![]()
![]()
![]()
Digestión, absorción y transporte
La mayor parte de las grasas alimentarias se suministran en forma de triacilglicéridos, que se deben hidrolizar para dar ácidos grasos y monoacilglicéridos antes de ser absorbidos. En niños y en adultos, la digestión de las grasas se produce de forma eficaz y casi completa en el intestino delgado. En los recién nacidos, la secreción pancreática de lipasas es baja. En los bebés, la digestión de las grasas mejora gracias a las lipasas segregadas por las glándulas de la lengua (lipasa de la lengua) y una lipasa presente en la leche materna. El estómago interviene en el proceso de digestión de las grasas debido a su acción agitadora, que ayuda a crear emulsiones. Las grasas que entran en el intestino se mezclan con la bilis y posteriormente se emulsionan. La emulsión es entonces tratada por las lipasas segregadas por el páncreas. La lipasa pancreática cataliza la hidrólisis de los ácidos grasos de las posiciones 1 y 3, generando 2-monoacilglicéridos (Tso, 1985). Los fosfolípidos son hidrolizados por la fosfolipasa A2, y los principales productos son lisofosfolípidos y ácidos grasos libres (Borgstrom, 1974). Los ésteres del colesterol son hidrolizados por la hidrolasa de ésteres de colesterol pancreática.
Los ácidos grasos libres y los monoglicéridos son absorbidos por los enterocitos de la pared intestinal. En general, los ácidos grasos con longitudes de cadena inferiores a 14 átomos de carbono entran directamente en el sistema de la vena porta y son transportados hacia el hígado. Los ácidos grasos con 14 o más átomos de carbono se vuelven a esterificar dentro del enterocito y entran en circulación a través de la ruta linfática en forma de quilomicrones. Sin embargo, la ruta de la vena porta también ha sido descrita como una ruta de absorción de los ácidos grasos de cadena larga (McDonald et al., 1980). Las vitaminas liposolubles (vitaminas A, D, E y K) y el colesterol son liberados directamente en el hígado como una parte de los restos de los quilomicrones.
Las enfermedades que perjudican a la secreción biliar, como la obstrucción biliar o los trastornos de hígado, conducen a graves deficiencias en la absorción de las grasas, como también sucede con las enfermedades que afectan a la secreción pancreática de las enzimas con actividad de lipasa, como la fibrosis cística. Como resultado, los triglicéridos con longitudes de cadena medias pueden tolerarse mejor en las personas que presentan una absorción deficiente de las grasas, y frecuentemente se utilizan como fuente de energía en la alimentación. La absorción intestinal completa de los lípidos puede verse afectada marginalmente por cantidades elevadas de fibra en la dieta. La absorción de las grasas se ilustra en la Figura 3.1.
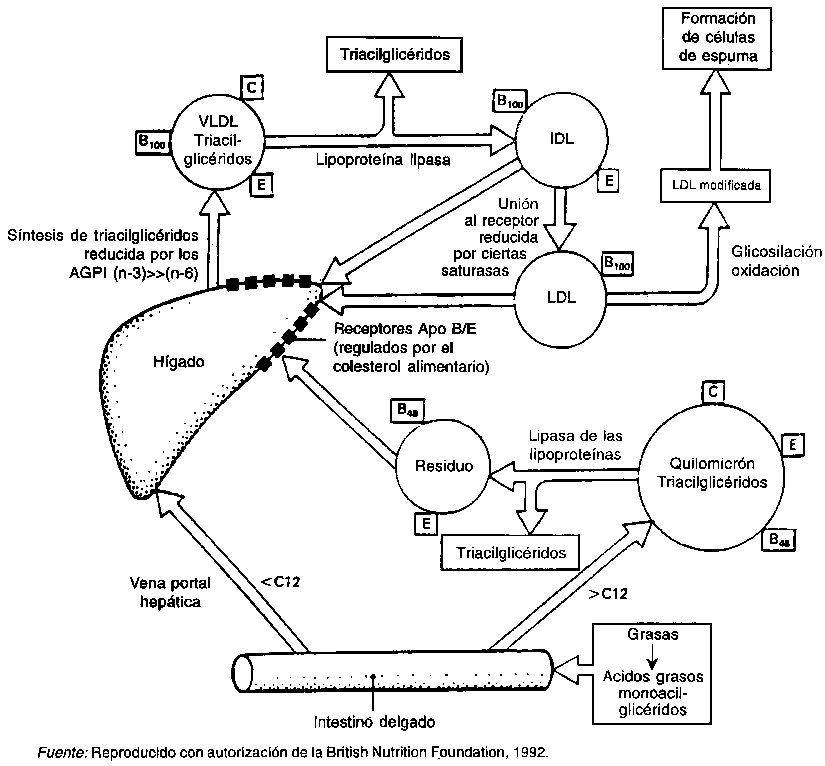
FIGURA 3.1 - Influencia de las grasas alimentarias en el metabolismo de las lipoproteínas
Los ácidos grasos son transportados en la sangre como complejos de albúmina o como lípidos esterificados en las lipoproteínas. Estas consisten en un núcleo de triacilglicéridos y ésteres ácidos grasos de colesterol, y un revestimiento formado por un estrato de fosfolípidos en el que se encuentran esparcidas moléculas de colesterol sin esterificar. Las cadenas plegadas de una o más apolipoproteínas se extienden por encima de la superficie y, con los fosfolípidos anfipáticos, permiten que los lípidos del núcleo sean transportados por la sangre. También regulan la reacción del conjunto lipídico con enzimas específicas, o unen las partículas a los receptores superficiales de las células.
Los quilomicrones son partículas lipoproteicas que proceden de las grasas alimentarias y son empaquetadas por las células de la mucosa. Entran en el torrente sanguíneo a través de los vasos linfáticos. La lipasa de lipoproteínas, que se encuentra en la pared interior de los capilares sanguíneos, hidroliza los triglicéridos, liberando ácidos grasos. Estos entran en el tejido adiposo, donde se almacenan, y en los músculos, donde se utilizan como combustible. Los restos de los quilomicrones son depurados por el hígado durante las primeras horas que suceden a la ingestión de una comida que contiene grasas.
Las lipoproteínas de muy baja densidad (VLDL) son partículas de gran tamaño ricas en triacilglicéridos que se producen en el hígado a partir de la grasa endógena, a diferencia de los quilomicrones, que transportan grasa exógena. Las VLDL son los principales portadores de triacilglicéridos que también son elaborados por la lipasa de lipoproteínas y proporcionan ácidos grasos a los tejidos adiposo y muscular.
Las lipoproteínas de baja densidad (LDL) son los productos finales del metabolismo de las VLDL. Su núcleo está formado principalmente por ésteres de colesterol y su superficie sólo presenta un tipo de apolipoproteína, apoB. Cerca del 60-80 por ciento del colesterol plasmático es transportado por las LDL. Los valores medios de LDL varían entre distintas poblaciones debido a factores genéticos y ambientales, siendo sin embargo la alimentación el principal factor determinante de estos valores.
Las lipoproteínas de alta densidad (HDL) transportan el 15-40 por ciento del colesterol del plasma. Probablemente se forman en el torrente circulatorio a partir de precursores generados en el hígado y en el intestino. La principal apolipoproteína de las HDL es apoA-1. En los seres humanos, las LDL conducen el colesterol al hígado, y las HDL pueden transferirlo a otras partículas LDL lipoproteicas. Existen pruebas de que las HDL protegen activamente las paredes de los vasos sanguíneos (Consenso del NIH, 1993). No se sabe si la manipulación de los niveles de HDL a través de la alimentación afecta al desarrollo de la aterosclerosis.
La lipoproteína(a) o Lp(a) es un complejo de LDL con apolipoproteína(a). Esta apoproteína presenta una homología de secuencia con la proenzima plasminógeno, que interviene en la disolución de los coágulos de sangre (Scanu y Scandiani, 1991). La concentración de la Lp(a) viene determinada principalmente por factores genéticos.
Metabolismo de los ácidos grasos
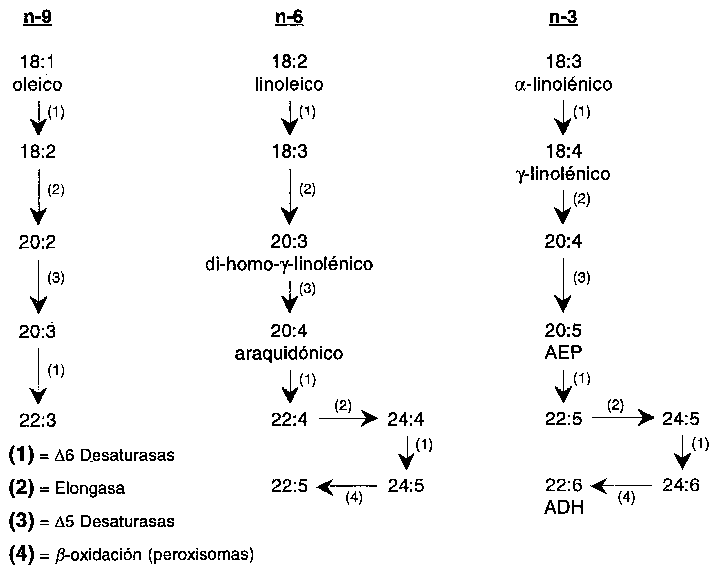
Los ácidos grasos saturados y monoinsaturados pueden biosintetizarse a partir de hidratos de carbono y proteínas. Los ácidos grasos insaturados pueden ser sustratos de las desaturasas y de las elongasas, como se muestra en el esquema de conversión de las familias de ácidos grasos n-9, n-6 y n-3 (Figura 3.2).
Los primeros miembros de cada familia de ácidos grasos (ácidos oleico, linoleico y a -linoleico) compiten por la misma 6-desaturasa, cuya velocidad de conversión aumenta con el número de dobles enlaces. Esta enzima limitante de la velocidad se encuentra bajo el control de muchos factores dietéticos y hormonales (Brenner, 1989; 1990), y se cree que es importante en la síntesis de 22:6 n-3. Este tipo de efecto puede explicar por qué las ingestiones elevadas de ácido linoleico reducen el nivel de 22:6 n-3. De un modo similar, la actividad 5-desaturasa está modulada por factores dietéticos y hormonales. Los miembros C20 y C22 de las familias n-6 y n-3 pueden inhibir antes la desaturación en la secuencia de la conversión de los ácidos grasos (Fisher, 1989).
Parece que la desaturación de la posición 4 no implica ninguna otra desaturasa específica, sino una elongación seguida de la acción de la 6-desaturasa, ambos procesos microsomales, seguidos de una retroconversión por la ruta de la b -oxidación peroxisomal (Voss et al., 1991). La desaturación es por tanto un proceso basado en la cooperación intracelular. El significado nutricional de este papel adicional de la 6-desaturasa todavía debe evaluarse. Las actividades de las elongasas parecen ser mayores que las de las desaturasas (Sprecher, 1989).
FIGURA 3.2 - Secuencia de conversiones de los ácidos grasos insaturados
Acidos grasos esenciales
Los ácidos grasos esenciales son aquellos que deben suministrarse en la alimentación, e incluyen miembros tanto de la serie n-6 como de la serie n-3. Cuando en la alimentación hay deficiencia de ácido linoleico, el ácido graso insaturado más abundante en los tejidos, el oleico, es desaturado y elongado para dar ácido eicosatrienoico, n-9, del que normalmente sólo se encuentran trazas. La acumulación de este ácido graso se considera un indicio de deficiencia de ácidos grasos (Holman, 1970). Cuando la alimentación tiene un bajo contenido de ácidos grasos n-3, en relación con los n-6, se produce una reducción de 22:6 n-3 junto con una acumulación compensatoria de 22:5 n-6 en los tejidos (Galli, Agradi y Paoletti, 1974).
El producto más abundante de la ruta n-6 es el ácido araquidónico que se encuentra en los glicerofosfolípidos, donde se esterifica selectivamente en la posición 2. Los alimentos que contienen grasas de origen animal proporcionan cantidades apreciables de ácido araquidónico preformado.
Se ha puesto en duda la eficacia de la ruta n-3 en los seres humanos. Sin embargo, los estudios realizados en seres humanos sobre la conversión de ácidos linoleico y a -linolénico deuterados en sus derivados de cadena larga han mostrado que la eficiencia de la conversión de linoleico en araquidónico era del 2,3 por ciento, mientras que la conversión del a -linolénico en su derivado de cadena larga era del 18,5 por ciento (Emken et al., 1992). Estos valores de conversión, medidos como productos acumulados en el plasma, se obtuvieron en personas que habían ingerido cantidades relativamente bajas de ácido linoleico (alrededor del 5 por ciento). Cuando la alimentación contenía alrededor del 9 por ciento de ácido linoleico, las conversiones se reducían al 1 por ciento en el caso del linoleato y al 11 por ciento en el del linolenato. Estos datos indican que en los adultos se produce la síntesis de los metabolitos de n-3 a partir de 18:3.
El aceite de pescado, que en comparación con el aceite de linaza es rico en ácido eicosapentanoico, aumenta en gran medida la concentración de derivados de cadena larga en la sangre humana (Sanders y Roshanai, 1983). En las plaquetas y otras células de la sangre sólo se alcanzan niveles elevados de ácido eicosapentanoico y docosapentanoico cuando se ingieren como tales en la alimentación. Los ácidos grasos n-3, así como los ácidos grasos n-6, se incorporan a la posición 2 de los fosfolípidos de membrana.
Los ácidos grasos poliinsaturados de los fosfolípidos de membrana proceden de los alimentos o de mentes endógenas. Su disponibilidad está influida por la desacilación-reacilación, en particular en la posición 2 de los fosfolípidos (Lands et al., 1982), por la especificidad de las kinasas de diacilglicéridos en la síntesis de los fosfolípidos (McDonald et al., 1988), y por el remodelado de los fosfolípidos donadores y aceptores. En todos estos procesos, existe competición entre los ácidos grasos de las series n-6 y n-3.
La importancia del ácido linoleico se demostró en estudios realizados con ratas. Cuando las ratas se alimentaban con alimentos carentes de grasas, empezaban a aparecer síntomas específicos que se evitaban suministrando ácido linoleico. La deficiencia en ácido linoleico, una condición poco común en los seres humanos, se había descrito en niños a los que se proporcionaban alimentos carentes de grasas (Hansen et al., 1963). En los niños, empezaban a aparecer trastornos de la piel similares a los que se producían en las ratas y se curaban por administración de ácido linoleico. Para evitar la acumulación de ácido eicosatrienoico (20:3 n-9), la cantidad de ácido linoleico que se requiere en la alimentación es de aproximadamente el 1 por ciento del aporte energético. Esto es aplicable a todas las especies animales probadas. Aportes comprendidos entre el 4 y el 10 por ciento de la energía son los consumidos normalmente por distintos grupos de población, y estos parecen ser compatibles con el estado de salud óptimo.
La deficiencia de ácido linoleico puede surgir como condición secundaria de otras alteraciones, como desnutrición proteico-energética o absorción deficiente de las grasas, o como consecuencia de una nutrición parenteral total con aportes insuficientes de ácido linoleico. En condiciones de aporte adecuado de ácido linoleico, el cociente entre trieno y tetraeno está por debajo de 0,2 (Mohrhauer y Holman, 1963). La deficiencia de ácido linoleico se puede evaluar mejor mediante un perfil de todos los ácidos poliinsaturados del suero (Holman y Johnson, 1981).
El ácido graso n-3, ácido docosahexanoico (ADH, 22:6), está presente en grandes concentraciones en el sistema nervioso central, en las membranas celulares y en el sistema visual (Tinoco, 1982; Neuringer et al., 1984; Bourre et al., 1989). Más adelante en este informe se discutirá el papel del ADH para tener una función neuronal óptima y agudeza visual, relacionándolo con las primeras etapas del desarrollo.
El equilibrio entre los ácidos grasos n-6 y n-3 de la alimentación es importante dada su naturaleza competitiva y sus funciones biológicas esenciales y diferentes. Se ha sugerido que las cantidades relativas de ácido linoleico y a -linolénico presentes en la alimentación deben estar por debajo de 10:1.
Funciones biológicas
Estructura de la membrana. Los ácidos grasos insaturados en los lípidos de membrana juegan un importante papel para mantener la fluidez (Lynch y Thompson, 1984). En la piel, el ácido linoleico tiene un papel específico, uniéndose a algunos ácidos grasos de cadena muy larga (C30-C34) en las acil-ceramidas. Estas forman una matriz intracelular para mantener la barrera de permeabilidad epidérmica (Hansen y Jensen, 1985).
En las membranas, las interacciones entre lípidos y proteínas pueden depender de un ácido graso poliinsaturado específico. Este parece ser el caso de los segmentos externos de los bastoncillos de los mamíferos que son muy ricos en ácido docosahexanoico.
Se han destacado tres ejemplos de interacciones entre lípidos y proteínas que controlan las funciones metabólicas de las membranas: el primero, las propiedades catalíticas de las proteínas de transporte; el segundo, actividades de enzimas como la Ca/Mg ATPasa del retículo sarcoplasmático, la adenilato ciclasa y la 5-nucleotidasa, en las que influyen los niveles de ácidos grasos n-6 y n-3 de los lípidos de membrana (Kinsella, 1990); y el tercero, las enzimas que intervienen en el ciclo de los fosfoinosítidos. Este ciclo, asociado con las respuestas de muchas células frente a una amplia gama de hormonas, neurotransmisores y factores de crecimiento celular, da lugar, a partir de la activación de una fosfolipasa C específica, a dos importantes segundos mensajeros: inositol 1,4,5-trifosfato (IP3), y diacilglicéridos. El IP3 es el encargado de modular los iones calcicos del citosol. El diacilglicérido, junto con los iones calcicos y la fosfatidil serina, interviene en la activación de una proteína kinasa C que fosforila varias proteínas intracelulares. Puesto que los polifosfoinosítidos son muy ricos en ácido araquidónico en la posición 2, el diacilglicérido generado a partir de ellos es también muy rico en este ácido graso. Una lipasa que actúe sobre el diacilglicérido puede en consecuencia liberar ácido araquidónico que podrá convertirse en eicosanoides.
El ciclo de los fosfoinosítidos controla algunos procesos de la división celular. Por tanto, la modulación del ciclo influye en la tasa de división de las células en la respuesta inmunitaria y en el crecimiento tumoral. Los ácidos grasos poliinsaturados de la alimentación y la proporción de ácidos grasos n-6 y n-3 parecen afectar al ciclo de los fosfoinosítidos.
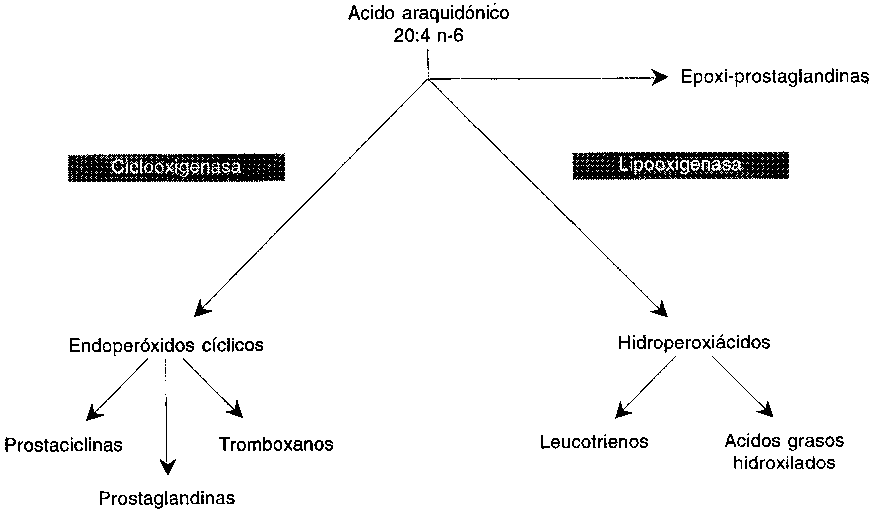
Formación de los eicosanoides. Los ácidos grasos n-6 y n-3 que forman parte de los fosfolípidos de membrana ejercen un control metabólico a través de su papel de precursores de los eicosanoides. Estos compuestos altamente activos de 20 átomos de carbono son liberados en cantidades muy pequeñas para actuar rápidamente en su entorno inmediato. Tras ser degradados enzimáticamente, los productos derivados de los eicosanoides que se encuentran en la orina son un índice de la producción corporal.
El primer paso de la biosíntesis de los eicosanoides consiste en la liberación de un ácido graso poliinsaturado de 20 átomos de carbono por acción de las fosfolipasas sobre los fosfolípidos, principalmente una fosfolipasa A2 o bien sobre los diacilglicéridos que se producen en el ciclo del fosfato de inositol. La secuencia de los eicosanoides consiste en derivados hidroxilados de los ácidos grasos poliinsaturados de 20 átomos de carbono: (a) productos cíclicos, generados por una ciclo-oxigenasa, incluyendo prostaglandinas, prostaciclinas y tromboxano; (b) productos de la lipoxigenasa como los derivados de la 12-lipoxigenasa, especialmente los productos de la 5-lipoxigenasa conocidos como leucotrienos; y (c), productos de la actividad del citocromo P450 (Figura 3.3). Los eicosanoides son en general muy potentes, sus efectos son muy diversos, y la acción de unos eicosanoides es con frecuencia contraria a la de otros diferentes. Además, los patrones de producción de los eicosanoides son diferentes en las distintas células y tejidos.
Entre los eicosanoides más activos se encuentra el tromboxano A2 (TxA2), derivado del ácido araquidónico y producido en las plaquetas y otras células a través de la ruta de la ciclo-oxigenasa. Este eicosanoide es un agente proagregante de plaquetas y de contracción del músculo liso, y se inactiva rápidamente originando tromboxano B2. La prostaciclina, producida mediante la ciclo-oxigenasa en las células de las paredes de los vasos sanguíneos, es un agente anti-agregante de las plaquetas y vasodilatador. Otros productos de la ruta de la ciclo-oxigenasa (como PEG2 y PGF2a) ejercen diversos efectos sobre las células del músculo liso, sobre las células inmunocompetentes, y así sucesivamente. Entre los productos de la ruta de la lipoxigenasa, los leucotrienos, producidos fundamentalmente por los leucocitos, actúan sobre los parámetros vasculares (permeabilidad, contractibilidad), y presentan propiedades quimiotácticas. Intervienen en la modulación de los procesos inflamatorios e inmunitarios.
FIGURA 3.3 - La secuencia de los eicosanoides
Los ácidos grasos poliinsaturados de 20 átomos de carbono y distintos grados de insaturación dan lugar a eicosanoides con distinto número y patrones de insaturación, y con actividades biológicas algo diferentes. Puesto que el ácido araquidónico (20:4 n-6) es el principal ácido graso poliinsaturado celular, la serie 2 de los eicosanoides es la más abundante y generalmente la más activa. Cuando se incorpora a los lípidos celulares un ácido graso poliinsaturado de 20 átomos de carbono con distinto número de dobles enlaces, por ejemplo, ácido di-homo-gamma linolénico (ADGL), 20:3 n-6, o ácido eicosapentanoico (AEP), 20:5 n-3, se producen respectivamente eicosanoides de la serie 1 o de la serie 3. Estos ácidos grasos también compiten con el ácido araquidónico por la ciclo-oxigenasa, y por lo tanto reducen la formación de eicosanoides de la serie 2. El AEP favorece la formación de eicosanoides de la serie 3 e inhibe la formación de eicosanoides de la serie 2.
Efectos sobre otros intermediarios derivados de los lípidos. El resultado de la hidrólisis de determinados fosfolípidos celulares es la formación de otros compuestos biológicamente activos adicionales como el agente de activación de las plaquetas, FAP. Este compuesto deriva de la 1-alquil, 2-acil fosfatidil colina. El FAP es un agente proinflamatorio sumamente poderoso, y un potente activador de varios tipos de células además de las plaquetas. Aunque los datos sobre los efectos de varios ácidos grasos en la ruta del FAP son limitados, se ha demostrado que la incorporación del ácido araquidónico a los fosfolípidos celulares potencia la producción del FAP (Suga et al., 1991). Los distintos ácidos grasos poliinsaturados de la posición 2 de los fosfolípidos precursores pueden modificar la formación del FAP como se demostró con la reducida producción del FAP por los monocitos de individuos que habían recibido ácidos grasos n-3 (Sperling et al., 1987). Los efectos opuestos de los ácidos grasos n-6 y n-3 pueden explicar algunas influencias de los ácidos grasos poliinsaturados sobre la función de ciertas células (Id.).
Efectos sobre otros parámetros. Los ácidos grasos n-3 parecen afectar a muchos otros procesos, como la producción de citoquinas y de otros factores. Las citoquinas son una familia de proteínas producidas y liberadas por las células implicadas en los procesos inflamatorios y en la regulación del sistema inmunitario. Incluyen las interleukinas y los factores de necrosis tumoral. El mecanismo mediante el cual los ácidos grasos n-3 afectan a la síntesis de las citoquinas no está claro, pero algunos estudios han demostrado un efecto sobre los niveles de ARNm, lo que sugiere un nivel de acción pretraslacional.
Los efectos de los ácidos grasos sobre la expresión de genes que codifican enzimas que intervienen en el metabolismo de los lípidos, así como sobre la expresión de genes que actúan en la regulación del crecimiento celular (genes de respuesta temprana inmediata) representan un importante aspecto adicional en los papeles biológicos de los ácidos grasos poliinsaturados. Parece que los ácidos grasos pueden interaccionar con un grupo de proteínas receptoras nucleares que se unen a ciertas regiones del ADN, alterando por tanto la transcripción de los genes reguladores.
Gran parte de la literatura relacionada con los ácidos grasos de cadena larga n-3 trata sobre dosis farmacológicas o se refiere a las lipoproteínas y su relación con las enfermedades coronarias del corazón. Se deben estimular otros estudios clínicos y metabólicos adicionales.
Oxidación de los ácidos grasos
Los ácidos grasos que no se utilizan para sintetizar eicosanoides ni se incorporan a los tejidos se oxidan para producir energía. Los ácidos grasos proporcionan energía a través de la beta oxidación que tiene lugar en las mitocondrias de todas las células, excepto las del cerebro y las del riñon. Entran en la mitocondria en forma de los correspondientes derivados de acil carnitina. Los ácidos grasos saturados de cadena corta, media y larga se someten al primer paso de la beta oxidación con distintas deshidrogenasas. El proceso va generando sucesivamente moléculas de acetil-CoA que entran en el ciclo de los ácidos tricarboxílicos o en otras rutas metabólicas. El producto final de los ácidos grasos con un número par de átomos de carbono es el acetato. Los ácidos grasos insaturados requieren dos pasos enzimáticos más que los ácidos grasos saturados para cambiar los dobles enlaces en cis a trans y para desplazarlos de la posición alfa a la beta. Aún así, la oxidación de los ácidos grasos insaturados, incluidos el ácido linoleico, es tan rápida o más que la del ácido palmítico. La reacción de la oxidación inicial es realizada por una enzima distinta de la que se encuentra en las mitocondrias; el acil-CoA graso entra directamente en esta organela. El proceso no conduce a la producción completa de acetato, sino que a la mitocondria se transfiere un ácido graso acortado para completar la oxidación. Los ácidos grasos de cadena larga (>20C) son oxidados preferentemente por los peroxisomas; también los ácidos grasos con menos de 14C se oxidan mediante este sistema. La oxidación peroxisomal es normalmente menos eficaz que la mitocondrial y produce más calor. Este tipo de oxidación puede inducirse con alimentos de alto contenido en grasas, así como con una gran variedad de xenobióticos.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}