![]()
![]()
![]()
Luis Martínez Machín[56], Nurys Valenciaga, Tomás E. Ruiz, Ciro Mora and Aida Noda
Key words: legume, sample size.
In recent years, interest to study shrub and tree legumes has increased, maybe because these are one of the few plants that could offer high quality animal forage during the whole year, besides other benefits as multipurpose plants that are essential to design real sustainable animal production systems (González, 1996).
At the beginning, leucaena plants were notable for being free of pests, probably due to the insecticide properties of the mimosine present in young leaves in active growing (Shelton 1996). The apparition of this legume psyllid (Heteropsylla cubana) created incertitude in its use as animal feeding source. However, it has been found that, in our conditions, there is an active permanence of bio-regulators that prevent this psyllid causes economic harms to leucaena crops, at least, with populations present in the activity (Valenciaga et al 1999).
The knowledge of spatial disposition of pests and phyto-pathogenic organisms is of great importance because it permits to foresee sample procedures to be used in future studies, help in selecting the optimum sample size and state and execute correctly a sequence sampling to monitor and control pest insects and diseases (Castillo 1998). The current study has as objective to contribute with the knowledge of behavior and distribution of Heteropsylla cubana in leucaena when determining spatial and time disposition of this psyllid in the pasture area, besides calculating the sample size to be used in similar experiments.
Sampling
Samplings were collected with the use of the entomological net, taking five samples of sixty trappings each using the zigzag diagonals method (Suárez et al 1986) in a 0.5 ha area with a 15-day frequency. Collections were deposited in nylon bags and later taken to the laboratory, where they were put in a stove at 40 ºC during 10 minutes to cause insects death and, then, facilitate their taxonomic classification.
To determine spatial as well as time disposition, the following aggregation indexes were used (Castillo, 1998):
Mean agglomeration index
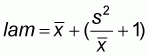 (1)
(1)
Alternative way
 (2)
(2)
Taylor index
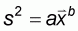 (3)
(3)
Where,
a: Parameter that determines the kind of agglomeration
b: Parameter that determines the kind of spatial disposition
S2: Variance of observations
To determine optimum sample size, the following in equations were used:
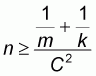
where,
 (4)
(4)
c: probability of the desired error
Spatial disposition is the way in which certain living organisms are distributed in a certain area (bidimensional space). This disposition could be affected by a great number of factors:
Feed availability
Adaptability to the environment for reproduction or survival
Interaction with other organisms of the same or different species
Others
With the information obtained through samplings, spatial disposition of H. cubana in each month of 1999 was determined using mean agglomeration index as observed in table 1. In 91.6% of months there was spatial disposition of “contagium” and only in April, spatial disposition was “regular”. Contagium of the psyllid increased even more from October to December when the rainy season finished and the dry season began. Months with least contagium were May, June and July, i.e., months of the beginning of the rainy season. Such results coincide with those of Valenciaga and Mora (1997).
Table 1. Kind of spatial disposition of H. cubana during the months of 1999 in a Leucaena leucocephala pasture area.
|
Months |
Mean |
Iam(1) |
Kind of spatial disposition |
|
January |
13.6 |
19.61 |
Contagium |
|
February |
60.4 |
69.89 |
Contagium |
|
March |
78.8 |
337.7 |
Contagium |
|
April |
41.8 |
41.21 |
Regular |
|
May |
12.6 |
13.85 |
Contagium |
|
June |
8.4 |
11.84 |
Contagium |
|
July |
2.8 |
4.01 |
Contagium |
|
August |
36.2 |
48.15 |
Contagium |
|
September |
12 |
12.29 |
Contagium |
|
October |
137 |
149.58 |
Contagium |
|
November |
441.6 |
648.53 |
Contagium |
|
December |
91.4 |
98.07 |
Contagium |
(1) Mean agglomeration index
We call time disposition to the way living organisms are distributed in a certain area, in different periods of time. In this experiment, considered data were month averages of the number of observed organisms during the period of analysis (2 years).
The index of mean agglomeration (alternative way) and the Taylor index were used to determine the time disposition. With one of these indexes we could really classify the kind of disposition and agglomeration of organisms, but both were used to confirm results because literature states that is common to observe contradiction with the kind of disposition of organisms when using those indexes for the same data group. Besides, some authors (Castillo, 1998) assure that the best way to describe the kind of spatial disposition is by the Taylor index. Others say that with a high number of data to calculate a and b, in some cases, there is a higher similarity in estimated values of these parameters.
In this experiment there is no contradiction among estimated parameters a and b when considering those indexes. In table 2, we observe that for parameter a, null hypothesis is non-significant, so a = 0, meaning the kind of agglomeration is of isolated individuals. For parameter b, hypothesis test was significant, that is, b ¹ 1 and as the estimated value is higher than 1 in both cases, time disposition is of “contagium”. Thus, sample data are associated to binomial negative probability distribution (Castillo, 1998).
Table 2. Simple lineal regression analysis to determine the kind of time disposition and agglomeration of organisms considering 2 aggregation indexes.
|
Aggregation index |
Parameters |
R2 |
Regression significance |
|
|
|
a |
B |
|
|
|
Mean agglomeration |
7.795 |
1.06 |
0.97 |
*** |
|
Taylor |
0.78 |
1.52 |
0.84 |
*** |
As probability distribution associated to sample data is binomial negative, sample size is calculated by in equation (4).
When substituting the values of m = 132.43, k = 62818.79 and C = 0.05, the optimum sample size obtained is 1433 trappings/ year. This value was calculate by annual mean and variance and later considering the sampling method used. This implies that for each month 24 trappings/ sample should be made because with the 5 samples selected from the field 120 trappings/ month are obtained.
We can conclude that H. cubana remained in the leucaena crop during the whole year with a high level of infestation with spatial disposition of “contagium” in the most part of the experimental period.
Time disposition was analyzed as “contagium” with agglomeration of isolated individuals.
Due to the kind of spatial and time disposition obtained we could say sample data come from a population with binomial negative probability distribution. This allowed to know that the optimum sample size is 1433 trappings/ year in similar experimental conditions.
Authors would like to thank the collaboration of Lic. Pedro J. Padilla in the translation and revision of this work.
Castillo, E. 1998. Notas del curso: Muestreo de poblaciones. Departamento de parasitología Agrícola. Universidad Autónoma Chapingo. México. pp. 86.
González, M. 1996. Leguminosas forrajeras en sistema de producción animal del nor-oriente de Venezuela. Centro de Transferencia en pastos y forrajes. Universidad de Zulia. Venezuela. pp. 115.
Shelton, M. 1996. El género Leucaena y su potencial para los trópicos. Centro de Transferencia en pastos y forrajes. Universidad de Zulia. Venezuela. Pág. 17.
Suárez, R.; Rodríguez, A. y Felipe, A. 1986. Protección de plantas. Editorial Pueblo y Educación. Ciudad de la Habana. pp. 22.
Valenciaga, N.y Mora, C. 1997. Estudio de la incidencia de insectos en una siembra de Leucaena leucocephala intercalada con maíz en dos momentos a partir de la siembra. Rev. Cubana de Cienc. Agric. 31:161.
Valenciaga, N.; Barrientos, A. y Mora, C. 1999. Comportamiento de la entomofauna beneficiosa en área de Leucaena leucocephala (Lam.) de wit. Rev. Cubana de Cienc. Agric. 33:339.
Camarão, Ari Pinheiro[57],
Rodrigues Filho José Adérito[58],
Rischkowsky Barbara[59],
Hohnwald, Stefan[60], Clécio
Mendonça[61]
Key words: Capoeira, cattle, Brachiaria humidicola, enriched pasture, legumes, secondary vegetation, smallholder
The traditional agriculture of the smallholders in northeastern Pará is based on the exploitation of the regenerative potential of the secondary vegetation (capoeira): a fallow period is followed by burning the accumulated biomass of the capoeira to fertilize the soil through the ash and to enhance the productivity of the subsequent food crops, mainly rice, maize, cowpea and cassava. In the last two decades, cattle husbandry has become an important additional farming activity in the Bragantina region (Billot, 1995). Cattle keeping is often a short-term engagement and the productivity and ecological stability of the conventionally managed pasture seems to be low. It is proposed that the integration of pastures as an intermediate stage in the agricultural cycle could be more appropriate for the smallholder situation. A pasture functioning as an intermediate stage could either precede or replace the traditional capoeira, but requires a higher biodiversity than a conventional pasture and still has to meet the nutritional demands of the cattle. The present study compares the botanical composition of the available forage and the cattle diets on conventional, Capoeira regeneration and legume-enriched smallholder pastures.
The experiment was established on a 3.2 ha field, owned by a smallholder in the municipality of Igarapé-Açu, which had been cultivated with annual crops (maize and cassava) for 1.5 years preceded by a 12-year-old capoeira. The area was divided in nine plots representing three types of Brachiaria humidicola pastures with three replications using a randomized block design. A traditional grass-only pasture (TP), is compared to a B. humidicola pasture, enriched with a combination of herbaceous (Arachis pintoi) and bushy legumes (Chamaecrista rotundifolia and Cratylia argentea) (LP). On the third type of pasture, a controlled natural regrowth of the capoeira is tolerated (CP). In the first and second grazing cycle (3/22 - 8/15/2000) three male crossbred cattle were rotated among the three replications resulting an initial stocking rate of 595 liveweight kg per ha. Forage availability was measured at the start of the grazing of each replication, taking five samples per plot. The botanical diet composition was evaluated by micro-histological analysis of feces samples taken from rectum of the three animals (Sparks & Malechek, 1968) at the end of the grazing in each replication.
In the second grazing cycle, the average grazing period per plot was 24 days. The average daily weight gain of the animals was 609 g, 684 g and 737 g on LP, CP, TP respectively. The availability of forage ranged from 9051 to 8229 kg DM/ha (Fig. 1), of which about 39 % was dead material. The botanical composition of the cattle diet showed a dominance of grasses (Fig. 2). The consumption of capoeira species was 11.40 %, 12.43 % and 36.12 % of the ingested forage on TP, LP and CP, consisting of 10, 8 and 21 species, respectively.
Fig. 1: Forage availability on three types of Brachiaria humidicola pasture
The species most frequently consumed by the cattle were Cecropia palmata, Borreria verticilata and Myrcia sp., the latter was found in the samples from all treatments. In addition, the animals fed on Coutoubea spicata, Solanum rugosum, Scleria pterota, Annona paludosa, Paspalum maritimum, Imperata brasiliensis, Memora flavida, Borreria verticillata, Lecythis lurida, Davilla kunthii, Guatteria poeppigiana, Rollinia exsucca, Connarus perottetii, Phyllanthus nobilis, Miconia eriodonte, Digitaria horizontalis, Cordia exaltata and Vismia guianensis. Among the legumes in LP, only Chamaecrista rotundifolia was found in the feces (18.09 %) during the two grazing cycle.
The available leaf biomass of 1470 kg dry matter per ha, the most nutritive part of the forage grass, was above the critical level determined for animal maintenance on pastures (Mott, 1980). The daily weight gains were similar to the performance documented for cattle on planted pastures in the region (Lourenço Júnior et al., 1993). The intake of Chamaecrista rotundifolia forage was not sufficient to increase the animal productivity. Nevertheless, C. rotundifolia persists very well on the pasture and looks a promising component for a multi-species legume pasture. While Cratylia argentea had not been consumed in the first grazing cycle, the animals defoliated the bushes nearly completely in the first three to five days of grazing during the second rotation cycle. However, the available C. argentea biomass is still quite low as the shrubs are not yet fully developed. The browsing of capoeira species, namely Cecropia palmata, Solanum rugosum, Annona paludosa, Memora flavida, Lecythis lurida, Davilla kunthii, Guatteria poeppigiana, Rollinia exsucca, was also observed in beef cattle on planted pastures in Paragominas, PA (Camarão et al., 1990). However, to maintain a balance between capoeira and forage grass, the vigorous regrowth of the herbaceous capoeira species needs to be controlled by selective slashing to prevent the suppression of the forage grass.
The research project was carried out under the auspices of the agreement on scientific-technological cooperation signed by the governments of Germnany and Brazil. The German partner was sponsored by the Federal Ministry of Education and Research (BMBF, FKZ: 01LT0002/9, SHIFT). The Brazilian Partner was sponsored by the Council for Scientific and Technological Development (CNPq, processo: 690076/98-3).
Billot A (1995) Agriculture et systèmes d’élevage en zone Bragantine (Pará-Brésil): diagnostic des systèmes de production familiaux a forte composant élevage. pp 140. CNEARC-EITARC, Montpellier.
Camarão AC., Simão Neto, M, Serrão, EAS, Rodrigues, IA and Lascano, L (1990): Identificação e composição química de espécies de invasoras consumidas por bovinos em pastagens cultivadas de Paragominas, Pará. Embrapa-CPATU, Boletim de Pesquisa, 104, pp61.
Sparks D and Malechek, JC, (1968) Estimating percentage dry weight in diets using a microscopic technique. Journal of Range Management, 21 (4), 264-265.
Mott, GO, 1980: Measuring forage quantity and quality in grazing trials. In: Proceedings of Southern Pasture and Forage Crop Improvement Conference, 37, Nashille, Tennessee, 3-6.
Ma. Jimena Esquivel S.[62], Zoraida Calle D.[63]
Key words: degraded pastures, isolated trees, pioneer species, restoration, seedlings, silvopastoral systems, succession catalyst
High population density, intensive agriculture and cattle ranching have significantly altered the highly diverse ecosystems of the Colombian Andes and have caused negative environmental effects related to forest fragmentation (i.e. species extinction, decrease of water sources and soil fertility). Research applicable to the restoration of mountain forests should be based on the region’s biodiversity and at the same time should provide alternatives for agriculture and cattle ranching.
This study describes the regeneration patterns of trees and shrubs in active pastures and shows how isolated trees enhance species addition during secondary succession in a Premontane Moist Forest locality. Farmers and high school students from the locality were involved as co-researchers.
Applications of this study attend both environmental (forest recovery) and social (re-conversion of cattle production into a sustainable system) needs.
Between January and November, 2000, the taxonomic composition and frequency of establishment of shrub and tree seedlings were evaluated monthly under six pioneer tree species commonly found in active pastures at Bella Vista (Valle del Cauca department, Colombia, 1750 m). The tree species evaluated were: Psidium guajava L. (guayabo) Myrtaceae, Myrsine guianensis (Aubl.) Kuntze (chagualo) Myrsinaceae, Montanoa quadrangularis Sch. Bip. (arboloco) Asteraceae, Nectandra lineatifolia (R. & P.) Mez. (laurel blanco), N. reticulata (R. & P.) Mez. (laurel) and Ocotea oblonga (Meisn.) Mez. (laurel jigua) Lauraceae.
A paired sample design was used for this study. for each tree, two 2m x 2m quadrants were sampled: one located beneath the canopy and the other one in open pasture (without woody vegetation), 5m away from the canopy border. 57 mature trees were evaluated (57 treatment and 57 control quadrants). These trees were heterogeneously distributed in 10 pasture plots within five farms including el Ciprés Natural Reserve.
Regeneration under trees was more abundant (2750 seedlings: 30 species belonging to 15 families) than in open pasture (575 seedlings of 9 species and 7 families). Species richness and seedling abundance were significantly higher under all tree species than in open pasture (control plots), except for Montanoa quadrangularis. No significant differences in species richness in the natural regeneration were found between the different species of isolated trees (Kruskall Wallis, P = 0.1), but differences in seedling abundance were highly significant (Kruskall Wallis, P<<<0.001).
Seedling abundance under LAURACEAE and M. guianensis was significantly different from seedling abundance under P. guajava and M. quadrangularis (LAURACEAE @ M. guianensis >>>P. guajava @ M. quadrangularis).
The distribution of tree and shrub species was not independent from the species of isolated tree (contingency chi-square, P<<<0.001). thus some isolated trees seemed to facilitate the regeneration of certain species and inhibit others. regeneration in open pasture (control) plots was dominated (87%) by P. guajava. M. guianensis was the only other tree species detected in these plots (3%). the remaining 10% of seedlings belonged to shrub species.
Seedling abundance could be higher under bird dispersed isolated trees, though only one wind dispersed species (M. quadrangularis) was studied. 93% of the species of seedlings found under isolated trees in pasture plots have zoochorous dispersal (mainly by birds) and 7% have anemochorous dispersal.
Natural regeneration was five times more abundant and three times more species-rich under isolated trees than in pasture plots without woody vegetation. This result stresses the role of the trees in pastures as centers of animal and plant activity and their importance in the natural succession dynamics of pastureland to forest.
Native pioneer trees commonly found in the Colombian Andes (eastern slope of Cordillera Occidental) such as Myrsine guianensis, Psidium guajava, Nectandra lineatifolia, N. reticulata and Ocotea oblonga, enhance the seed rain and constitute microsites favorable for germination of seeds that cannot arrive or survive in open pasture. The potential of these microsites to enhance species richness suggests the importance of incorporating the protection of isolated trees in pasture management strategies. Propagation of such pioneers is thus recommended in areas assigned to forest recovery.
Though the role of isolated M. quadrangularis as a succession catalyst in pasture is not so clear, this species plays a significant role in the enlargement of mountain forest remnants given its abundant regeneration in forest edges (Calle, in prep.). Its moderate facilitation of tree and shrub regeneration can be viewed as a useful attribute for its incorporation in silvopastoral systems.
These results suggest that maintaining or introducing isolated trees in pasture plots is an efficient way to enrich the vegetation in degraded areas. The complete removal of cattle is not a necessary requisite for forest restoration in degraded pastures. Cattle browsing in areas with reforestation purposes controls excessive pasture growth and at the same time speeds up the invasion of woody species like Psidium guajava. Once the initial barrier to woody species colonization is overcome with the introduction of this tree, other pioneer species like Myrsine guianesis can easily regenerate in active degraded grasslands. P. guajava in grazing areas has productive benefits such as additional food for livestock, birds and humans, good quantity firewood, wood for the farm needs, soil improval through organic matter addition and shade for cattle.
After the initial invasion of this pioneer species, seedlings of secondary growth species become established underneath their crowns increasing the supply of fruits and perches for more specialized bird species that are the most important seed vectors in these habitats. This facilitator system allows the quick recovery of tree and shrub diversity in species-poor habitats.
Typically cattle is completely removed from degraded areas destined to forest restoration. However, the vigorous growth of pasture inhibits the regeneration of woody vegetation. This study suggests that it is possible to take advantage of cattle activity, of the invasive capacity of Psidium guajava, the establishment of native pioneer species, the high biodiversity of the Andes, and animal-plant interactions to catalyze forest recovery in former pasturelands, with low costs and work effort, while producing additional benefits.
T.E. Ruiz, G. Febles, H. Díaz[64]
Key words: agroforestry, biodiversity, environment,
Plant diversity is an important goal that should be considered if we intend to change the livestock environment. Recent data indicate that in Latin America cattle production is characterized by a low productivity, since animals are fed native pastures of poor quality and stocking rates below 0.7 heads-ha (Szott et al 2000). This carrying capacity has also decreased by the degradation of more than 40% of the grazing areas as a consequence of inadequate management practices (Szott et al 2000). Paddock degradation leads to a poor production efficiency, losses of biodiversity and also increases of gases affecting total heating of the planet (Veldkamp 1993, Kaimowitz 1996)..
Thus, the previous statements are of great importance, but at the same time are very complex and their final results cannot be obtained in a short period of time. Therefore, all efforts should be directed to attain these objectives. At present, silvopastoral systems have been only characterized by the presence of Leucaena leucocephala, as tree component and only one grass species. To overcome this situation and increase biodiversity, tree species are being introduced to obtain plant shade, habitat for birds, litter deposition and pods for animal feeding.
Lysiloma bahamensis, Albizia falcataria, Gmelina arborea, Terminalia sp., Azadirachta indica, Erythrina mysorensis were the plants assessed. They were planted in the field from plastic bags at the same time than the system of Leucaena and one grass species was introduced in the paddock area to be later grazed by bovine cattle. The assessment during several years of Lysiloma or Gmelina plant densities (150-200-300 plants/ha) and the combination of other species were also studied.
The available information indicated that with different plant populations (150-200-300 plants/ha) of Lysiloma or the combination of Lysiloma-Albizia the best survival (91%) is obtained with the former tree followed by 60% in Albizia. Growing was also different in these species. Thus, Albizia attained the lowest development at 90 days after sowing with only 41 cm height and 8 leaves/plant. Lysiloma grew up to 125 cm height and 5 branches/plant and 53 leaves/plant. After one year of grazing, the population of 200 plants/ha of Lysiloma showed 78% of survival, decreasing only 12% regarding the initial density. Plants had a height of 1.88 m. Lysiloma-Albizia combination showed the worst performance with only 50 and 5% of survival, respectively.
The study indicated after three years that 200 plants/ha was the best treatment maintaining a survival of 78% and a plant height of 6 m showing a stability in time in an indicator as reliable as survival.
Following this line of work, but with Gmelina, it was observed that a population of 300 plants/ha had the worst survival behavior with a value of only 66% 1 year after the beginning of the experiment. Other treatments had figures between 83 and 87%. It should be emphasized that in all treatments a decrease in plant population after the start of grazing was not evident. With 150 plants/ha a height of 2.6 m was obtained and in the others this measurement varied between 1.74 m and 1.87 m. After 3 years of exploitation with animals, the population survival was maintained for the population of 150 and 300 plants/ha with 82 and 67%, respectively while that of 200 plants decreased by 10 percentile units, attaining only 77%. Height was maintained with 2.4, 1.7 and 1.6 m in the studied populations. This showed that animals consumed these plants when biomass production decreased (Leucaena-grass) in the dry season.
Other experiments but with the same objective included the combinations Terminalia-Albizia; Neem-Erythrina; Gmelina-Albizia; Neem-Albizia and Neem-Gmelina. In these cases Albizia showed again a poor survival of approximately 0-35% obtaining this latter value with the Neem-Albizia combination 1 year after the beginning of grazing. It should be stressed the survival achieved by the Neem tree that was higher than 80% and 3 m height after 1 year of planting. Gmelina confirmed its high survival as in other experiments with more than 77% and a height of more than 2.2 m for the same period. Neem-Gmelina is an interesting combination.
The previous mentioned performance was confirmed two years after this measurement when Neem-Gmelina attained a survival above 70%, as well as a height of 6.7 m and 4.4 m, respectively. On the other hand, Albizia disappeared in the combination with Neem, Gmelina and Terminalia. However, Gmelina presented a very positive and interesting stage of association with the species under evaluation.
We must indicate that when more than one species is planted in near holes, high survival values can be obtained than when only one is planted alone. The alternative of planting all the components at the same time allow the development of the system from early stages and also can decrease the necessary period to introduce animals to make use of the combination and to contribute to a better land utilization. With all the results discussed in this paper we can conclude that it is possible to develop silvopastoral systems with different plant species. Hence, each plant component can be able to carry out the right complementary function in the stratum in which it grows regarding the others and as a consequence a balanced and non-competitive situation can be obtained.
Kaimowitz, D. (1996). Livestock and deforestation. Central America in the 1980s. A Policy Perspective. CIFOR, Jakarta, Indonesia
Szott, L., Ibrahim, M. & Beer, J. (2000). The hamburger connection hangover: cattle, pasture land degradation and alternative land use in Central America. CATIE, Costa Rica
Veldkamp, E. (1993). Soil organic carbon dynamics in pastures established after deforestation in the humid tropics of Costa Rica. Ph.D. Thesis University of Wageningen, N.L.
Zaragoza H C[65],
Clemente S F[66], Mendoza M G[67],
Aguilar V B C[68], Tarango L A[69],
Gómez R.B.[70]
Montezuma quail is a species that uses range vegetation. It has been estimated that near 60% of agricultural land in Central America is covered by rangelands, and overgrazing by livestock has been the main cause of the increasing habitat deterioration (Solano, 1994). Particularly, overgrazing is negative for Montezuma quail, due to competition for food and destruction of vegetation cover that is required by quail has nesting areas and escape cover (Brown, 1982; Albers y Gehlback, 1990). The objective of this study, was to determine relationships between habitat use by Montezuma quail, biodiversity, and biomass production in forested and range land areas in the Northwestern portion of the State of Mexico. This research will contribute to conserving and providing mechanisms for sustainable management of silvopastoral ecosystems.
The study area is located in the Northwestern region of the State of Mexico, and cover 37% of the State within the Game Hunting Region II which encompasses 8,600 km2 (INEGI, 1993). Vegetation was sampled in 1 m2 plots (Bonham, 1989) randomly selected from maps of soil use and vegetation type. Sampling plots (22) covered 2,740 ha at 61 sites within 15 counties. All sampling points were located by GPS (Magellan-350). Vegetation sampling was carried out in winter 1999-2000 and spring 2000. Vegetation variables recorded were abundance, species, ground cover, density, biomass and vegetation height (Bonham, 1989). Biodiversity was estimated by using the Shannon-Wiener and Fisher (alpha) indexes which were determined with the EstimateS501 oftware (Colwell, 1997). To identify differences among sites, indexes were compared with Tukey test (a=0.05) as suggested by Ramírez (1984) using the GLM procedure, including factors and unbalanced subclasses (SAS, 1986) with the following model:
yij = µ +esp +sit+est+sit*est+est*esp+ eij
Where: yij = observed value of abundance, µ = general mean, espi = species effect, sit i = site effect, esti = season effect, sit*estij = site x season interaction, eij = random error.
We found a richness of 148 species with a total population of 9,630 individuals. Predominant vegetative communities were pine-oak forest (Pinus spp. and Quercus spp.) and grasslands. The main plant species found within the study area were Muhlenbergia spp. (zacaton), Bouvardia spp. (trompetilla), Gnaphalium americanum (gordolobo), found at a mean elevation of 2,650 m in a temperate subhumed climate C(w2)(w) according to Koppen modified by García (1989) where the mean temperature varied from 12 to 18°C. The Shannon - Wiener diversity index in the pine-oak forest was between 0.665 and 1.0 whereas the alpha diversity index between 9.927 and 24.684. In grasslands, the Shannon-Wiener index varied from 0.34 to 0.927 while the from 1.997 to 3.531. The diversity index varied from 0.34 to 1.0, representing intermediate to high diversity. Diversity index of the overall habitat was 3.87 (Shannon-Wiener), with 0.54 as an intermediate value for the habitat used by the Montezuma quail. Both indexes had a Poisson distribution and were highly sensitive to detect rare species; however, Fisher’s alpha differs in the evaluation of the size of a natural reserve and to understand the dynamic of species extinction in fragmented habitats. On the other hand, the Shannon-Wiener index is sensitive to changes in distribution and frequency of species to compare communities in an specific and uniform way (Begossi, 1996; Magnussen and Boyle, 1995). Magnussen and Boyle (1995) found that diversity indexes follow the same model with a strong linear correlation (r=0.98, P= 0<0.001) with the number of expected species. In this study, both indexes presented a high linear correlation r= 0.97, (P =0<0.001), which is confirmed by the regression coefficient (bo = -10.3128 and b1 = 7.41.
The main source of heterogeneity abundance was the species type (P<0.01). No effects were found (P>0.05) for site, season or its interaction. According to this results, an understanding of the relationships between species and site are important for the of biological diversity in the Montezuma quail’s habitat. Halffter and Ezcurra (1995) indicate that the relationship site-species is being revaluated because of the necessity to conserve natural resources in forests with grassland. It is important to emphasize that species diversity is the main parameter to measure direct or indirect effects of human activities in the ecosystems (Halffter and Ezcurra, 1995). The state of Mexico has the highest human population density of the country. Regarding biomass production, Oxalis spp., and Muhlenbergia spp. were the main plant species in the Montezuma quail’s habitat. Oxalis spp. was the main component of the diet, with a density of 7.01 bulbes/m2 and 3.54 g/m2 of biomass, covering 6.1% of the sampling site. Zacaton, serves as a nesting cover, escape, and protection, occupying 7.1%. Our results contrast to those reported by Brown (1982) in Arizona where Oxalis bulbs yielded a biomass of 2.72 g/m2 . Mean of vegetation density was 17.21 plants/m2, where Muhlenbergia spp., Eragrostis spp., Bouteloua spp., Piptochaetium spp., Stipa spp., Lycuros spp., Bromus spp., Brachiaria spp., Heteropogon spp., and Vulpia spp., were the most common grasses used by Montezuma quail. The herbaceous strata yielded a biomass of 21.37 g/m2. The arboreal strata was composed of 54.1 trees/400m2 with a mean height of 5.9m, being Quercus spp. the most common tree. This is important, because it provides refuge and resting areas for quail (Leopold and McCabe, 1957; Brown, 1978, 1982). A combination of grassland and forest land provides appropriate conditions for maintenance, reproduction and escape cover for Montezuma quail.
Management recommendations include the protection of forests, edge habitats and grasslands, where the high human population and domestic sheep overgrazing are the main treats, and agriculture and livestock production are important source of income for rural people. For these reasons, management strategies in agrosilvopastoral systems must be focused in using the natural resources in a sustainable manner.
Since world human population demands more food production and utilization of other natural resources in a sustainable bases, it is a necessity to implement conservation and management strategies for vegetative communities in an integrated silvopastoral concept. These management strategies will allow the use of forested and grassland areas for agricultural and livestock activities in an integrated fashion. Our challenge as resource managers is to look forward increasing the production efficiency of traditional systems without increasing the rate of fragmentation of Montezuma quail’s habitat.
Begossi, A. 1996. Use of ecological methods in ethnobotany: Diversity indices. Economic-Botany 50 (3): 280-289.
Bonham, Ch. D., 1989. Measurements for Terrestrial Vegetation. Ed. John Wiley & Sons. Fort Collins, Colorado, USA. 19 - 46 p.
Brown, R. L., 1982. Effects of livestock Grassing on populations of mearns quail in southeasterm Arizona. Jour Rang. Manag. 35 (4): 727 -732.
Colwell. 1999. Statistical Estimation of Species Richness and Shared Species from Samples. Department of Ecology and Evolutionary Biology, University of Connecticut.
Halffter G. y Ezcurra E. 2000. La Diversidad Biológica de Iberoamérica. ¿Qué es la Biodiversidad?. www.mesoamerica.org.mx/biodiv.htm
INEGI, 1993. Estudios del Estado de México. Instituto Nacional de Estadística, Geografía e Informática, Aguascalientes, México.
Leopold S. and McCabe A. 1957. Natural History of the Montezuma Quail in Mexico. The Condor 59(1): 3-26.
Magnusses, S. and Boyle, T.J.B. 1995. Estimating sample size for inference about the Shannon-Wiener and the Simpson indices of species diversity. Forest Ecology and Management 78:71-84.
Ramírez, B. 1984. Estudio de las zonas de rechazo del análisis de varianza y algunas pruebas de comparaciones múltiples para el caso de tres medias. Tesis de Maestría en Ciencias. Colegio de Posgraduados, Chapingo, México, 210 p
SAS INSTITUTE. 1986 SAS user’s guide. SAS INSTITUTE INC. Raleigh.NC.
Solano, A.R.1994. La Ganadería: ¿Actividad destructora del medio ambiente?. Agroforestería en las Américas. ICRAF, Danida, CATIE. No.3. 25p.
Johnny Montenegro, Sergio Abarca[71]
Key words: carbon dioxide, emissions balance, global warming, methane, nitrous oxide.
The greenhouse effect is the result of the gases accumulation in the atmosphere, mainly carbon dioxide (CO2), methane (CH4) and nitrous oxide (N2O), that they increase annually 0.5, 0.6 and 0.35% respectively. Approximately 30% of these gases take place for the change in the use of the soil. It is considered that the tropics emit of 10 to 30 million of carbon (C) year-1 as a result of decreasing of the organic matter in the soils deforested (Detwiller and Hall 1988). In this sense, the soils with pastures play a very important roll for the great extension that they cover, for the retention and reduction of the emission of C to the atmosphere (Minami et al. 1993, Fischer et al. 1994).
On the other hand, the bovine when converting material with great content of cellulose in food of high nutritive quality, they produce CH4, a gas that contributes with the global antropogenic heating. The CH4 represents nutritious energy that gets lost in gas form, instead of being become milk or meat. Fortunately it is feasible reduce the emissions in the cattle means of the application of appropriate technology (Montenegro and Abarca 2000).
The antropogenics activities and the natural ones emit similar amounts of N2O, contributing the sector agricultural until the 22% of the emissions, that it is generated like consequence of the application of nitrogenous fertilizers.
The agroforestry systems in the tropic have great potential in order to contribute in the mitigation of the problem of the Global Heating, since the combination of species C3 and C4 acts like an efficient sink of C in the silvopastoril systems (SSP). In this sense they should be evaluated in order to quantify and determine their contribution in the improvement of the atmosphere.
With the purpose of carrying out a balance of gases emissions with greenhouse effect in SSP in three several zones of life of Costa Rica, the emission of CO2 and N2O was determined in gas form, the amount of C stored in the soil and in the arboreal component was quantified, and was estimated the emission of CH4 that produce the bovine.
This investigation was carried out in three ecologic zones, Low Montano Forest (LMF), Very Humid Premontano Forest (HPF) and Humid Tropical Forest (HTF) according to Holdriedge (1996). The soil was classified like andosol with franc texture in the LMF, and like inceptisol of franc-sandy texture in the HPF and in the HTF.
The determination of the gas emission of N2O and CO2 was carried out by means of the technique of the close camera (Veldkamp 1994), locating these at random, analyzing it in a gas chromatograph Hewlett Packard 5890 previous preparation of a standard curve of calibration. In addition was determined the temperature (0-10 cm), humidity (0-10 cm), level of ammonium (N-NH4), of nitrates (N-NO3) and the compaction (0-10 cm) of the soil (Forsythe 1980). With the obtained data was calculated the factor of emission (gas amount emitted by unit of area) and the net emission (factor of emission less the natural ecosystem emission). The content of carbon in the soil (0-10 cm) and in the arboreal component was calculated according to Veldkamp (1994) and Cubero and Rojas (1999).
For purposes of this investigation more amount of jaul (Alnus acuminata) trees in the LMF and laurel (Cordia alliodora) trees in the HPF and HTF that the really present were simulated. In the three strata was considered a density of 75-trees ha-1 with an age average of 15 years and a total volume of 0.287 m3 tree-1 in the BMB and 0.392 m3 tree-1 in BPH and BHT.
The grasses kikuyu (Pennisetum clandestinum, fertilized with 485 kg of N ha-1 year-1), african star (Cynodon nlemfuensis, fertilized with 250 kg of N ha-1 year-1) and ratana (Ischaemum indicum, without fertilization), where the bovine grazing, were sampled and they were analyzed in the Laboratory of Animal Nutrition in CATIE.
The emission of methane in Jersey cows of the properties of the LMF and HPF, and bovines typical (Bos indicus) of double purpose in the HTF was considered by means of simulation. Was calculated the factor of emission and the efficiency of emission (amount of CH4 emitted by kg of produced milk). This information was proceeded to carry out the balance of emissions in equivalent units of C.
In order to compare the variables in the SSP, an ecosystem of natural forest in each ecologic zone was used, where the same determinations were made that in the SSP. Using the SAS they were carried out analysis of variance and of lineal correlation.
Emission of nitrous oxide and carbon dioxide of the soil
In the LMF the factor of emission for the N2O and the CO2 was bigger for the SSP (6.38 kg of N ha-1 year-1 and 9.06 kg of C ha-1 year-1) that the natural ecosystem (0.72 kg of N ha-1 year-1 and 2.19 kg of C ha-1 year-1). The bigger emission of the N2O is explained for the application of nitrogen fertilizer (Magalhaes et al. 1987) and the level of ammonium (N-NH4) in the soil (r2=0.77 and P=0.0439). This coincides with the informed by Davidson et al. (1996), who’s determined that the emission of this gas increased when the concentrations of N-NH4 were high. The nitrogen fertilizer applied altered the relationship C/N of the soil and this affected the populations and the activity of the denitrifies bacterias. The net emission was 5.66 kg of N ha-1 year-1 and 6.87 kg of C ha-1 year-1.
The emissions of N2O in the SSP of the HPF (3.14 kg of N ha-1 year-1) were 50% less that those of the natural forest (4.72 kg of N ha-1 year-1). The bigger levels in the natural forest were due rate of nitrogen recycled (Vitousek and Matson 1988) and to high contents of N-NO3 detected in the soil (r2=0.64, P=0.0892), which is in agreement to the results obtained by Weier et al. 1991. The pasture presented lower levels of N-NO3 that the natural forests. Emissions of CO2 obtained in the natural forest (2.51 kg C ha-1 year-1) were lesser then the SSP of the HPF (6.03 kg C ha-1 year-1), without the evaluated variables present influence in the levels of emission of this gas.
In consequence, the net emission in this SSP was -1.58 kg N ha-1 year-1 and 3.51 kg C ha-1 year-1. This demonstrates a favorable balance for the case of N2O, while the natural forest emits minor amount of C. This is in agree with informed for other investigators (Sanhueza et al. 1990, Keller et al. 1993) who’s determined bigger emissions in the natural forest that in pastures. In respect to C, this is emitted in bigger amount for covered soil for the grass due to the discharge and quick rate of growth of this specie, that occasion the deposit of important amounts of organic matter on the soil due to the grazing, which causes the mortality of superficial roots also. The decomposition of organic matter for microorganisms of the soil, which is stimulated for the amount of nitrogen in the system, explains the bigger emissions of this gas in the ecosystem of pasture.
In the SSP the HTF emissions were 2.28 kg N ha-1 year-1 and 6.17 kg C ha-1 year-1, while in the natural ecosystem, the same corresponded 1.12 and 2.18 kg N and C ha-1 year-1, respectively. In both cases the emissions were higher in the SSP due to the more amount of nitrogen in the soil, product of the recycle that they carry out the animals in grazing, and that in accordance with Velthof et al. (1996) the percentage of N2O that comes from the excretes of the animals are able to be more than the coming from the nitrogen of the fertilizer. The rate of denitrification is bigger in the recycled nitrogen, because this only affects between the 10% and 15% of the total area in grazing, then the rate of liberation of N2O is high (Ruz-Jerez et al. 1994). The emission of N2O correlated with the level of N-NH4 (r2=0.83, P=0.0214) and with those of N-NO3 (r2=0.89, P=0.0072). Davidson et al. (1996) obtained similar results. The net emission was 1.16 kg N ha-1 year-1 and 3.99 kg C ha-1 year-1.
Methane emission
High emission of CH4 ha-1 year-1 was determined in the SSP of the HPF (654 kg) followed by the LMF (360 kg) and of the HTF (183 kg). The differences in the level of emission are due to the handling of the SSP; the property located in the HPF manages more animal load (6.5 AU ha-1) than the LMF (3.9 AU ha-1) and HTF (2.2 AU ha-1).
The efficiency of emission was 19, 22 and 31 g CH4 kg-1 of milk, respectively for the SSP of the LMF, HPF and HTF. Better efficiency presented the SSP of the LMF due to a series of conditions that they favor the production of milk, such like climate, quality of the available forage and genetic quality of the animals. The efficiencies of the SSP of the LMF and HPF are similar to them reported by Montenegro and Abarca (1998), while those of the HTF could be improved by means of a better handling (nutritional and genetic) of this type of animal production system.
Carbon in the soil
The levels of carbon determined in the soil of the SSP (LMF: 41.2 t C ha-1, HPF: 20.9 t C ha-1, and HTF: 42.8 t C ha-1) was higher than that those of the ecosystem of natural forest (LMF: 35.2 t C ha-1, HPF: 15.7 t C ha-1, and HTF: 24.8 t C ha-1). This is in agreement for the results of other investigations that have demonstrated the importance of the grasses in order to capture and retain C in the soil (Minami et al. 1993, Fischer et al. 1994). This was possibly due to the great radical biomas of the grasses, being the proportion of sand present in these soil a factor that stimulates the radical development (Veldkamp 1994).
Carbon in the arboreal component
The estimated amount of fixed C ha-1 annually in the arboreal component of the SSP was bigger in the HPF ecosystems and HTF (313 kg) then the LMF (229 kg). It is due to the high rate of growth that present the laurel in these ecosystems compared to the jaul.
Emissions balance
The emissions balance of the SSP was positive, in all cases they act like nets emissors. The lesser emissions were estimated for the HTF (557 kg C ha-1 year-1) and the higher for the HPF (3376 kg of C ha-1 year-1), locating the LMF in an intermediate range (2144 kg C ha-1 year-1). The factor that affects negatively the emissions balance is the methane, since it is the gas that takes place in high amount in these production systems.
Exists influence of the nitrogen applied like fertilizer and the recycled by the animals on the amount of N2O emitted, and with the exception of the HPF the total emission was higher in the natural forest. The concentration of C in the soil was bigger in the SSP that the certain in the ecosystem of natural forest. In the three evaluated SSP the gases emissions with greenhouse effect was higher that the fixed C, this is a result mainly the emissions of CH4; however, is feasible diminish the emissions of this gas and increase the amount of fixed C, since the technology for this exists, which would affect the final balance positively obtaining a SSP more friendlier with the ambient.
Cubero, J.; Rojas, S. 1999. Fijación de carbono en plantaciones de melina (Gmelina arborea Roxb.), teca (Tectona grandis L.f:) y pochote (Bombacopsis quinata Jacq.) en los cantones de Hojancha y Nicoya, Guanacaste, Costa Rica. Tesis Lic. Ciencias forestales. Universidad Nacional. Heredia, Costa Rica.94 p.
Davidson, E.; Matson, P.; Brooks, P. 1996. Nitrous emission controls and inorganic nitrogen dynamic in fertilized tropical agricultural soils. Soil Sci. Soc. Am. J. 60:1145-1152.
Detwiller, R.; Hall, C. 1988. Tropical forest and the global carbon cycle. Science 239:42-47
Fischer, M.; Rao, I.; Ayarza, M.; Lascano, C.; Sanz, J.; Thomas, R.; Vera, R. 1994. Carbon storage by introduced deep-rooted grasses in the South American savannas. Nature (London) 371 (6494): 236 - 238.
Forsythe, W. 1980. Física de suelos. IICA, San José, Costa Rica. 212 p.
Holdriedge, L. 1996. Ecología basada en zonas de vida. IICA, San José , Costa Rica. 216 p.
Keller, M.; Veldkamp, E.; Weitz, A.; Reiners, W. 1993. Effects of pasture age on soil trace-gas emission from a deforested area of Costa Rica. Nature. 365:244-246.
Magalhaes, A.; Nelson, D.; Chalk, P. 1987. Nitrogen transformations during hydrolysis and nitrification of urea. I. Effect of soil properties and fertilizer placement. Fertilizer Research 11:161-172.
Minami, K; Goudriaan, J; Lantinga, E; Kimura, T; Baker, M. 1993. Significance of grasslands in emission and absorption of greenhouse gases. Grasslands for our world. (New Zealand) 444-450:60.
Montenegro, J; Abarca, S. 2000. Alternatives to methane emissions reduction in the livestock of Costa Rica. In Second International Conference Methane Mitigation, 18-23 June, Novosibirk, Rusia. pp. 175-182.
Montenegro, J.; Abarca, S. 1998. Estimación de la emisión de metano en la ganadería bovina de Costa Rica, 1990 y 1996. In David Conneely, Michael Gibbs, Mark Orlic (ed) Methane Measurements Workshop, Washington State University. Pullman, WA. pp. 19-29.
Ruz-Jerez, B.; White, R.; Roger, P. 1994. Long-term measurement of dinitrification in three contrasting pastures grazed by sheep. Soil Biol. Biochem. 26(1):29-39.
Sanhueza, E.; Min Hao, W.; Scharffe, D.; Donoso, L.; Crutzen, P. 1990. N2O and NO emissions from soils of the Northern Part of the Guyana Shield, Venezuela. Journal of Geophysical Research 95:22481-22488. VELDKAMP, E. 1994. Organic carbon turnover in three tropical soils under pasture after deforestation. In: Soil organic carbon dynamics in pastures established after deforestation in the humid tropics of Costa Rica. Thesis Ph. D. Wageningen University, Netherlands.
Veldkamp, E. 1994. Organic carbon turnover in three tropical soils under pasture after deforestation. In: Soil organic carbon dynamics in pastures established after deforestation in the humid tropics of Costa Rica. Thesis Ph. D. Wageningen University, Netherlands
Velthof, G.; Brader, A.; Oenema, O. 1996. Seasonal variations in nitrous oxide losses from managed grasslands in The Netherlands. Plant and Soil 181:263-274.
Vitousek, P.; Matson, P. 1988. Nitrogen transformations in a range of tropical forest soils. Soil Biol. Biogeochem. 20(3):361-367.
Weier, K.; Macrae, I.; Myers, R. 1991. Seasonal variation in denitrification in a clay soil under a cultivated crop and a permanent pasture. Soil Biol. Biochem. 23(7):629-635
Adriana Morales Trejo[72], Luis Ramírez Y Avilés[73], Juan Carlos Ku Vera[74].
Key words: botanical composition, forage availability, tree density.
In many countries of Latinamerica, the population increase, exhausted cultivable lands and wood extraction has promoted the destruction of tropical forest. This problem demand the development of sustainable systems of land use that may increase and diversify production and contribute to mantain the ecological balance.
To create a sustainable livestock production system it is necessary to know the species diversity in the ecosystems, use polycultures, wooded rangelands and use rangelands with secondary vegetation, and in this way optimize the use of water and soil nutrients to prevent the erosion with an increase in forage production (Pimentel et al., 1992).
The present work was aimed to evaluate the botanical composition of silvopastoral rangelads with secondary vegetation, the forage availability and animal intake in comparision with another system of grass monoculture.
The present work was carried out at south of Yucatan, Mexico, between 20°1’18” N and 89°5’27” W, from April 1999 to January 2000. The experiment consisted of two treatments: 1) Silvopastoral system in transition (SPS): with association of guinea grass (P. maximum) and tree species of secondary vegetation; and 2) Grass monoculture system (GMS), only with guinea grass. The trials were carried out during the three seasons of the year: dry (April to June), rainy (July to September) and late rainy (October to January). For each system were used 80 paddocks of 1000 m2, which were grazed by cows (Bos taurus x Bos indicus) in lineal rotation. A stocking rate of 1.0, 1.8 and 1.2 AU/ha was employed for dry, rainy and late rainy seasons, respectively. The occupation time in the paddocks was 16 h/day, with a 40 days rest period. Four heifers cannulated in the dorsal sac of the rumen were used to evaluate intake and botanical composition of the ingesta.
Botanical composition and forage availability in the paddocks
For each season and system, the botanical composition and forage availability was determined by the quadrant technique (Cox, 1980); paddocks was sampled one day before they were grazed. The botanical composition of the paddocks were estimated with two 100 m2 quadrants located in a randomized position into the paddocks at the SPS. All the species in every quadrant were sampled, and then, they were classified at the herbarium of the University of Yucatan. The same quadrants were used to determine the density of forage trees per hectarea. The availability of guinea grass was evaluated with 0.50 m2 quadrants; and tree foliage availability with 25 m2 quadrants, in this case, all the material considered forage, like leaves and green stems, between high of zero and 1.70 m were trimed, dried and weighed and divided into different species.
Forage intake and botanical composition of ingesta
In each season, grazing time was determined by direct observation of every cow in the paddocks during one complete day, recording the activity (grazing, ruminating, idling) at 10 minutes intervals. The following day, the rumen of the four cannulated heifers, adapted to grazing in the same experimental paddocks, were completely emptied; then heifers were taken to graze during 45 minutes. Effective grazing time was evaluated during this period. Once this period was over, the rumen was completely emptied again and ingesta was weighed, dried and used to determine the dry matter (DM) intake and botanical composition of the ingesta by histological technique (González, 1990). Finally, all the ingesta emptied at the beggining was returned to the rumen of each heifer. Total forage intake per animal per day was estimated with the average intake per hour multiplied by the total grazing time at day. To classify the plant species consumed the guinea grass, trees and herbs were taken in consideration.
Statistical analysis
A completely randomized design with a factorial arrangement of treatments was employed (factor 1: type of system; factor 2: season of the year). The Tukey test was used for means when significant differences were detected, statistical analysis of results was performed with the statistical package SPSS 9.0 for Windows.
Botanical composition and forage availability at paddocks
Plant species observed in the SPS differed along the year. 89 species were found in total, 34 of them were of the Leguminosae family, 5 of Rubiaceae, 4 of Compositae families and the rest 46 species belonged to 27 different families. Only the species Acacia farnesiana, Hybanthus yucatanensis, Leucaena leucocephala, Piscidia piscipula and Vigueira dentata were observed during all year. Forage tree density at SPS was variable, between 1850 to 3400 trees/ha. Acacia gaumerii, Bauhinia divaricata, L. leucocephala, P. piscipula and Ehretia tinifolia were the most preferred species by cows. Availability of guinea grass was significantly different (P<0.05) among systems. MCS showed higher availability of grass (3.6 vs 1.7 t DM/ha/grazing cicle) (Table 1). There were not statistical differences (P>0.05) for total availability of tree species in the SPS among seasons. MCS always showed the highest forage production along the year.
Table 1. Forage availability (t DM/grazing cicle) at SPS and MCS during three seasons of the year.
|
|
Silvopastoral system |
Monoculture system |
|
|
||||
|
Season |
Dry |
Rainy |
Late rainy |
Dry |
Rainy |
Late rainy |
S.E. |
Sig. |
|
|
|
|
|
|
|
|
|
|
|
Trees |
0.18 |
0.07 |
0.10 |
0.00 |
0.00 |
0.00 |
0.03 |
|
|
Guinea grass |
2.32 |
1.87 |
1.12 |
3.00 |
3.49 |
4.30 |
0.46 |
§ |
|
|
|
|
|
|
|
|
|
|
|
Total |
2.49 |
1.94 |
1.22 |
3.00 |
3.49 |
4.30 |
0.45 |
§ |
|
|
|
|
|
|
|
|
|
|
§ Statistically difference (P<0.05) among systems.
Forage intake and botanical composition of ingesta
Grazing time was different statistically (P<0.05) during the rainy season (6.2 h) compared to the dry and late rainy season (8.0 h). In general, there were not statistical difference (P>0.05) in dry matter intake 9.3 kg DM/cow/day among systems and seasons. Botanical composition of ruminal ingesta in SPS was different among seasons (P<0.05) and showed the following ratios 58:28:14; 72:15:13 and 43:41:16 for guinea grass:trees:herbs during dry, rainy and late rainy seasons, respectively. The tree species included in higher proportion in the ingesta were L. leucocephala and P. piscipula. At the MCS, guinea grass was the only source of forage for cows.
Botanical composition and forage availability at paddocks
Diversity of species at SPS can show that this areas (now in regeneration) were initially conformed by a high botanical diversity, and many of them were legumes and seasonal species. Nevertheless, only a few of this species has the ideal features to be considered forage species. In relation to the tree density parameter, it has to be considered that the vegetation is in a natural succesion and therefore is common to found a large variation at this area.
At SPS, forage availability showed a trend to diminish, opposite to that observed at MCS. This could be due to the development of many secondary species, wich competed for water and nutrients (Chávez et al., 1983) and probably had higher adaptation through a lower productivity (Jurado et al., 1990). Therefore, is advisable to performance regular handling to allow for the selective establishment of desirable species (Parra, 1996).
The low availability of tree foliage was due to its low density inside paddocks, this effect has been shown several times in the scientific literature where the availability varies with the sowing density and with the associated species (Vizcaíno y Palma, 1997; Jordán et al., 1998).
Forage intake and botanical composition of ingesta
The results of forage intake in this work are similar to that of Roothaert’s (1999) who evaluated a diet of P. purpureum supplemented with tree foliage in cows. Some authors have found that the tree foliage in the diet can increase the intake of the basal diet (Valdivia, 1996). Nevertheless, this is determined by several factors such as availability, fibre and nutrient contents, climatic stress, digestibility (Stobbs, 1975); and with forage trees, palatability and tannin contents are very important (Maasdorp et al., 1999).
Botanical composition of ruminal ingesta at SPS was different among seasons due to climatic conditions that affected the availability of many species (De Alba and Holechek, 1996). Maximun intake of tree foliage relative to the grass was observed during the late rainy season, when it seems the grass remained wet for a longer period of time and cows preferred to browse, similar observations have been described by Mahecha (1998). Possibly, during the late rainy season trees had a higher protein and lower fibre content, which may render them more palatable to cows. The maximum grass intake was observed during the rainy season, since in this season grass is tender, has a better quality and has a high leaf availability.
Natural SPS is comformed by a great number of species, most of them are available throughout the year and are consumed by the animals. However, forage biomass was low compared with the grass monoculture.
Cox GW (1980) Laboratory Manual of General Ecology. William C. Brown publishers, San Diego, USA. pp 41-45.
Chávez A, Soltero S, Márquez J, Villalobos C (1983) Estudio de dietas de animales en pastoreo y su aplicación en la producción de carne. Memorias del 3er. Simposium Internacional sobre Ganado Productor de Carne. Chihuahua, México.
De Alba BR, Holecheck J (1996) Identificación de pastos en la dieta de tres razas de ganado bovino en una localidad de Nuevo México. Tec Pec Mex 34:116-120.
González MH (1990) Manual de Técnica Histológica para determinar la composición de la dieta de herbívoros. Departamento de Recursos Naturales Renovables. Universidad Autónoma Agraria Antonio Narro. 15 p.
Jordan H, Traba JDL, Ruíz T, Febles G (1998) Utilización de las leguminosas para cubrir el déficit de biomasa en la seca con vacas Holstein en pastoreo. Memorias del III Taller Internacional Silvopastoril. Indio Hatuey. Cuba. pp 230-231.
Jurado P, Negrete F, Arredondo T, García R (1990) Evaluación de especies forrajeras nativas e introducidas bajo condiciones del altiplano central mexicano. Tec Pec Mex 28:40-44.
Maasdorp BV, Muchenje V, Titterton M (1999) Palatability and effect on dairy cow milk yield of dried fodder from forage trees Acacia boliviana, Calliandra calothyrsus and Leucaena leucocephala. Animal Feed Sci Tech 77:49-59.
Mahecha L (1998) Análisis de la relación planta-animal desde el punto de vista nutrición en un sistema silvopastoril de pasto estrella africana Cynodon plectostachyus, Leucaena Leucaena leucocephala y algarrobo Prosopis juliflora en el Valle del Cauca. Tesis de maestría. Escuela de Posgrado Universidad Nacional de Colombia 134p.
Parra L (1996) Control de malezas en legumbres forrajeras arbóreas. In: Leguminosas forrajeras arbóreas en la agricultura tropical. Clavero, T. editor. Venezuela pp 147-153.
Pimentel D, Stachow U, Takacs D, Brubaker HW, Dumas AR, Meaney JJ, O’neil AS, Onsi DE, Corzilius DB (1992) Conserving biological diversity in agricultural and forestry systems. BioSc 42:5.
Roothaert R (1999) Feed intake and selection of tree fodder by dairy heifers. Animal Feed Sci Tech 19:1-13.
Stobbs TH (1975) Factors limiting the nutritive value of grazed tropical pastures for beef and milk production. Tropical Grasslands 9:141-150.
Valdivia SV (1996) Efectos del follaje de Brosimum alicastrum Suv sobre el consumo, degradación ruminal de Panicum maximum Jacq y suministro de nitrogeno microbiano al duodeno en ovinos. Tesis de Maestria. Facultad de Medicina Veterinaria y Zootecnia, Universidad Autónoma de Yucatán. México 188 p.
Vizcaíno A, Palma JM (1997) Evaluacion de la produccion de biomasa de G. sepium en asociación con otros forrajes. Memorias del XXI Congreso Nacional de Buiatria. Colima México. pp 277.
David Morales[75], Christoph Kleinn[76]
Forests as multifunctional ecosystems and renewable resources at the same time have been one of the principal themes in many discussions about natural resources, their state, quality and sustainable management. However, another tree resource, the trees outside forests are less recognized, and little taken into account in larger area studies. While trees outside the forests comprise actually all trees that do not fall under the definition of forest, in Central America the trees on pasture lands (“potreros”) play a prominent role, simply because of the large area of pastures. In Costa Rica, for example, about 40% of land area is reported to be under pasture.
The trees on pasture lands have a series of ecological and economic functions which are similar to the functions of forest trees: carbon sequestration, conservation of biodiversity, soil improvement, soil erosion control, timber and fuelwood production, fodder and shade for cattle, among others.
From these general considerations one may directly conclude that trees on pasture land must play a relevant role from an ecological and from an economic perspective. While this is clearly acknowledged in general terms and on a farm basis, there are few studies which look at this topic from a large scale point of view. In the present document, some data is compiled about the relevance of trees on pasture land in Costa Rica, coming from a general review of information and literature and from preliminary results of the EU funded TROF project[77].
Cattle raising started in Costa Rica with the arrival of the Spaniards, when the first forest areas were converted into pastures and other cultivated lands, at this time for subsistance production (Montenegro and Abarca 1998). Land was considered more productive the more the vast “adversary” forest was pushed back (this custom prevailed until the 20th century when the legislation still fostered colonization of new areas). In fact, there is not too much known about cattle breeding during the early colonial years but certainly there was no rapid growth in the number of livestock. This is likely due to the natural scarcity of grasses suitable for forage and the absence of extended natural savannah lands (Montenegro and Abarca 1998). Only from the mid-20th century, the expansion of cattle breeding became significant, and from the 1950s onwards it was the most decisive driving force behind the land use and landscape development, and forest destruction. In the development of pasture land area in Costa Rica from 1961 to 1994 is shown as reported in the FAOSTAT database. Negative developments on the meat market stopped the rapid increase in the mid 1980s.
Figure 1. Development of pastrure land area in Costa rica from 1961 to 1994 (FAOSTAT database).
There are not many sources giving large scale information about quantitative and qualitative aspects of tree resources outside forests, or specifically on pastureland. Interesting baseline data would be species distribution, diameter distribution, density, basal area, volume, regeneration, functions and socio-economic attributes. At the end, of course, the actual management and options of sustainable management are the most relevant issues: trees are an important component of cultivated (non-natural) landscapes and it may be implicitly assumed that conservation of a certain tree cover on pastures is advantageous from an ecological and economic perspective.
Data can be collected either by review of existing sources, by interviews or by direct observations (inventories). From interviews with farmers, one tends to receive biased results on tree density etc., as most of them are not well aware of the quantity of trees that they have on their non-forest land. Van Leeuwen and Hofstede (1995) state that in their study in the Atlantic Region of Costa Rica it was practically impossible to obtain a good estimate of the average number of scattered trees on pastures. Corresponding interviews resulted consistently in an underestimation of the tree density. As mentioned, there is hardly any published data source available; large area forest inventories usually do not include those trees that are not in the forest - and agricultural surveys usually do not either, as the trees are not classed as an agricultural crop. In a Coseforma (1995) forest inventory trees on pasture land were also sampled wherever tree density was greater than 6 trees per hectare. Trees were tallied from a diameter at breast height of 10cm onwards (Table 1)
Van Leeuwen and Hofstede (1995) present results for study areas in the Atlantic Zone of Costa Rica. They report densities between 3 and 20 trees per hectare of pasture and emphasize the importance of the tree component by listing it as one of the three major elements of this type of land use, which are cattle, grass and trees.
Table 1: Comparison of key attributes of trees in forest and on pastureland (COSEFORMA, 1995).
|
Vegetation type |
Number of trees/ha >10cm dbh |
Basal area/ha [m2] |
Volume/ha [m3] (>=30 cm dbh) |
Number of species |
|
|
Identified |
% unknown individual trees |
||||
|
Forest |
540.07 |
40.29 |
296.12 |
145 |
15% |
|
Pastures |
11.22 |
2.39 |
23.57 |
79 |
1% |
|
Relation pastures/forest (%) |
2.07 |
5.93 |
7.95 |
54.4 |
|
Harvey and Haber (1999) carried out a study on 24 dairy farms in the Monteverde Region, covering a total of 237ha of pasture. 5,583 trees were found (190 species), when all trees with a minimum height of 2m were recorded. This results in an average tree density of 25 trees (2m and higher) per hectare of pasture, which does not include trees in forest patches and live fences that were also present. 3813 of all tallied trees (180 species) had a height of more than 5m (average 10m) and a mean diameter of 38.9cm.
In the TROF project, 5 study areas of 100 ha each were surveyed in the northwestern Province of Guanacaste. Non-forest trees with a diameter greater than 10cm or a height greater than 5m were recorded. The area that corresponded to pastures summed 157 ha and for analysis purposes a distinction was made between scattered trees and trees in lines (Table). A total number of 60 different species were found on pasture land in those study areas. The structure of the diameter distribution is interesting; the smaller diameter classes are clearly underrepresented (Figure 2). This is a clear indication of a non-sustainable tree population as regeneration is missing.
Figure 1. Diameter distribution for trees in pasture land excluding trees in lines (TROF project)
Table 2. Summary mensurational characteristics of pasture trees as found in the TROF project in the region of Cañas, Guanacaste, Costa Rica (the mean diameter is calculated as the diameter of the circle corresponding to the mean basal area)
|
|
Number of trees n/ha |
Volume m3/ha |
Basal Area m2/ha |
Mean Diameter cm |
|
Scattered trees |
10.48 |
4.617 |
1.45 |
41.91 |
|
Trees in lines |
11.29 |
3.50 |
1.05 |
34.41 |
|
TOTAL |
19.17 |
6.99 |
2.13 |
37.59 |
A comparison of these different studies is presented in Table 3.
Table 3. Comparison of the Costa Rican studies evaluated. (n.a.: not available).
|
Reference |
No. of trees |
Vol. |
Basal Area |
Number of species |
Region |
Estimated total pasture within region |
|
n/ha |
m3/ha |
m2/ha |
n |
ha |
||
|
Coseforma* (1995) |
10.10 |
23.57 |
2.33 |
76 |
Huetar Norte |
430.000 |
|
Van Leeuwen and Hofstede (1995) |
8.5 |
n.a. |
n.a. |
n.a. |
Huetar Atlántico |
386.000 |
|
Harvey and Haber 1998) |
16.08 |
n.a. |
n.a. |
n.a. |
Transición |
--- |
|
TROF project (2000) |
10.48 |
4.61 |
1.446 |
60 |
Chorotega |
773.000 |
*In this inventory, only pastures with more than 6 trees per ha were included. The figures given here must therefore be seen as lower limits.
According to the Forest Act No. 7174, all tree harvesting has to be approved by the national forestry authorities in SINAC (National System of Conservation Areas). This explicitly also extends to trees that are not in forest. However, the information available is obviously far from being complete and in addition, it is not possible to separate clearly the pasture tree harvesting within the statistics of non-forest tree harvesting.
However, one might assume that the greater part of the registered non-forest tree harvesting took place on pastures. Some data were found for the period of 1990 to 1998 (Table 4), indicating that for this relatively short period the volume share of non-forest trees was in the range of about 30-45% of the total volume harvested. Though the statistics are not complete, one may tentatively conclude that the timber harvest from natural forests is decreasing, while the harvest from forest plantations is increasing. The data available for timber harvesting of trees outside forests suggest that this quantity is less variable over the years.
Table 4. Data on timber harvest in 1990 to 1998 in Costa Rica according to forest service statistics (González y Lobo (1999).
|
Year |
Outside forests |
Forest |
Plantations |
Total |
||||
|
No of permits |
Volume (m³) |
No of permits |
Volume (m³) |
No of permits |
Volume (m³) |
No of permits |
Volume (m³) |
|
|
1990 |
3,442 |
269,928 |
384 |
393,995 |
161 |
19,667 |
3,987 |
683,590 |
|
1991 |
3,113 |
288,729 |
421 |
369,881 |
221 |
73,118 |
3,755 |
731,728 |
|
1992 |
2,948 |
225,202 |
415 |
377,778 |
308 |
50,211 |
3,671 |
653,191 |
|
1993 |
2,760 |
223,816 |
375 |
279,680 |
197 |
163,384 |
3,332 |
666,880 |
|
1994 |
|
|
|
473,691 |
|
|
|
473,691 |
|
1995 |
|
|
|
296,278 |
|
|
|
296,278 |
|
1996 |
|
|
|
221,729 |
|
|
|
221,729 |
|
1997 |
|
|
|
247,814 |
|
|
|
247,814 |
|
1998 |
|
228,982 |
|
248,362 |
|
- |
|
447,344 |
Obviously, trees on pasture land form an interesting, yet very diverse, “forestry” resource. The limited large scale data available, suggest that in the ecological and economic relevance is high. However, it appears that the tree resource on pasture land is not systematically and sustainably managed; i.e., what other authors call “living dead”. The diameter distribution of trees on pastures in several studies is alarming insofar as the regeneration is virtually completely missing. Only longer term silvicultural management and artificial establishment of tree regeneration can guarantee sustainability of the relatively high density of trees in Costarrican pastures.
Coseforma 1995. Inventario Forestal de la Región Huetar Norte. Resumen de resultados. II Edición. Basado en el informe de la consultoría GWB (Gesellschaft für Walderhaltung and Waldbewirtschftung, mbH) y en el Documento del Proyecto COSEFORMA # 43 “Evaluación de la calidad de plantaciones forestales en la Región Huetar Norte de Costa Rica. Cooperación Tecnica entre la República Federal de Alemania y la República de Costa Rica.
González F & S Lobo. 1999. Breve descripción del estado de los recursos forestales de Costa Rica. In Kleinn, C y Davis, R (eds) 1999. Memoria del Taller sobre el programa de evaluación de los recursos forestales en once países latinoamericanos. Memoria (CATIE, Turrialba, Costa Rica 17-21 Mayo, 1999. 65-81
Harvey C & W Harber 1999. Remanant trees and the consevation of biodiversity in Costa Rica Pastures. Agroforestry Systems 44(¿?):37-68.
Montenegro J & S Abarca. 1998. La ganadería en Costa Rica, tendencias y proyecciones: 1984-2005. Ministerio de Agricultura y Ganadería. Turrialba, Costa Rica.
Sistema Nacional de Areas de Conservación. 1996. Información Estadística relevante sobre el Sector Forestal de Costa Rica de 1972-1995. Documento interno, no publicado. San José, Costa Rica.
TROF Project::http://www.forst.uni-freiburg.de/TROF.
Van Leeuwen A & M. Hofstede. 1995. Forests, Trees and foarming in the Atlantic zone of Costa Rica. An evaluation of the current and future integration of trees and forest in farming systems in the Atlantic Zone of Costa Rica. Serie Técnica. Informe técnico/ CATIE: No. 257.
Mildrey Soca[78],
Leonel Simón[79], Yaima Roche[80],
Saray Sánchez[81], Anobel
Aguilar[82], Edelfidio Gómez[83]
Parasitic infestations are considered among the most important problems that affect bovine production all over the world, specially in tropical countries where pastures constitute the feeding base of ruminants, and climatic conditions favor the development of these parasitic infestations (Bianchin, 1996). Regarding this, droppings constitute real “incubators”, on the grazed pasture grounds, in which larvae are developed until they reach the infective state. According to Almería & Uriarte (1999) the particular microclimatic conditions of the droppings turn them into a reservoir for infectious larvae from which they migrate to the pasture, as the external conditions grow favorable.
On the other hand, the use of silvopastoral systems is acquiring more interest and importance for animal production in the tropics. According to Pezo & Ibrahim (1998), the use of trees in pastures is considered to have direct effects on the survival of the animals under grazing conditions because they decrease the presence of parasites and vectors that spread diseases. These systems supply edafoclimatic conditions that favor the development of a rich and varied edafic fauna, which participates actively in the decomposition of droppings (Rodríguez et al, 1998), and during this process it exerts noxious effects on the eggs and larvae of parasites. Reineck (cite by: Lobo & Veiga, 1990) states that if contaminated feces are buried by coleoptera before the parasites reach the reach the infective stage, the potential parasitism of every dropping would be reduced. The objective of this work is to evaluate the decomposition rate and its relationship to the parasitological dynamics in droppings of young bovines under silvopastoral system conditions.
The study was carried out on experimental areas of the EEPF “Indio Hatuey”. A completely randomized design was used and two treatments were studied: A. Silvopastoral systems and B. Traditional systems (pastures as monocrops). The studies were performed monthly during a year, 250droppings were used in each treatment, which were selected and marked by means of the method of systematic sampling, at the exit of the animals from the grazing areas. The studies were carried out every 24 hours during the 7 days following the bowel movement, between 8:00 and 9:00 a.m. The droppings at the beginning of the study had en average weight of 525 g, a diameter of 16 cm and depth of 3.73 cm. Every dropping occupied an average area of 0.013 m².
The following measurements were performed:
a) Temperature (of the droppings, under the droppings and of the environment).
b) Dropping weight, diameter and depth.
c) % of dry matter.
d) Biological structure of the dropping.
e) Type and height of the pasture surrounding the dropping.
f) Biological structure of the soil under the dropping (0-10 cm of depth).
The parasitological studies corresponded to the gastrointestinal nematodes and were carried out through the fecal counting of eggs (EPG) using the method of the McMaster Chamber and for larvae culture, the method of feces culture. Every dropping was evaluated in three parts: A. edge of the dropping with the soil, always pointing north, B. crust of the dropping and moist zone of the dropping. Young bovines (5/8 Holstein x 3/8 Zebu), with an average age of 6 months and a weight higher than 80 kg, grazed in the systems with an average stocking rate of 12 animals/ ha.
For the statistical analysis the STATGRAPHICS package (see 7,0) was used for the variance analysis, and the LSD for average comparison. In order to determine the relations between dropping weight and the studied variables the Method of Principal Components was used by means of the statistical package STATIFS. The proportion comparison test was performed to determine the differences regarding reduction of the fecal counting of eggs in the droppings.
When studying the decomposition dynamics of the droppings of young bovines in silvopastoral systems, we could appreciate that there is a rapid decomposition of the droppings in these systems, which showed significant differences (P<0,001), with weight losses of 45 % (227 g) after 48 hours of bowel movement (Fig. 1), reaching after 7 days a decomposition higher than 94 % (493. 27 g). While in the traditional system the droppings shower slower decompositions, mummifying from the third day, when it had lost only 18. 1 % (95 g) of its weight and increasing crust thickness to 0.4 cm.
Fig. 1. Decomposition dynamics of the droppings in systems with and without trees.
According to the results of the Principal Component Analysis, carried out in the silvopastoral system, in the first component, with maximum variability, a positive relationship was found among dropping weight (r² = 0.56), with depth (r²= 0.72), diameter (r² = 0.39) and the biological structure of the dropping (r² = 0.73), and a negative relationship was found with environment temperature (r² = 0.59). Nevertheless, it did not exert any influence on dropping weight, type and height of the pasture surrounding the dropping (r² = 0.02 and r² = 0.07, respectively).
The rapid disappearance of the droppings was much related to the presence, in these systems, of a rich and varied edafic fauna (table 1), which showed a significant increase (P< 0.01) in the number of individuals/m2 (1025) regarding the control treatment (680), and a better performance of ecological indicators. These results are similar to those found by Sánchez et al. (1998).
According to Barth et al. (1995) and Crespo & Rodríguez (2000), the presence of these fauna, specially coleoptera, worms and diptera larvae play an important role in dropping decomposition, because when they remove high amounts of excrement, they promote aeration and microbial activity.
Table 1. Behavior of the edafic macrofauna in the studied systems.
|
Phylum |
Class |
Order |
Stage |
Amount of individuals/m2 |
SE ± |
|
|
Silvopastoral systems |
Traditional systems |
|||||
|
Arthropoda |
Insecta |
Coleoptera |
Adults |
182a |
42b |
1.30 ** |
|
Larvae |
313a |
212b |
|
|||
|
Orthoptera |
Adults |
22a |
4b |
|
||
|
Dermaptera |
Adults |
30a |
4b |
|
||
|
Lepidoptera |
Larvae |
4 |
- |
|
||
|
Diptera |
Larvae |
4 |
- |
|
||
|
Myriapoda |
Diplopoda |
Adults |
112 |
217a |
|
|
|
Crustaceae |
Isopoda |
Adults |
88a |
72b |
|
|
|
Arachnida |
Araneae |
Adults |
4 |
- |
|
|
|
Annelida |
Oligochaeta |
- |
Adults |
266a |
129b |
|
|
TOTAL |
1025a |
680b |
|
|||
a, b, values with different letters differ at P< 0,05
** P<0,01
Fecal studies showed similar results to the decomposition dynamics in the silvopastoral system (Fig. 2) with a significant decrease (P< 0.01) in the fecal counting of eggs (EFG) and in the reduction of the infection percentage of the dropping, which was 59 % after 72 hours (413 EPG) and 100 % 7 days after bowel movement, neither eggs nor larvae were found in the remains of the dropping and on the surrounding soil. Nevertheless, in the traditional system, the infection percentage had only been reduced by a 29.13 % in the third day on the other hand, the permanence of the droppings in the field permitted to have fecal countings of 408 EPG (168 hours), which were higher than 200 EPG 50 days after bowel movement .
Figure. 2. Performance of egg counting (EPG).
The decrease in fecal counting and in the reduction of the infection percentage of the droppings was very related to the variability of organisms in the soil (table 1), and specially to the presence of coprophage coleoptera in the droppings, which was significantly (P< 0.01) higher in the silvopastoral system: 5.33, 8.25, 10.01, 7.50, 6.25, 5.25 and 3.75 individuals/ dropping, while for the traditional system there were: 0, 1.66, 2.34, 3.8, 4.7, 4.1 and 3.6 individuals/dropping after 24, 48, 72, 96, 120, 144 and 168 hours respectively. The role of coprophage coleoptera in the control of parasitic infections has been studied by several authors. According to Lobo and Veiga (1990), these organisms constitute natural enemies of nematodes because during the process of dropping decomposition, they exert noxious effects on the eggs and larvae, which are destroyed in the feeding process or buried deeply in the soil. They also modify the quality and quantity of the excrement, exposing eggs and larvae to the action of other predators, thus interrupting the biological cycles and limiting the access of cattle to the infective stages of these parasites.
The studies performed showed the effect of the silvopastoral system on the dropping decomposition rate, which was 33 % higher than in the traditional system after 168 hours. This increase was accompanied by a decrease of the fecal counting of eggs, showing significant differences between both systems. Within the main causes of this situation are: the significant increase of the edafic macrofauna and a higher presence of coprophage coleoptera in the droppings in silvopastoral systems as compared to the traditional system. All the above_ mentioned facts lead us to state that the silvopastoral system constitutes a natural control source of gastrointestinal parasite populations, as the dropping is their main reservoir.
Almería, Sonia & Uriarte, J. 1999. Papel de las heces bovinas como reservorio de las poblaciones larvarias de nemátodos gastrointestinales ante su migración al pasto. ITEA. Producción Animal. Vol. 95ª. No.3: 209-220
Barth, D.; Karrer, M. & Heinze-Mutz, E.M. 1995. Significance of moisture content of dung pats for colonization and degradation of cattle dung. Appl. Parasitol. 36: 11
Bianchim, I. 1996. Epidemiología dos nematódes gastrointestinais em bovinos de corte nos cerrados e o controle estratégico no Brasil. In: Padilha, Terezinha (ed). Controle dos nematodeos gastrointestinais em ruminantes. Coronel Pacheco: EMBRAPA-CNPGL. p. 113
Crespo, G. & Rodríguez, Idalmis (Eds.). 2000. Contribución al conocimiento del reciclaje de nutrientes en el sistema suelo-pasto-animal en Cuba. EDICA. La Habana, Cuba. 72p.
Lobo, J. M. & Veiga, C. M. 1990. Interés ecológico de la fauna coprófaga en pastos de uso ganadero. Ecología. No. 4, p. 313-331
Pezo, D & Ibrahim, M. 1998. Módulos de enseñanza agroforestal. Los sistemas silvopastoriles. Ediciones CATIE/GTZ. Turrialba, Costa Rica. 258 p.
Rodríguez, Idalmis; Crespo, G. & Fraga, S. 1998. Nota sobre el efecto de la acumulación de bostas vacunas en la macrofauna del suelo. Rev. Cubana Cienc. Agríc. 32: 321
Sánchez, Saray; Hernández, Marta & Simón, L. 1998. Diversidad de los organismos del suelo bajo un sistema silvopastoril. Memorias III Taller Internacional Silvopastoril. EEPF “Indio Hatuey”. Matanzas, Cuba. p. 295-297
Odalys Toral Pérez[84], Rey Machado Castro[85], Eliel González Gómez[86]
Genetic diversity of plants is a key element for agricultural development. Plant breeders, bio-technologists and researchers are interested in genetic variation of local cultivars, primitive strains and wild forms to produce new cultivated variety that are better adapted to local conditions and are higher-yielding. Nevertheless, it is very important to acquire and conserve a wide range of germplasm for their use in breeding cultivars for low input systems
Many factors, such as extension and change in the use of land, introduction of modem techniques in agriculture and the use of chemical fertilizers, pesticides and fungicides, make traditional crops become obsolete and provoke a high need of their rapid replacement by improved cultivars. On the other hand, these factors, together with the introduction of new and uniform cultivars, deforestation, the under taking of big hydraulic projects, warming and desertification, soil salinization, highway construction and urbanization may lead to a high rate of erosion of genetic resources of a country for all those economically important crops and their related ancestors. That is why, under these circumstances, there is an urgent need of trying to collect and conserve all genetic diversity still available.
The great genetic diversity existing in the heap of genes, maintains a wide potential for current and future use for the benefit of mankind. Genetic resources are irreplaceable and it is essential to carry out their collection and conservation, at the level of species, and even at the level of ecosystems, as one of the unavoidable ways for human current and future survival.
The main objective of this methodology is to show to the professionals involved in the task of collecting, characterizing and conserving plant genetic resources useful for the livestock sector, those methodological elements needed and essential for channeling this work on highly - strict scientific - technical bases.
In the family of legumes, the endemic or naturalized material shows a potentially high richness that has not been fully exploited yet in spite of the efforts developed in the labors of collection in different regions of the country. There are 433 legume species in Cuba. From these, 305 are endemic of neotropical flora and 155 are autochthonous, in addition to 15 "Cuban endemic” species from the genera Belairia (6), Notodon (4), Behaimia (2), Bembicidiom (1), Herpiza (1) and Sauvallella (1). That is why, in the case of legumes (herbaceous as well as trees and shrubs) the effort should be greater in comparison to grasses, due to the important role played by these plants in the volume and quality of animal diet and the improvement of other physical-chemical and biotic factors of the environment (widely known), including shade and incorporation of OM as litter.
Three ways will be used to collect (leguminous and non-leguminous) trees and/or shrubs:
I. Surveys addressed to producers in order to ask about ligneous species that are normally craved by the animals.
II. Direct observation of the animals during grazing or browsing in order to determine, by means of frequency studies, the most used species.
III. Use of literature for obtaining secondary information about species that have been reported in other studies.
Besides botanical identification, information should be collected about other uses given to these species at the farm and about traditional forms of agronomic management. Many of the species, in addition to being used as forage, are also used to obtain firewood, as ornamental plants, living fence posts, for human food and as medicinal plants. The information given by the producers will also allow to know simple and easy-to-implement techniques of agronomic management.
With the purpose of obtaining preliminary information on the biomass production capacity of the identified species, it is advisable to prune trees that grow naturally o that are present in the farms with this, it is possible to determine if the plants will survive the pruning and to obtain a first approximation on their production capacity, a time period long enough to pre-select the best. Nevertheless, these possibilities will be reassured by the studies performed in the arboretum.
Generally speaking, tree or shrub species should have a set of characteristics that must be taken into account during their characterization and even some of them during collection.
They may not be colonizing or invading species.
They should have few or no damaging parts (thorns).
Adequate intake and nutritive value.
Good regrowth capacity.
Good biomass production.
Good establishment and rapid growth.
Good edaphic-climatic adaptation.
Deep root system.
Acceptable foliage architecture.
They should not be heliophilous plants and must tolerate competence for light.
They must produce posts with regrowth capacity from stems and branches.
They should offer complementary productions. v The characterization of the collections of tree and shrub species will be carried out in the nursery and in the arboretum (where potentiality in determined for different usage). Nevertheless, it is very important that all the information of “passport data” from these accessions will be registered in the database of the germplasm bank (introduction book, cards, lists and computerized programme).
The process of characterization of trees and shrubs will be carried out as fast as possible. It is important to introduce a wide number of seeds from each accession (according to what has been previously described for this kind of plant), so that the number of individuals needed for this process is obtained (nursery and arboretum). v The later fate of the potentially useful material, in the case of trees, will correspond to the work flow for this type of plant. That is why, the methodology described for trees “Evaluation of trees and shrubs”, pointed out in the Agile Methodology for the Flow of Forage Germplams of the EEPF “Indio Hatuey” (1997).
Seed benefit and conservation.
After seeds are germinated in the fields used this objective, it is necessary to keep a special cultural attention on the plants, following plant health rules for the control of weeds, insects and diseases. Fertilization and correct irrigation should also be used so that healthy and well-developed plants are guaranteed in order to produce good-quality seeds.
The efficient control of the varietal purity of each planted accession is necessary.
The harvest should be performed at the optimum moment of fruit ripening. In order to achieve this, the specific characteristics of the different families and species and their behavior after ripening their seeds should be taken into consideration. Some of these characteristics are: the degree of dehiscence for a group of legumes, as well as the amount of fallen seeds and the place of disarticulation of the spikelets in the case of grasses. These aspects must be known and well-managed by the technicians and/or specialists in charge of this activity.
The pruning will be performed with the necessary care, in order to avoid the shedding of seeds from legumes and inflorescence.
Besides keeping varietal purity and plots without weeds, these aspects should be taken into account in the harvest, in order to ensure the purity and quality of the material.
The seeds from each plot, individually harvested, will be put in envelopes with their corresponding card identifying accession and harvest date. This envelopes should be securely closed in order to avoid the falling of a seed from one envelope into another, thos producing contamination.
At the place of drying and benefit, the legumes or inflorescence with the seeds will be dried in boxes covered with thin wire mesh, which guarantee aeration at a temperature of around 25-40ºC, during 3 or 4 days, in order to avoid that the rapid drying at a higher temperature damages viability.
The identification card of each accession should continue with it during the process.
The threshing and cleaning should be carried out in such a way that no mechanical damage is produced which could facilitate the entrance of disease-causing agents.
All plant residues or any impurity and the damaged or irregular seeds must be separated.
After cleaning, the seeds will be dried by means of the most adequate techniques in order to reduce moisture to 3-7 %; then they will be considered ready for packaging and storage.
Before packaging and storing the seeds, it is necessary to perform the corresponding viability or germination tests, with the objective of finding out if they possess adequate germination values (between 75 and 95 % in legumes).
The results of initial germination will be annexed to the identification card of the accession, because they are very useful for determining the time of viability. They will also be annexed to the (manual or computerized) bases of passport data. Germination will be carried out by means of periodical tests, so that the moment in which they should be regenerated again may be determined, depending on the species and conservation conditions.
Conservation of germplasm
The seeds received by any or the possible ways (regeneration or refreshing, introduction from abroad or national collection) will belong to the base and active collection kept in the chamber or in the genebank (collection in the field of those species which do not produce or produce few or no viable seeds).
The supply items stored in the base collection should have at Kats 1000 viable seeds. Yet one should not be too strict: if less than 1000 seeds are available, they can be kept under good storing conditions (-18ºC) waiting for regeneration. The active collection may be formed by a lower number of viable seeds (<300), but when it is very low (<100) it should be regenerated and afterwards kept in the chamber (2-4ºC). If this chamber is not available, both collections should be kept under short-term storing conditions (5-12ºC) and regeneration should be performed every time germination rate has been reduced to 65 % of the initial value. This value will also be taken for the base collection when it is kept in the chamber at 18ºC. If the seed has a high initial germination rate, the first test (in the base collection kept at -18ºC) may be carried out between 5 and 10 years of storing. Later tests will be performed regarding experience and existing conditions in the germplasm bank (temperature and moisture): if the conditions are optimum, every 5 to 10 years; if the conditions are bad, every 2 or 3 years.
Packaging should be carried out after seed moisture has been reduced to around 3-7 %, proceeding to introduce them right away in the container used for this purpose. This container should be perfectly hermetic in order to avoid the possible penetration of moisture. To achieve this, it is necessary to use tightly-sealed envelopes made of laminated aluminum paper, which will guarantee conservation for any of the (short, medium or long) terms.
After the envelopes are sealed and with the corresponding data (taxonomic name, name of the cultivar or variety or accession number, introduction key, harvest date, germination percentage and origin), they will be taken to the conservation chamber placing them in their corresponding (base and active) collection, in an organized way for facilitating their localization.
The field collection (genebank) should be kept free from other plants or varietal mixtures. To achieve this, periodical weeding and negative selection will be performed every time they are needed and pruning will be carried out according to the requirements of each species. For the latter, the biological cycle and the development of every accession should be taken into account. Fertilization, irrigation and treatments for the control of pests and diseases will be performed whenever needed.
In the field collection the same controls and data as for the seed collection will be kept.
Francisco Casasola, Muhammad Ibrahim, Celia Harvey, Christoph Kleinn
Inventory of forest and other plant species provides information on the richness and abundance of the species within a given plant population and it helps to determine what are the distribution of plants within the different sucessional stages (magurran, 1983). In traditional silvopastoral systems there are many multipurpose trees and shrubs that supply high value feed to animals in the dry season and farmers are keen improving the management of these systems to optimize production (casasola, 2000).
This study reports data on richness, density and distribution of successional stages of species in different category vegetations in Moropotente, Esteli, Nicaragua.
The study was conducted in the protected area of Miraflor, Moropotente in Esteli, Nicaragua which has geographical coordinates of 13o 3' 22” y 13º 7' 30” north latitude and 86o 29' 15” y 86º 29' 50” west longitude. Mean annual temperature of the area is 21.4 0C and mean annual precipitation is 874 mm which is distributed between the months November and April.
A random survey was carried with thirty cattle farmers of the area to collect bio-physical and socio-economic data on the land use and cattle production systems in the study area. Base on data collected from surveys and aerial photographs five main categories of vegetation was identified on cattle farms: 1) Pastures with low density of carbon trees (< 30 adult carbon trees/ha, PLDC); 2) Pastures with high density of carbon trees (> 30 adult carbon trees/ha, PHDC); 3) Pastures with oak trees (Quercus spp) (PO); 4) Scrub vegetation (SV); and 5) Forest (F). Four livestock farms were randomly selected to make an inventory of species in each vegetation category identified. For PLDC, PHDC and PO square sample plots of 1 ha (100 x 100 m) were marked in each of the farm selected whereas for the other classes of vegetation (SV and F) square sample plots of 0.1 ha (10 x 10 m) were used. Measurements were made on: species richness, similarity of species, plant density, and distribution of plants by sucessional stages (i.e sapling, juvenile and old trees) in the different categories of vegetation). A t test was used to determine differences in species richness between defined categories of vegetation (i.e, PLDC vs PHDC, PLDC vs PO, PHDC vs PO, SV vs F) and Jaccard coefficient of similarity was used to compare similarity of species. The density of plants between vegetation were compared using the Tukey test.
In the study area (5584 ha) 1323 plants of 60 species were collected. In table 1 is present data on species richness in the different categories of vegetation. The t test did not detect differences in species richness between the different categories of vegetation that were compared (PLDC vs PHDC, PLDC vs PO, PHDC vs PO, SV vs F) though PLDC and PHDC had greater (50%) species richness than that observed with PO.
Table 1. Species richness per category of vegetation in cattle farms of Moropotente, Nicaragua, 2000.
|
Vegetation category |
Área of sampling (ha) |
Species richness |
|
PO |
1 |
5 |
|
PLDC |
1 |
10 |
|
PHDC |
1 |
10 |
|
SV |
0.1 |
7.5 |
|
F |
0.1 |
11.5 |
Species richness = Woody and herbaceous species; PLDC = Pastures with low density of carbon trees (< 30 adult carbon trees/ha); PHDC = Pastures with high density of carbon trees (> 30 adult carbon trees/ha); PO = Pastures with oak trees (Quercus spp); SV = Scrub vegetation; and F = Forest.
An analysis of variance was not carried out to determine species richness between the five categories of vegetation because the sample plots of PLDC, PHDC and PO had different size to those of SV and F. The analysis of variance showed that the density of plants was significantly (p < 0.05) different between vegetation categories; the density of plants in SV and F were greater than that of PLDC, PHDC and PO but there were no differences between SV and F (Figure 1). In SV the main species that animals preferred were Hedyozmun mexicanum and A. pennatula (Casasola, 2000).
Figure 1. Mean density of plants/ha
in five vegetation categories in Moropotente, 2000. Lines represent the standard
deviation; bars with the same letter are not significantly different (p <
0.05).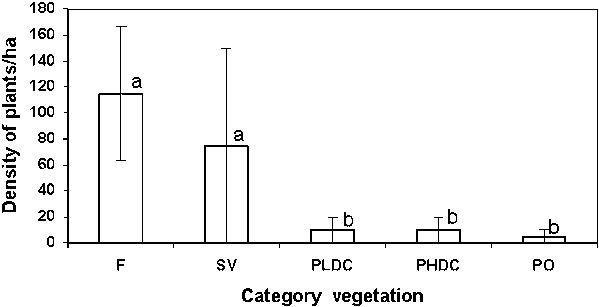
In the dry season farmers usually put the cattle in the SV because of high quality forage produced in this vegetation class and large quantities of fruits produced by A. pennatula that are consumed by animals (Jiménez, 2000; Nieto, 2000).
A low Jaccard coefficient of similarity was observed for comparisons between SV and F and between PO and PHDC indicating that there was a small number of species that were common in the vegetation compared, but a greater coefficient was observed for comparisons between PLDC and PHDC indicating that there were higher number of species that were common (Table 2).
Table 2 Comparisons of the density of species presents in different category vegetation base on the Jaccard index of similitude, Moropotente, Nicaragua, 2000.
|
Jaccard (SV vs F) |
0.19 |
|
Jaccard (PO vs PHDC) |
0.17 |
|
Jaccard (PHDC vs PLDC) |
0.43 |
|
Jaccard (PO vs PLDC) |
0.26 |
PLDC = Pastures with low density of carbon trees (< 30 adult carbon trees/ha); PHDC = Pastures with high density of carbon trees (> 30 adult carbon trees/ha); PO = Pastures with oak trees (Quercus spp);
SV = Scrub vegetation; and F = Forest.
With respect to successional stages, saplings and old trees had a greater percentage than that of juvenile stage irrespective of vegetation category. In PO and PHDC a better distribution of plants in the different successional stages was observe. In this area livestock farmers traditionally manage pastures by burning and this may explain the low number of juvenile plants found in pastures with low tree density. On the other hand, the scrub vegetation are characterized with a high density of adult plants (2649/ha) which apparently creates ecological conditions not favorable for the development of juvenile plants.
Forest and scrub vegetation had a greater specie richness than pastures with low and high density of Acacia pennatula and with Quercus spp. The categories of vegetation that had the greater similarity of species were pastures with low and high densities of A. pennatula. The density of plants varied significantly between vegetation categories observing a greater density of plants in scrub and forest vegetation.
Casasola, F. 2000. Productividad de los sistemas silvopastoriles tradicionales en Moropotente, Estelí, Nicaragua. Tesis M Sc. CATIE, Costa Rica, 99 p.
Magurran, A. 1983. Diversidad ecológica y su medición. Ediciones VEDRA. 199 p.
Jiménez, G. 2000. Potencial de árboles y arbustos forrajeros en la región Maya Tzotzil del norte de Chiapas, México. Tesis Ph. D. Mérida, Yucatán Méx. Universidad Autónoma de Yucatán. 229 p.
Nieto, H. 2000. Contribución de Acacia pennatula carbón a la productividad agroforestal sostenible de la reserva natural Miraflor-Moropotente, Esteli, Nicaragua. Tesis M Sc. CATIE, 50p.
|
[56] Instituto de Ciencia
Animal, Cuba [57] Embrapa Amazônia Oriental, CP 48, CEP 66017-970, Belém, Pará, Brazil [58] Embrapa Amazônia Oriental, CP 48, CEP 66017-970, Belém, Pará, Brazil [59] Georg-August-University of Göttingen, Kellnerweg 6, D-37077, Göttingen, Germany, [60] Georg-August-University of Göttingen, Kellnerweg 6, D-37077, Göttingen, Germany, [61] PIBIC/CNPq, scholarship, Graduate student of Agronomy FCAP,CP 917,CEP 66077-530, Belém, Pará, Brazil [62] Universidad del Valle – CIPAV, Colombia. [63] Agoforestry Systems Area, CIPAV, Colombia [64] Instituto de Ciencia Animal, P.O. Box 24, San José de las Lajas, La Habana, Cuba [65] Colegio de Postgraduados (Area de Fauna Silvestre).Carretera México-Texcoco km. 35.5, Montecillo, Méx. Tel: 0158045900 Ext. 1712 y 1716 Fax: 01-595-20279.E-mail : [email protected]. [66] Colegio de Postgraduados (Area de Fauna Silvestre).Carretera México-Texcoco km. 35.5, Montecillo, Méx. Tel: 0158045900 Ext. 1712 y 1716 Fax: 01-595-20279.E-mail : [email protected]. [67] Colegio de Postgraduados (Area de Fauna Silvestre).Carretera México-Texcoco km. 35.5, Montecillo, Méx. Tel: 0158045900 Ext. 1712 y 1716 Fax: 01-595-20279.E-mail : [email protected]. [68] Universidad Autónoma Chapingo, Departamento Forestal. [69] Colegio de Postgraduados (Area de Fauna Silvestre).Carretera México-Texcoco km. 35.5, Montecillo, Méx. Tel: 0158045900 Ext. 1712 y 1716 Fax: 01-595-20279.E-mail : [email protected]. [70] Colegio de Postgraduados (Area de Fauna Silvestre).Carretera México-Texcoco km. 35.5, Montecillo, Méx. Tel: 0158045900 Ext. 1712 y 1716 Fax: 01-595-20279.E-mail : [email protected]. [71] Ministerio de Agricultura y Ganadería, Dirección de Sanidad Animal y Protección Fitosanitaria, Costa Rica. E-mails:[email protected]; [email protected] [72] Facultad de Medicina Veterinaria y Zootecnia. Universidad Autónoma de Yucatán Apdo. Postal 4-116 Itzimná, CP 97100, Mérida, Yucatán, MÉXICO [73] Facultad de Medicina Veterinaria y Zootecnia. Universidad Autónoma de Yucatán Apdo. Postal 4-116 Itzimná, CP 97100, Mérida, Yucatán, MÉXICO [74] Facultad de Medicina Veterinaria y Zootecnia. Universidad Autónoma de Yucatán Apdo. Postal 4-116 Itzimná, CP 97100, Mérida, Yucatán, MÉXICO [75] Statistics Subunit, Tropical Agricultural Research and Higher Education Center (CATIE), Turrialba 7170, Costa Rica; E-mail: [email protected], [email protected] [76] Statistics Subunit, Tropical Agricultural Research and Higher Education Center (CATIE), Turrialba 7170, Costa Rica; E-mail: [email protected], [email protected] [77] The TROF Project (Methodology development for inventory and assessment of Tree Resources Outside the Forest – with study areas in Central America) is being carried out 1998-2001 within the INCO-DC framework of the European Commission (Project No. ERB3514PL973202). The consortium of project partners includes institutions from Costa Rica, Guatemala, Honduras, and from Austria, Germany, and The Netherlands [78] Experimental Station of Pastures and Forages “Indio Hatuey”. Matanzas, Cuba. E- mail: [email protected] [79] Experimental Station of Pastures and Forages “Indio Hatuey”. Matanzas, Cuba. E- mail: [email protected] [80] Experimental Station of Pastures and Forages “Indio Hatuey”. Matanzas, Cuba. E- mail: [email protected] [81] Experimental Station of Pastures and Forages “Indio Hatuey”. Matanzas, Cuba. E- mail: [email protected] [82] Experimental Station of Pastures and Forages “Indio Hatuey”. Matanzas, Cuba. E- mail: [email protected] [83] National Center of Agricultural Health. Havana, Cuba. [84] Experimental Station of Pastures and Forages “Indio Hatuey”, Matanzas, Cuba [85] Experimental Station of Pastures and Forages “Indio Hatuey”, Matanzas, Cuba [86] Experimental Station of Pastures and Forages “Indio Hatuey”, Matanzas, Cuba |
![]()
![]()
![]()
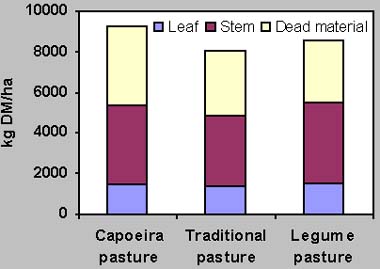{kind=link}
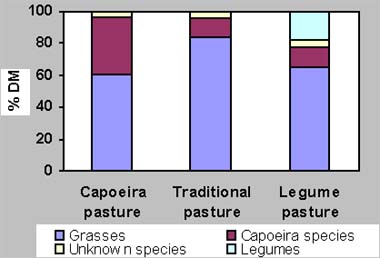{kind=link}
{kind=link}
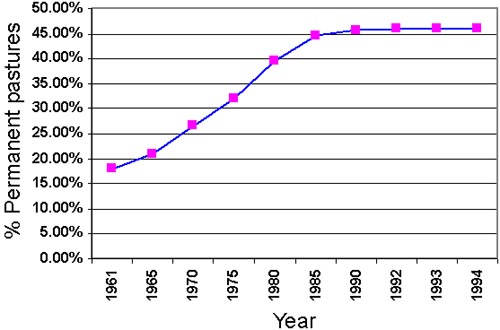{kind=link}
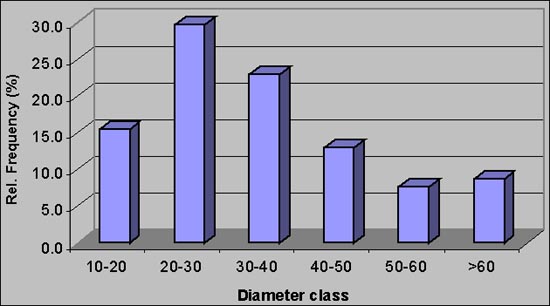{kind=link}
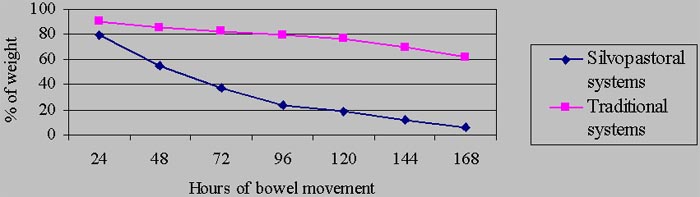{kind=link}
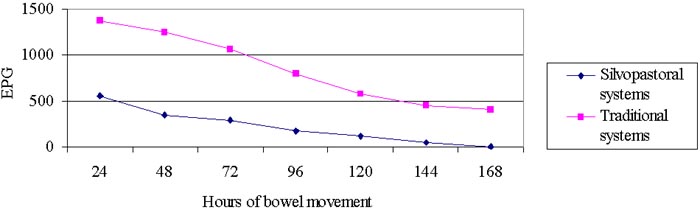{kind=link}