Nature Conservancy [online].
[Cited 15 February 2020].
https://www.nature.org/en-us/about-us/who-we-are/how-we-work/policy/tropical-forest-conservation-act/
Tracewski, Ł., Butchart, S.H.M., Donald, P.F., Evans, M., Fishpool, L.D.C.
& Buchanan, G.M.
2016. Patterns of twenty-first century forest loss across a global network of
important sites for biodiversity.
Remote Sensing in Ecology and Conservation, 2(1): 37–44.
TRAFFIC. 2019. African elephants: elephant conservation and the
global trade in ivory. In: TRAFFIC [online]. Cambridge, UK. [Cited 5
January 2020].
www.traffic.org/what-we-do/species/elephants-ivory
Triguero-Mas, M., Dadvand, P., Cirach, M., Martínez, D., Medina, A.,
Mompart, A., Basagaña, X., Gražulevičienė, R. & Nieuwenhuijsen,
M.J.
2015. Natural outdoor environments and mental and physical health: relationships and
mechanisms.
Environment International, 77, 35–41.
Tropical Forest Alliance. 2017.
The Role of the Financial Sector in Deforestation-Free Supply Chains.
Tropical Forest Alliance and World Economic Forum, Geneva. [also available at
https://www.vivideconomics.com/wp-content/uploads/2019/08/TFA2020_Framing_Paper_030117.pdf].
Turner,
I. 1996. Species loss in fragments of tropical rain forest: a review of
the evidence. Journal of Applied Ecology, 33: 200–209.
Turner, B.L. & Sabloff, J.A. 2012. Classic Period collapse of
the Central Maya Lowlands: Insights about human–environment relationships for
sustainability. PNAS, 109(35):13908–13914.
UAESPNN. 2005.
Plan de manejo Parque Nacional Natural Macuira 2005–2009
[National Natural Park Makuira, Management Plan 2005–2009]. Bogota, Parques
Nacionales Naturales de Colombia.
Udawatta, R.P., Rankoth, L.M. & Jose, S.
2019. Agroforestry and biodiversity.
Sustainability, 11(10): 2879 [online]. [Cited 5 January 2020].
https://doi.org/10.3390/su11102879
UN. 1992a. United Nations Convention
on Biological Diversity. New York, USA. [also available at
https://www.cbd.int/doc/legal/cbd-en.pdf].
UN. 1992b. United Nations Framework Convention on Climate
Change. New York, USA. [also available at https://unfccc.int/resource/docs/convkp/conveng.pdf].
UN. 1992c. United Nations Convention on to Combat
Desertification. New York, USA. [also available at
https://www.unccd.int/sites/default/files/relevant-links/2017-01/UNCCD_Convention_ENG_0.pdf]
UN. 2008a. United Nations Declaration on the Rights of
Indigenous Peoples. New York, USA. [also available at https://www.un.org/esa/socdev/unpfii/documents/DRIPS_en.pdf].
UN.
2008b. World urbanization prospects: The 2007 revision. New York, USA.
UN. 2015. Paris Agreement. New York, USA. [also available
at
https://unfccc.int/files/essential_background/convention/application/pdf/english_paris_agreement.pdf].
UN. 2017a. United Nations Strategic Plan for Forests 2017–2030. In:
United Nations Department of Economic and Social Affairs – Forests
[online]. New York, USA. [Cited 5 January 2020].
www.un.org/esa/forests/documents/un-strategic-plan-for-forests-2030/index.html
UN. 2017b. New York Declaration on Forests (list of endorsers
updated in July 2017). New York, USA. [also available at
https://www.undp.org/content/dam/undp/library/Environment%20and%20Energy/Forests/New%20York%20Declaration%20on%20Forests_DAA.pdf].
UN.
2019a. The Sustainable Development Goals Report 2019. New York, USA.
UN. 2019b. General Assembly. Seventy-third Session. 107th
plenary meeting, Monday, 16 September 2019, 10 a.m., New York. A/73/PV.107.
New York, USA. [also available at https://undocs.org/en/A/73/PV.107].
UN. 2020. SDG indicators: Metadata repository. In:
United Nations, Department of Economic and Social Affairs, Statistics
Division
[online]. New York, USA. [Cited 5 January 2020].
https://unstats.un.org/sdgs/metadata
UN, European Commission, FAO, IMF,
et al. 2014a. System of Environmental Economic Accounting 2012 —
Central Framework. New York. [also available at
http://unstats.un.org/unsd/envaccounting/seeaRev/SEEA_CF_Final_en.pdf]
UN, European Commission, FAO, OECD,
et al. 2014b.
System of Environmental Economic Accounting 2012 — Experimental Ecosystem
Accounting.
New York. [also available at
http://unstats.un.org/unsd/envaccounting/seeaRev/eea_final_en.pdf]
United Nations General Assembly. 2008. 62/98. Non-legally
binding instrument on all types of forests. Resolution adopted by the General
Assembly on 17 December 2007. A/RES/62/98. New York, USA. [also available
at
https://undocs.org/en/A/RES/62/98].
United Nations General Assembly. 2015a. Transforming our world:
the 2030 Agenda for Sustainable Development. Resolution adopted by the General
Assembly on 25 September 2015. A/RES/70/1. New York, USA. [also available
at
https://undocs.org/en/A/RES/70/1].
United Nations General Assembly. 2015b. Tackling illicit
trafficking in wildlife. Resolution adopted by the General Assembly on 30 July
2015. A/RES/69/314. New York, USA. [also available at
https://undocs.org/en/A/RES/69/314].
UNCCD. n.d. The LDN Fund – An impact investment fund for land
degradation neutrality. In:
United Nations Convention to Combat Desertification
[online]. Bonn, Germany. [Cited 2 January 2020].
https://www.unccd.int/actions/impact-investment-fund-land-degradation-neutrality
UNCCD. 2018. Decision 7/COP.13. The future strategic framework
of the Convention. Bonn, Germany. [also available at
https://www.unccd.int/sites/default/files/relevant-links/2018-08/cop21add1_SF_EN.pdf].
UNCCD. 2019a. The LDN target setting programme. In:
United Nations Convention to Combat Desertification
[online]. Bonn, Germany. [Cited 5 January 2020].
www.unccd.int/actions/ldn-target-setting-programme
UNCCD. 2019b. The GGW aims to restore Africa’s degraded landscapes
and transform millions of lives in one of the world’s poorest regions. In:
United Nations Convention to Combat Desertification
[online]. Bonn, Germany. [Cited 5 January 2020].
https://knowledge.unccd.int/publications/ggw-aims-restore-africas-degraded-landscapes-and-transform-millions-lives-one-worlds
UNCTAD. 2006. International Tropical Timber Agreement,
2006. TD/TIMBER.3/12. Geneva, Switzerland. [also available at https://treaties.un.org/doc/source/docs/tdtimber3d12_en.pdf].
UNDESA. 2016. Documents – UN forest instrument. In:
United Nations Department of Economic and Social Affairs
[online]. [Cited 18 December 2019].
https://www.un.org/esa/forests/documents/un-forest-instrument/index.html
UNDP. 2017. What is biodiversity finance? In:
United Nations Development Programme – BIOFIN – The Biodiversity Finance
Initiative
[online]. New York, USA. [Cited 5 January 2020].
www.biodiversityfinance.net/about-biofin/what-biodiversity-finance
UNECE & FAO. 2018.
Forests and Water. Valuation and payments for forest ecosystem services.
Geneva. [also available at
https://www.unece.org/fileadmin/DAM/timber/publications/sp-44-forests-water-web.pdf]
UNEP. 1979. Convention on the Conservation of Migratory Species
of Wild Animals. Nairobi. [also available at
https://www.cms.int/sites/default/files/instrument/CMS-text.en_.PDF].
UNEP. 2019. Global environment outlook GEO-6. Summary for
policy makers. Cambridge, UK, Cambridge University Press.
UNEP-WCMC. 2007. A spatial analysis approach to the global
delineation of dryland areas of relevance to the CBD Programme of Work on Dry
and Subhumid Lands. Cambridge, UK.
UNEP-WCMC. 2020. Welcome to the
global ICCA Registry [online]. Cambridge, UK. [Cited 5 January 2020].
http://www.iccaregistry.org/
UNEP-WCMC & IUCN. 2019. World Database on Protected Areas. In:
Protected Planet [online]. Cambridge, UK. [Cited 31 December 2019].
https://www.protectedplanet.net/c/world-database-on-protected-areas
UNEP-WCMC, IUCN & NGS. 2020. Protected Planet Digital
Report [online]. Cambridge, UK, Gland, Switzerland and Washington, DC.
[Cited 18 December 2019]
https://livereport.protectedplanet.net
UNEP-WCMC & UNSD. 2019.
Assessing the linkages between global indicator initiatives, SEEA Modules and
the SDG Targets.
Working Document. [also available at
https://seea.un.org/sites/seea.un.org/files/seea_global_indicator_review_methodological_note_post_workshop_0.pdf]
UNESCO. 1971. Convention on wetlands of international
importance especially as waterfowl habitat. Paris. [also available at
https://treaties.un.org/doc/Publication/UNTS/Volume%20996/volume-996-I-14583-English.pdf].
UNFCCC. 2011.
Report of the Conference of the Parties on its 16th session, Cancun, Mexico, 29
November – 10 December 2010. Addendum: Part Two: Action taken by the Conference
of the Parties at its sixteenth session. Decision 1/CP.16. The Cancun
Agreements: Outcome of the work of the Ad Hoc
Working Group on Long-term Cooperative Action under the Convention.
FCCC/CP/2010/7/Add.1. Bonn, Germany. [also available at https://unfccc.int/resource/docs/2010/cop16/eng/07a01.pdf].
UNODC. 2016. World wildlife crime report: Trafficking in
protected species 2016. Vienna.
USAID. 2017. Countries with TFCA Programs. In: UNAID
[online]. Washington, DC. [Cited 2 January 2020].
https://www.usaid.gov/biodiversity/TFCA/programs-by-country
USDA. n.d.a. Northwest Forest Plan. In:
United States Department of Agriculture, Forest Service
[online]. Washington, DC. [Cited 2 January 2020].
https://www.fs.usda.gov/detail/r6/landmanagement/planning/?cid=fsbdev2_026990
USDA. n.d.b. 5022: Wild crop harvesting. In:
United States Department of Agriculture, Agricultural Marketing Service
[online]. Washington, DC. [Cited 2 January 2020].
https://www.ams.usda.gov/rules-regulations/organic/handbook/5022
US Fish
& Wildlife Service. 1998. Recovery plan for the Oregon
chub (Oregonichthys crameri). Portland, OR, USA.
US Fish & Wildlife Service. 2018. North American Model of
Wildlife Conservation. In: US Fish & Wildlife Service, Hunting
[online]. Washington, DC. [Cited 1 January 2020].
https://www.fws.gov/hunting/north-american-model-of-wildlife-conservation.html
US/ICOMOS. 2019. Heritage trees: international research and
registries. In: US/ICOMOS [online]. Washington, DC. [Cited 5 January 2020].
https://usicomos.org/heritage-trees-international-research-and-registries/
Uusivuori, J., Lehto, E. & Palo, M.
2002. Population, income and ecological conditions as determinants of forest area
variation in the tropics.
Global Environmental Change, 12: 313–323.
Valencia, R., Balslev, H. & Paz y Miño, G.C.
1994. High tree alpha-diversity in Amazonian Ecuador.
Biodiversity & Conservation, 3:21– 28, cited by Dirzo, R. & Raven,
P. H. 2003. Global state of biodiversity and loss. Annual Review of Environment
and Resources, 28: 137–167.
van Lierop, P., Lindquist, E., Sathyapala, S. & Franceschini,
G.
2015. Global forest area disturbance from fire, insect pests, disease and severe
weather events.
Forest Ecology and Management 352: 78–88.
Van Vliet, N., Muhindo, J., Nyumu, J.K. & Nasi, R.
2019. From the forest to the dish: A comprehensive study of the wildmeat value chain
in Yangambi, Democratic Republic of Congo. Frontiers in Ecology and
Evolution, 7: 132 [online]. [Cited 5 January 2020].
https://doi.org/10.3389/fevo.2019.00132
Verdone, M. & Seidl, A. 2017. Time, space, place, and the Bonn
Challenge global forest restoration target. Restoration Ecology, 25(6):
903–911. [also available at http://dx.doi.org/10.1111/rec.12512]
Verissimo,
D. 2013. Influencing human behaviour: an underutilised tool for
biodiversity management. Conservation Evidence, 10: 29–31.
Verschuuren,
B. & Brown, S. eds. 2018. Cultural and spiritual significance
of nature in protected areas: Governance, management and policy. Abingdon,
UK, Routledge.
Vié, J.-C., Hilton-Taylor, C. & Stuart, S.N.
eds. 2009.
Wildlife in a changing world: an analysis of the 2008 IUCN Red List of
Threatened Species. Gland, Switzerland, IUCN.
Vlam, M., van der Sleen, P., Groenendijk, P. & Zuidema, P.A.
2017. Tree age distributions reveal large-scale disturbance-recovery cycles in three
tropical forests.
Frontiers in Plant Science, 7: 1984 [online]. [Cited 5 January 2020].
https://doi.org/10.3389/fpls.2016.01984
Vogt, P. 2019a. Image object Accounting
(Available in the free JRC software
GuildosToolbox). Ispra, Italy, European Commission Joint Research Center,
Directorate for Sustainable Resources. [also available at
https://ies-ows.jrc.ec.europa.eu/gtb/GTB/psheets/GTB-Objects-Accounting.pdf].
Vogt, P. 2019b. Measuring Forest Area Density
to quantify forest fragmentation. (Available in the free JRC software
GuidosToolbox). Ispra, Italy, European Commission Joint Research Center,
Directorate for Sustainable Resources. [also available at
https://ies-ows.jrc.ec.europa.eu/gtb/GTB/psheets/GTB-Fragmentation-FADFOS.pdf
Vogt, P., Riitters, K.H., Caudullo, G., Eckhardt, B.
2019.
FAO – State of the World’s Forests: Forest fragmentation. JRC Technical
Report, EUR 29972 EN. Luxembourg, Publications Office of the European Union. [also
available at
https://publications.jrc.ec.europa.eu/repository/bitstream/JRC118594/technicalreport_fao_frag.pdf].
Vorontsova, M.S., Clark, L.G., Dransfield, J., Govaerts, R. & Baker,
W.J.
2016. World checklist of bamboos and rattans. INBAR Technical Report No.
37, Beijing, INBAR.
Walker, X.J., Baltzer, J.L., Cumming, S.G., Day,
N.J., Ebert, C., Goetz, S., Johnstone, J.F. et al. 2019.
Increasing wildfires threaten historic carbon sink of boreal forest
soils. Nature, 572: 520–523.
Watson, E.E. 2005. Gender-sensitive natural resource management
(NRM) research-for-development. DFID NRSP Programme Development Report
PD123: Gender Sensitive Research for Development. Cambridge, UK, University of
Cambridge, Department of Geography.
Watson, J.E.M., Dudley, N., Segan, D.B. & Hockins, M.
2014. The performance and potential of protected areas.
Nature, 515: 67–73.
Watson, J.E.M., Evans, T., Venter, O., Williams, B., Tulloch, A., Stewart,
C., Thompson, I.
et al. 2018. The exceptional value of intact forest ecosystems. Nature
Ecology & Evolution 2: 599–610.
WEF. 2020. One trillion trees – World Economic Forum launches plan
to help nature and the climate. In: World Economic Forum [online]. Geneva,
Switzerland. [Cited 15 February 2020].
https://www.weforum.org/agenda/2020/01/one-trillion-trees-world-economic-forum-launches-plan-to-help-nature-and-the-climate/
West, P., Igoe, J., & Brockington, D.
2006. Parks and peoples: the social impact of protected areas. Annual Review of
Anthropology, 35: 251–277.
White, M.P., Alcock, I., Wheeler, B.W. & Depledge, M.H.
2013. Would you be happier living in a greener urban area? A fixed-effects analysis
of panel data.
Psychological Science, 24(6): 920–928.
WHO. 2002. WHO Traditional medicine strategy: 2002–2005.
Geneva, Switzerland.
WHO. 2016. Ambient air pollution: a global assessment of
exposure and burden of disease. Geneva, Switzerland.
WHO. 2017. 5. Annex 5. Guidelines for the production, control
and regulation of snake antivenom immunoglobulins. Replacement of Annex 2 of WHO
Technical Report Series, No. 964. Geneva, Switzerland.
WHO. 2018a. Household air pollution and health. In: World
Health Organization [online]. Geneva, Switzerland. [Cited 5 January 2020].
www.who.int/news-room/fact-sheets/detail/household-air-pollution-and-health
WHO. 2018b. Air pollution: Maps and databases. In: World Health
Organization [online]. Geneva, Switzerland. [Cited 5 January 2020].
www.who.int/airpollution/data/en
WHO. 2019. Traditional, complementary and integrative medicine:
About us. In: World Health Organization [online]. Geneva, Switzerland.
[Cited 5 January 2020].
www.who.int/traditional-complementary-integrative-medicine/about
WHO. 2020. Q&A on coronaviruses (COVID-19). In: World
Health Organization [online]. Geneva, Switzerland. [Cited 1 April 2020].
www.who.int/news-room/q-a-detail/q-a-coronaviruses
WHO/UNICEF. 2000. Global water supply and sanitation assessment
2000 report. Geneva, Switzerland, WHO/UNICEF Joint Monitoring Programme for
Water Supply and Sanitation.
Wilcox, B.A.
& Ellis, B. 2006. Forests and emerging infectious diseases of
humans. Unasylva, 224: 11–18.
Wilkie, D.S., Wieland, M., Boulet, H., Le Bel, S., van Vliet, N., Cornelis,
D., BriacWarnon, V., Nasi, R. & Fa, J.E.
2016. Eating and conserving bushmeat in Africa.
African Journal of Ecology, 54: 402–414.
Willett, W., Rockström, J., Loken, B., Springmann, M., Lang, T., Vermeulen,
S., Garnett, T.
et al. 2019. Food in the Anthropocene: the EAT–Lancet Commission on healthy
diets from sustainable food systems. Lancet, 393(10170): 447–492.
Willis, K.J.
ed. 2017. State of the World’s Plants 2017. Richmond, Surrey,
Kew Publishing.
Willis, K.J.
ed. 2018. State of the World’s Fungi 2018. Richmond, Surrey,
Kew Publishing.
Winfree, R., Williams, N.M., Dushoff, J. & Kremen, C.
2007. Native bees provide insurance against ongoing honey bee losses. Ecology
Letters, 10: 1105–1113.
Winfree, R., Aguilar, R., Vazquez, D.P., LeBuhn, G. & Aizen,
M.A.
2009. A meta-analysis of bees’ responses to anthropogenic disturbance.
Ecology, 90: 2068–2076.
Witt, K.A. 2013. The nutrient content of Moringa oleifera
leaves. In: ECHO Community [online]. North Fort Myers, FL, USA. [Cited 5
January 2020].
https://www.echocommunity.org/resources/a7ee06e3-40f2-4ef0-859e-4e64b90a56c8
World Agroforestry. 2009. The Agroforestree Database. In: World
Agroforestry [online]. Nairobi. [Cited 13 January 2020].
http://www.worldagroforestry.org/output/agroforestree-database
World
Bank. 2002. A revised forest strategy for the World Bank
Group. Washington, DC.
World Bank. 2017. Guidebook on Ecosystem Accounting.
Washington, DC. [also available at https://elibrary.worldbank.org/doi/pdf/10.1596/29829].
World Bank. 2019.
Global Wildlife Programme Phase 2: Summarized version of child projects
[online]. [Cited 5 January 2020].
https://www.thegef.org/sites/default/files/webdocuments/10200_PFD_Wildlife_Annex_ChildProjects.pdf
World Cocoa Foundation. 2017. Cocoa & Forests Initiative:
Statement of intent. In: World Cocoa Foundation [online]. Washington, DC.
[Cited 5 January 2020].
www.worldcocoafoundation.org/cocoa-forests-initiative-statement-of-intent/
World Land Trust. n.d. Golden-headed lion tamarin. In: World
Land Trust [online]. Halesworth, Suffolk, UK. [Cited 5 January 2020].
www.worldlandtrust.org/species/mammals/golden-headed-lion-tamarin
WWF. 2018. WWF Tanzania set to implement debt for nature swap
programme. In: WWF [online]. [Cited 5 January 2020].
wwf.panda.org/?324230/WWF-Tanzania-Set-to-implement-Debt-for-Nature-Swap-Programme
WWF
China. 2012. Standards for Giant Panda friendly products.
Chengdu, China.
Yearsley. 2019. FairWild project in India is a win-win-win for
Terminalia trees, people, and hornbills. HerbalEGram, 16(6) [online].
[Cited 5 January 2020].
http://cms.herbalgram.org/heg/volume16/06June/FairWildTerminalia.html
Zhang, D.
& Pearse, P.H. 2011. Forest economics. Vancouver, UBC
Press.
Zhu, H., Xu, Z.F., Wang, H. & Li, B.G.
2004. Tropical rain forest fragmentation and its ecological and species diversity
changes in southern Yunnan.
Biodiversity and Conservation, 13(7): 1355–1372.
Zomer, R.J., Trabucco, A., Coe, R. & Place, F.
2009.
Trees on farm: analysis of global extent and geographical patterns of
agroforestry. ICRAF Working Paper 89. Nairobi, Kenya, World Agroforestry
Centre.
ZSL & WWF. 2014. Living Planet Index [online]. [Cited
26 December 2019].
http://www.livingplanetindex.org/home/index