![]()
![]()
![]()
El interés sobre la participación de los lípidos en la respuesta inmunitaria empezó cuando se reconoció su efecto modificador del sistema del retículo endotelial (Di Luzio, 1972). Un estímulo adicional para este interés fue la sugerencia de que podía darse una asociación entre algunos tipos de cáncer y la cantidad y calidad de los lípidos de la alimentación (Carroll y Khor, 1975; Tannenbaum y Silverstone, 1953). Además, se ha demostrado que las grasas influyen en la gravedad de las enfermedades autoinmunitarias, así como en la duración de la aceptación de homo-injertos (Mertin y Hunt, 1976; Ring et al., 1974). Además, la adición de ácidos grasos a los cultivos in vitro de linfocitos modificaban su respuesta mitogénica (Mertin y Hughs, 1975).
Para la elaboración de una respuesta inmunitaria satisfactoria, es esencial la cooperación entre las distintas células del sistema inmunitario a través de diferentes lípidos y proteínas intermediarios. Los lípidos de la alimentación pueden afectar al sistema inmunitario influyendo en la disponibilidad del sustrato de la ciclooxigenasa y de la lipooxigenasa. Estos productos, a su vez, actúan como intermediarios lipidícos en el control del sistema inmunitario (Rola-Plaszczynski, 1985, y Goodwin, Messner y Peake, 1974). Por otra parte, las células del sistema inmunitario dependen en gran medida de la función de la membrana celular para realizar operaciones tales como secreción de linfoquinas y anticuerpos, recepción de antígenos, transformación de linfocitos y lisis por contacto. La importancia de los lípidos en el mantenimiento de la integridad de la membrana (Stubbs y Smith, 1984) indica que pueden ser nutrientes críticos en la regulación de la función inmunitaria.
Metabolitos del ácido araquidónico (AA)
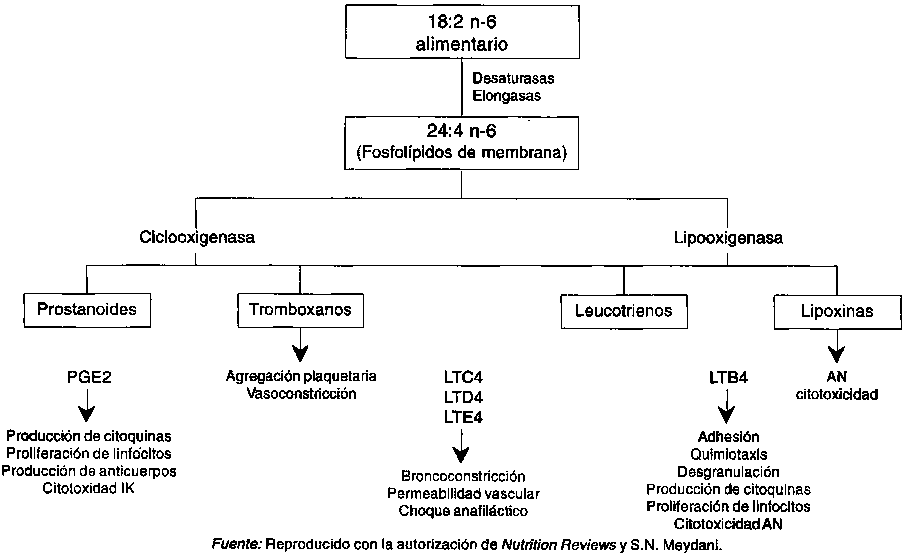
Los metabolitos del ácido araquidónico (AA) intervienen en el control del sistema inmunitario. Los metabolitos del ácido araquidónico (AA), las prostaglandinas (PG), el ácido hidroxieicosatetraenoico (HETE) y los leucotrienos (LT) son producidos por las células mononucleares de la sangre periférica humana (CMSP) y por los esplenocitos del ratón en respuesta a los estímulos de los mitógenos o de los antígenos. También inhiben la subsiguiente proliferación de las células T. De manera similar, la inhibición in vitro de las PG aumenta la proliferación de las células T. Las respuestas inmunitarias celular y humoral se producen bajo el control negativo de las PG. In vitro, la PGE2 inhibe la proliferación de las células T. (Goodwin, Messner y Peake, 1974; Webb, Rogers y Nowowiejiski, 1980; Rola-Plaszczynski y Lemaire, 1985; Metzger, Hoffeld y Oppenheim, 1980; Fisher y Bostic-Bruton, 1982; Muscoplat, Rakich y Thoen, 1978), la producción de linfoquinas (Gordon, Bray y Morley, 1976), la generación de células citotóxicas (Plaut, 1979), y la actividad de las células asesinas naturales (AN) (Roder y Klein, 1979).
Se ha demostrado que, además de la PGE2 los productos de la lipooxigenasa, esto es, LT y HETE, inhiben la proliferación de los linfocitos en los esplenocitos del ratón y en los linfocitos periféricos humanos (Goodman y Weigle, 1980). Este efecto puede estar mediado por un descenso de la proliferación de los linfocitos T ayudadores y por un aumento de la proliferación de las células T supresoras/citotóxicas (Payan Missirian-Bastian y Goetzl, 1984; Gualde et al., 1984). En algunos estudios, pero no en todos ellos, se ha demostrado que el LTB4 aumenta la producción de la interleuquina (IL)-1 así como la de la IL-2, y la proliferación de los linfocitos (Rola-Plaszczynski y Lemaire, 1985). Se ha visto que las lipoxinas (Lx), que son los productos de la 15-lipoxigenasa del AA, inhiben la actividad AN humana medida frente a las células diana K562 (Ramstedt et al., 1985). Por otra parte, se ha visto que el LTB4 aumenta la actividad AN humana (Rola-Plaszczynski, Gagnon y Sirois, 1983). El efecto de los metabolitos del AA sobre la inmunidad celular se esquematiza en la Figura 12.1
Acidos grasos poliinsaturados (AGPI)
Algunos investigadores han evaluado el efecto de las grasas en la respuesta inmunitaria (Johnson y Marshall, 1984; Erickson, 1986). El examen de la bibliografía presenta un cuadro confuso debido principalmente a que el efecto de los AGPI en la respuesta inmunitaria varía dependiendo de diversos factores como son: concentración de las grasas; duración de la suplementación; variación genética; existencia de enfermedades infecciosas o autoinmunitarias e inflamatorias; edad del animal; niveles de otros nutrientes, como vitamina E, cuyas necesidades dependen del grado de saturación de los ácidos grasos; deficiencia o no de los ácidos grasos esenciales en la dieta de control; pruebas inmunológicas empleadas en el experimento; y serie de AGPI probadas, esto es, AGPI n-6 o n-3.
AGPI (n-6) de origen vegetal. Recientemente se han revisado los efectos de los AGPI n-6 de origen vegetal en la respuesta inmunitaria (Meydani et al., 1991b; Dupont et al., 1990). En la mayoría de los estudios realizados con animales, los ácidos grasos esenciales eran necesarios para desempeñar una función inmunitaria normal. La carencia de ácidos grasos esenciales es perjudicial para las respuestas mediadas por las células B y T. Esta carencia se normaliza incluyendo ácidos grasos esenciales en la alimentación. Los regímenes alimentarios con gran contenido de grasas (alrededor del 45 por ciento), independientemente de su grado de saturación, suprimen los índices del sistema inmunitario tanto in vitro como in vivo. Con escasas excepciones, cuando los animales que no presentaban carencias de ácidos grasos se alimentaron con AGPI de origen vegetal (alto porcentaje de AG n-6), respondieron de forma similar a la de los animales que se habían alimentado con una concentración equivalente de ácidos grasos saturados de origen animal o vegetal. Esto es particularmente evidente con niveles moderados de grasa en la alimentación (alrededor de 22 por ciento). Con concentraciones mayores de grasa (alrededor de 45 por ciento), o cuando los ratones estaban expuestos a agentes infecciosos o carcinógenos químicos, los que se habían alimentado con AGPI tendían a dar respuestas más bajas frente a algunas pruebas inmunológicas, pero no frente a todas, que los ratones que se habían alimentado con la misma concentración de ácidos grasos saturados. Estas conclusiones no pueden extrapolarse fácilmente al ser humano porque los sistemas inmunitarios de distintas especies o estirpes dentro de una misma especie responden de diferente manera frente a las alteraciones de las grasas alimentarias. Se necesitan pruebas bien controladas en el hombre para determinar los efectos de las concentraciones y de la saturación de los ácidos grasos en las respuestas inmunitarias humanas.
Los escasos estudios realizados sobre los efectos de las grasas de la alimentación en la respuesta inmunitaria humana han sido estudios epidemiológicos retrospectivos. Los estudios de exploración sobre la esclerosis múltiple (EM), una posible enfermedad autoinmunitaria desmielinizante con cambios de lípidos (Bornstein, 1973), sugieren que se deba a carencia de los AGPI (Bernsohn y Stephanides, 1967; Mertin y Meade, 1977). Un estudio mostró que los pacientes con EM tenían menos ácido linoleico de lo normal en los lípidos de sus linfocitos, y que los pacientes de EM complementados con aceite de cártamo durante dos años presentaban recaídas menos frecuentes y menos graves que los complementados con aceite de oliva (Millar et al., 1973). Otro estudio mostró que los linfocitos de pacientes con EM daban respuestas mitogénicas inferiores que los individuos de control, y que la complementación con aceite de semillas de prímula (que contiene un 8 por ciento de ácido oleico, un 75 por ciento de linoleico, y un 9 por ciento de g -linoleico C18:3n-6) durante 85 días mejoraba la respuesta mitogénica (Offner, Konat y Clausen, 1974).
La suplementación de los pacientes con EM y de individuos sanos con 25 g/día de aceite de cártamo durante cinco semanas aumentó la aglutinación tanto de un antígeno del sarampión como de la concanavalina A (Con A), pero la suplementación con aceite de oliva no tuvo ningún efecto (Utermohlen et al., 1981). La suplementación con AGPI, combinada con una terapia inmunosupresora convencional, facilita la aceptación de los trasplantes de riñón en los seres humanos (Uldall, 1974).
En un estudio transversal sobre la mitogénesis de los linfocitos frente a los mitógenos de las células B y T y a la actividad AN realizada en 94 personas de vida libre, no se encontró correlación entre el grado de insaturación de los ácidos grasos del tejido adiposo (que posiblemente sea el reflejo del consumo dietético) o las lipoproteínas del plasma y los índices inmunológicos (Berry et al., 1987). La variación entre distintas personas y la influencia de otros factores de la alimentación podría enmascarar un posible efecto de las grasas.
Un estudio reciente mostró que los regímenes alimentarios con un contenido de grasas bajo y con moderado AGPI (n-6) (6 por ciento) no tenían ningún efecto en la respuesta de hipersensibilidad retardada (HSR) de la piel que puede medir in vivo las funciones mediadas por las células T en las personas sanas. Sin embargo, aumentó significativamente la respuestas proliferativa de los linfocitos frente a los mitógenos de las células T, la Con A y la producción ex vivo de IL-1b.
AGPI (n-3) de origen vegetal. El ácido linolénico (C18:3n-3) puede convertirse en AEP (C20:5n-3), que puede reemplazar al ácido araquidónico de los fosfolípidos de membrana y ser utilizado de modo preferente por la ciclooxigenasa. Esto ocasiona una producción reducida de PG de las dos series y de leucotrienos de las cuatro series. Así, se ha propuesto que el consumo de aceites que contienen C18:3n-3 podría estar asociado con cambios inmunológicos. Esta propuesta se ve apoyada por muchos estudios que han mostrado una modulación inmunológica tras el consumo de aceite marino. Existen muy pocos estudios inmunológicos sobre los aceites vegetales que contienen AGPI (n-3). Los aceites vegetales que contienen AGPI (n-3) incluyen al aceite de semilla de perilla que tienen el mayor contenido de AGPI (n-3) (64 por ciento), seguido por el aceite de linaza (62 por ciento), el aceite de canola (10 por ciento) y el aceite de soja (7 por ciento). Al extrapolar los resultados obtenidos con el AGPI (n-3) de los aceites marinos a los aceites que contienen C18:3n-3, se deben considerar dos aspectos: primero, cuál es la tasa de conversión de C18:3n-3 en AGPI (n-3) con cadenas más largas (esto es, C20:5n-3 y C22:6n-3); y segundo, ¿el porcentaje del AGPI (n-3) en el aceite es el adecuado para modificar el metabolismo del ácido araquidónico y, por consiguiente, los procesos inmunológicos e inflamatorios?
Un examen bibliográfico reciente (Meydani et al., 1991b) indica que el consumo de aceites que contienen C18:3n-3 está relacionado con una ligera disminución de la producción de eicosanoides y carece de efecto estimulador alguno sobre la respuesta inmunitaria, o este efecto es muy débil. Los consumos a largo plazo de alimentos con contenido de grasas bajo y con un 1,8 por ciento de 18:3n-3 producen un descenso significativo del nivel de AA plasmático asociado a un aumento significativo de la respuesta mitógena de las células T frente a la Con A y la producción de IL-1b (Meydani et al., 1993) en los seres humanos. No afectó a otros índices inmunológicos.
AGPI (n-3) procedentes de aceites marinos. La PGE2 controla distintos aspectos de la respuesta inflamatoria e inmunológica celular. La producción de PGE2 disminuye tras consumir AGPI (n-3). Algunos investigadores han estudiado el efecto producido por un aumento del consumo de estos ácidos grasos sobre la producción de citoquinas y sobre las respuestas inmunitarias celulares. Los estudios en el ser humano han demostrado de manera coherente que la producción de citoquinas proinflamatorias disminuye cuando se ingieren por vía oral concentraciones entre moderadas y altas de AGPI (n-3) de origen marino (Meydani et al., 1993; Kremer et al., 1987; Meydani et al., 1991a; Endres et al., 1989). La disminución de la producción de citoquinas proinflamatorias y de eicosanoides contribuye al efecto beneficioso del aceite de pescado en la reducción de la patogénesis de las enfermedades inflamatorias y ateroscleróticas.
Los primeros estudios epidemiológicos mostraron una incidencia baja de las enfermedades ateroscleróticas, inflamatorias y autoinmunitarias en los esquimales de Groenlandia que consumen alimentos ricos en AGPI (n-3) de origen marino. En los estudios sobre la artritis humana se ha comprobado un ligero efecto beneficioso de los aceites de pescado (Kremer et al., 1990). La suplementación con aceites de pescado redujo los síntomas clínicos de la psoriasis, una enfermedad inflamatoria de la piel (Ziboh et al., 1986). También se demostró un efecto ligeramente beneficioso en la colitis ulcerosa (Lorenz et al., 1989).
Algunos estudios han evaluado el efecto de la suplementación con aceite de pescado sobre la inmunidad mediada por la célula. También se ha observado que la suplementación con aceite de pescado in vivo reduce la proliferación de las células T y B y la DTH, una medida de la inmunidad mediada por la célula in vivo (Meydani et al., 1991; Kramer et al., 1991; Santoli y Zurier, 1989; Virella et al., 1989).
También se ha constatado el efecto inmunosupresor del aceite de pescado en personas (de más de 40 años de edad) que se alimentaban siguiendo las recomendaciones del National Cholesterol Education Panel (NCEP) (NCEP Etapa 2) enriquecidas con AGPI (n-3) procedentes de pescado (que aportaban el 0,54 por ciento de la energía o 1,23 g/día de AEP y ADH) durante seis meses (Meydani et al., 1993). Este consumo a largo plazo de la dieta de la Etapa 2 del NCEP enriquecida con pescado causó un pequeño, pero significativo, descenso del porcentaje de células T ayudadoras, mientras que el porcentaje de células T supresoras aumentó. Este cambio se acompañó de una reducción significativa del DHT y de la respuesta mitogénica frente a la concanavalina A. Se observó una correlación significativa entre los cambios en la DHT y los niveles plasmáticos de AEP. Como se había descrito en el caso de la suplementación con aceite de pescado, se reducía también la producción de citoquinas, IL-1b, TNF-a e IL-6 en las CMSP (Id.). Por el contrario, una alimentación prolongada con la dieta de la Etapa 2 del NCEP con bajo contenido de pescado (0,13 por ciento de la energía o 0,27 g/día de AEP y ADH, pero manteniendo el mismo nivel de los AGPI totales, esto es, el 10 por ciento) aumentó la respuesta mitogénica de las CMSP frente al mitógeno de las células T, de la concanavalina A, e incrementó la producción de IL-1b y TNF-a pero no tuvo ningún efecto sobre la producción de la IL-6 ni de DTH. Aunque la dieta de la Etapa 2 del NCEP enriquecida con AGPI (n-3) procedentes de pescado producía un descenso significativo del cociente plasmático entre el tocoferol y los AGPI, la dieta de la Etapa 2 del NCEP con bajo contenido de pescado no produjo un descenso significativo de este cociente. Los resultados de este estudio, así como los que se habían obtenido en los estudios con animales anteriormente descritos, indican que el nivel de AGPI (n-3) y el de tocoferol son determinantes importantes de la naturaleza del efecto sobre la respuesta inmunitaria que se había observado tras consumir estos ácidos grasos.
La reducida producción de citoquinas proinflamatorias y de eicosanoides proagregatorios puede resultar beneficiosa en los casos en que se produzcan niveles excesivos de estos intermediarios, por ejemplo, en el caso de las enfermedades inflamatorias. Sin embargo, en personas sanas o en aquellas que presentan un estado inmunitario comprometido, la reducción de la producción de citoquinas o de eicosanoides podría comprometer sus funciones biológicas normales, lo que perjudicaría a las defensas del huésped o a la agregación normal de las plaquetas. Esto puede comprenderse mejor si se tiene en cuenta la doble función de estos compuestos, esto es, el papel de regulación fisiológica, así como la contribución a la patogénesis de las enfermedades.
A lo largo de estas líneas, se ha mostrado que la suplementación con aceite de pescado disminuye la producción de IL-2, la proliferación de las células T inducida por los mitógenos, y el DTH, tanto en modelos animales como humanos. También se ha observado un aumento de la hemorragia cerebral asociada al consumo de aceite de pescado. Además, la alimentación con aceite de pescado inhibe la capacidad de las células accesorias del ratón para presentar el antígeno a las células T ayudadoras (Fujikawa et al., 1992), y los ratones que habían sido alimentados con aceite de pescado tenían reducida la capacidad citotóxica de las células de los mastocitomas estimuladas con LPS o INF-g (Hubbard, Somers y Erickson, 1991).
Se ha demostrado que los consumos prolongados de niveles moderados de AGPI (n-3) marinos disminuían el porcentaje de las células T ayudadoras, la respuesta mitogénica ante los mitógenos de las células T (alrededor de un 34 por ciento) y el DTH (alrededor del 50 por ciento) (Meydani et al., 1993). La disminución del DTH tiene un interés especial, ya que se ha visto que esta prueba se puede utilizar para pronosticar morbilidad y mortalidad. El significado clínico de este descubrimiento se ve apoyado por las observaciones respecto a que la alimentación con aceite de pescado reduce la resistencia natural de los ratones frente a la infección con Salmonella typhimurium (Chang et al., 1992), y las ratas alimentadas con dietas que contenían un 9 por ciento de aceite de menhaden tenían una vida más corta que las que se alimentaban con aceite de maíz o con sebo de buey (Berdanier et al., 1992). En los conejos a los que se había suministrado un suplemento rico en aceite de pescado o en aceite de cártamo (5 g/kg.d) durante los siete días siguientes al nacimiento, la eliminación por los pulmones de los Staphylococcus aureus inspirados disminuyó aproximadamente en un 50 por ciento, en comparación con el control salino o con la administración de pequeñas dosis de los aceites (0,22 g/dg.d) (D'Ambola et al., 1991). Las dietas ricas en aceite de pescado incrementaron también la respuesta broncoconstrictora no mediada por la histamina en la anafilaxis pulmonar (Lee et al., 1985). Se requieren estudios adicionales para determinar el efecto de la suplementación con aceite de pescado sobre la respuesta del huésped en los seres humanos.
Sin una protección antioxidante adecuada, la sustitución de los ácidos grasos de la membrana con los AGPI (n-3) fácilmente oxidables de los aceites de pescado, esto es, AEP y ADH, puede potenciar la peroxidación de las membranas celulares. Se ha demostrado un aumento de la actividad de los radicales libres en animales de laboratorio alimentados con dietas de alto contenido de AGPI (n-3) (Nalbone et al., 1989; Herbert y Wills, 1987; Hu M-L et al., 1989). Estos compuestos pueden contribuir al efecto supresor de las funciones mediadas por las células T del aceite de pescado. Estos riesgos, asociados con el consumo de AGPI (n-3), pueden minimizarse mediante la ingestión de los niveles adecuados de un nutriente antioxidante como la vitamina E, sin que se vean comprometidos sus efectos beneficiosos.
Conclusión
Los estudios realizados con animales o cultivos de tejidos, así como los realizados con los seres humanos, indican que tanto el nivel como el grado de saturación de los lípidos de la alimentación influyen en las respuestas inflamatorias e inmunológicas. La naturaleza del efecto depende del tipo de ácido graso, la edad, el antioxidante y el estado de salud de la persona. Algunos de estos cambios inmunológicos están relacionados con efectos clínicos beneficiosos, como la reducción de la citoquina proinflamatoria con los AGPI (n-3) marinos. Otros pueden ser perjudiciales, como la disminución de la función de intermediario de las células T producida por los AGPI (n-3) marinos en la gente de más edad.
Así, deberían considerarse los efectos de los lípidos en la respuesta inmunitaria antes de recomendar que se cambien los lípidos de la alimentación para mejorar otros aspectos de la salud. Además, la recomendación de aumentar el consumo de AGPI debe acompañarse con las adecuadas recomendaciones para aumentar el consumo de antioxidantes, y en particular de vitamina E.
En general, en individuos sanos, la recomendación de disminuir la ingesta de grasas totales, y la inclusión de cantidades moderadas de AGPI (n-6) y (n-3) junto con los nutrientes antioxidantes adecuados, debe mantener una respuesta inmunitaria competente.
![]()
![]()
![]()
{kind=link}