![]()
![]()
![]()
Bunvong Thaiutsa and Orman Granger
BUNVONG THAIUTSA is a member of the Faculty of Forestry, Kasetsart University, Bangkok, Thailand, and ORMAN GRANGER is a member of the Department of Geography, University of California, Berkeley, California.
The importance of the forest floor as an integral part of the ecosystem has been recognized for a long time. One of the major activities occurring there is ''decomposition'', a term widely used to refer to any changes in biochemistry, appearance and weight. Wood (1974) defined decomposition of Eucalyptus leaves as weight losses due to a number of factors, including the removal and/or consumption of tissues by leaf-feeding invertebrates, leaching, biochemical degradation by microorganisms and biochemical degradation during passage through the guts of invertebrates. Decomposition occurs largely on the forest floor, where bacteria and fungi apparently are among the earliest colonizers of fresh litter.
Wildland ecologists have paid considerable attention to litter decomposition in relation to nutrient cycling and soil productivity. The obvious reason is that litter decay has a pronounced effect on the availability of nutrients, and nutrient availability is a basic determinant of tree growth and timber production.
There are three principal techniques used to study litter decay: (1) the measurement of carbon dioxide evolution or oxygen uptake (Reiners, 1968; Wiant, Jr., 1967a; Wiant, Jr., 1967b; Witkamp, 1966, (2) the observation of the disappearance of leaf disc and a specific constituent, such as cellulose and lignin (Edwards and Heath, 1963; Heath et al., 1964; Heath et al., 1966; Madge, 1965); and (3) the macro-microfaunal activities under the particular substrata! and environmental conditions.
Physico-chemical properties of litter
The foliage of coniferous trees generally decays more slowly than that of' deciduous species (Daubemire and Prusso, 1963; Gosz et al., 1973; Mikola, 1960; Ovington, 1954) because broad-leaved litter contains more potassium, more phosphorus, less lignin and nearly always less ether-soluble fraction than coniferous litter. Differences between hardwood species are also significant (Edwards and Heath, 1963, Heath et al., 1966). Pradhan (1973) reported that decomposition of teak litter was more rapid than that of Acacia arabica litter. Furthermore, leaf litter disappears much faster than twigs and branches (John, 1973; Rochow, 1974), and shade leaves which are softer disappear more rapidly than sun leaves (Willams and Gray. 1974).
Generally speaking, the rate of decomposition is highest in species with maximum ash and nitrogen contents and minimal C/N ratios and lignin contents. The lowest rate of break-clown occurs in species which are relatively poor in ash and nitrogen contents and show the highest lignin content and C/N ratios. Species showing medium ash, nitrogen and lignin contents and an average C/N ratio seem to decompose at an intermediate rate (Singh, 1969). Broadfoot and Pierre (1939) found a highly significant correlation between litter decomposition and each of five independent variables: excess base, water-soluble organic matter, total nitrogen, total ash and total calcium. The multiple coefficient of correlation between percent decomposition and the three variables-total nitrogen, water-soluble organic matter, and excess base-- was found to be 0.86. Kucera (1959) also reported a positive correlation between both rapidity of decay and high ash content of hot-water-soluble materials. Since the chemical composition of litter affects its rate of decomposition, it is assumed also to have great significance in determining the release of nutrients.
Microbial activity
The dominant factors affecting microbial activity on leaf litter are plant species and climate. The effect of leaf species on microflora is determined by the physical and chemical properties of the leaves, both of which are reflected in the C/N ratio. Crossley and Hoglund (1962) found a high correlation between the number of micro-arthropods in litter bags and the rate of litter decomposition. During winter there were few animals on the leaf discs and less leaf tissue was eaten as compared to the summer, when more litter was eaten by more animals on the soil surface.
The number of arthropods in litter also depends on the moisture content of litter. Thus Madge (1965) concluded that there were more animals on the leaf discs during the wet season than in dry season. When animals were completely excluded for nine months, no visible breakdown of oak and beech leaf discs occurred (Edwards and Heath, 1963). The same investigators reported that earthworms removed discs three times faster than smaller invertebrates, the most important of the latter being springtails, enchytraeids and dipterous larvae. A common carbamate insecticide (Carbofuran), when applied at recommended rates, reduced the decomposition rate of red maple to between 0.99 and 1.26 g/m²/day. This is chiefly because such insecticides have been found to be highly toxic to earthworms (Weary and Merriam, 1978).
Climatic conditions
Climate markedly modifies the nature and rapidity of decomposition of plant remains on soil surface and thus exerts an important influence upon the nature and abundance of the organic matter. Moisture and temperature are among the most crucial variables (Brinson, 1977, Singh, 1969) because they affect both the development of plant cover and the activities of microorganisms which are highly critical factors in soil formation. Kononova (1975), citing several other publications, concluded that the highest intensity of organic matter decomposition was observed under conditions of moderate temperature (about 30°C) and soil moisture content of about 60-80 percent of its maximum water-holding capacity. Simultaneous increase or decrease of temperature and moisture beyond the optimal levels brought about a decline in the rate of organic matter decomposition. Therefore, studies in litter decay should not be compared unless temperature and moisture regimes are the same.
Wood (1974) presented linear regressions between 1) microbial decomposition and rainfall (r = 0.80), 2) decomposition and organic carbon (r = 0.75), and 3) decomposition and pH (r = 0.60). The pH result reflected the interaction between rainfall and temperature, revealing slow decomposition both at low pH (low temperature and high rainfall) and high pH (high temperature and low rainfall), and rapid decomposition at intermediate pH levels.
A TROPICAL FOREST How does it become soil? And how rapidly?
Results of a 4-year study of litter decomposition in a tropical evergreen forest
Differences in decomposition rates at various altitudes, due to variation in temperature, were reported by Williams and Gray (1974). Shanks and Olson (1961) compared litter decay beneath natural stands at various elevations and concluded that there was an average decrease in breakdown of nearly 2 percent for each 1°C drop in mean temperature. This result is similar to Mikola's (1960), who found that the decomposition rate decreased as latitude increased. The influence of temperature on the decomposition of lignin is especially marked. At 37°C, lignin decomposes rapidly, with 5060 percent of it disappearing within nine months (Waksman and Gerretsen, 1931). More recently, Meentemeyer (1978) used annual actual evapotranspiration as the index predictor variable of decomposition rate.
Reiners (1968) correlated environmental factors with carbon dioxide evolution rate and found that average temperatures at five and 15-cm soil depth were correlated highly with carbon dioxide production. For each 10°C increase in temperature between 20 and 40°C, the rate of carbon dioxide production doubled (Wiant, Jr., 1967a). No carbon dioxide production at all was detected at 10°C, and at 50°C or above it declined markedly. Carbon dioxide evolution is also strongly influenced by moisture content. In unincorporated organic matter where moisture content fell below 20 percent or in mineral soil where it had fallen to only 5 percent, no carbon dioxide production could be detected. (Wiant, Jr., 1967b).
However, some controversial conclusions on the effects of climatic environment have been reported. Van der Drift (1963) pointed out that precipitation was more important than temperature in litter decomposition, although mild winters and warm springs, when combined with relatively normal soil moisture content, might speed decomposition. Moisture passing through the detritus may be important in speeding decomposition.
The percolating water from rainfall may leach the excrements and remains of organisms down to the lower horizons, where other specialized microbes will attack remaining organic molecules. Therefore, litter decay in dry years is slower than it is in an average year. On the other hand, Jenny et al. (1949) and Witkamp (1966) emphasized that temperature was the most important factor, followed by rainfall.
Since climate is the main factor controlling decomposition, the breakdown rates vary with season. Gosz et al. (1973) and Loomis (1975) found that decomposition of plant residues was rapid in summer, whereas Lang (1974) estimated the leaf litter decay to be 3 75 g/m²/day during the autumn months and 0.80 g/m²/day during the remainder of the year. Boonyawat and Ngampongsai (1974) supported Lang's result when they found that the highest decomposition of hill evergreen forest litter occurred in the late rainy season and early winter (0.36 t/ ha/month) and the lowest rate in summer (0.14 t/ha/month). Madge (1965) concluded that litter disappearance in Nigeria occurred mainly during the wet season, owing to the activity of mites and Collembola.
Even though the differences in decay rates between temperate and tropical regions were studied a few decades ago by Jenny et al. (1949), few investigations have been reported from Southeast Asia. Most previous studies on production and decomposition of forest litter in Thailand (Aksornkoae et al., 1972; Boonyawat and Ngampongsai, 1974, Paovongsar, 1976; Thaiutsa et al., 1978) recorded climatic environment but ignored analytical interpretation. The objective of this study, therefore, is to develop regression models for predicting decomposition rates from climatic information.
Study area
The investigation was carried out at Kgma Watershed Research Station in northern Thailand, located between 18°45'N latitude and 98°54'E longitude. The slope is less than 20 percent, and the elevation between 1 2,50 and 1 500 m. The soil has been developed from granite, gneiss and quartzite, and is classified in the Reddish Brown Lateritic Great Soil Group. It has a sandy loam texture associated with high leaching, resulting in a deep B horizon. According to Spurr and Barnes (1973), the subtype of evergreen forest found here is referred to as "montane rain forest". The principal tree species are those belonging to the Fagaceae family, including Castanopsis acuminatissima, Lithocarpus polystachyus, Quercus kerrii and Quercus kingiana. Crown cover is about 85 percent.
Procedure
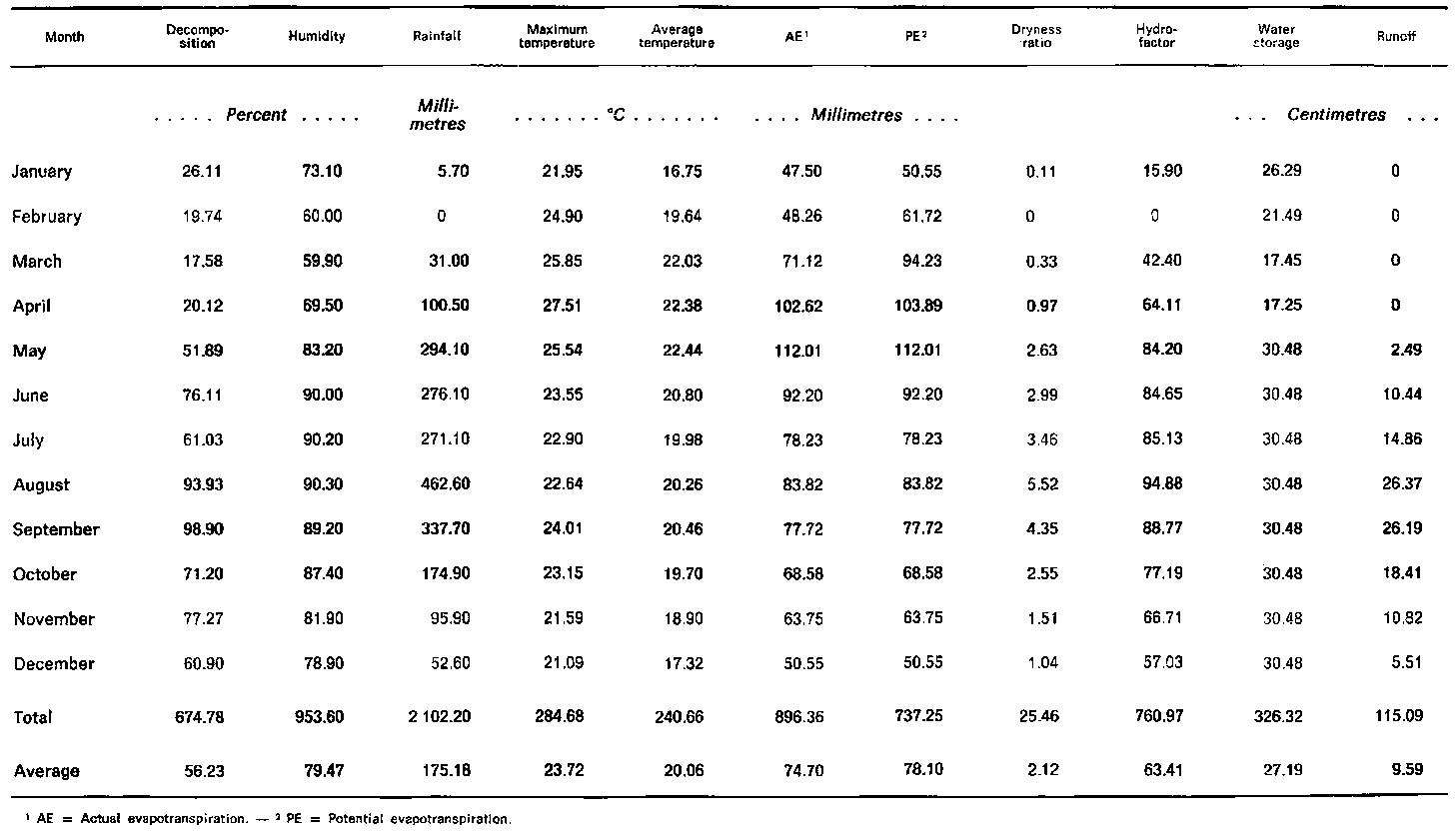
One hundred and fifty-six wooden traps were randomly placed on the forest floor throughout the watershed area of 0.65 km² to measure the litter input and output. Each trap was 0.75 × 0.75 m in size. Twelve of them were controlled for decomposition. Litter collection was made monthly from January 1968 to December 1972. The litter was dried at 70°C for 24 hours, and the weight recorded, the loss in weight being assumed to represent the monthly decomposition rate. Temperature, rainfall and relative humidity were all recorded.
Components of water balance were calculated from the "Water Balance Computer Programme" of Stone, Jr. (1971). Such computation was based on the tables of Thornthwaite and Mather (1957), which gave the values of unadjusted potential evapotranspiration from mean temperature and I (heat index) values. Their tables also provided the mean monthly duration of sunlight at this latitude.
Two secondary variables were derived from the primary climatic data. Hydrofactor value was estimated from the empirical equation cited by Kononova (1975): Hf = 43.2 log P-- T. where Hf is hydrofactor, P is monthly rainfall in mm, and T is mean temperature in °C. The other variable, dryness ratio (DR), was approximated from the equation DR = P/PE, where P is monthly rainfall in mm and PE is potential evapotranspiration in mm.
Simple and multiple regressions between percent decomposition of forest litter and the primary and secondary climatic variables were developed. A selective stepwise technique was, employed in the case of the multiple regressions. In this approach, a sequence of multiple regression equations was computed in a stepwise manner for each relationship. At each step, the variable added was the one that made the greatest reduction in the error sum of squares; or the one which had the highest partial correlation with the dependent variable partialled on the variable which had already been added; or the one which, ii it had been added, would have had the highest F-value. The sequence of interactions was broken by predetermined criteria based on F-values and a tolerance level.
Table 2. Linear regression equations between percent decomposition (D) and relative humidity (RH, %), rainfall (R. mm), mean temperature (T. C), maximum temperature [Tmax, C), monthly actual evapotranspiration (AEm, mm), dryness ratio (DR), and hydrofactor (Hf) of hill evergreen forest, Chiangmai, Thailand
|
Equations |
r |
r² |
|
D = 2286 RH - 125.481 |
0.89 |
0.79 |
|
D = 30.096 + 0.149 R |
0.77 |
0.59 |
|
D = 242.712 - 7.860 Tmax |
- 0.52 |
0.27 |
|
D = 26.267 + 14.122 DR |
0.85 |
0.72 |
|
D = 0.891 Hf - 2.103 |
0.75 |
0.56 |
|
D = 1.626 RH - 0.466 AEm + 0.899 R -- 53.948 |
0.93 |
0.86 |
|
D = 2.284 RH - 0.910 AEm + 6.222 T + 0.607 R - 192.715 |
094 |
0.88 |
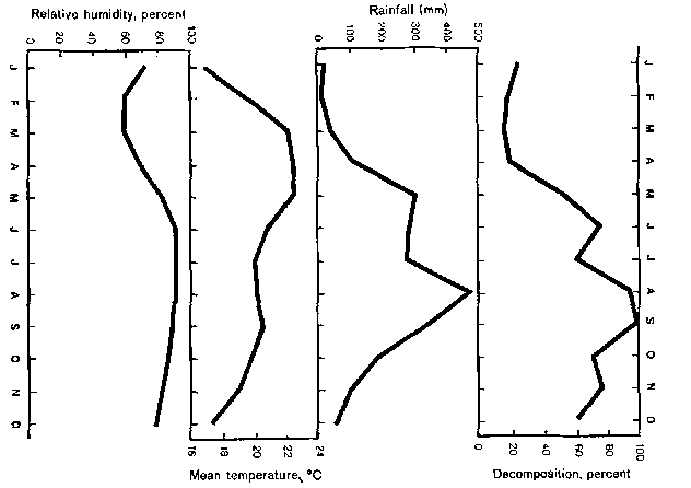
The original and derived climatic data and monthly decomposition rates of hill evergreen forest are presented in Table 1. The primary climatic data are relative humidity, rainfall, maximum temperature and average temperature. The percentage of decay is found to increase with increasing amounts of rainfall and humidity. A reverse trend is true for temperature. Figure 1 shows that the breakdown rate closely follows rainfall intensity, with a lag of about a month. For example, the first peak of decomposition occurred in June while the rainfall peak occurred in May. The second peaks of decay and rainfall were found in September and August respectively.
An important climatic component derived from the recorded rainfall and temperature is evapotranspiration. "Actual evapotranspiration," the amount of water actually evaporated and transpired, is the only term which describes the simultaneous availability of energy and moisture to natural systems. "Potential evapotranspiration," on the other hand, refers to a theoretical loss of water if water supply is unlimited. In the tropics the reason that temperature is not considered a limiting climatic element is because of its role in the consistently high potential evapotranspiration rates. It is this feature of temperature which makes precipitation the more critical and limiting among climatic elements.
Meentemeyer (1974) cited a hundred papers on the role of evapotranspiration in litter decomposition, and developed positive regressions between percent decomposition and actual evapotranspiration. However, the result of the current study on decomposition of forest litter does not indicate as strong a relationship with actual evapotranspiration. The r value of this study was only 0.17, whereas those of the previous investigators were above 0.90. It is generally recognized that the Thornthwaite formulation underestimates actual evapotranspiration in tropical climatic regimes, and this could have influenced the results. It might also be that the soil moisture capacity value used in the computer algorithm (10 inches) was too low for the tropical soil.
Simple and multiple regression equations derived in this study are summarized in Table 2. It was found that percent litter decomposition of hill evergreen forest in northern Thailand was significantly correlated with four climatic components: relative humidity, rainfall, dryness ratio and hydro-factor. Maximum temperature is a much better correlation coefficient value (r = - 0.52) than mean temperature (r = - 0.10). Also, it was found that there was an average increase in litter breakdown of 2.29 percent for each 1 percent increase in relative humidity. Similarly, decay rate at this study site increased by 0.15 percent per 1 mm increase in rainfall.
The climatic factors which accounted for the largest percentage of the variance in decomposition, based on an application of the: stepwise regression technique, were isolated and are presented in Table 3. Humidity alone accounted for 79 percent of the total variance in observed decay rates. Monthly actual evapotranspiration added 2 percent, and rainfall added 5 percent. The r² values of both multiple linear regressions showed that multi-climatic factors led to better prediction than those using only single variables. Neither average nor maximum temperature had the important influence on litter decay that might have been expected from the results of previous investigators. It is therefore clear that precipitation plays a greater role in decomposition than temperature.
Table 3. Correlation matrix and summary table of stepwise multiple correlation
|
Variable |
X1 Decomposition |
X2 Humidity |
X3 Monthly AE |
X4 Rainfall |
|
|
Percent
|
Millimetres
|
||||
|
X1 |
1.00 |
0.89 |
0.17 |
0.77 |
|
|
X2 |
|
1.00 |
0.35 |
0.80 |
|
|
X3 |
|
|
|
0.61 |
|
|
X4 |
|
|
|
1.00 |
|
|
Step number |
Variable entered |
r |
r2 |
r2 increase |
Significance |
|
1 |
X2 |
0.89 |
0.79 |
0.79 |
0.000 |
|
2 |
X3 |
0.90 |
0.81 |
0.02 |
0.001 |
|
3 |
X4 |
0.93 |
0.86 |
0.05 |
0.001 |
Table 4. First-year decomposition rates of forest litter in different locations
|
Forest types or tree species |
Location |
First-year decomposition |
References |
|
Percent | |||
|
Hill evergreen |
Thailand |
56 |
Current study |
|
Teak plantation |
Thailand |
95 |
Aksornkoae et al., 1972 |
|
Teak stand |
Nigeria |
100 |
Egunjobi, 1974 |
|
Tropical rain forest |
Trinidad |
100 |
Cornforth, 1970 |
|
Moist tropical forest |
Ghana |
100 |
Nye, 1961 |
|
Beech and oak |
England |
75 |
Edwards & Heath, 1963 |
|
Annual vegetation |
New Jersey |
58 |
Lang, 1973 |
|
Perennial grasses and shrubs |
New Jersey |
35 |
Lang, 1973 |
|
Oak, red maple, red cedar |
New Jersey |
39-48 |
Lang, 1973 |
|
Oak |
New Jersey |
85 |
Lang, 1974 |
|
Red maple |
Canada |
100 |
Weary & Merriam, 1978 |
|
Eucalyptus delegatensis |
Australia |
35-97 |
Wood, 1974 |
It should be kept in mind, however, that the amount of rainfall measured from rain-gauges may be less significant than soil water, especially surface soil moisture and litter moisture. Soil water plays a more direct role in faunal activities than gauge-recorded rainfall, in part because its role is independent of water surplus or surface runoff. Attempts in' the experiment to show relationship between surface runoff and litter decomposition obtained an equation
Y = 30.869 + 2.644 X,
where Y is percent decomposition and X is runoff in cm. (The r value is 0.90.) This correlation, however, seems to be only empirical, since during the highest months for runoff (August and September) the majority of litter disappearance may have been caused by transportation rather than decomposition. This is the major limitation of the tray technique in approximating weight loss of litter. Soil moisture and litter moisture are therefore expected to be more important than aggregate rainfall. They can be thought of as "effective precipitation," or total moisture available after immediate surface runoff is accounted for.
Since the decomposition rate varies with plant species, climate and microorganisms, the. period required for complete mineralization is different. Table 4 gives information on the first year decay rate of plant residues from various sites. 'the decomposition rate during the first 12 months (56.23 percent of litter production) seems to be lower than those obtained by previous studies from the tropics. However, using an equation for a steady state proposed by Olson (1963):
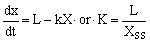
this watershed area is found to have a k value of 2.28. The recomposition parameter k for tropical forests, according to Olson, ranges from slightly more than one to slightly more than four in any given stand. Therefore, the k value of 2.28 found here is within this range. At this rate, complete mineralization would occur within 1.8 years. Differences in such turnover periods affect the development of wildland ecosystems, especially in the tropical zones where forest litter influences wildfire and leaching activity. Therefore, appropriate methods of forest litter management are required.
When the effects of rainfall, temperature and some modified components on litter decomposition of hill evergreen forest in northern Thailand were examined. it was found that 56.23 percent of the annual litterfall decomposed within 12 months, with a decomposition parameter k of 2.28. Complete mineralization of this ecosystem would occur within 1.8 years. Percent decomposition showed significant correlation with relative humidity, rainfall, dryness ratio and hydrofactor. A negative relationship between percent decomposition and maximum temperature was found. A selective stepwise regression indicated that 86 percent of the first-year decay was dependent on humidity, rainfall and actual evapotranspiration. When temperature, either mean or maximum, was added as the fourth independent variable, this percentage was not significantly changed. It was also found that precipitation played a greater role in litter recomposition than temperature. There was an average increase in litter breakdown of 0.15 percent for each l-mm increase in rainfall amount. Surface soil water and moisture content of litter should not be ignored in future investigations.
In order to develop the mathematical models for litter decomposition in relation to climatic factors, certain laboratory studies may be required. However, only field investigation can provide a practical understanding of ecosystem development. Field experiments should cover the wide range of climatic conditions. Surface soil water and moisture content of litter should be accounted for. Detailed studies on production and decomposition of forest litter in various forest types throughout Thailand are expected to be carried out soon.
AKSORNKOAE, S., C. KHENMARK & T. KAEWLA-IAD. 1972. Study on organic matter in teak plantation. Kasetsart Univ. For. Res. Bull., 23. 36 p. (In Thai with English summary)
BOONYAWAT, S. & C. NGAMPONGSAI. 1974. An analysis of accumulation and decomposition of litter fall in hill evergreen forest, Doi Pui, Chiangmai. Kasetsart Univ. Kogma Watershed Res. Bull., 17. 21 p. (In Thai with English summary)
BRINSON, M.M. 1977. Decomposition and nutrient exchange of litter in an alluvial swamp forest. Ecol., 58: 601-609.
BROADFOOT, W.M. & W.H. PIERRE. 1939. Forest soil studies: I. Relation of rate of decomposition of tree leaves to their acid-base balance and other chemical properties. Soil Sci., 48: 329-348.
CORNFORTH, I.S. 1970. Leaf-fall ill a tropical rain forest. J. Appl. Ecol., 7: 603 -608.
CROSSLEY, D.A. & M.P. HOGLUND. 1962. A litter-bag method for the study of microarthropods inhabiting leaf litter. Ecol., 43: 571-574.
DAUBEMIRE, R. & D.C. PRUSSO. 1963. Studies of the decomposition rates of tree litter. Ecol., 44: 589-592.
EDWARDS, C.A. & G.W. HEATH. 1963. The role of soil animals in breakdown of leaf material. In Soil organisms, eds. J. Doeksen and J. V an der Drift. Amsterdam, North-Holland Publ. Co., pp. 76-84.
EGUNJOBI, J.K. 1974. Litter fall and mineralization in a teak (Tectona grandis) stand. Oikos, 25: 222-226;.
GOSZ, J.R., G.E. LIKENS & F.H. BORMANN. 1973. Nutrient release from decomposing leaf and branch litter in the Hubbard Brook Forest, New Hampshire. Ecol. Monogr., 43: 173-191.
HEATH, G.W., M.K. ARNOLD & C.A. EDWARDS. 1966. Studies in leaf litter breakdown. I. Breakdown rates of leaves of different species. Pedobiol., 6: 1-12.
HEATH, C.W., C.A. EDWARDS & M.K. ARNOLD. 1964. Some methods for assessing the activity of soil animals in the breakdown of leaves. Pedobiol., 4: 80-87.
JENNY, H., S.P. GESSEL & F.T. BINGHAM. 1949. Comparative study of decomposition rates of organic matter in temperate and tropical regions. Soil Sci., 68: 419-432.
JOHN, D.M. 1973. Accumulation and decay of litter and net production of forest in tropical West Africa. Oikos, 24: 430-435.
KONONOVA, M.M. 1975. Humus of virgin and cultivated soils. In Soil components, Vol. I, ed. J.E. Gieseking. New York, Springer-Verlag. pp. 475-526.
KUCERA, C.L. 1959. Weathering characteristics of deciduous leaf litter. Ecol., 40: 485-487.
LANG, G.E. 1973. Litter accumulation through ecosystem development. Ph.D. Thesis, Rutgers Univ., New Brunswick, New Jersey. 105 p.
LANG, G.E. 1974. Litter dynamics in a mixed oak forest on the New Jersey Piedmont. Bull. Torrey Bot. Club, 101: 277-286.
LOOMIS, R.M. 1975. Animal changes in forest floor weights under a southeast Missouri oak stand. USFS Res. Note. NC- 184. 3 p.
MADGE, D.S. 1965. Leaf fall and litter disappearance in a tropical forest. Pedobiol., 5: 273-288.
MEENTEMEYER, V. 1974. Climatic water budget approach to forest problems. II. The prediction of regional differences in decomposition rate of organic debris. Publ. Climatol., 27 (1): 35-74.
MEENTEMEYER, V. 1978. Macroclimate and lignin control of litter decomposition rates. Ecol., 59: 465-472.
MIKOLA, P. 1960. Comparative experiment on decomposition rates of forest litter in southern and northern Finland. Oikos, II: 161-166.
NYE, P.H. 1961. Organic matter and nutrient cycles under moist tropical forest. Plant & Soil, 13: 333-346.
OLSON, J.S. 1963. Energy storage and the balance of producers and decomposers in ecological systems. Ecol., 44: 322-331.
OVINGTON, J.D. 1954. Studies of development of woodland conditions under different trees. IT. The forest floor. J. Ecol., 42: 71-80.
PAOVONGSAR, S. 1976. Litter fall and mineral nutrient content of litter in dry dipterocarps forest. M.S. Thesis, Kasetsart Univ. 75 p. (In Thai with English summary)
PRADHAN, I.P. 1973. Preliminary study of interception through leaf litter. Indian For., 99: 440-445.
REINERS, W.A. 1968. Carbon dioxide evolution from the floor of three Minnesota forests. Ecol., 49: 471-483.
ROCHOW, J.J. 1974. Litter fall relations in a Missouri forest. Oikos, 25: 80-85.
SHANKS, R.E. AND J.S. OLSON. 1961. First-year breakdown of leaf litter in southern Appalachian forests. Sci., 134: 194-195.
SINGH, K.P. 1969. Studies in decomposition of leaf litter of important trees of tropical deciduous forest at Varanasi. Trop. Ecol., 10: 292-311.
SPURR, S.H. & B.V. BARNES. 1973. Forest ecology. New York, Ronald Press, Co. 571 p.
STONE, JR., L.A. 1971. Water balance computer program using consecutive monthly data. In Computer program for the climatic water balance, eds. L.A. Stone, Jr., J.C. Albrecht and G.A. Yoshioka. Publ. Climatol., 24(3): 1-10.
THAIUTSA, B., SUWANNAPINUNT & W. KAITPRANEET. 1978. Production and chemical composition of forest litter in Thailand. Kasetsart Univ. For. Res. Bull., 52. 32 p.
THORNTHWAITE, C.W. & J.R. MATHER. 1957. Instructions and tables for computing potential evapotranspiration and the water balance. Publ. Climatol., 10(3): 185-311.
VAN DER DRIFT, J. 1963. The disappearance of litter in mull and mor in connection with weather conditions and the activity of the macro-fauna. In Soil organisms, eds. J. Doeksen and J. Van der Drift. Amsterdam, North-Holland Publ. Co. pp. 124- 133.
WAKSMAN, S.A. & F.C. GERRETSEN. 1931. Influence of temperature and moisture upon the nature and extent of decomposition of plant residues by microorganisms. Ecol., 12: 33-60.
WEARY, G.C. & H.G. MERRIAM. 1978. Litter decomposition in a red maple woodlot under natural conditions and under insecticide treatment. Ecol., 59: 180-184.
WIANT, JR., H.V. 1967a. Influence of temperature on the rate of soil respiration. J. For., 65: 489-490.
WIANT, JR., H.V. 1967b. Influence of moisture content on soil respiration. J. For., 6:3, 902-903.
WILLIAMS, S.T. & T.R.G. GRAY. 1974. Decomposition of litter on soil surface. In Biology of litter decomposition, Vol. II, eds. C.H. Dickinson and G.J.E. Pugh. New York, Academic Press. pp. 611-630.
WITKAMP, M. 1966. Decomposition of litter in relation to environmental factors. Plant & Soil, 15: 295-311.
WOOD, T.G. 1974. Field investigations on the decomposition of leaves of Eucalyptus delegatensis in relation to environmental factors. Pedobiol., 14: 343-371.
Forestry for people

publications you need

Available from
The Community Forestry Officer Forestry Department
Food and Agriculture Organization of the United Nations
Rome 00100 Italy
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}