C.D. Allen
Craig D. Allen travaille avec l’United States Geological Survey, Fort Collins Science Center, Jemez Mountains Field Station, Los Alamos, Nouveau- Mexique (États-Unis).
Une introduction à de nouveaux modèles mondiaux de mortalité des forêts due au climat.
Les forêts, qui couvrent aujourd’hui 30 pour cent des terres émergées de la planète (FAO, 2006), sont transformées rapidement et directement, dans de nombreux endroits, par les impacts de la croissance démographique et de l’essor des économies. Moins évidents sont les effets profonds des changements climatiques en cours sur l’état et la situation des forêts du monde. Des exemples récents de stress et de dépérissement des forêts (défini ici comme un taux de mortalité des arbres clairement plus élevé que les taux habituels), liés à la sécheresse et à la chaleur, font l’objet de documentation dans tous les continents boisés, ce qui permet de commencer à détecter des modèles mondiaux. Le présent article introduit ces modèles et envisage la possibilité que de nombreuses forêts et terres boisées soient menacées aujourd’hui de façon croissante de dépérissement dû au climat. Un article plus exhaustif (Allen et al., 2009) traite de la question de façon beaucoup plus détaillée.
Bien que les événements climatiques puissent endommager les forêts de multiples façons, allant des tempêtes de verglas aux tornades et aux ouragans, l’accent est mis ici sur le stress hydrique climatique, provoqué par la sécheresse et les températures élevées.
Il est reconnu que le climat de la planète subit des changements significatifs causés par l’homme; la température moyenne globale a désormais dépassé la fourchette des températures des 1 300 dernières années au moins (GIEC, 2007). Des variations des modèles de climat bien plus marquées sont prévues pour les décennies à venir dans de nombreuses régions, y compris des températures beaucoup plus élevées et des modèles altérés des précipitations qui assurent la disponibilité d’eau pour les plantes.
Comme la plupart des forêts du monde sont situées dans des zones où la température, la lumière et les éléments nutritifs limitent la croissance et la productivité des arbres, le réchauffement récent de la planète, les changements de la composition atmosphérique (c’est-à-dire des concentrations accrues de composés azotés et d’anhydride carbonique provenant d’émissions massives d’origine humaine) et les augmentations locales de l’ensoleillement et des précipitations ont favorisé la croissance de nombreuses forêts, au cours des dernières décennies, dans des lieux ne souffrant pas de pénurie d’eau (Boisvenue et Running, 2006).
Par ailleurs, le tiers environ des terres émergées du globe sont actuellement trop arides pour soutenir la croissance d’arbres, et des superficies étendues de forêts et de terres boisées se situent dans des zones climatiques marginales, où la productivité nette de la végétation primaire est fortement limitée par la pénurie d’eau (Boisvenue et Running, 2006). Dans ces zones semi-arides, les forêts peuvent accuser de fortes baisses de croissance ou des augmentations de mortalité dues à la sécheresse ou aux températures en hausse (Peñuelas, Lloret et Montoya, 2001, par exemple), comme c’est le cas pour les espèces forestières aux extrémités les plus arides de leur aire de répartition (Jump, Hunt et Peñuelas, 2006, par exemple).
Cependant, la croissance et la mortalité dans les forêts plus humides du globe – des forêts tropicales humides aux systèmes boréaux – sont aussi très dépendantes de la sécheresse (Clark, 2004; Nepstad et al., 2007; Soja et al., 2007). Les forêts tempérées présentes sur des sites productifs pourraient manifester des baisses majeures de croissance, des niveaux élevés de mortalité et des effets pluriannuels retardés en raison de l’extrême sécheresse et du stress thermique, comme on l’a observé partout en Europe depuis la sécheresse et la vague de chaleur de 2003 (Ciais et al., 2005; Breda et al., 2006). Les températures plus élevées peuvent, à elles seules, augmenter le stress hydrique des forêts, indépendamment du volume des précipitations (Barber, Juday et Finney, 2000; Angert et al., 2005). Cela étant, il semble qu’aucune forêt au monde n’échappe aux impacts de la sécheresse.
La mortalité des arbres dépend normalement de multiples facteurs interactifs, allant de la sécheresse aux attaques d’insectes nuisibles et aux maladies, rendant souvent irréaliste la recherche d’une cause unique. Toutefois, ce sont les facteurs de stress abiotiques qui sont normalement à la base des problèmes de santé de la forêt, le stress climatique étant considéré comme le principal facteur de déclenchement de nombreuses grandes attaques d’insectes et de maladies (Desprez-Loustau et al., 2006; Raffa et al., 2008).
Le stress hydrique déterminé par le climat peut provoquer directement la mortalité des arbres par des effets aigus à court terme, comme la rupture irréversible des colonnes d’eau dans le fût et les feuilles de l’arbre (cavitation). Les espèces arborescentes présentent des variations marquées aux plans de leur résistance et de leur vulnérabilité à la cavitation, un facteur déterminant de la résistance à la sécheresse. Sous l’effet du stress hydrique, les arbres réduisent au minimum le risque de cavitation par la fermeture des stomates, ce qui diminue les pertes d’eau et la tension consécutive à l’intérieur du xylem. La fermeture des stomates a toutefois son prix, car elle empêche la diffusion du CO2 dans le feuillage, réduisant par là même la photosynthèse. Le stress hydrique chronique sur de longues périodes affaiblira l’arbre et finira par le tuer, soit directement par la privation de carbone, soit indirectement par les attaques de ravageurs comme les scolytes qui abattent les défenses décroissantes d’arbres frappés de privation chronique (McDowell et al., 2008). Les conditions climatiques influencent aussi directement la dynamique des populations d’insectes et d’agents pathogènes mycosiques (Hicke et al., 2006, par exemple). Ainsi, certaines infestations massives d’insectes forestiers tueurs d’arbres pourraient être attribuées à des facteurs climatiques (Raffa et al., 2008). Indépendamment du mécanisme précis, le dépérissement est souvent un processus non linéaire; il peut se manifester soudainement à l’échelle régionale lorsque les conditions climatiques dépassent les seuils physiologiques de tolérance de l’espèce, ou déclencher des infestations d’insectes nuisibles (Allen, 2007).
De nombreux rapports relient la mortalité accrue des forêts à diverses combinaisons climatiques comprenant la chaleur et/ou la sécheresse, par exemple la sécheresse qui a sévi dans les zones tropicales à la suite d’événements graves liés au phénomène El Niño en 1988 et 1997-1998, le réchauffement persistant et diffus d’une grande partie de l’ouest de l’Amérique du Nord depuis les années 1990, et la sécheresse et vague de chaleur extrêmes de l’été 2003 en Europe occidentale.
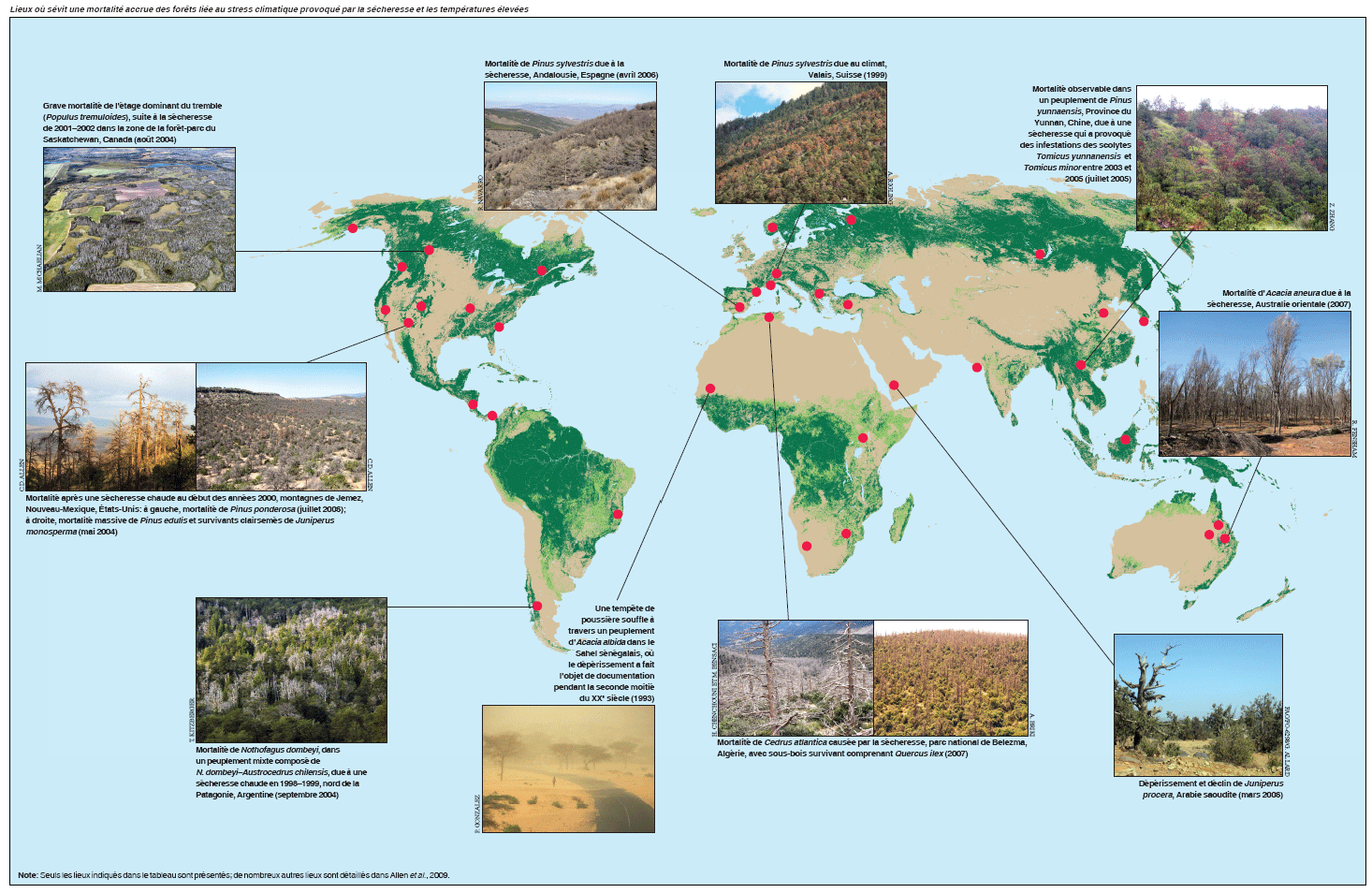
La mortalité des forêts associée à la sécheresse a fait récemment l’objet de documentation dans tous les continents boisés (figure, p. 46); elle concerne divers types de forêts et de zones climatiques. Le dépérissement des forêts généralement communiqué a lieu près des marges géographiques ou altitudinales d’un type de forêt ou d’une espèce forestière (Jump, Hunt et Peñuelas, 2006), vraisemblablement près des seuils historiques de son adaptation climatique, où on peut s’attendre à la sensibilité la plus marquée aux fluctuations climatiques.
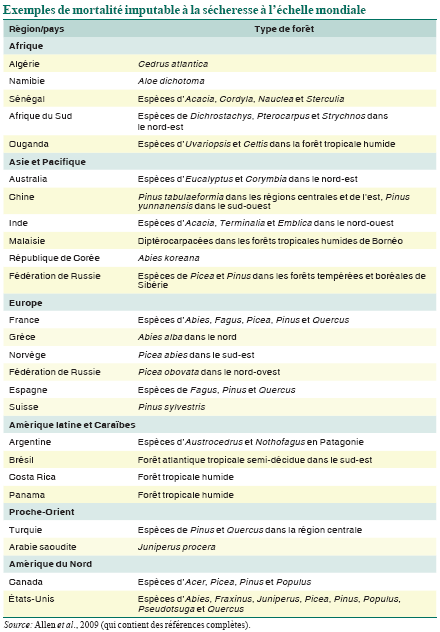
Quelques exemples de mortalité des forêts provoquée par le stress climatique hydrique et thermique depuis 1970, sur la base d’un examen de plus de 120 exemples mondiaux documentés (Allen et al., 2009), sont présentés dans le tableau. Bien que le dépérissement des forêts soit couramment noté dans des régions semi-arides où les arbres sont proches des limites physiologiques de la sécheresse pour la croissance des plantes ligneuses (Fensham, Fairfax et Ward, 2009, par exemple), il est clair que la sécheresse et le stress thermique provoqués par le climat peuvent causer le dépérissement des forêts dans une vaste gamme de types de forêts et de terres boisées dans le monde. Sont particulièrement bien documentés des exemples concernant les zones méridionales d’Europe (Peñuelas, Lloret et Montoya, 2001; Breda et al., 2006) et les forêts tempérées et boréales de l’ouest de l’Amérique du Nord, où les taux de mortalité communiqués ont augmenté rapidement au cours des dernières décennies (van Mantgem et al., 2009) et où la mort diffuse de nombreuses espèces d’arbres dans de multiples types de forêts a touché plus de 10 millions d’hectares depuis 1997 (Breshears et al., 2005; Raffa et al., 2008).
 |
Il est fondamental d’évaluer les possibilités et les conséquences des dépérissements très étendus des forêts provoqués par le climat, car les arbres poussent assez lentement mais peuvent vite mourir. Un arbre de 100 ans pourrait être tué par une sécheresse grave en l’espace de quelques mois à quelques années. De ce fait, la mortalité des forêts déclenchée par la sécheresse peut déterminer des changements rapides de l’écosystème sur d’immenses superficies bien plus rapidement que les transitions progressives qui ont lieu grâce à la régénération et à la croissance de l’arbre. Les impacts sur les utilisations des terres, comme les incendies allumés par l’homme et la fragmentation des forêts, interagissant avec le stress dû au climat, sont susceptibles d’amplifier le dépérissement de la forêt dans certaines régions, par exemple dans le bassin amazonien (Nepstad et al., 2008). Si les écosystèmes forestiers actuels sont forcés de s’adapter brusquement à de nouvelles conditions climatiques du fait d’un dépérissement massif de la forêt, il en résultera de nombreuses conséquences écologiques et sociales profondes et persistantes, provoquées par la perte de produits forestiers et de services écosystémiques – y compris le piégeage du carbone atmosphérique.
L’une des conséquences d’un dépérissement marqué est la redistribution des réservoirs de carbone au sein de l’écosystème et la libération rapide du carbone dans l’atmosphère. Par exemple, les effets du dépérissement des forêts dus au climat et leur mortalité causée par les insectes et les maladies, ainsi que les impacts des incendies, ont récemment transformé les forêts tempérées et boréales du Canada, autrefois un puits net de carbone, en une source nette de carbone (Kurz et al., 2008). De même, il est possible qu’un effondrement diffus des forêts dû à la sécheresse transforme les forêts tropicales humides de la terre en une vaste source nette de carbone au cours de ce siècle, alors qu’elles étaient un puits net dans le passé (Lewis, 2005).
Vu les risques potentiels du dépérissement des forêts provoqué par le climat, on peut s’attendre à ce que la gestion accorde une attention particulière aux systèmes d’adaptation visant à accroître la résistance et la résilience des forêts à des stress climatiques prévus; citons par exemple la réduction de la densité des peuplements à l’aide de coupes d’éclaircie pour affaiblir la concurrence, l’amélioration de différents génotypes (résistance à la sécheresse) ou le déplacement des espèces pour les adapter aux changements climatiques attendus.
Les forestiers et les écologistes savent de longue date que le stress climatique a des effets prononcés sur la santé des forêts. La prise de conscience de ces questions et l’intérêt porté au dépérissement des forêts produit par le climat ne sont pas nouveaux (Auclair, 1993; Ciesla et Donaubauer, 1994). Il est notoire que la variation naturelle du climat a déclenché dans le passé des épisodes de mortalité répandue des forêts (Swetnam et Betancourt, 1998). On pourrait donc se demander si quelque chose de nouveau ou de différent a lieu maintenant. Certes, la Terre connaît à l’heure actuelle un changement climatique marqué, rapide et directionnel, impulsé par des altérations humaines considérables et profondes de l’atmosphère et de la superficie des terres et des eaux de la planète (GIEC, 2007). En même temps que ces changements, la mortalité des forêts liée au climat augmente apparemment dans de nombreuses parties du monde. Bien que les preuves disponibles ne soient pas encore concluantes, il est possible que les notifications croissantes de dépérissement ne représentent que le début de fortes intensifications des problèmes mondiaux relatifs à la santé et au dépérissement des forêts. Étant donné les signes de dépérissement déjà communiqués dans le cadre des hausses récentes relativement modestes de la température mondiale moyenne (environ 0,5 °C depuis 1970) et du climat plus sec dans certaines zones (Seager et al., 2007, par exemple), on peut s’attendre à des risques de stress et de mortalité chroniques bien plus importants dans les forêts, car des augmentations beaucoup plus fortes de la température moyenne (de 2 à 4 °C dans l’ensemble et davantage dans certains endroits) et de graves sécheresses régionales et à long terme dans certaines zones sont prévues d’ici à 2100 (GIEC, 2007). Au-delà des variations des conditions climatiques moyennes, d’autres changements climatiques tels que des sécheresses extrêmes, la hausse des températures maximales et des vagues de chaleur prolongées, qui devraient augmenter en fréquence et intensité (GIEC, 2007), pourraient aggraver le dépérissement des forêts.
À l’heure actuelle, un certain nombre d’incertitudes scientifiques et de lacunes dans l’information limitent les conclusions qui peuvent être tirées quant aux tendances de la mortalité des forêts et aux prévisions qu’on peut formuler sur le dépérissement futur des forêts déclenché par le climat. On observe tout d’abord, malgré de nombreux efforts mondiaux et même régionaux de monitorage des forêts, une absence de données mondiales adéquates sur l’état de santé des forêts (FAO, 2006). Il faudrait une surveillance fiable à long terme de la santé des forêts à l’échelle de la planète, associée à des mesures prises par télédétection et sur le terrain, pour déterminer avec précision l’état et les tendances du stress et de la mortalité des forêts du monde, ainsi que pour comprendre les réactions de l’écosystème après des événements causant le dépérissement.
En deuxième lieu, des données quantitatives adéquates sur les seuils physiologiques de la mortalité d’arbres individuels due au stress hydrique chronique ou aigu ne sont disponibles que pour quelques espèces d’arbres (McDowell et al., 2008), et la vulnérabilité à la température est largement inconnue. En outre, ne sont guère compris de façon détaillée les séquences propres aux lieux et l’écart entre les conditions climatiques moyennes et extrêmes pouvant déclencher la mortalité d’espèces forestières particulières dans des paysages réels, et entraîner potentiellement le dépérissement à grande échelle de la forêt.
Troisièmement, les scientifiques ne connaissent pas assez les réactions en retour et les interactions non linéaires entre le stress des forêts provoqué par le climat et les autres processus climatiques de perturbation, comme les infestations d’insectes et les incendies, qui peuvent causer la mortalité diffuse de la forêt (Allen, 2007).
Ces incertitudes scientifiques concernant les processus fondamentaux de mortalité des arbres représentent une limitation importante à la modélisation quantitative plus précise du dépérissement futur des forêts dû au climat (Huntingford et al., 2008, par exemple). En conséquence, elles limitent aussi la capacité de prévoir les retombées du dépérissement sur l’aptitude potentielle des forêts mondiales à piéger l’excès de carbone atmosphérique ou, autrement, à devenir des sources de carbone et contribuer ainsi à amplifier les changements climatiques (Lucht et al., 2006).
Dans l’ensemble, une surveillance accrue de la santé des forêts du monde et de nouvelles recherches sont nécessaires pour améliorer la certitude scientifique concernant les dangers d’un dépérissement futur des forêts attribuable au climat, afin de fournir des informations plus précises aux décisions stratégiques et à la gestion des forêts dans le monde.
![]() Bibliographie
Bibliographie
Allen, C.D. 2007. Cross-scale interactions among forest dieback, fire, and erosion in northern New Mexico landscapes. Ecosystems, 10: 797–808.
Allen, C.D., Macalady, A., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., Gonzales, P., Hogg, T., Rigling, A., Breshears, D.D., Fensham, R., Zhang, Z., Kitzberger, T., Lim, J.-H., Castro, J., Running, S.W., Allard, G., Semerci, A. et Cobb, N. 2009. Drought-induced forest mortality: a global overview reveals emerging climate change risks. (à publier)
Angert, A., Biraud, S., Bonfils, C., Henning, C.C., Buermann, W., Pinzon, J., Tucker, C.J. et Fung, I. 2005. Drier summers cancel out the CO2 uptake enhancement induced by warmer springs. Proceedings of the NationalAcademy of Sciences of the United States of America, 102(31): 10823–10827.
Auclair, A.N.D. 1993. Extreme climatic fluctuations as a cause of forest dieback in the Pacific Rim. Water, Air and Soil Pollution, 66(3–4): 207–229.
Barber, V.A., Juday, G.P. et Finney, B.P. 2000. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature, 405: 668–673.
Boisvenue, C. et Running, S.W. 2006. Impacts of climate change on natural forest productivity – evidence since the middle of the 20th century. Global Change Biology, 12(5): 862–882.
Breda, N., Huc, R., Granier, A. et Dreyer, E. 2006. Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Annals of Forest Science, 63: 625–644.
Breshears, D.D., Cobb, N.S., Rich, P.M., Price, K.P., Allen, C.D., Balice, R.G., Romme, W.H., Kastens, J.H., Floyd, M.L., Belnap, J., Anderson, J.J., Myers, O.B. et Meyer, C.W. 2005. Regional vegetation die-off in response to global-change-type drought. Proceedings of the NationalAcademy of Sciences of the United States of America, 102(42): 15144–15148.
Ciais, P., Reichstein, M., Viovy, N., Granier, A., Ogee, J., Allard, V., Aubinet, M., Buchmann, N., Bernhofer, C., Carrara, A., Chevallier, F., De Noblet, N., Friend, A.D., Friedlingstein, P., Grunwald, T., Heinesch, B., Keronen, P., Knohl, A., Krinner, G., Loustau, D., Manca, G., Matteucci, G., Miglietta, F., Ourcival, J.M., Papale, D., Pilegaard, K., Rambal, S., Seufert, G., Soussana, J.F., Sanz, M.J., Schulze, E.D., Vesala, T. et Valentini, R. 2005. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature, 437(7058): 529–533.
Ciesla, W.M. et Donaubauer, M.E. 1994. Decline and dieback of trees and forests: a global overview. Étude FAO: Forêts n° 120. Rome, FAO.
Clark, D.A. 2004. Sources or sinks? The responses of tropical forests to current and future climate and atmospheric composition. Philosophical Transcripts of the Royal Society of London, Series B, 359: 477–491.
Desprez-Loustau, M.-L., Marcais, B., Nageleisen, L.-M., Piou, D. et Vannini, A. 2006. Interactive effects of drought and pathogens in forest trees. Annals of Forest Science, 63: 597–612.
FAO. 2006. Évaluation des ressources forestières mondiales 2005 – Progrès vers la gestion forestière durable. Étude FAO: Forêts n° 147. Rome.
Fensham, R.J., Fairfax, R.J. et Ward, D.P. 2009. Drought-induced tree death in savanna. Global Change Biology. (Sous presse)
GIEC (Groupe d’experts intergouvernemental sur l’évolution du climat). 2007. Climate change 2007 – the physical science basis. Contribution du Groupe de travail I au quatrième rapport d’évaluation du GIEC. Cambridge, Royaume-Uni, Cambridge University Press.
Hicke, J.A., Logan, J.A., Powell, J. et Ojima, D.S. 2006. Changing temperatures influence suitability for modeled mountain pine beetle (Dendroctonus ponderosae) outbreaks in the western United States. Journal of Geophysical Research, 111: G02019.
Huntingford, C., Fisher, R.A., Mercado, L., Booth, B.B.B., Sitch, S., Harris, P.P., Cox, P.M., Jones, C.D., Betts, R.A., Malhi, Y., Harris, G., Collins, M. et Moorcroft, P. 2008. Towards quantifying uncertainty in predictions of Amazon «die-back». Philosophical Transcripts of the Royal Society of London, Series B, 363(1498): 1857–1864.
Jump, A., Hunt, J.M. et Peñuelas, J. 2006. Rapid climate change-related growth decline at the southern range edge of Fagus sylvatica. Global Change Biology,12: 2163–2174.
Kurz, W.A., Stinson, G., Rampley, G.J., Dymond, C.C. et Neilson, E.T. 2008. Risk of natural disturbances makes future contribution of Canada’s forests to the global carbon cycle highly uncertain. Proceedings of the National Academy of Sciences of the United States of America, 105: 1551–1555.
Lewis, S.L. 2005. Tropical forests and the changing earth system. Philosophical Transcripts of the Royal Society of London, Series B, 361: 195–210.
Lucht, W., Schaphoff, S., Erbrecht, T., Heyder, U. et Cramer, W. 2006. Terrestrial vegetation redistribution and carbon balance under climate change. Carbon Balance and Management, 1: 6.
McDowell, N., Pockman, W.T., Allen, C.D., Breshears, D.D., Cobb, N., Kolb, T., Sperry, J., West, A., Williams, D. et Yepez, E.A. 2008. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? Tansley Review. New Phytologist, 178: 719–739.
Nepstad, D.C., Stickler, C.M., Soares-Filho, B. et Merry, F. 2008. Interactions among Amazon land use, forests and climate: prospects for a near-term forest tipping point. Philosophical Transcripts of the Royal Society of London, Series B, 363(1498): 1737–1746.
Nepstad, D.C., Tohver, I.M., Ray, D., Moutinho, P. et Cardinot, G. 2007. Mortality of large trees and lianas following experimental drought in an Amazon forest. Ecology,88(9): 2259–2269.
Peñuelas, J., Lloret, F. et Montoya, R. 2001. Severe drought effects on Mediterranean woody flora in Spain. Forest Science, 47: 214–218.
Raffa, K.F., Aukema, B.H., Bentz, B.J., Carroll, A.L., Hicke, J.A., Turner, M.G. et Romme, W.H. 2008. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. Bioscience,58(6): 501–517.
Seager, R., Ting, M., Held, I., Kushnir, Y., Lu, J., Vecchi, G., Huang, H.-P., Harnik, N., Leetmaa, A., Lau, N.-C., Li, C., Velez, J. et Naik, N. 2007. Model projections of an imminent transition to a more arid climate in southwestern North America. Science, 316: 1181–1184.
Soja, A.J., Tchebakova, N.M., French, N.H.F., Flannigan, M.D., Shugart, H.H., Stocks, B.J., Sukhinin, A.I., Varfenova, E.I., Chapin F.S. et Stackhouse, P.W. Jr. 2007. Climate-induced boreal forest change: Predictions versus current observations. Global and Planetary Change,56(3-4): 274–296.
Swetnam, T.W. et Betancourt, J.L. 1998. Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest. Journal of Climate, 11: 3128–3147.
van Mantgem, P.J., Stephenson, N.L., Byrne, J.C., Daniels, L.D., Franklin, J.F., Fulé, P.Z., Harmon, M.E., Larson, A.J., Smith, J.M., Taylor, A.H. et Veblen, T.T. 2009. Widespread increase of tree mortality rates in the western United States. Science, 323: 521–524.