![]()
![]()
![]()
• Difficulties in experimental hatchery
By Mr. Ksouri Jamel (INSTOP - Tunisia)
• Final maturation and ovulation in marine fish
By Mr. Ramos M.A. (Instituto Nacional de Investigacao das Pescas - Portugal)
• Water management in marine fish hatchery
By Mr. Ayari Abdelaziz (AST-Tunisia)
• Nutritional management in hatchery
By Mr. Patrick Lavens (Belgium)
• Planning, dimensionning & maintenance criteria
for sea water hatchery building and equipment
By Mr. Alessandro Moretti (STM - Italy)
• Management of clams hatchery
By Mr. Jean-Pierre Band (IFREMER - France)
• MAROST experience in shellfish hatchery
By Mr. Jamel Kasmi (MAROST - Morocco)
• Shrimp hatchery in MAROST
By Mr. Rabah Boukabous (MAROST - Morocco)
By Mr. Lazaro Rosa
SPAIN
TECHNICAL MANAGEMENT
Reproduction Area
1-GENERATING
Quick Stable
Natural Stable
Preparation to spawn
2-QUALITIES
Eggs
Larval
3-METHODOLOGY
Water
Air
Worked systems
FOOD AREA
1.1- PHITO
Work production line
1.2- ROTIFERO (Brackionus Picadilli)
Alternation systems
1.3- ARTEMIA
1.4- INERT FEED
1.5- COORDINATION
Effectiveness
Valuation about economic
1 - ANALISIS TEORICO DE COSTES, PARA LA PRODUCCION DE ALEVINES DE DORADAS EN HATCHERY.
La obtencion de una production potencial de 4 a 5 milliones de alevines de doradas, requierirà los siguientes volumenes de inversion y gastos de explotacion:
| Inversion: | ||||||
| A- | Construccion de una HATCHERY (Edificacion, instalaciones y equipos) presupuesto: | |||||
| 2 milliones de Dolares | 206 milliones de pesetas. | |||||
| Explotacion: (Supuesto de produccion de 4 millones de Alevines de 2–3 grs | ||||||
| B- | Costes de Personal | |||||
| (Incluida Segoridad Social) | Pesetas | |||||
| Direccion Tecnica | 7,300,000 | |||||
| 6 Tecnicos. | 29,215,800 | |||||
| 12 Especialistas | 25,615,800 | |||||
| 4 Guardas o Vigilantes | 9,576,000 | |||||
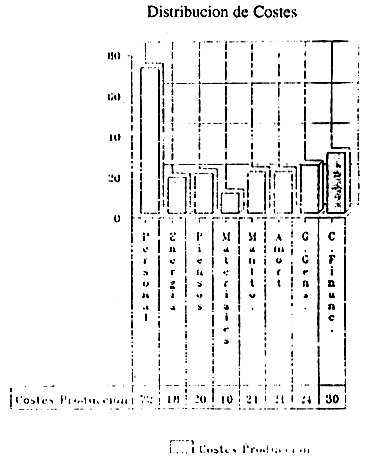
| Total Coste de Personal | 71,707,600 | |||||
| C- | Energia eléctrica | 18,000,000 | ||||
| D- | Piensos (5 ptas/Und.) | 20,000,000 | ||||
| E- | Material y productos sanitarios | 10,000,000 | ||||
| F- | Mantenimiento de Instalaciones | 20,600,000 | ||||
| G- | Amortizacion Instalaciones | 20,600,000 | ||||
| H- | Gastos Generales | 24,000,000 | ||||
| I- | Costes de financiacion | 29,585,000 | ||||
| Total Costes de Explotacion 214,492,600 | ||||||
Distribucion del Personal de Produccion:
| Tecnicos | Especialtas | Total | |
| Direccion | 3 | - | 3 |
| Alimentacion | 2 | 4 | 6 |
| Quimico | 1 | 1 | 2 |
| Prod. y Engorde | 1 | 7 | 8 |
| Totales | 7 | 12 | 19 |
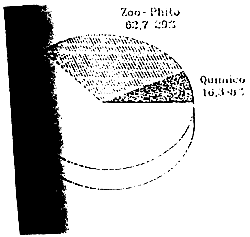
Distribucion de los costes de produccion por departamentos:
| Direccion | % | Almentacion | Quimico | Prod-Bngorde | Total | % | ||||
| Personal Técnico | 17,038,000 | 35.7 | 18,277,600 | 37.8 | 7,003,950 | 47.2 | 19,812,050 | 19.1 | 62,131,600 | 28.97 |
| Guarderias | - | - | 2,872,800 | 5.9 | - | - | 6,703,200 | 6.5 | 9,576,000 | 4.46 |
| Energia | - | - | 6,300,000 | 13.1 | 900,000 | 6.1 | 10,800,000 | 10.4 | 18,000,000 | 8.39 |
| Piensos | - | - | 4,000,000 | 8.3 | - | - | 16,000,000 | 15.5 | 20,000,000 | 9.32 |
| Materiales | 100,000 | 0.2 | 2,000,000 | 4.1 | 4,000,000 | 26.9 | 3,900,000 | 3.8 | 10,000,000 | 4.66 |
| Mantenimiento | - | - | 4,120,000 | 8.5 | 300,000 | 2.0 | 16,180,000 | 15.6 | 20,600,000 | 9.60 |
| Amortizacines | - | - | 4,120,000 | 8.5 | 600,000 | 4.0 | 15,880,000 | 15.3 | 20,600,000 | 9.60 |
| Castos Generales | 24,000,000 | 50.3 | - | - | - | - | - | - | 24,000,000 | 11.19 |
| Costes Financieros | 6,582,032 | 13.8 | 6,670,415 | 13,8 | 2,.048,617 | 13.8 | 14,283,936 | 13.8 | 29,585,000 | 13.79 |
| SUBTOTALES | 47,720,032 | 100 | 48,360,815 | 14,852,567 | 100 | 103,559,186 | 100 | 214,492,600 | 100 | |
| Porcentaje S/total Gastos | 22.2 | 22.5 | 6.92 | 48.3 | ||||||
| Reclasificacion Direccion | 14,316,010 | 30.0 | 1,431,600 | 3,0 | 31,972,422 | 67,0 | ||||
| 62,676,825 | 29,2 | 16,284,167 | 7.59 | 135,531,608 | 63.2 | |||||
| Costes Totale de Produccion | |||||
| 1- | Costes Cadena Alimentaciion | 62,676,825 | |||
| 2- | Costes Quimico-sanitarios | 16,284,167 | |||
| 3- | Costes Cadena de Engorde | 135,531,608 | |||
| Total Costes de Produccion 214,492,600 | |||||
| Numéro de Unidades Estimadas Producidas | 4,000,000 | ||||
| Coste Unitario de Produccion | 53,62 ptas | ||||
| Precia Estimadeo de Ventas | 70 Pts/und. | ||||
| Margen de Beneficio por unidad | 16.38 Ptas | ||||
| Rentabilidad de las Ventas | 23,40 % | ||||
Costed por Sessiones
| Costes Totale de Producción | |||||
| 1.- | Costes Cadena Alimentación | 62,676,825 | |||
| 2.- | Costes Quimico-Sanitarios | 16,284,167 | |||
| 3.- | Costes Cadena de Engorde | 135,531,608 | |||
| Total Costes de Producciion 214,492,600 | |||||
| Numéro de Unidades Estimadas Producidas | 4,000,000 | ||||
| Coste Unitario de Produccion | 53,62 ptas | ||||
| Precia Estimadeo de Ventas | 70 Pts/und. | ||||
| Margen de Beneficio por unidad | 16.38 Ptas | ||||
| Rentabilidad de las Ventas | 23,40 % | ||||
| Rentabilidad Económica Neta | 31.81% | ||||
 |  |
Élevage larvaire du loup Dicentrarchus labrax à la station de Ghar El Melh - Tunisie
By Mr. Jamel KSOURI
et Mr. Fredj KARTAS
TUNIS
RÉSUMÉ
Dans cette étude, nous présentons les techniques adoptées et les résultats enregistrés à la station aquacole de Ghar El Melh au cours de la campagne 1988–1989 d'élevage larvaire et de prégrossissement de loup Dicentrarchus labrax.
La ponte induite a engendré un pourcentage de viabilité des oeufs de 72,5% et le nombre d'oeufs viables émis par une femelle de 1 kg a été égal à 224.000.
Le taux d'éclosion était de 60%. Au terme de 80 jours d'élevage, 42.000 alevins sevrés pesant 0,3 gr. et représentant un taux de survie du 7% ont été produits.
Le prégrossissement a duré 40 jours et a débouché sur des alevins de 1 à 2 gr. représentant du 83,3% du nombre initial d'alevins.
ABSTRACT
In this study we present the techniques and the results of Ghar El Melh Station during the compaign of 1988–1989 concerning the larval rearing and the preswelling of sea bass Dicentrarchus labrax.
One female of 1 kg gave 224.000 viables eggs. This represents a percentage of 72.5% viability 60% of eggs hatched.
Àfter 80 days of larval rearing 42.000 fry of 0,3 gr were produced. This represents 7% of the larvaes of one day.
The preswelling lasted 40 days and gave 83,3% as survivial rate for fry of 1 to 2 gr.
INTRODUCTION
En pisciculture marine, le grossissement intéresse en général des juvéniles sauvages piégés lors des migrations trophiques de la mer vers les estuaires ou les lagunes ou des alevins produits en conditions contrôlées en écloseries. L'étape «écloserie» regroupe de nombreux stades interdépendants et nécessite des techniques aussi sophistiqués que coûteuses. Elle concerne, de ce fait, des espèces «nobles» à haute valeur commerciale telles que le loup, la daurade, la sole et la crevette pouvant supporter les charges afférentes à la nourriture, au fonctionnement des structures d'élevage et à la main d'oeuvre spécialisée. L'aboutissement, au terme de 3 à l'élevin de loup de 1 á 2 gr. implique la passage par les principales étapes suivantes: l'obtention des oeufs, leur incubation et leur éclosion, l'élevage larvaire, le sevrage et le prégrossissement en plus de la culture du plancton. Les techniques pratiquées en écloserie sont encore en pleine évolution et, à l'exception de la sole, espèce particulièrement rustique, le taux de mortalité dépasse généralement 50% un mois après l'éclosion (Person-Leruyet et Bedier, 1984).
En Tunisie, la première écloserie a été construite en 1973 par une campagne française, puis acquise en 1977 par l'Institut National Scientifique et Technique d'Océanographie et de Pêche (INSTOP).
Initialement, l'activité de cette écloserie s'est limitée au stockage des géniteurs de loup et de daurade dans des bassins en ciment de 90 m3, à la production du plancton dans des conditions extensives, à l'élevage larvaire dans des bassins en ciment de 17m3.
Les conditions d'élevage s'étant avérées aléatoires, un changement progressif, entrepris à partir de 1982, a hissé les activités à un niveau de recherche et d'expérimentation marqué notamment par:
Nous développons dans les paragraphes ci-dessous les procédés de l'élevage larvaire et du prégrossissement tels qu'ils sont pratiqués durant la saison 1988–1989.
STOCKAGE DES GÉNITEURS
Le rôle de l'unité des géniteurs est de subvenir aux besoins en oeufs de l'écloserie. Mais dans certains cas, il est possible de faire recours à d'autres sources d'approvisionnement en oeufs fertilisés telles que les géniteurs sauvages, les autres écloseries et l'extraction des oeufs de poissons fraîchement pêchés.
D'après Barnabé (1976), la charge en géniteurs de daurade et de loup dans des bassins de volume supérieur à 20m3 ne devrait pas dépasser un seuil estimé à 4 km/m3 car elle agit négativement sur la ponte compte tenu de la compétition et de l'agressivité.
Du point de vue alimentation, Giavenni (1987) remarque qu'en zootechnie, des poissons suralimentés ou sous-alimentés ne peuvent pas constituer de bons géniteurs. D'après Barnabé (1973), une alimentation trop homogène amènerait certaines carences chez les reproducteurs. L'alimentation des géniteurs par des poissons frais ou décongelés distribués deux fois par semaine et à des quantités de l'ordre de 5% du poids du cheptel est suffisante (Barnabé, 1986).
Pour Devauchelle (1987), la ration hebdomadaire de ce type d'aliment équivalente à 4–24% du poids des reproducteurs est fournie en 2 ou 3 fois.
Les géniteurs que nous avons utilisés sont nés à l'écloserie de Ghar El Melh et maintenus captifs depuis au moins cinq ans. Ils sont stockés à une charge de 4 kg/m3 dans un bassin en ciment rectangulaire de 13 cm3 doté d'une couverture légère évitant le développement d'algues et l'exposition prolongée des géniteurs aux rayons lumineux.
Le pompage de l'eau est effectué à partir de la mer avec un débit qui assure un renouvellement de 15 à 20% du volume de bassin en une heure. Ce mode de nouvellement continu d'eau engendre des conditions physico-chimiques caractérisées au cours de l'année par une température variable entre 10 et 29°C, une salinité comprise entre 36,5 et 39,8% et un taux d'oxygène dissous allant de 4,2 à 7,5 ppm. C'est au cours du mois d'août que ces paramètres ont atteint des valeurs critiques.
De surcroît, les bassins de stockage des géniteurs font régulièrement l'objet d'un siphonnage de nature à éviter l'accumulation des déchets sur le fond.
Il est à noter que ces conditions de stockage n'ont occasionné aucune mortalité du cheptel même après 4 à 6 heures d'arrêt de pompage.
L'alimentation de nos géniteurs est basée essentiellement sur des sardines et des saurels et, secondairement, sur des poissons provenant des rebuts de chalutage qu' on distribue après décongélation, en petit morceaux en deux à trois fois par semaine à une quantité globale de 5 à du poids des reproducteurs.
OBTENTION DES OEUFS
La ponte induite par voie hormonale ne conduit pas à des oeufs viables lorsque la vitellogénèse n'est pas achevée (Barnabé, 1976). A Ghar El Melh, la température de l'eau de mer atteint, au cours du mois de janvier, la valeur optimale de ponte, soit environ 30°C. A cette époque, les géniteurs dont les gonades sont à un stade de maturation avancée sont sélectionnés d'après le gonflement abdominal pour les femelles et l'émission du sperme à la suite d'une légère pression sur les flancs pour les mâles. Ceux jugés aptes à pondre sont placés dans une bassine contenant du phénoxy-2-éthanol à la dose de 0,3 ppm, anesthésiant destiné à réduire les mouvements des poissons. Les géniteurs sont mesurés, pesés et injectés. Le poids des femelles est en moyenne de 0,9 kg et varie de 0,6 à 1,3 kg. La poids des mâles est de 0,7 kg en moyenne et est compris entre 0,6 et 1 kg.
La ponte est provoquée par une injection de gonadotrophine chorionique humaine pratiquée à la base de la deuxième nageoire dorsale à une dose de 800 à 1.000 U.I. par kg de poids vif pour les femmelles et 500 U.I. pour les mâles.
Les reproducteurs sont alors transférés, à raison de deux mâles pour trois femelles et à une charge de 4 kg/m3, dans un bassin de ponte circulaire de 5m3 ombragé par une bâche et où le renouvellement horaire d'eau est de 30 M du volume.
Au terme d'une période de latence d'environ trois jours après l'injection, une première ponte est observée; la fécondation ayant eu lieu naturellement dans le bassin de ponte. Les oeufs sont récupérés par surverse dans un collecteur cylindrique en toile de 400 μm de maille, puis dénombrés par la méthode volumétrique sur la base de l'équivalence entre 1 ml de 800 oeufs.
La séparation des oeufs viables se fait dans une éprouvette de 1 à 21; ces oeufs remontent à la surface au bout de 5 à 15 mm tandis que les oeufs morts sédimentent.
Les oeufs viables hémisphériques et limpides possèdent un disque germinatif et présentent un diamètre moyen de 1.118 μm. Ils peuvent contenir jusqu'à 7 gouttelettes lipidiques; mais 75% d'entre eux n'en contiennent qu'une ou deux. La plus grosse gouttelette mesure 350 μm. Le tableau 1 illustrant la qualité de la ponte montre qu'une femelle de 1 kg émet par acte de ponte environ 224.000 oeufs viables (75,2% su total) à raison de 32.000 par jour. Le nombre d'oeufs viables que nous avons obtenu est nettement supérieur à celui cité dans la littérature: 100.000 (CNEXO, 1983; France Aquaculture, 1984) et 180.000 (Barnabé, 1986).
Tableau 1 - Qualité de la ponte issue d'un kg de femelle
| Date d'injection: | 4/1/1989 |
| Durée de la ponte (jour) | 7 |
| Nombre moyen d'oeufs viables émis par 24 h. | 32.000 |
| Nombre d'oeufs viables émis pendant la durée de la ponte | 224.000 |
| Pourcentage d'oeufs viable | 72,5 |
OBTENTION DES LARVES
L'incubation des oeufs peut se faire dans des unités spécialement prévues à cet effet ou directement dans les bacs d'élevage larvaire à 18–20°C (Freddi, 1985) ou à une température stable comprise entre 13 et 17°C (CNEXO, 1983).
A l'écloserie de Ghar El Melh, l'incubation s'effectue dans des bacs d'élevage larvaire, cylindroconiques en polyester armé et de 1m3 de volume. Le débit d'eau assure un renouvellement horaire de 40 à 70% du volume du bac et l'aération, émanant des diffuseurs placés au fond de l'incubateur; permet une bonne distribution des oeufs et évite leur stratification. L'incubation se déroule à l'obscurité pendant 3–4 jours à la température de 13°C.
A la naissance des prélarves, l'estimation du pourcentage d'éclosion est réalisée par comptage sur cinq échantillons de 1.1 prélevés sur toute la surface du bac; l'aération étant augmentée momentanément pour homogénéiser le milieu. Le taux d'éclosion moyen était de 60% contre des valeurs de 90, 80, et 60% signalées par Barnabé (1976), CNEXO (1983) et France Aquaculture (1984).
ÉLEVAGE LARVAIRE
Chez les poissons marins, l'élevage larvaire concerne la phase allant de la larve de 4–5 jours à l'alevin âgé de 60–90 jours. La délicatesse de cette étape est due à la conjugaison de plusieurs facteurs tels que la qualité de l'eau, la photopériode, la charge et la nourriture. Leur synergie conditionne la croissance, la survie, les maladies et les malformations larvaires.
Il est établi que la température optimale de l'eau d'élevage se situe autour de 18–22°C. La forme, les dimensions, le mode de construction et le principe de fonctionnement des bassins destinés à recevoir les larves sont très variables (Brunel, 1987). La forme peut être circulaire, sub-carrée, raceway ou cylindro-conique; cette dernière étant actuellement la plus répandue. Le volume varie du 1m3 pour les élevages très intensifs à 100m3 pour les élevages extensifs. Ils sont construits en béton armé, en polyester, en polyéthylène ou en contre plaqué avec une bâche en PVC, le polyester étant prédominant. L'élevage peut être très intensif (plus de 150 oeufs/litre) ou extensif (10 oeufs/litre); des densités moyennes de 100 oeufs/litre sont les plus couramment utilisées.
UNITÉ D'ÉLEVAGE LARVAIRE
A l'écloserie de Ghar El Melh, l'élevage larvaire se déroule dans des bacs de 1m3 à la densité moyenne initiale de 60 larves au litre. Comme le montre le schéma 1, nous disposons de deux unités d'élevage regroupant au total 19 bacs et formant une partie intégrante avec le biofiltre et l'unité de production et de stockage des proies vivantes. Une couverture légère en plastique permet de minimiser les déperditions thermiques au niveau de chaque unité; l'ensemble est abrité dans un hangar à toit translucide permettant la pénétration de lumière tamisée. L'eau des bacs est recyclée à travers un filtre biologique. Le schéma 2 illustre une vue générale de ce filtre et met en relief les différentes parties le constituant:
un décanteur comportant des chicanes servant à ralentir la circulation de l'eau à ce niveau et à amener la plus grande parties des particules solides en suspension à sédimenter;
un lit bactérien composé de deux parties: une première (a) constituée d'une couche de coquilles d'huîtres au-dessus de laquelle trois canalisations en PVC de 75 mm de diamètre perforées sur toute la longueur assurent l'étalement de l'eau et son infiltration à travers toute la couche de coquilles; i'eau traverse ainsi dans un sens descendant toute la colonne de support du filtre; une deuxième partie (b) formée de tubes en PVC de 250 mm de diamètre remplis de morceau de tuyau en plastique d'environ 5 cm. de longueur, mise en place récemment pour empêcher la dégradation de la qualité de l'eau observée au cours des campagnes précédentes;
une enceinte renfermant des thermoplongeurs de 1 à 2 kW assurant le chauffage de l'eau;
un compartiment de stérilisation dans lequel l'eau circule sur une lame mince au voisinage immédiat d'une série de lampes ultraviolet pour éliminer les éventuels germes pathogènes et éviter la contamination des bacs indemnes;
une pompe refoulant l'eau vers un château d'eau alimentant par écoulement gravitaire les deux unités d'élevage.
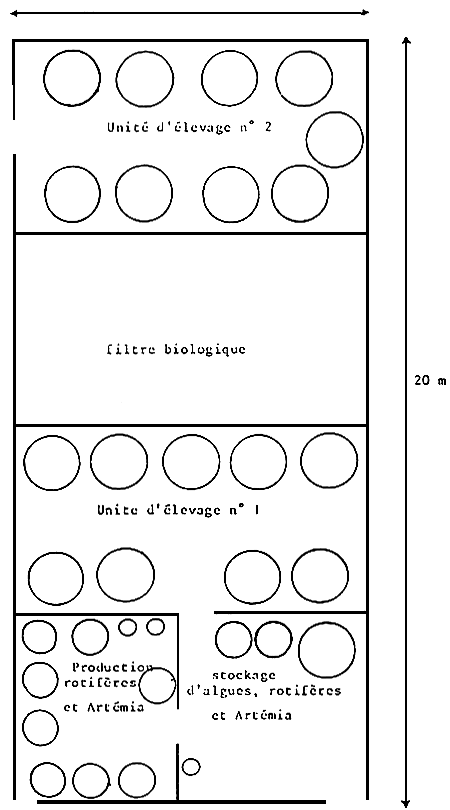
Schéma 1: Unité d'élevage larvaire en intensif
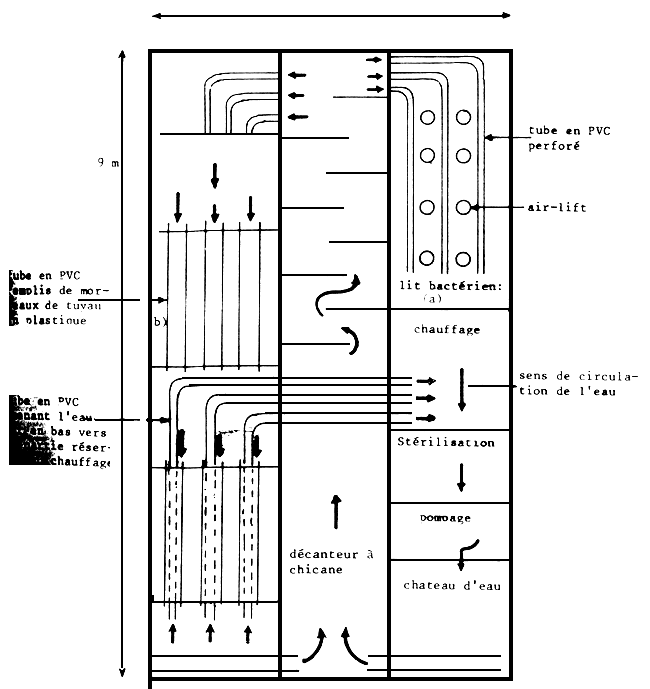
Schéma 2 : vue générale du filtre biologique.
CONDITIONS DU MILIEU
L'élevage larvaire est effectué à une température comprise entre 16 et 18°C. Le taux d'oxygène dissous est de 5 à 7 ppm jusqu'au 45ème jour et varie entre 4 et 5 ppm au-delà. L'aération est assurée par des diffuseurs d'air placés au fond des bacs et dont le débit est modulé en fonction de la phase d'élevage : ce débit est élevé au cours de l'incubation des oeufs afin d'éviter leur stratification et modéré durant l'élevage pour éviter la turbulence de l'eau. Le renouvellement d'eau. Le renouvellement d'eau est ajusté en fonction de l'étape d'élevage : à l'éclosion, il est faible et égal à 15–20 % du volume par heure et ceci pour éviter que le courant d'eau n'entraîne les larves encore fragiles et passives vers le centre du bac où elles s'écrasent contre les parois du filtre d'évacuation et le colmatent; ensuite, il augmente progressivement pour attendre 70 % en début de sevrage et 100 % à la fin de l'élevage larvaire (Tabl. 2).
Le nettoyage et le changement des filtres d'évacuation, ainsi que le siphonnage des bacs, se font d'une façon régulière.
Tableau 2 - Alimentation en eau des bacs de l'élevage larvaire du loup.
| Age (jour) | Étape | Filtre d'évacuation | Aération | Renouvellement d'eau (% du volume/h) | Circuit d'eau | |
| Vide de maille | Nettoyage | |||||
| Incubation | 0,1 mm | +++ | 40 – 70 | |||
| 0 | Eclosion | Tous les jours | + | Fermé | ||
| 15 – 20 | ||||||
| 10 | ||||||
| 45 | Début de sevrage | 30 – 40 | ||||
| 1 à 2 fois par jours | ++ | |||||
| 0,4 mm | 70 – 90 | Ouvert | ||||
| 55 | Fin de sevrage | |||||
| Tous les 2–3 jours | ||||||
| 60 | ||||||
| 2 mm | Tous les 5 jours | 100 | ||||
Il importe de signaler qu'une obscurité total a été maintenue au niveau des bacs d'élevage pendant les dix premiers jours à une température de l'ordre de 16°C. En effet, parmi différents protocoles lumineux, l'équipe MEREA (1984) a obtenu le meilleur résultat de normalité des alevins (par référence à une vessie gazeuse fonctionnelle) lorsque la période d'obscurité est prolongée pendant 160°C - jours après l'éclosion. Pour la suite de l'élevage et dans le but de déterminer l'effet de la nature de la lumière sur la croissance et la survie des larves, trois bacs sont éclairés naturellement et sept autres artificiellement pendant 14 heures par jour grâce à des lampes à incandescence de 60 W placées à 50 cm de la surface de l'eau.
RÉSULTATS
La phase d'élevage larvaire a duré 80 jours, au terme desquels les 42.000 alevins dénombrés dans les 14 bacs représentent 7 % du nombre de larves de 1 jour et ont en moyenne 3.,4 cm de longueur et un poids de 0,290 gr. (tableau 3); ce qui concorde avec les observations de Barnabé (1986) qui signale que, lorsque les alevins atteignent un poids individuel de 0,3 gr., ils sont théoriquement aptes à être placés en prégrossissements.
Notons que les présents résultats sont nettement meilleur que ceux obtenus en 1987 (Ksouri, 1989) dans des conditions physico-chimiques et alimentaires défavorables ayant engendré au bout de 80 jours d'élevage larvaire une survie de 4,9 % et une croissance anormalement faible caractérisée par une longueur de 1,9 cm et un poids de 0,07 gr.
D'après la littérature, ce chiffre de 7 % que nous avons obtenu est inférieur à ceux cités par CNEXO (1983 et Barnabé (1986) pour qui l'élevage larvaire dure entre 60 et 90 jours et aboutit à un taux de survie de 9 et 10 %
Tableau 3 - Résultats global de l'élevage larvaire.
| Nombre de bacs | 14 | ||
| Nombre de larves de 1 jour (x 103) | 600 | ||
| Nombre d'alevins de 80 jours (x 103) | 42 | ||
| Taux de survie (%) | 7 | ||
| Croissance des alevins | en longueur (CM) | moyenne extrêmes | 3,4 2,8 – 4,2 |
| en poids (gr.) | moyenne extrêmes | 0,290 0,200 – 0,400 | |
Le suivi des bacs ayant fait l'objet d'une expérimentation concernant l'effet de la nature de la lumière sur la croissance et la survie des larves est consigné dans le tableau 4.
Tableau 4 - Impact de la lumière sur l'élevage larvaire.
| Nature de la lumière | Naturelle | Artificielle | |
| Nombre de bacs | 3 | 7 | |
| Nombre de larves de 1 jour (x 103) | 137 | 342 | |
| Nombre d'alevins de 80 jours (x 103) | 14 | 21 | |
| Taux de survie (%) | 10,2 | 6,1 | |
| Croissance des alevins | en longueur (cm) | 3,7 3 – 4,2 | 3,2 2,4 – 3,5 |
| en poids (gr.) | 0,310 0,230 – 0,400 | 0,260 0,200 – 0,350 | |
Nous constatons que dans les trois bacs soumis à la lumière naturelle et chargés initialement avec 137.000 larves, la production a atteint au bout de 80 jours 14.000 alevins; ce qui représente une survie de 10,2 %. La longueur et le poids moyen étaient respectivement de 3,7 cm et 0,310 gr. En revanche, les sept bacs, maintenus sous un éclairage artificiel et totalisant 342.000 larves de 1 jour, ont engendré une production de 21.000 alevins mesurant 3,2 cm et pesant 0,260 gr., ce qui représente une survie de 6,1 %. Notons qu'en remplaçant la lumière artificielle par la lumière naturelle, Coves (1986) obtient une progression de la survie de 15 % à 40–50 %. Ces résultats de survie et de croissance mettent en évidence tout l'intérêt qu'il y a à réaliser l'élevage larvaire avec un éclairement naturel d'une photopériode de 9 à 12 heures.
PRÉGROSSISSEMENT
Pour le prégrossissement, les 42.00 alevins sont répartis dans les bacs d'élevage larvaire à raison de 2.500 par bac.
Les bacs sont alimentés en eau de mer filtrée dont la température varie entre 14 et 16°C. Le transit de l'eau, selon un circuit fermé par les structures du filtre biologique, a permis de contourner les difficultés inhérentes à l'interruption fréquente du débit d'eau au niveau de pompage. Car nous avons assisté pendant au moins deux mois à une baisse générale du niveau d'eau de mer; ce qui a souvent occasionné la mise à découvert de la canalisation reliant le puit duquel se fait l'aspiration de l'eau à la mer.
L'alimentation des alevins est constituée de granulés secs (alevinage 1,2 et 3) fournis par de distributeurs automatiques et par un autre aliment composé de 20 % de farine de poissons, 65 % de broyât de poissons, 5 % de gélatine et 10 % de vitamines et distribué sous forme de pâte sur des mangeoires. Au terme de 40 jours de prégrossissement, on a obtenu 35.000 alevins représentant une survie de 83,3% II est à remarquer que lorsque la durée de prégrossissement est limitée à 40 jours, la survie est de I' ordre de 80% (France Aquaculture, 1984). Trente mille de ces alevins âgés alors de 120 jours et pesant de 1 à 2 gr., ont été livrés des privés.
CONCLUSION
A l'éclosion de Ghar El Melh, l'obtention des oeufs ne pose pas de problèmes particuliers étant donné que les géniteurs sont stockés en permanence à la station en nombre satisfaisant en général la demande en oeufs des unités d' élevage larvaire. De surcroît, nous essayons d'avoir deux lots de reproducteurs d'âge différents pour pouvoir éliminer les vieux géniteurs et les remplacer par des jeunes, comme c'était le cas pour cette campagne. Des juvéniles sont, en effet, prélevés dans les lots de tête à la fin de chaque cycle de prégrossissement en vue de les élever comme futurs géniteurs.
La ponte induite par traitement hormonal a engendré un pourcentage de viabilité de 72,5% et le nombre d' oeufs viables émis par un kg de femelle étant de 224,000 peut être considéré comme bon.
L'incubation, faite à la température de eau de mer, a donné lieu à un taux d'éclosion de 60%. La phase d'élevage larvaire, effectuée dans les bacs de 1m3, a abouti au bout de 80 jours à un taux de survie de 7 % et à des alevins pesant 0,3 gr.
Nous signalons à cet égard I'intérêt de réaliser l'élevage larvaire à la lumière naturelle puisque dans ce cas, le taux de survie et de 10,2% contre 6,1 % en éclairage artificiel. Pour la croissance et la survie des larves, des améliorations peuvent être obtenues en rehaussant la température d'élevage aux normes optimales établies autour de 18–22°C et en mieux alimentant les larves du 30éme jour jusqu'au sevrage au 45éme jour. En effet, les nauplii d'Artemia stockés dans de l'eau verte pendant un jour ne satisfont plus à cette échéance les besoins des larves et de grandes mortalités sont alors observées. L'Artemia adulte ou le zooplankton collecté du milieu naturel sont de nature à augmenter le rendement de cette étape. Il est à souligne que l'extension du lit bactérien du filtre biologique a permis de maintenir notamment l'oxygène dissous dans des normes acceptables et d'éviter la dégradation de la qualité de l'eau enregistrée précocement au cours des campagnes précédentes.
En l'absence d'une unité appropriée, le prégrossissement des 42,000 alevins issus de l'élevage larvaire a été effectué dans des bacs de lm3 à faibles charges. Au bout de 40 jours, la survie était alors de 83,3%. L'unité de prégrossissement gagnerait à être dotée de raceways en polyester et d'un circuit fermé d'eau à même de soustraire l'unité aux aléas de la station de pompage.
BIBLIOGRAPHIE
• BARNABE G. (1973) - Etude morphologique du loup Dicentrarchus labrax (L.) de la région de Sète. Rev. Trav., Inst. Pêches Maritimes, 37 (3): 397–410
• BARNABE G. (1976) - Contribution à la connaissance de la biologie du loup Dicentrarchus labrax (L.) de la région de Sète. Thèse Doctorat d'État, Univ. Sci. Tech. Languedoc - Montpellier - France : 426 p.
• BARNABE G. (1986) - L'élevage du loup et de la daurade. In : Aquaculture, 2 BARNABE G. - Lavoisier - Paris : 628–666
• BRUNEL G. (1987) - Écloserie de poissons : structures, équipements, aspects économiques In : production contrôlée en écloserie - MEDRAP - Yougoslavie : 53–63
• CENTRE NATIONAL POUR L'EXPLOITATION DES OCÉANS (CNEXO) (1983) Le loup. Fich. Biotech. d'Aquaculture : 102 p.
• COVES D. (1986) - Améliorations récentes des techniques d'élevage et de la qualité des animaux produits en écloserie de loup, Dicentrarchus labrax In : Techniques d'élevage intensif et d'alimentation de poissons et de crustacés, II MEDRAP - Italie : 342–356
• DEVAUCHELLE N; (1987) - Ponte en captivité et incubation de quatre poissons marins In : Production contrôlée en écloserie - MEDRAP - Yougoslavie : 307–352
• ÉQUIPE MÉDITERRANÉENNE DE RECHERCHE AQUACOLE (1986) Maîtrise de la qualité des alevins de loup Dicentrarchus labrax produit en élevage intensif Pisc. Fr. 85 : pp. 17–23
• FRANCE AQUACULTURE (1984) - Réalisation d'une écloserie de poissons marins Deuxième offre pour la conception et la réalisation de l'écloserie de Monastir : 50 p.
• FREDI A; (1985) - Loup Dicentrarchus labrax et daurade Sparus aurata Élevage larvaire - Cours de formation en aquaculture - MEDRAP : 77 P.
• GIAVENNI R; (1987) - Pathologie des loups et des daurades In : Pathologie des espèces élevées en aquaculture marine en Méditerranée MEDRAP, Italie : 139–147
• KSOURI J (1989) - Élevage larvaire et prégrossissement de la daurade Sparus aurata et du loup Dicentrarchus labrax à la station de Ghar El Melh Thèse Doctorat Spéc., Facsc. Sci., Tunis : 264 p.
• PERSON-LE RUYET J., BEDIER E. (1984) La production de juvéniles de poissons marins en écloserie. La Pêche Maritime, 1273 : 215–222
By Mr. RAMOS M.A.
PORTUGAL
INTRODUCTION
In aquaculture technology and fishing activities, the knowledge of the relations between behaviour, growth and reproduction of fish are of scientific and practical interest. Aquatic animals are different from land animals, particularly fish, with the usual seasonal production of tiny eggs emitted in large quantities. Also, the different phases of fish embrionary development shows that there is no morphological or behavioral relation with the adult. The advancement of aquaculture technology has allowed the potential increase of fish species, through the control of reproduction and intensive larvae culture. The selection of breeders for genetic improvement of species as well as hybridization or gametes manipulation technics are of interest, specially for growth optimization.
MORPHOLOGICAL ASPECTS OF GAMETOGENESIS IN TELEOSTS CYTOLOGICAL BASE
In teleosts there is a great variety of reproductive behavior and many ways of gonad differentiation. The sex is genetically determined, but teleosts species can be gonocoric or hermaphrodite. The gonads morphogenesis regulates the stages of gametogenesis and reproduction. The interpretation of the morphological evolution of the female germinal cells is one of the aspects that expresses the dynamic of oocyte growth. The interpretation of the spermatogenesis shows that the male gametes viability is interrupted in certain light and temperature conditions. In the particular case of the protandric hermaphroditism of Sparus aurata, the involution of the testicular component of the gonad is related with sex reversal which depends on the age and growth of the animals, and their sexual and reproductive behavior in the group. In relation to the specific oocyte «timing» of the development events we can distinguish in the passage from one stage to the next that in Sparus aurata (hermaphrodite), in Dicentrarchus labrax, in Solea sp. (gonocoric) and in other fish species, the oocytes are , for most part of the time in meiotic prophase, with the chromosome at diploten and ring shape nucleolus (Azevedo, 1984).
The oogonia, after mitotic divisions, originate primary oocytes that remain in meiotic prophase. The first phase of oocytary growth or pre-vitelogenesis involves two morphological stages : the nuclear chromatin phase and the phase in which the nucleoli are placed at the periphery of the nucleus (Fig. 1).
ENDOCYTOSIS FINAL MATURATION
The second oocytary growth phase involves the endocytosis which occurs in particular conditions, after the organization of the endocytic complex, with the normal evolution of the ooplasma organella, deposition of the pellucid zone (microvilosities in which pinocitosis occurs) and the development of the follicular and these cells Fig. 2 (Roth, 1962). It is known that this phase depends on the hypophise action and involves the production of vitellogenine of hepatic origin under the influence of steof steroids. The 17B estradiol stimulates the synthesis of the lipophosphoprotein (vitellogenin, specific of females) being transported in the blood and incorporated in the oocyte as fat vesicles (acid mucopolissacharides) and vitellin globules (phospholipoprotein) (Goldstein, 1979 - Fostier, 1983) table 1.
By histoquemical methods and by ultra structural studies of the gonads it was demonstrated that, in the females, follicular cells are those responsible for the steroid hormones. In the males, the interstitial cells of the testis would be those responsible for the androgen synthesis (Fostier et al., 1983).
INVOLUTION OF GERMINAL SEX CELLS
It is known that environmental changes or adaptation to a new diet with metabolic changes can inhibit final maturation and reproduction. In the particular case of the Dicentrarchus labrax breeders, maintained in laboratory conditions, during the first year of their adaptation to captivity conditions. most of the females did not enter in vitellogenesis. Some females with secondary oocytes present the ovary in different atresia stages (Fig. 3). Gonadotrophin production can be affected by the decrease of the activity, when in particular stress conditions. During the second year of adaptation to captivity conditions the final maturation occurred in good conditions in the three groups and spontaneous ovulation took place. Probably the adaptation of the breeders in the group to each other and to environmental conditions has a fundamental importance in relation to the physiological regulation of reproduction. The sex reversal in Sparus aurata, with the involution and degeneration of the spermatozoids in the spermoduct, takes place at the same time that the oocytes in meiotic prophase are developed in the ovigerous lamellae in the female gonad component.
The ultra structural cytological study of the germinal male tissue in this phase shows that the spermatozoids motility is low because of the degeneration of the nuclei of the germinal male cells fagocited by the gonad epithelium and by the Sertoli cells. This fact is evident from the abundance of residual bodies in the cytoplasm of these cells in this phase (Fig. 4).
ENDOCRINE REGULATION OF FISH GAMETOGENESIS GAMETAS RELEASE
It is now well known that the teleosts reproduction is regulated by a large number of hormones. The gonad seasonal cycle (circanual) is developed according to the endocrine rythm of hypotalamo-hypophyse-gonad. Maturation and releasing of the gametes is connected with endocrine activity (Hoar, 1983).
The weight of the gonads is usually presented as an indication of endocrine activity, what we do not consider absolutely correct. The measurement of the gonadotrophin plasmatic levels with the circanual fluctuation shows a particular rythm of reproductive activity as well as a variation of releasing hypothalamic hormone. The hypophysis ablation inhibits maturation (Peter, 1978) but the gonadotrophin hormones induces ovulation. Additionally, final maturation and ovulation is due to the action of prostaglandin's. Pheromones are also important in what concerns sexual signals, with physiological and physical importance synchronizing the ritual response of the reproductive act.
Environmental factors, such as light or temperature, are also responsible for the regulation of the reproductive mechanism. They are specific for each species and have the capacity to stimulate or inhibit the endocrine response.
The reproductive regulation of fish is possible using hormones either to obtain final maturation and ovulation or to synchronize the gametes maturation. Administration of those hormones (HOG, LHRH, DOCA, SG100) involves biological knowledge and care in practice because it can cause death of a large number of breeders. Instead of these techniques the control of final maturation and ovulation, in a continuous and spontaneous way, was developed specially because it is easier and more practical for industrial purposes than the administration of hormones (table 2).
MATURATION DIAGNOSIS
In Sparus aurata, Dicentrarchus labrax or Solea sp. anatomic characteristics, such as abdomen enlargement, are not necessarily an indication of maturation. In many cases females with large abdomen present oocytes in the initial maturation stages. The maturation stage of the oocytes can be determined according to the methods we used as a way of applying maturation diagnosis in fish culture.
In anaesthesed females oocyte fragments are removed from the ovary using a polyethylene tube with the dimension of the genital pore. The oocyte dimension is measured using a micrometer in the microscope ocular and a maturation scale is used (table 3).
In the diagnosis process the definition of the endocytosis is fundamental to determine the hormone doses for maturation (Ramos, 1986).
ENVIRONMENTAL PARAMETERS
In general, species survival and in particular, fish species survival, depends on reproduction in the appropriate season. The fotoperiod and temperature have been recognized as important in sexual cycle regulation. In fish it is possible to establish a calendar in which each specie has its own place, because the biological regulation of reproduction through the environmental factors (temperature, light, oxygen, pH, salinity, social and of the group) is genetivaly determined in each species.
ARTIFICIAL FERTILIZATION
Artificial fertilization has been developed as a method of fish propagation. This has to be performed using diagnosis technics and by technics to induce final maturation and ovulation at the same time, synchronizing gametes releasing.
SPONTANEOUS AND CONTINUOUS REPRODUCTION
This method is now currently used to produce viable eggs every day, for larvae rearing, specially in Sparus aurata for industrial purposes. Other marine fish species can also present the same behavior, using the appropriate control of the environmental parameters.
LARVAE GROWTH
Different questions are of interest in larvae development of marine fish. When do the larvae differentiate the eyes and acquire capacity to see the food? Light and temperature have an important effect in relation to larvae survival. The perception of light and the transmission of photo information depends on the pineal gland and on the eye. The visual acuity of larvae is very poor but it increases when the pigmentation of the eye differentiates, with the evolution of photo sensibility (Ellertsen, et al., 1981).
The most important factor for larvae survival is the administration of the living or the inert food establishing a biochemical equilibrium between the evolution of the larvae (vitellin reserves) and the quality of the trophic chain.
GENETIC IMPROVEMENT
In order to produce better adapted genotypes to aquaculture, a scientific and experimental base is necessary. In fish, as in other vertebrates, hybrids, consanguineous and poliploids as well as sterile have been created (Dujakovié et al., 1990 - Purdum, 1983 - Wohlfarth, 1983).
Specimens selections is however a problem that exceeds the geneticists considering that in practice the density of fish, the manner of feeding and the water quality, as well as the temperature and oxygenation, are responsible for the growth.
GROWTH ACCELERATION
Controlled by the physiological clock, the growth of fish can be changed by administration of somatotrophin of exogenous origin. Used in salmon, the bovine somatotrophin recombinant produce a growth acceleration during the winter, the animals usually grow more slowly, due to a reduction of the anabolism as result of low temperature (Gill et al., 1985 - Shelbourn et al., 1992).
The bovine somatotrophin recombinant used in our experiences in Sparus aurata did not accelerate growth as expected.
Possessing a growth rate superior to other knows species it is probable that the sensibility of this species to external stimulus is lacking because it is already genetically prepared to grow faster (table 4). All the above mentioned technics to increase fish production present advantages and disadvantages, mostly because the genotype-environmental relation conditionates the results. Additionally, when fast growing species are used, the amount of food distributed also increases, and so the economic value of the production has to be carefully studied.
CONCLUSIONS
Aquaculture is a pluridisciplinary activity. Basic information about the anatomy and physiology of fish and how is the interplay between the external environment and the animals physiology leads to the establishment of optimum environmental conditions for different species.
The development of Technological-Biology at the experimental level is important in order to understand the functional requirements of the animals to be produced.
The manipulation of gametes to achieve final maturation, ovulation and fertilization in marine fish according to specific technologies can contribute to increase production.
REFERENCES
• AZEVEDO C., 1984 - Development and ultra structural auto radiographic studies of nucleolus like bodies (nuages) in oocyte of a viviparous teleost (Xiphophorus helleri). CELL TISSUE RES., 238 : 121–128
• DUJAKOVIC J.J., - GLAMUZINA B., 1990 - Inter generic hybridization in Sparidae 1, Sparus aurata ♀ - Diplodus puntazzo ♂ and Sparus aurata ♀ - Diplodus vulgaris ♂, Aquaculture, 86 269–378
• ELLERTSEN B.- MOKNESS E.- SOLEMDAL P.- STROEMME T.- TILSETH S.- WESTA-GARD T.- Oesiestad V, 1981 The early life history of fish : fish studies Rapp. P.V. Reun. Ciem. Vol. 178
• FOSTIER A.- JALABERT B.- BILLARD R.- BRETON B.- ZOAR Y.- 1983 The gonadal sterois. Fish Physiology - Vol. IX Reproduction Part. A. - Academic press - New York - London
• GILL J.A.- SUMPTER J.P.- DONALDSON M.E.- DYE M.H.- SOUZA L.- THOR. B.- WYPYEH J.- LANGLEY K.- 1985 Recombinant chicken and bovine growth hormones, accelerate growth in aquaculture juvenile, pacific salmon Onchorhynchus kisutch. Biotechnology Vol. 3
• GOLDSTEIN J.L. - ANDERSON R.G.W.-BROWN M.S.- 1979 Coated vesicles and receptor mediated endocytosis Nature 279: pp. 679–685
• HOAR W.S.-RANDAL D.C.-DONALDSON E.M.- 1983 Reproduction part B. Behavior and fertility control Academic Press-New York - In Fish Physiology Vol. 9
• JONHSTONE R.- KNOTT R.M.- MACDONLAD A.G. WALSINGHAM M.V. - 1989 Triploidy induction in recent fertilized Atlantic salmon ova using anaesthetics Aquaculture 78: 229–236
• PETER RE.-CHANG J.P.-COOK A.F.- 1981 Neuroendocrine regulation of gonadotropin and growth hormone secretion in goldfish Abst r. Pap. Int. Symp. Comp. Endocrinology 9th, p. 64
• PURDUM C.E.- 1983 Genetic engineering by the manipulation of chromosomes Aquaculture 33: 287–300
• RAMOS M.A.- 1986 Contribuiçäo para o conhecimento da ultrastructura do oocito e do funcionamento da gonada ha dourada (Sparus aurata L., PISCES, PERCIFORMES) Tese, INIP
• ROTH T.F.- PORTER K.R.- 1962 Yolk protein up take in the oocyte of mosquito Aeds aegypty J. Cell. Biol., 20: pp. 313–331
• SHELBOURN E.J.-CLARK C.W.- McBRIDE J.R.- FAGUERLUND U.H.M.-DONALDSON EM.- 1992 The use of 17-methyltestosterone, and 5, 5, 3'-triiodo-L-thyronine for sterilizing and accelerating the growth of zero-age coho salmon smolts (Onchorhynchus kisutch) aquaculture 103: 85–99
• WOHLFARTII G.W. - 1983 Genetics of fish applications to warm fishes Aquaculture 33: 373–381
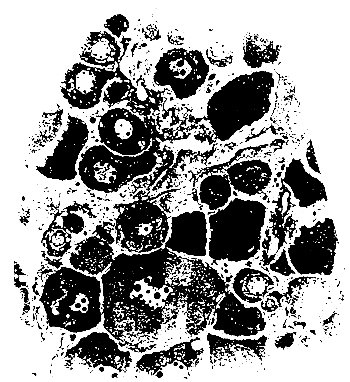 |  |
| Fig. 1 - Primary oocytes 1.500 X | Fig. 2 - Secondary oocytes 100 X |
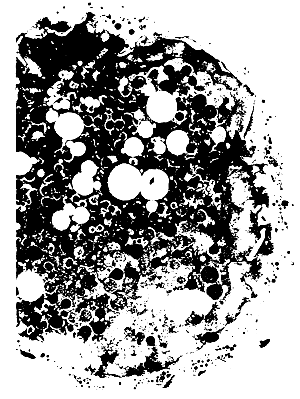 |  |
| Fig. 3 - Oocyte envolution 100 X | Fig. 4 - Spermatozoids envolution Residual bodies 6.000 X |

Table 1 - Endocytic activity of the Sparus aurata oocyte
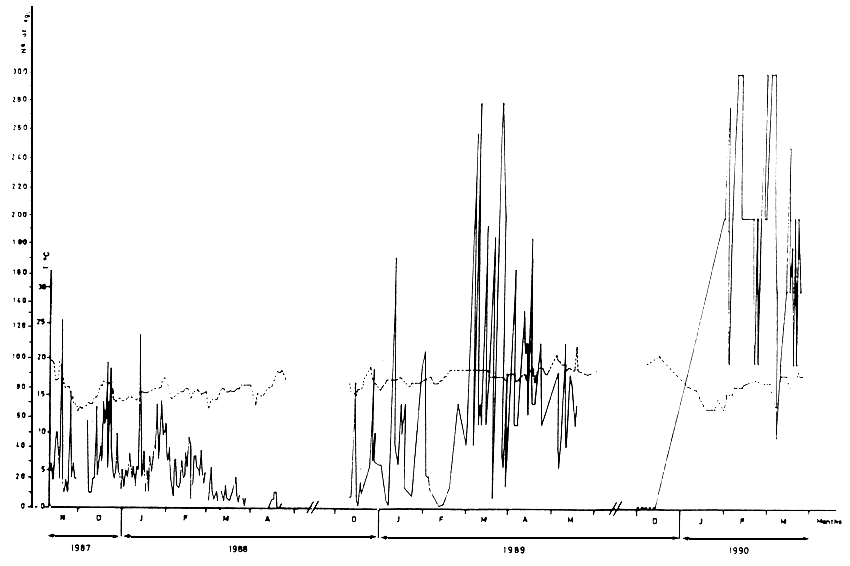
Table 2 - Production of Sparus aurata eggs - Relation with temperature.
DIFFERENTIATION OF THE AMBISSEXUAL GONAD
| ECLUSION | ||||||||
| GONOCITES | ||||||||
| 1st YEAR | Oogônia | Spermatogônia | ||||||
| Oocytes in meiotic prophase | Spermatozoid mature | Functional male | ||||||
| 2nd/3rd YEAR | Slow accumulation of RNA intermitochondrial | Reabsorption of spermatozoids in part of the population | ||||||
| Functional female | Functional male | |||||||
| Oocyte diameter in functional females in captivity | G.S.I. the wild females | |||||||
| 4th/5the YEAR | ||||||||
| Feb. | 12.5–97 | 1.0 | Slow RNA accumulation | 16–20 million Cells per ml, from October till April | ||||
| March | 24.25–99 | 0.92 | ||||||
| April | ---------- | 0.72 | ||||||
| May | 38.8.99 | 0.73 | ||||||
| Oocyte diameter | Values of the G.S.L. | |||||||
| Jun. . | 29.1–106.7 | 0,99 | Micravilosities Follicular cells Golgi at the periphery | |||||
| Jul. | 24.1–106.7 | ---------- | Vacuolar nucleoles long mitochondria dispersed | |||||
| A.ug. | ---------- | ---------- | Multivesicular bodies | |||||
| Sept. | 19.1–135 | 1.0 | Pinocitoses Depositions Pellucid zone M.V.D. accumulation | |||||
| Oct. | 19.1–488 | 1.3 | Deposition of pellucid zone. Pinocitosis cortical alvedi multivesicular bodies | |||||
| Nov. | 26.1–600 | 1.3 | Nuclear membrane invagination. Cortical alveoli at the surface. Lisosoms Pinocitosis | |||||
| Dec. | 0.5–600 | 1.5 | Nuclear membrane invagination. Cortical alveoli at the surface. Lisosoms Pinotosis | |||||
| Jan. | ---------- | 4.97 | Nuclear membrane invagination. Reticulate nucleolus cortical alveoli at the surface. Pinocitosid | |||||
| Hydratation 900–1.200 | Lipid agglutination vitelin globuli cortical alveoli at the periphery. Follicular cells separate. There is no pinocitic activity | |||||||
Sequential activity of the gonad. | ||||||||
Oocytes dimension in functional females. | ||||||||
G.S.I. in wild population. | ||||||||
Table 3 - Scale of oocyte evolution - Diagnostic of maturation
SPARUS AURATA GROWTH ACCELERATION
Table 4 - Evolution of Sparus aurata growth in weight using recombinant of somototropin
La gestion de l'eau dans une écloserie de poissons exemple: L'A.S.T.
By. Mr Abdelaziz AYARI
TUNIS
INTRODUCTION
Présentations Générale de l'A.S.T
La société AST (Aquaculture du Sud Tunisien) a été créé en 1984 sous l'impulsion d'un promoteur technique français, SEPIA International.
L'exploitation de cette ferme aquacole a démarré en 1986 avec un objectif de production de 2.400.000 alevins de 2 gr. de loup (Dicentrarchus labrax) et une production finale de 400 t.
Cette ferme est située au sud tunisien dans le golfe de Gabès, au bord d'une lagune de 50.000 ha de superficie séparée de la Méditerranée par l'île de Jerba (la lagune de Bougrara).
Présentation Technique
La ferme aquacole d'A.S.T. comprend:
une unité de géniteurs
une unité plancton
une écloserie
une unité de conditionnement
une unité de grossissement en mer (cages en mer).
LA GESTION DE L'EAU DANS UNE ÉCLOSERIE
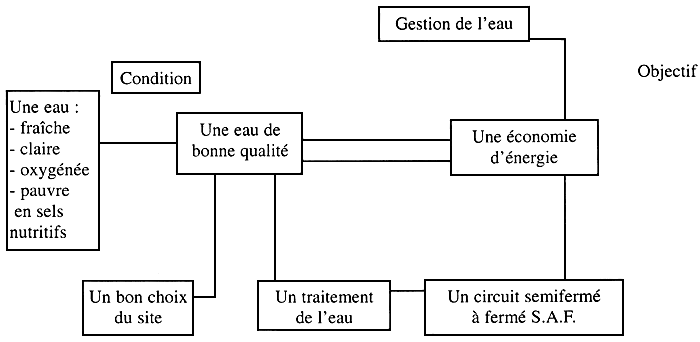
Schéma № 1
Choix du site d'une écloserie
Plusieurs critères rentrent en jeu dans le choix du site d'implantation d'une écloserie. Parmi ces critères, il y a la qualité de l'eau.
Pour avoir une de pompage de bonne qualité il faut éviter les zones suivantes:
| • | Les zones polluées: | |||
| * | Pollution chimique: | - - | les rejets industriels les ports | |
| * | Pollution organique: | - - - - | les milieux eutrophes (lagunes) les estuaires les rejets urbains les ports | |
| • | Les zones d'ensablement et de sédimentation | |||
| • | Les zones de très profondeur. | |||
Traitement de l'eau dans une écloserie de poissons (exemple: le loup)
Dans une écloserie, les besoins en eaux sont représentés sur ce schéma:
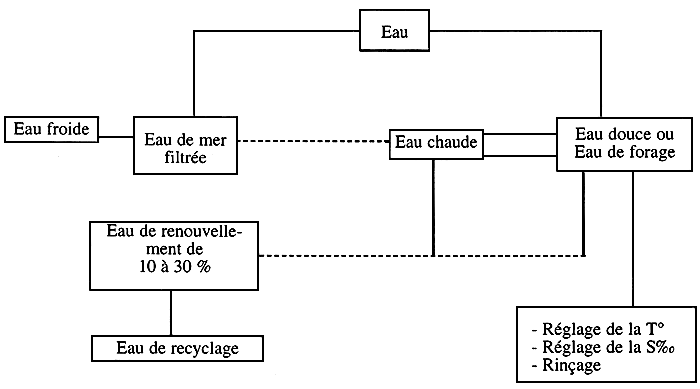
Schéma № 2
Ces différentes eaux qui alimentent l'écloserie sont traitées en fonction des exigences en qualité d'eau.
Le circuit général d'alimentation en eau dans une écloserie ainsi que les ouvrages de traitement sont représentés sur le schéma № 3:
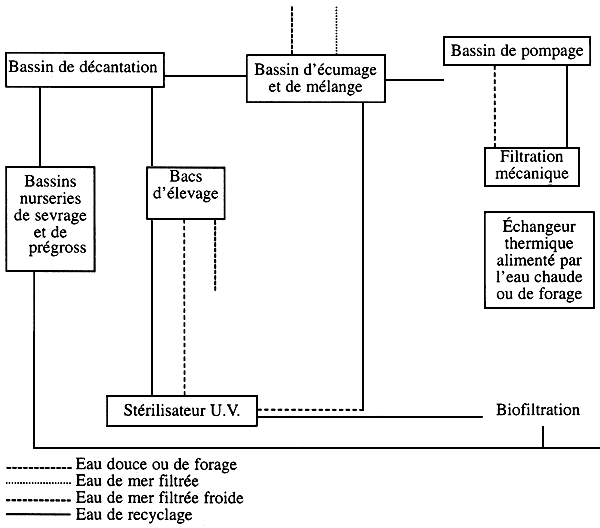
Schéma № 3
Rôle des ouvrages de traitement de l'eau
La décantation: elle consiste à vitesse de l'eau de recyclage pour permettre la sédimentation des particules en suspension les plus denses.
L'écumage: il consiste à injecter de l'air en bulles dans une colonne d'eau. Ces bulles d'air brasées avec l'eau de recyclage riche en m.e.s. s'accumulent dans le cône de la colonne d'eau et forment des mousses qui, poussées par le flux d'air sous-jacent, sont jetées à l'égout par une cheminée d'évacuation. Ce système d'épuration permet d'extraire une bonne partie de la m.e.s. la plus fine.
La filtration mécanique: elle consiste à filtrer l'eau de recyclage (également l'eau froide) sur des filtres à sables ou sur des filtres à cartouches selon les besoins en degrés de filtration et surtout en fonction du volume d'eau à filtrer. La filtration mécanique nous donne une eau de bonne transparence par l'élimination de la quasi-totalité de la m.e.s. qui a pu s'échapper à la décantation et à l'écumage. En plus de la bonne transparence de l'eau d'élevage, cette filtration poussée (décanteurs, écumeurs et filtres à sables) constitue une protection de l'échangeur thermique, des biofiltres et du stérilisateur U.V. (ouvrages capitaux dans le traitement de l'eau dans une écloserie).
L'échangeur thermique: menu d'une électrovanne de réglage thermique, l'échangeur permet le réchauffement de l'eau et la maintien de la température désirée pour l'élevage.
La biofiltration: c'est une procédure d'épuration indispensable dans le traitement des eaux de recyclage, elle assure la transformation des substances azotées toxiques (NO2 et NH4) excrétées par les larves et décomposition de la matière organique, en une forme azotée tolérée par les espèces en élevage (NO3).
L'oxydation de l'ammoniaque en nitrites et des nitrites en nitrates est assurée par les bactéries nitrifiantes fixées sur le support bactérien du biofiltre (gravier, biolithe,…).
Cette opération s'appelle la nitrification. Le cycle de l'azote est représenté sur le schéma №:
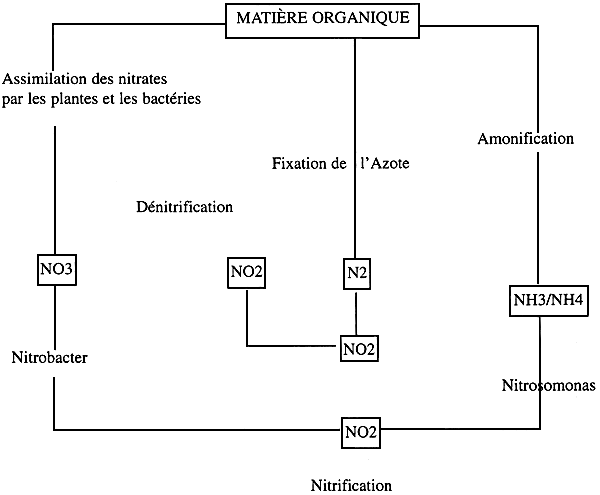
Schéma № 4
Pour bien fonctionner, le biofiltre a besoin d'une grande quantité d'oxygène dissous. Par conséquent, en sortie du biofiltre, l'eau s'appauvrit en oxygène, d'où la nécessité de réoxygéner l'eau soit par l'aération, soit par l'injection de l'oxygène pur dans le circuit de distribution d'eau en cas de déficit important en oxygène.
Un dysfonctionnement des biofiltres est marqué par une élévation du taux d'ammoniaque et du pH, par conséquent un suivi de ces deux paramètres permet de vérifier le rendement des biofiltres.
La stérilisation U.V.: elle consiste à détruire les agents pathogènes contenus dans l'eau (bactéries, virus, champignons,…) par les radiations Ultraviolet (U.V.). Ce type de stérilisation a l'avantage de ne pas modifier la composition physico-chimique de l'eau traitée.
Contrôle de la qualité d'eau et entretien des ouvrages de traitement
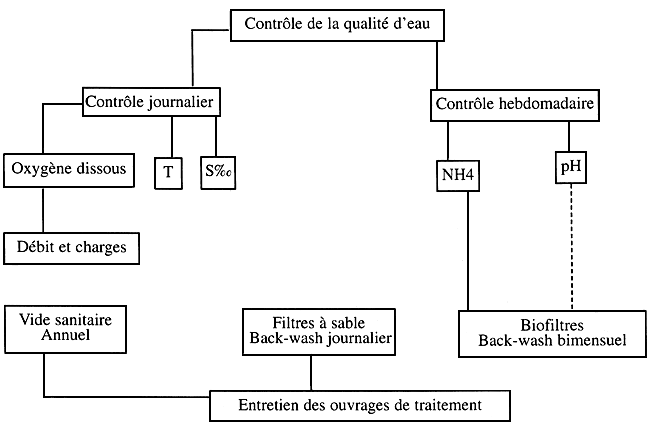
Schéma № 5
Contrôle de la qualité d'eau
Contrôle journalier: un suivi journalier est effectué sur les paramètres physico-chimiques les plus déterminants dans un élevage et connus sous le nom des paramètres de l'élevage. Ces paramètres sont : la température (T°), la salinité (S%0) et l'oxygène dissous.
- La température: la température optimale de croissance est fonction de l'âge et de l'espèce. Un bon choix de la fourchette de température optimale permet d'obtenir des alevins de bonne qualité (des lots homogènes, un box taux de vessie natatoire) et de meilleurs croissance. Pour le loup (Dicentrarchus labrax), la température d'élevage varie de 14–50° (à l'incubation des oeufs) à 21–30° (en fin d'élevage larvaire).
- La salinité (S%0): la salinité optimale de l'élevage est variable en fonction de l'espèce, de l'âge et des techniques d'élevage adoptées. La salinité de l'eau joue un rôle important, surtout aux premiers jours d'èlevage. Il est important de rappeler que les oeufs du loup deviennent benthiques en-dessous de 33%o, d'où la nécessité de maintenir une salinité supérieure à 33%o, pendant l'incubation des oeufs pour pouvoir séparer les oeufs viables (qui flottent en surface) des oeufs non viables et obtenir un bon taux d'éclosion.
- L'oxygène dissous: c'est un élément indispensable à a vie de toute les espèces aquatiques. Il assure la respiration et, par conséquent, l'activité métabolique.
Les besoins en oxygène des organismes aquatiques dépendent de l'espèce, de l'âge et du niveau métabolique (à jeun, en métabolisme de routine ou au moment de la digestion).
Dans une écloserie, en circuit de recyclage, l'apport en oxygène est assuré par les moyens de réoxygénation suivants:
l'apport d'eau de mer neuve (eau de renouvellement de 10 à 30 %,
l'oxygénation de l'eau de recyclage par l'installation de colonnes d'oxygénation et de dégazage à l'arrivée des bassins d'élevage:
les aérations installées dans les bassins d'élevage;
l'injection d'oxygène pur dans le circuit d'alimentation d'eau des bacs d'élevage larvaire (en cas de déficit en oxygène dissous).
Étant donné que la solubilité de l'oxygène dans l'eau est limitée, on est amené à connaître un élément plus significatif que la teneur en oxygène dissous l'eau , à savoir le taux de saturation
Dans le cas de l'élevage du loup, les besoins en oxygène dissous sont élevés (de 80 á 110% de saturation) pour les jeunes larves (dans les bacs d'élevage larvaire).
Dans les bassins nurseries, on peut tolérer des taux d'oxygène dissous plus bas (allant jusqu'à 50% de la saturation) sans à l'appétit des larves ni à leurs croissances.
La consommation moyenne en oxygène dissous des larves (pour le loup, elle est de 1200 à 1,000 mg/kg/h pour an poids moyen de 0.3 à 0.5 gr. et à une T°=22°C) ainsi que la teneur en oxygène dissous à l'arrivée des bassins, déterminent le débit d'eau nécessaire pour la survie et la bonne croissance des larves.
Les différents débits d'eau à détermine dans une écloserie sont:
Le débit de renouvellement: e'est la quantité d'eau mer (ou de mélange eau de mereau douce) nécessaire au renouvellement de l'eau de recyclage de 10) 30%, et ce, dans le but d'améliorer en partie la qualité de l'eau dans un circuit S.A.F. Le débit de renouvellement est fonction du débit de l'eau de recyclage.
Le débit de l'eau de recyclage: ce débit est fonction de la taille de l'écloserie, donc de sa capacité de production par cycle d'élevage.
Pour une meilleure gestion de l'eau, il est préférable d'avoir plusieurs pompes de recyclage de même débit pour les raisons suivantes:
- faire fonctionner une seule pompe pour de faibles besoins en eau (en débit de saison), et deux pompes où plus en pleine production. Par sécurité on peut garder une pompe en arrêt;
- le fait d'avoir le même type de pompe avec un même débit permet de minimiser le stock des pièces de rechange et de bien adapter le personnel aux réparations.
Pour protéger le circuit d'eau de recyclage des systèmes de sécurités, sont installés:
- un anti-bélier: c'est un ballon d'air sous pression qui permet d'absorber les chocs de pression (les coups de bélier) produits lors du démarrage des pompes, ou d'une fermeture brutale des vanne de réglage;
- un système de by-pass: installé sur presque tous les ouvrages de traitement (filtres à sable, biofiltres, stérilisateurs U.V. et échangeurs thermiques), ce système permet d'isoler ces ouvrages pour une intervention de nettoyage on de réparation, sans gêner à la circulation normale de l'eau.
Le débit d'alimentation en eau des bassins d'élevage: ce débit est calculé en fonction de la charge d'élevage et de l'âge des larves. le taux de renouvellement varie de 20% (à l'éclosion )à 300% (dans les bassins nurseries).
Pour le loup, les charges d'élevage préconisées sont de 100 larves de 1j/1 dans les bacs d'élevage larvaire et de 5 à 7kg/m3 dans les bassins nurseries.
Il est important de noter que l'amélioration de la qualité de l'eau de recyclage par une élévation thermique et par la réoxygénation (pour obtenir une meilleure croissance et des larves de meilleure qualité) peut conduire à une sursaturation de l'eau en gaz dissous.
Ce phénomène, en cas d'aggravation, peu conduire à des mortalités, d'où la nécessité d'installer les dispositifs de dégazage (les colonnes de dégazage, les cascades d'eau, l'écoulement gravitaire…).
Contrôle hebdomadaire : ce contrôle est effectué sur les paramètres indicateurs du rendement des biofiltres qui sont l'ammoniaque et le pH.
- L'Azote ammoniacale (NH3/NH4) :il est présent dans l'eau sous deux formes en
équilibre : NH3, H2O NH4, OH. La forme la plus toxique pour les poissons est
l'ammoniac (NH3); il provoque une gène respiratoire, un arrêt de croissance, et il peut
être mortel à de fortes doses, La toxicité de l'ammoniac commence à partir d'une
concentration d 0.05 mg/l. Étant donné que le dosage de l'azote ammoniacale par la
méthode colorimétrique détecte seulement l'ion ammonium NH3, il faut se référer à
un tableau ou à une courbe donnant le pourcentage de l'ammoniac (NH3), en fonction
du pH du milieu.
NH4, OH. La forme la plus toxique pour les poissons est
l'ammoniac (NH3); il provoque une gène respiratoire, un arrêt de croissance, et il peut
être mortel à de fortes doses, La toxicité de l'ammoniac commence à partir d'une
concentration d 0.05 mg/l. Étant donné que le dosage de l'azote ammoniacale par la
méthode colorimétrique détecte seulement l'ion ammonium NH3, il faut se référer à
un tableau ou à une courbe donnant le pourcentage de l'ammoniac (NH3), en fonction
du pH du milieu.
- Le pH: l'eau de mer possède un effet tampon sur le pH GRÂce à sa forte charge ionique. Par conséquent, la variation du pH de l'eau de mer en recyclage est fiable en cas de fonctionnement normal du biofiltre (pH=8). La variation du pH peut devenir importante par un dysfonctionnement des biofiltres (production importante d'ammoniaque et d'autres formes carbonées).
Les suivi de l'ammoniaque et du pH permet de contrôler le rendement des biofltres.
Entretien des ouvrages de traitement
Entretien journalier: les filtres à sables retiennent la plus grande partie de la m.e.s. Par conséquent, ils se colmatent vite, surtout en pleine production et au moment du sevrage Le colmatage des filtres à sable provoque un manque d'eau et peut être contrôler par des manomètres de pression installés à l'entrée et à la sortie des filtres à sables (ou tout simplement un seul manomètre installé entre le pompage et les filtres à sable). Pour assurer une bonne circulation d'eau à travers le sable des filtres, un back-wash est effectué une à deux fois/jour, Le back-wash consiste à faire circuler l'eau dans le sens inverse à celui de la filtration pour éliminer le dépôts solides de la m.e.s. retenue par le sable.
Entretien bimensuel: cet entretien est effectué sur les biofiltres qui, au bout d'un certain temps de fonctionnement accumulent une grande quantité de la m.e.s la plus fine. En plus, les supports bactériens des biofiltres (gravier, biolithe,…)peuvent s'agglutiner les uns contre les autres et former des caillots, ce qui réduit la surface de contact eau-bactéries et affecte par conséquent le rendement des biofiltres. Pour chasser la m.e.s. retenue par le biofiltre et pour empêcher la formation de caillots de biolithe ou de ravier, on procède à un back-wash avec de)'air sous pression deux fois/mois.
Entretien annuel: cet entretien est effectué sur toutes le installations et les circuits d'eau de l'écloserie à la fin de chaque saison larvaire. II consiste à faire les travaux d'entretien suivants:
- nettoyer les plaques de l'échangeur thermique à l'eau douce sous pression;
- nettoyer les stérilisateurs U.V. selon la description de la notice technique du modèle utilisé.
- faire le vide sanitaire de toute l'écloserie.
Le vide sanitaire est une mesure d'hygiène qui consiste à nettoyer et à désinfecter toute l'écloserie au début et à la fin de chaque saison larvaire.
Les avantages du circuit semi-fermé à fermé
Les avantages du circuit semi-fermé à fermé dans une écloserie sont:
assurer une régulation thermique de l'eau au moindre coût (une économie d'énergie calorifique);
éliminer la nécessité des grandes sources d'eau (une économie de l'énergie de pompage);
assurer une certaine autonomie de fonctionnement (isoler le circuit d'eau de l'écloserie);
réduire les risques de contamination par des sources d'eau polluée (réduire les problèmes de maladies dans les élevages);
améliorer la croissance des larves en travaillant dans les conditions optimales.
CONCLUSION
La gestion de l'eau dans une écloserie consiste à chercher un moyen terme entre l'obtention d'une eau de meilleure qualité et le coût du traitement appliqué, et ce dans le but de produire des alevins au moindre coût pour être compétitif sur le marché
Cependant, l'obtention d'une eau de meilleure qualité n'est pas seulement liée aux ouvrages de traitement installés, mais, également aux mesures d'hygiène prises dans la gestion des bassins d'élevage. En fait, un traitement d'eau, même poussé (Onéreux), peut ne pas être suffisant si les bassins d'élevage ne sont pas maintenus propres (siphonnage du reste d'aliment non ingéré, des feces, des cadavres de larves…)
Il est à signaler que les problèmes majeurs rencontrés dans un élevage larvaire (une fois que les techniques d'élevage sont maîtrisées) sont les problèmes pathologiques et les maladies de stress.
Par conséquent, une bonne gestion de l'eau peut, aide en grande partie à résoudre ces problèmes en assurant une eau de meilleure qualité physique, chimique et biologique.
RÉFÉRENCES BIBLIOGRAPHIQUES
• AYARI A., 1985 Application du contrôle en continu des paramètres physico-chimiques à l'étude de l'eutrophisation dans une lagune d'épuration. Thèse de troisième cycle - Montpellier-U.S.T.L. : 218 p
• JOHNSON, D.W. et Katavic, I, 1986 Survival and growth of se bass (Dicentrarchus labrax L.) larvea as influenced by temperature, salinity and delayed initial feeding. Aquaculture 52:11–19
• HAZEL, C.R., TIIOMPSON W. ET MEITS S. J., 1971 Sensitivity of striped bass and stickleback to ammonia in relation to temperature and salinity, Calif Fish Game 57:138–153
• HUSSENOT JET LECLERCQ D, 1987 La saturation des gaz dissous, un phénomène souvent mal connu en aquaculture Aqua Revue № 11 - Fev-Mars: 138–153
• LEMARIE G, GASSET E. CAM D. ET DELAFONCHAIS E., 1991 Modélisation de la consommation en oxygène du loup (Dicentrarchus labrux L.) et de la daurade (Sparus aurata L.) Recherche aquacole - Station IFREMER de Palavax, N/REF.GL/DL - 91/08/629 : 14 p.
• SUBB J, 1981 Manuel d'initiation aux méthodes de mesure des paramètres physiques et chimiques dans l'eaux de mer. I.U.T.(CREUTOP) U.S.T.L. - Station de biologie marine et lagunaire de Sète : 51 P.
By Dr. Patrick LAVENS
and Mr. Patrick SORGELOOS
GENT - BELGIUM
INTRODUCTION
Dependable availability of fry, fingerlings or post-larvae is one, if not the single most critical factor in the commercial success of the industrial production of marine fish or crustaceans (FAO, 1990). For only a few species with unique ecological and ethological characteristics, wild sources of seed can be used for the stocking of grow out ponds or cages : e.g.
in Ecuador Penaeus vannamei and P. stylirostris post-larvae are being coastal areas by the billions per year, and in fact this cheap source of readily available seed has been at the origin of the present multi-million US dollar shrimp farming industry in Ecuador (O'Lee, 1989);
wild fry is the single source of seed for annual productions of several hundred thousand tons each of the milk fish Chanos chanos in the Philippines and lndonesia, and the yellowtail Seriola quinqueradiata in Japan.
When a number of years ago in Ecuador, wild penaeid seed provisions suddenly decreased, extended pond areas could not be kept in operation (Spurrier, 1988). Milk fish and penaeid shrimp farming in the Philippines are often restricted because of seasonal fry shortages. The Japanese government does restrict yellowtail cage farming to prevent over fishing of wild fry resources.
When applying intensi9ve grow out techniques, shrimp farms need to operate at maximum capacities in order to show good profit. This involves guaranteed provisions of shrimp postlarvae on a year round basis.
In recent years different species of marine fish (e.g. several bass and bream species, dolphin fish mahimahi, turbot, halibut, etc…) have been identified as new aquaculture candidates, i.e. their growout in cages or land-based systems have proven to be commercially attractive provided a regular supply of fry can be guaranteed. For most fish and crustaceans with aquaculture potential this can only be realized by the domestication of the species, i.e. involving the development of appropriate techniques for controlled maturation and larviculture.
Larviculture nutrition, more particularly start-feeding in the early larval stages, appears to be the major bottle-neck for the industrial upscaling of the culture of fish and shellfish. With a few selected species such as salmon, minimal problems had to be overcome and since a few years already, billions of hatchery produced fry are used in the commercial production of several 100,000 tons of marketable salmon per year. Early development in salmon does not involve feeding problems as the babies at hatching carry a big yolk sac with enough food reserves for the first three weeks of their development. Once yolk is consumed and exogenous feeding can start, larvae already have a large mouth and can thrive on formulated feeds. With most other marine fish, egg production might not pose the major problem but larval size at hatching and first feeding is at the origin of farming difficulties (sec table 1).
Bass, bream, turbot, mahimahi and many other marine fish with aquaculture potential, all have very limited yolk reserves at hatching mostly lasting for no more than one or two days. At first feeding they still have small mouths, often with an opening size of less than 0.1 mm.
In shrimp, the size of the larval food is not the only problem, larvae furthermore pass through different larval stages eventually changing from a herbivorous filter-feeding behavior to become a carnivorous hunter.
Table 1 - Size of eggs and larval length at hatching in different species of fish
(after Jones and Houde 1981)
| Species | Egg diameter (mm) | Length of larvae (mm) |
| Salmon (Salmo salar) | 5.0 – 6.0 | 15.0 – 25.0 |
| Trout (Salmo gairdneri) | 4.0 | 12.0 – 20.0 |
| Carp (Cyprinus carpio) | 0.9 – 1.6 | 4.8 – 6.2 |
| Bass (Dicentrarchus labrax) | 1.2 – 1.4 | 7.0 – 8.0 |
| Bream (Sparus aurata) | 0.9 – 1.1 | 3.5 – 4.0 |
| Turbot (Scophthalmus maximus) | 0.9 – 1.2 | 2.7 – 3.0 |
| Sole (Solea solea) | 1.0 – 1.4 | 3.2 – 3.7 |
| Milk fish (Chanos chanos) | 1.1 – 1.25 | 3.2 – 3.4 |
| Grey mullet (Mugil cephalus) | 0.9 – 1.0 | 1.4 – 2.4 |
| Grouper Epinephelus tauvina) | 0.77 – 0.90 | 1.4 – 2.4 |
| Bream (Acanthopagrus cuvieri) | 0.78 – 0.84 | 1.8 – 2.0 |
The natural diet of most aquaculture fish and crustacean species consists of wide diversity of phytoplankton species (diatoms, flagellates, etc…) and zooplankton organisms (copepods, cladocerans, decapod larvae, etc…), found in great abundance in the plankton. This abundance and maximal diversity of food organisms of different sizes and biochemical composition provide maximal chances for meeting all the nutritional requirements of the shrimp larvae.
The first culture trials on a laboratory scale were carried out feeding freshly-collected wild plankton, aiming at imitating the natural habitat, free of predators. Collecting and feeding natural plankton may appear evident but hardly dependable beyond laboratory scale. On an industrial scale, similarly to intensive cattle and poultry farming, a readily and consistently available, practical and performing diet is to be selected.
Selection of a suitable and nutritious diet may be carried out according to number of criteria selected (see Fig. 1)
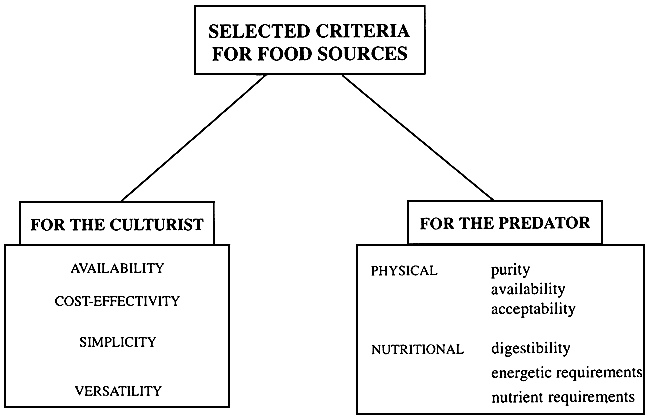
Figure 1 - Selection criteria for larval food sources front the viewpoint of the culturist and the cultured larvae (from Léger et al., 1987a)
From the practical viewpoint of the culturist, a good diet should be readily available, cost-effective, simple as well as versatile in application.
The consistent availability of sufficient numbers of food organisms is of utmost importance for continuous cultures. In this respect, the collection and feeding of wild plankton has proven unreliable and not practical. Over the past two to three decades, trial and error approaches have resulted in the adoption of selected larviculture diets, taking into account the different criteria listed in Figure 1. Today three groups of live diets are widely applied in industrial larviculture of marine fish and crustaceans:
different species of 2 to 20 μm large micro-algae,
the 50 to 200 μm rotifer Brachionus plicatilis, and
the 200 to 500 μm brine shrimp Artemia.
In recent year different formulations of supplementation and substitution product have been added to this list.
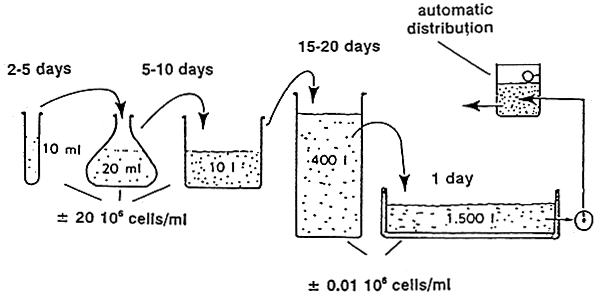
Figure 2 - Schematic outline of steps involved in the mass production of micro-algae
MICRO ALGAE
Diatoms and green algae are the two dominant groups of cultured micro-algae (De Pauw and Pruder, 1986). Food species have been selected on the basis of their mass culture potential, cell size, digestibility, and overall food value, much more by trial and error than any other scientific selection process. Easy to culture «weeds», such as Chlorella sp. appear to have poor food value (i.d. poor digestibility) for many species with aquaculture potential. The most suitable species still pose many problems for large scale culture, i.e. their culture implies the use of complex culture media, made up with germ-free seawater treated by microfiltration and/or U.V.-irradiation.
Algae are progressively cultured in a multi-stage back-up system (Figure 2). Starting from sterile test-tubes the algae are inoculated in larger glass containers and carboys, then to bigger vessels or plastic bags and finally to large, indoor or outdoor tanks. As a result most farms still apply labor intensive and expensive batch production systems. Even when production targets can be maintained with regard to cell numbers produced, shrimp farmers for example have example have experienced temporal variations in algal food values resulting in inconsistent hatchery outputs (Chamorro, pers. comm.). Olsen (1989) reviewed the literature and illustrated that the content of the (n-3) highly unsaturated fatty acids (HUFA's) eicosapentaenoic acid (EPA) 20:5(n-3) and docosahexaenois acid (DHA)22:6(n-3) can greatly vary among algal species but even from culture to culture within a given species (Figure 3). Using Penaeus stylirostris as a test organism, Léger et al (1985a) could demonstrate that the content of EPA and DIIA in the zoea diet had a major impact on survival and growth in later stages, when animals had already been switched to another diet (Figure 4, black bars only). This provided the rationale to look for alternatives or supplements to live micro-algae. Today different approaches and formulations are being applied at commercial level already, and many new developments in producing more cost-effective products are to be expected: i.e. freeze-dried algae (Taylor, 1990) manipulated yeast's (Léger et al., 1987b, Léger and Sorgeloos, 1990-Coutteau et al., 1990 - see Figure 4), micro-encapsulated feeds (Jones et al., 1979), and different kinds of microparticulate diets (Langdon and Waldock, 1981 - Kanazawa et al., 1982 - Chamberlain, 1988 - Sorgeloos and Van Stappen, 1991) are gradually reducing the need for and might eventually totally replace the micro-algae in commercial larviculture in the near future.
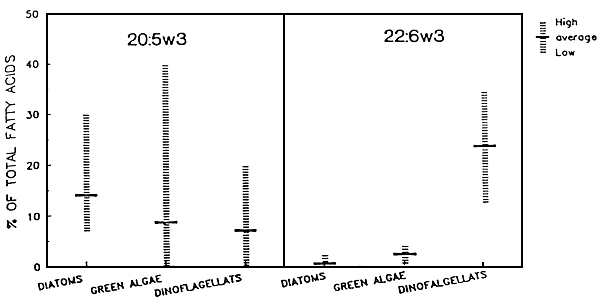
Figure 3 - Content of the essential fatty acids 20:5(n-3) and 22:6(n-3) in marine diatoms, green algae and dinoflagellates (from Oslen, 1989)
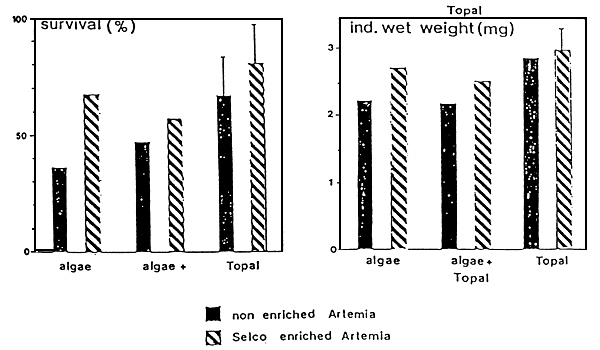
Figure 4 - Larviculture outputs with Penaeus vannamel reared up to the PL-8 stage in 200 l tanks on diets consisting of only algae (mixture of Chaetoceros and Tetraselmis; HUFA-composition : 3–9 % 20:5n-3 and 0– l % 22:6n-3; (first two columns», only Topal (HUFA-composition : 13.2 % 20:5n-3 and 8.7% 22:6n-3; last two columns) or a mixture of both (columns in Middle) up to the M-2 stage; each treatment was, split up as of the M-3 stage in a group fed only freshly hatched Artemia (HUFA-composition: 5.6% 20:5n-3; no 22:6n-3: dark bars), and a group receiving 12-hr SELCO-enriched Artemia (6.4% 20:5n-3 and 3.3% 22:6n-3) in M-3 and PL-l stage, followed by 24-h SELCO enriched Artemia (21.3% 20:5n-3 and 12.7% 22:6n-3) in the later PL-stages; striped bars (modified after Léger et al., 1987b)
THE ROTIFFER BRACHIONUS PLICATILIS
This brackish water zooplankton (Figure.5) was identified by Japanese aquaculturists as a suitable starter diet in marine fish larviculture. (Fukusho, 1989). In function of the larval mouth size of the cultured fish, a small Brachioun strain (50-10 μm length) or a large one (100–200 μm) is used.
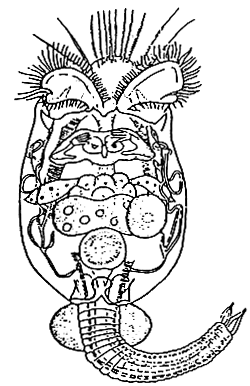
Figure 5 - Microscopic drawing of Brachionus plicatilis (from Walker, 1981)
Under optimal culture conditions, Brachionus plicatilis reproduces by parthenogenesis, i.e. each female produces several eggs at a time which upon hatching reach reproductive stage in a few days only. The widely-applied batch culture procedure appears to be simple, using micro-algae often Chlorella and/or bakers' yeast or other formulated products as food source. Cylindro-conical tanks are inoculated at a start density of approximately 200 animals per liter (20 ppt seawater). Homogeneous mixing and oxygenation of the water is performed by using airstones. When feeding yeast or other formulated feeds, airstones should be placed at a certain distance above the tank's bottom as to allow sedimentation of waste particles that can be siphoned off once per day.
Food should be distributed over several rations per day. Best results are obtained when applying continuous pumping of concentrated food from a refrigerated stock suspension via a centrifugal pump into the culture tanks. Daily added food quantities start at about 55 mg dry matter (example of yeast-based product : CULTURE SELCO, ARTEMIA SYSTMS NV/SA, Gent, Belgium) per million rotifers and are progressively reduced by 5 mg per day. Cultures are harvested on day 5 when the rotifer density has reached a maximum of about 450 animals per ml (Léger et al., 1991).
Many fish hatcheries have experienced significant problems in maintaining large cultures and producing on a predictable basis the massive numbers of rotifers that are needed to feed the hundred thousands to millions of baby fish they have in culture (different commercial fish hatcheries in the Mediterranean, pers. comm.). The rotifers' food appeared to be one of the key elements in the successful mess production of rotifers; i.e. for convenience fresh bakers' yeast is mostly used as the main diet ingredient. However, its freshness, a criterion which is difficult to evaluate by the farmer, can greatly influence the dietary value of the yeast for the rotifers, and as a consequence determine rotifer culture success. Many farmers supplement the baker's yeast with micro-algae, a procedure which at the same time ensures an increase of the level of (n-3) essential fatty acids in the rotifers. This (n-3) HUFA enrichment is critical in optimizing the food value of the rotifers for marine fish larvae (Fukusho, 1989, see further).
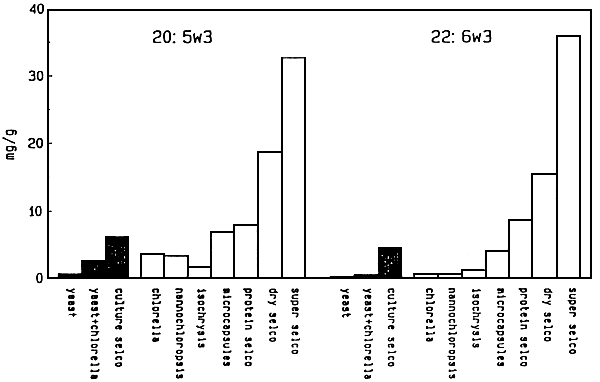
Figure 6 - Levels of EPA and DHA in rotifers cultured (black bars) or enriched (open bars0 with different micro-algae, bakers' yeast, microcapsules and different formulated SELCO-products (Artemia Systems NV/SA, Gent, Belgium) (from Komis et al., 1989)
Different microparticulate (Rimmer and Reed, 1990) and emulsified formulations (Watanabe et al., 1989 - Léger et al., 1987c - Léger et al., 1989) are used for this boosting with essential fatty acids and other components. The (n-3) HUFA enrichment of rotifers is performed either in the culture vessel or mostly in a separate tank. The enrichment diets are fed during 6 to 8 hours at a dose of 100 to 250 ppm, in two rations, depending on the type of product. Rotifer density is about 200 to 500 per ml and temperature is kept between 20 to 25° C. Maximal (n-3) HUFA levels that can be reached today in 6 hours enriched rotifers amount to 77 mg ∑(n-3)HUFA level of 5.8 mg/g respectively 8.3% (Léger et al., 1989). EPA and DHA levels in rotifers cultured or enriched on different diets are given in Figure 6.
THE BRINE SHRIMP ARTEMIA SPP
Of the live diets used in larviculture, brine shrimp Artemia nauplii constitute the most widely used species. Technically speaking the advantages of using Artemia is that one can produce live food «on demand» from a dry and storable powder, i.e. dormant Artemia cysts (embryos) which upon immersion in seawater regain their metabolic activity and within 24 hours release free-swimming larvae (nauplii) of about 0.4 mm length (Figure 7). Actually more than 500 metric tons of dry Artemia cysts are marketed annually for worldwide production of freshly hatched Artemia nauplii to be used as food in the hatchery phase of fish and crustacean aquaculture.

Figure 7 - Early stage of hatching of Artemia cyst (pre-nauplius surrounded by the hatching membrane still attached to the cyst shell of chorion) and free-swimming nauplius (instar 1 stage) (from Léger et al., 1986)
Although hatching of Artemia cysts appears to be simple, several parameters need to be taken into consideration when hatching out kilogram quantities of cysts as needed on a daily basis in the large fish/shrimp hatcheries, i.e. a constant temperature between 25 and 28°C, salinity of 15 to 35 ppt, pH Z 8.0 (in order to increase the buffer capacity of the hatching medium about 1 g NaHCO3/1 should be added), oxygen levels close to saturation (best hatching results can be achieved with funne Ishaped containers that are aerated from their bottom), cysts densities of not more than 2g/l, and a strong illumination of the cysts (about 2,000 lux at the water surface) is essential, at least during the first hours following their complete hydration, in order to trigger the start of their hatching metabolism (Sorgeloos et al., 1986). These factors will all affect the hatching rate and maximum output, and hence the production cost of the harvested Artemia nauplii. Attention should be paid to select Artemia cyst-lots with a good hatching synchrony and a high hatching efficiency (i.e. the number of nauplii that can be produced under standard conditions from one gram cyst product), as considerable variation has been demonstrated between cysts of various origin, and even between batches from the same strain (Sorgeloos et al., 1986).
After hatching, and prior to feeding them to the fish/shrimp larvae, Artemia nauplii should be separated from the hatching wastes (empty cyst shells, bacteria and hatching metabolities). When switching of the aeration in the hatching tank, cyst shells will float and nauplii will concentrate at the tank's bottom from where they should be siphoned off within 5 to 10 minutes. Nauplii should be thoroughly rinsed with freshwater, preferentially in submerged filters as to prevent physical damages of the nauplii.
Considerable progress has been made in the past decade in improving and increasing the value of Artemia as a larval diet. A better knowledge of the biology of Artemia was at the origin of the development of methods for cyst disinfection and decapsulation (Sorgeloos et al., 1986 - Léger and Sorgeloos, 1991). Artemia cyst shells indeed may be loaded with germs of bacteria and fungi. These may be removed by disinfection using leach. A minimum treatment consists of a 30 minute soak of the cysts in a 200 ppm hypochlorite solution (fresh or seawater) prior to incubation for hatching. This treatment however may not kill germs present in the alveolar and cortical layer of the chorion. A more effective treatment consists in the decapsulation of the cysts, i.e. the complete removal of the chorion by short exposure of the hydrated cysts to a 2-% hypochlorite solution at high pH. Provided the different steps in the decapsulation treatment (cyst hydration, exposure to hypochlorite solution, washing, chlorine deactivation in hydrochloric acid, washing and/or brine dehydration) are performed following recommendations (Sorgeloos et al., 1986) sterile embryos are produce that have not lost their viability. These procedures are being applied at most large fish and shrimp hatcheries to sterilize the cysts and at the same time to reduce the problems of separating the empty cyst shells at naupliar harvest.
For a long time farmers have overlooked the fact that an Artemia nauplius in it a first stage of development can not take up food and thus consumes its own energy reserves (Benijts et al., 1976 - see Figure 8). At the high water temperatures which are applied for cyst incubation, the freshly hatched Artemia nauplii develop into the second larval stage within a matter of hours. It is important to feed first-instar nauplii rather than starved second-instar metanauplii which are less visible as they transparent. Instar II Artemia are also larger and swim faster than first instars, as a result of which they are less acceptable as a prey. Furthermore they contain lower amounts of free amino acids so they are less digestible and their lower individual organic dry weight and energy content will reduce the energy uptake by the predator per hunting effort. All this will be reflected in a reduced growth of the larvae (Figure 9), for an increased Artemia cyst bill (as about 20 to 30 % more cysts need to be hatched to feed the same weight of starved metanauplii to the predator). Molting of the Artemia nauplii to the second instar stage may be avoided and their energy metabolism greatly reduced (see Figure 8) by storage of the freshly hatched nauplii at a temperature below 10°C in densities of up to 8 million nauplii per litre, for periods up to 24 h (Léger et al., 1983). A slight aeration is needed in order to prevent the nauplii of accumulating at the bottom of the tank where they would suffocate. Applying 24 h cold storage (using styrofoam insulated tanks and blue ice packs for cooling) commercial hatcheries are able to economize their Artemia cyst hatching efforts (e.g. fewer tanks, bigger volumes, maximum one hatching and harvest per day). Cold storage furthermore allows the farmer not only to ensure the availability of a better quality product but the same time to consider more frequent food distributions. This appeared to be very beneficial for fish and shrimp larvae as food retention times in the larviculture tanks can be reduced and hence growth of the Artemia in the culture tank can be minimized; e.g. applying one or maximum two feeding per day, shrimp farmers often experienced onfsgrowth of the Artemia in their larviculture tanks. With poor hunters such as the larvae of turbot Scophthamus maximus and tiger shrimp Penaeus monodon, feeding cold-stored Artemia results in much more efficient food uptake.
Figure 8 - Change in energy content and dry weight of different forms of Artemia (newly-hatched instar I nauplii are considered to have 100% values for those variables). The present decrease or increase from 100% is shown for consequently instar II-III metanauplii, cold-stored inst-I nauplii, and decapsulated cysts (from Léger et al., 1987)
Figure 9 - Effects of feeding starved instar II-III Artemia nauplii to fish of crustacean larvae (from Léger and Sorgeloos, 1991)
Easy hatching and disinfection procedures, however, are not the sole parameters in ensuring the success of using Artemia as a larval food source. Several other Artemia characteristics can influence the suitability of a particular brine shrimp product for one or another type of larviculture. One of these is nauplius size which can greatly very from one geographical source of Artemia to another. This is especially critical with several species of marine fish that have a very small mouth size and swallow their prey in one bite; i. e. using the marine silverside Menidia menidia as a test-organism Beck and Bengtson (1982) are able to illustrate a high correlation between Artemia nauplii size (eight Artemia strains varying in size from 440 to 520 ωm in nauplius length) and larval mortality during early development; with the largest strains of Artemia up to 50% of the fish could not ingest their prey and starved to death (Figure 10).
Figure 10 - Correlation of mortality rate of Menidia menidia larvae and naupliar length of Artemia from eight different geographical sources offered as food to the fish larvae (from Beck and Bengtson, 1982)
Another important dietary characteristic of Artemia nauplii was identified in the late '70's, early '80's when many fish and shrimp farmers started to go commercial and reported unexpected problems when switching from one source of Artemia to another.

Figure 11 - Survival of Mysidopsis bahia larvae fed freshly hatched Artemia nauplii from different geographical sources (left) and from different collections from the San Francisco Buy (S.F.B., California, USA; (right) : results expressed as percentage of the control treatment fed with freshly hatched Reference Artemia Cysts (RAC) (from Sorgemoos and Léger, 1991)
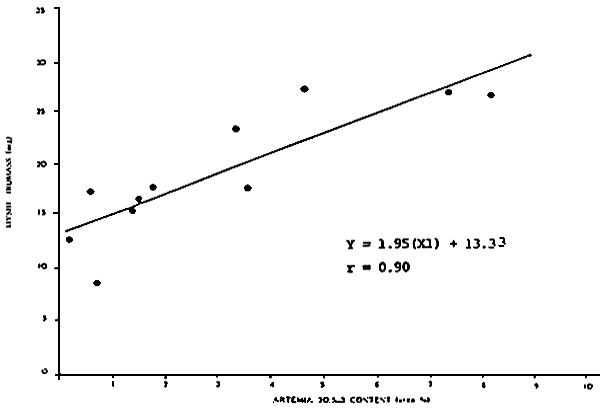
Figure 12 - Linear relationship between the 20:5 (n - 3) content Artemia collections from San Francisco Bay origin and the biomass of Mysidopsis bahia to which the freshly hatched Artemia were fed (from Sorgeloos and Léger, 1991)
Table 2 - Intra-strain variability of 20;5 (n-3) content in Artemia. Data represent the range (ares percent) and coefficient of variation of data as complied by Léger et al. (1986 and 1987a)
| Artemia geographical strain | 20:5(n-3) range (area %) | Coefficient of variation (%) |
| USA-California : San Francisco Bay | 0.3 – 13.3 | 78.6 |
| USA-Utah Great Salt Lake (S arm) | 2.7 – 3.6 | 11.8 |
| USA-Utah Great Salt Lake (N arm) | 0.3 – 0.4 | 21.2 |
| Canada-Chaplin Lake | 5.2 – 9.5 | 18.3 |
| Brazil-Macau | 3.5 – 10.6 | 43.2 |
| PR China-Tientsin | 1.3 – 15.4 | 50.5 |
(Sorgeloos and Persoone, 1980), Japanese, American and European researchers studied these problems and soon confirmed variations in nutritional value when using different geographical sources of Artemia for fish and shrimp species (see reviews in Watanabe et al., 1983 and Léger et al., 1986). The situation became more critical when very significant differences in production yields were obtained with distinct batches of the same geographical origin of Artemia (Figure 11) Multidisciplinary studies in Japan and by the International Study on Artemia (Léger et al., 1987) revealed that the concentration of the essential fatty acid FPA in the Artemia nauplii is determining the nutritional value of this particular batch of Artemia for the larvae of various marine fish and crustacean species. Figure 12 illustrates the results obtained with different batches of the same geographical Artemia source, containing very different amounts of EPA and yield proportional results in growth and survival of Mysidopsis bahia shrimps fed these Artemia
As can be seen in Table 2, EPA levels in Artemia can greatly vary, even from one batch to another within the same strain. Cyst products from inland resources appear to be more constant in composition, be it however at suboptimal low levels. As a result concentrations of the (n-3) HUFA EPA need to be taken into consideration when selecting the most appropriate batch or Artemia cysts. Commercial provisions of Artemia cysts containing high EPA-levels are limited. Their use, however, should be restricted to the feeding period when size of the prey is most critical. Indeed even the best natural Artemia products do not meet all the nutritional requirements of the predator larvae, most particularly with regard to the other essential fatty acid for marine organisms, namely DHA 22;6(n-3), which is never available in significant amounts in Artemia cysts (Léger et al., 1986).
It is fortunate that Artemia because of its primitive characteristics facilitates a very convenient way to manipulate its biochemical composition. Since Artemia is non-selective in taking up particulate matter once it has molted into the second larval stage, i.e. about 8 hours following hatching, simple methods could be developed to incorporate any kind of product into the Artemia prior to offering it as a prey to the predator larva, this method of bioencapsulation, also called Artemia enrichment of boosting, is widely applied at marine fish and crustacean hatcheries all over the world for enhancing the nutritional value of Artemia with essential fatty acids.
British, Japanese and Belgian researchers developed enrichment products and procedures using selected micro algae and/or micro-encapsulated products, yeast and/or emulsified preparations, respectively self-emulsifying concentrates and/or micro-particulate products (see review of Léger et al., 1986). Highest enrichment levels are obtained when using emulsifies concentrates (Figure 13): freshly hatched nauplii are transferred to the enrichment tank at a density of 100 (for enrichment periods of > 24 hours) to 300 nauplii/ml (max. 24 hr enrichment period). The enrichment medium consists of disinfected seawater that is maintained at 25°C. The enrichment emulsion is added at consecutive doses of 300 ppm every 12 hours. A strong aeration using airstones is required to maintain dissolved oxygen levels above 4 ppm. Enriched nauplii are harvested after 24 or 48 hours, thoroughly rinsed and stored at temperatures below 10/degree/c in order to assure that HUFA's not metabolized during storage (see Figure 14).
PROGRESS IN LARVICULTURE NUTRITION
The use of (n-3) HUFA-enriched Artemia as a more adequate food source has without any doubt been at the origin of a real breakthrough in the larviculture of many marine fish species (Watanabe et al., 1983 - Sorgeloos et al., 1987 and 1988), e.g. for the European bass Dicentrarchus labrax and seabream Sparus aurata, the adoption of this bio-encapsulation methodology has allowed the transition from pilot to commercial larviculture of these species (Frenzos and Swaatman, 1989). The effects of feeding (n-3) HUFA-enriched Artemia and Brachionus indeed are significant; e.g. Dicentrarchus larvae die off before day 35 when fed (n-3) HUFA-deficient freshly-hatched Artemia nauplii from Great Salt Lake cysts (Van Ballaer et al., 1985 - Franicevic et al., 1987); switching to enriched Artemia of the same batch resulted not only in increased survival but also in the production of bigger larvae which better resisted stress conditions.

Figure 13 - HUFA-levels in Great Salt Lake (UT-USA) Artemia (meta) nauplii enriched with the self-emulsifying concentrate SUPER SELCO (Artemia Systems NV/SA, Gent, Belgium) (from Léger et al., 1991)
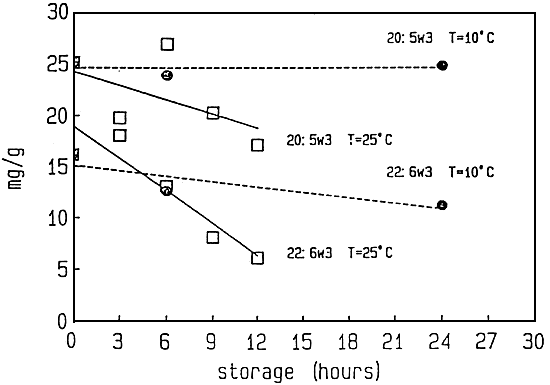
Figure 14 - HUFA-levels in SUPER SELCO-enriched Artemia metanauplii during storage at 10 and 25°C (from Léger et al., 1991)
Similar observation of increased survival and growth when feeding (n-3) HUFA-enriched diets have been confirmed for several species of penaeid shrimp were sometimes effects of diet composition only come to expression in later stages (Léger at al., 1987b - Léger and Sorgeloos, 1990 - see Figure 4, striped bars). A good illustration of this is the resistance to salinity stress in PL-10 stages of a batch of Penaeus monodon larvae fed on three different larval diets that varies in (n-3)HUFA levels (Tackaert et al., 1989). Differences in survival between the three diets were not significant at PL-10 before the stress test. However, differences in PL-quality (expressed as their ability to survive the salinity stress applied) were very pronounced (Figure 15) This criterion of resistance to salinity shocks which can easily be applied at the hatchery level is now being used as a quality criterion for determining the appropriate time for PL-transfer from the hatchery to the ponds (Sorgeloos, 1989).
With the freshwater prawn Macrobrachium rosenbergii (n3)HuFA-requirements of the larvae were anticipated not to be very critical in view of the fact that they sped most of their life in freshwater. These assumptions, however, were largely contradicted by Devresse et al. (1990) and Romdhane et al. (1991) who used Artemia enriched with different (n-3)-HUFA emulsions. Besides the improved growth rate, a distinct difference having an important impact for the commercial farmer is the more precocious and more synchronous metamorphosis as well as the higher stress resistance of these Macrobrachium postlarvae that had received (n-3) HUFA-enriche Artemia in the larval stages (Figure 16).
Whereas for a number of species larviculture outputs have been improved thanks to these developments in nutritional manipulation of the live feeds (various papers presented at the Larviculture Special Session of Aquaculture 1990, Halifax, Canada), with other species such as mahimahi, turbot, halibut and others, research is still in progress to better define qualitative and quantitative (n-3) HUFA requirements; it appears that for many species of marine fish optimal dietary levels have not been reached yet in Brachionus and Artemia. Furthermore (n-3) HUFA's might have proven most critical, other nutrients, e.g. other lipid classes, particular peptides, free amino acids, pigments, sterols and vitamins might appear equally important and in some species may be more critical. In view of the better results obtained when using natural plankton, for example consisting of marine copepods in culturing turbot and mahimahi (Stottrup, respectively Kraul, pers. communications), the challenge remains to identify these vital components in the copepods in order to have these incorporated in the convenient dietary system consisting of Brachionus and Artemia.
FORMULATED SUBSTITUTION PRODUCTS
Off the shelf dry products (Brachionus and Artemia substitutes) are being developed and commercialized as a much more user friendly application for the farmer (Sorgeloos and Léger, 1991). With some species such as penaeid shrimp the moment is approaching of reducing live algae and Artemia feeding to virtual elimination from the hatchery operation. Other species such as marine fish species impose much more constraints than shrimp not only in terms of nutritional requirements and digestibility but also with regard to physical properties of the feed, e.g. water stability, buoyancy, palatability, etc… Appropriate process technologies will certainly be developed but their commercial applicability and price competitiveness might not be obvious.
Feeding strategies in hatcheries of marine fish and crustaceans will probably never turn into worldwidely standardized blueprint methods because of species differences and geographical discrepancies. The cost-effectiveness of live and formulated feeds will dictate their proportional use (Sorgeloos and Léger, 1991).
ACKNOWLEDGMENTS
This research has been sponsored by the Belgian National Science Foundation, the Belgian Administration for Development Cooperation, the Belgian Ministry for Science Policy, and the Belgian Company ARTEMIA SYSTEMS NV/SA.
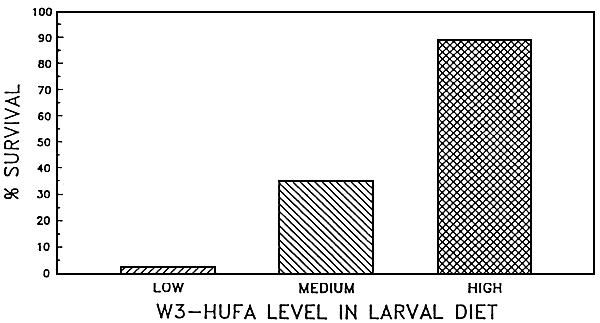
Figure 15 - Survival of Penaeus monodon PL-10 cultured on larval diet combinations that contained low, medium and high levels of (n-3) HUFA's after 60 min, transfer from 35 ppt to 7 ppt seawater (from Sorgeloos and Léger, 1991)

Figure 16 - Results of a 28-day culture test with Macrobrachium rosenbergii larvae fed Artemia nauplii enriched with low, medium and high (n-3)HUFA diets (from Romdhane et al., 1991)
![]()
![]()
![]()