![]()
![]()
![]()
Salvador Hernández-Daumás[87],Graham Russell[88]
In the last decade, there has been a considerable amount of research in the field of sustainable resource management. Governmental and international agencies, universities and research institutes have strongly encouraged many scientific projects on environmentally sustainable social and economic development. Despite the importance of sustainable development, neither the amount of money invested nor the quantity of publications produced are satisfactorily reflected in either the farmers’ standards of living or in the ecological conditions of agro-ecosystems. One of the main causes of such imbalance can be found in the process of technology transfer. Because research is carried out either on experimental fields or on privately owned lands, it is difficult to extrapolate research results to small scale farming systems. This is because environmental variability and the fragmentation of land between tenants produce an agroecological mosaic in which long-term developmental programmes are difficult to introduce. Such variability must be considered in order to achieve a broader adaptability for research results. We consider such variability to be a consequence of the agricultural system - environmental supra-system interaction. Later in this note we shall address the importance of the interaction between system and supra-system.
Incorporating nitrogen fixing trees into extensive grazing systems could bring advantages such as an additional source of forage during the dry season, a high protein supplement for grass based feeds, nitrogen fixation from the atmosphere, more effective cycling of nutrients, improvement of soil structure and better microclimatic environment for grass and animals. Nevertheless, introducing high yielding germplasm (improved trees and pasture species) into low input farming systems is inevitably bound to changing management and eventually the rundown of initial production (Myers and Robbins, 1991). Beyond the need for a plan to tackle the technical and economic requirements of the new scenario, awareness of the environmental impact caused must be gathered. In this context, the study of the tree-grass-soil interaction is necessary for the understanding of experimental and on-farm validation results.
There exist a large number of aspects that result altered from mono-crop pastures when introducing trees in the system. We decided to concentrate in those we consider determinant for grazing systems with nitrogen fixing shrubs in the tropics. However, most of this material can be readily extrapolated to other silvopastoral alternatives. The first half of this section comprises some of the state-of-the-art theories on the tree-grass-soil interaction. The second half suggests some hypothesis based on our own experiences in a silvopastoral experiment carried out in Oaxaca, México.
Brachiaria decumbens withstands moderate shade and maintain high biomass yield and yet persists in densely shade environment. Whilst this grass reduces its forage production under 50% shade down to 70% of that under full sun, total leaf nitrogen increases up to twofold and the amount retained by the animal grows fivefold whereas leaf cell wall retards its lignification under shade (Norton, et al., 1991). Ludlow (1978) found that shaded plants allocate a larger proportion of their dry weight increments to leaf blades. Near infrared radiation penetrates the upper-storey canopy better than PAR (Ludlow, 1978) and the ratio of the quantum fluxes in 660 nm and 730 nm has been suggested to determine such a shift in resource allocation. Even so, this is insufficient to counteract the slower assimilation rate. However, long term yield increments under shade suggest an effect of the interaction with soil such that after the initial available nitrogen is depleted, the plant gains access to the N released by the augmented organic nitrogen mineralisation in the shade (Wilson, et al., 1990; Norton, et al., 1991).
The mineral nutrition of shaded pastures is better than full sun, particularly under limiting water and mineral conditions (Cruz, 1997). Complementarily, enhanced water use efficiency has been demonstrated to occur under shaded conditions, this reducing the evaporative demand, thus improving net photosynthesis and reducing the periods of water deficit (Wilson and Ludlow, 1991).
Cadisch and co-workers (1994) calculated the nitrogen outputs (animal, excreta and soil) of B. decumbens swards to be of about 80 kg ha-1 yr-1. Low quality litter such as that from B. decumbens, with C to N ratio as high as 117 (above ground litter), is likely to cause mineral nitrogen immobilisation, thus reducing pasture productivity in the long term (Killham, 1994). When no additions are made, the system relies only in the diazotropic bacterial fixation, organic nitrogen mineralisation and the atmospheric deposition of nitrogen. Such sources are not sufficient in mono-crop pastures, leading to soil degradation. Thus, an input of high quality litter is required (polyphenol + lignin to nitrogen ratio lower than 12.0, Handayanto et al., 1995). Associating legumes whose foliage is mainly entered to the litter pool rather than browsed is a feasible way to satisfy pasture demand. In addition, nodule mortality after pruning can be quite high and often constitutes itself sufficient reason to introduce nitrogen-fixing species in inter-cropping or during the fallow. The principle behind green-manuring with nitrogen-fixing trees prunings is that the nutrients from one species (roots) can eventually be utilised by a second species. Nevertheless, care must be taken, as frequent pruning will act against symbiotic nitrogen fixation.
Tree - pasture systems are sought as a means for diversifying pasturelands. Finding the appropriate tree and grass species is determinant for the success of the production system. We worked on a system in which prunings of tree legumes are used as green manure in improved pastures. Gliricidia sepium, Leucaena leucocephala, Lysiloma auritum and Delonix regia were used in association with B. decumbens. On doing so, several effects of the tree - grass -soil interaction had to be identified. We named that work the Silvopastoral Experiment and We pretend to use its results to illustrate some aspects of this talk.
Pasture productivity in our experiment was high and significantly different between plots, both in the dry and wet seasons. Yield being higher in the rains. At the highest tree density used (c. 800 t ha-1), grass in inter-crop yielded the same or above as in mono-crop during most of the dry season. In very dry spells, mono-crop and G. sepium associations produced less forage than the average inter-cropping. G. sepium trees could have presented strong competition for water that offset the beneficial effects observed in the rest of the mixtures; compared with the other tree species, this mixture presented a healthier root status. Every significant difference between mono-crop and the high yield mixtures disappeared in the wet season. Yielding results are in agreement with grass leaf area index measurements, which suggest a morphological response of B. decumbens to water deficit that is different in different mixtures. Treatments of poorer tree development, along with those of less tree density, produced lower grass LAI during the period of hydric deficit. No statistical differences were found in LAI when rain was abundant. Again, the reduction in grass LAI under G. sepium when compared with the control could be attributable to the extent of soil water competition. Even though both low and high density plots experienced competition between trees and pasture, there was a year round beneficial effect of the trees in the higher density treatments that offset competition and, moreover, enhanced resource availability during the dry season.
The span of the dry period should be a reference for the adequate selection of tree species in silvopastoral systems. L. leucocephala higher tree population allowed for higher grass biomass production during the period of maximum hydric stress, perhaps because of its small fine roots pool. Apparently, such a blessing in disguise entailed a shift in the partitioning of resources within the grass plant in favour of leaf lamina due to lower temperature under the canopy and less water competition. L. auritum, of similar leaf morphology to L. leucocephala, produced the same effect but with more modest results.
Of the four species established, only G. sepium maintained most of the original population. Many L. leucocephala and L. auritum trees survived but the bad shape of many individual trees resulted in very low mulch production. This put forward that limiting grass growth by planting more trees per unit area would reach a balance point between the two populations. L. leucocephala and L. auritum are higher in cell wall than G. sepium. L. leucocephala combines low carbon to nitrogen ratio with relatively high lignin and phenolics, resulting in ideal mulch for cropping systems in which there is a permanent, rather than seasonal, demand of nitrogen, such as tropical pastures. Upon the hypothesis of limited grass competitiveness under higher tree density, species of high content of nitrogen, such as L. leucocephala that did not withstand inter-cropping under the current conditions, could become an alternative sustained source of mulch. Choosing species that combine adaptation to pasture competition, high growth rate and nutrient contents a mulch decay rate that is in agreement with crop demand enables the adequate design of silvopastoral systems.
As to the nutritive value of forage grass, mono-crop was always worst, albeit not statistically different than the better treatments as in 38 day interpolations. Also, the role of the trees on preserving the good nutritional status of the grass for longer was demonstrated, and clearly, different tree species produced different results on grass crude protein and cell wall content. However, chlorophyll readings suggest that the effect of the distance from the nearest tree is of smaller scale than the layout of our experiment. It seems that factors deriving from the above ground parts of the trees, such as shade or litter fall have been restricted to the crown cover area. Tree species of well developed root system (G. sepium) would have affected the nutritional status of the soil, hence the nutritive value of the grass near the trees.
Root systems in agroforestry can be described as a spatial and temporal array of biomass where tree roots hypothetically occupy the full soil profile, whilst crop roots only reach a fraction of it. This spatial distribution is consistent with the assumption that trees improve capture or utilisation of growth resources. Such array has been observed for agroforestry systems with annual crops (Jonsson et al., 1988; Hairiah et al., 1992; Rao et al., 1993; Vidhana Arachichi and Liyanage, 1998). However, it is not clear whether the same pattern occurs in silvopastoral systems with improved perennial pastures. The risk of implementing land use technologies based on uncertain assumptions is that yields and rent can drop as resource competition overcomes the benefits of the inter-crop depleting soil and biotic resources faster than in the traditional system. A better understanding of rooting patterns in tree-grass inter-cropping systems is required for designing sustainable agroforestry systems. In our experiment B. decumbens roots were found deeper than any of the tree species roots. Woody roots were scarce and normally occurred only in the top 30 cm of the soil. The apparently poor growth of the tree roots can be explained by both the negative effect of lopping on the below-ground development, and on the effect of B. decumbens whose roots are strong competitors that prevented tree roots from capturing resources. Allelopathic effects of Brachiaria roots are no discarded, albeit the four tree species showed different levels of tolerance. Finally, rodents attack was determinant for the survival of D. regia and also affected L. leucocephala and to a lesser extent G. sepium. B. decumbens opposes a permanent pressure against trees growth, this put forward the idea of growing trees at closer spacing so as to restrict grass growth and reach full establishment of the association. Unlike the agroforestry hypothesis of the safety net from deep tree roots (Van Noordwijk et al., 1996), grass roots established a solid network for resource capture and transport, reaching more than 120cm deep in the soil. Tree roots systems in this experiment would have played the roll of weak competitors to grass roots that operates mostly in the topsoil.
Grass roots of high turnover rate, as B. decumbens roots are, cause more biomass and more nitrogen turnover occurring below than above ground. This situation put forward the hypothesis that the benefit of an extended rooting system is, at least, partially offset by the increased hazard of nitrogen leaching by deep roots. This process is opposite to the hypothesis of nitrogen sequestration sought by planting leguminous trees in inter-crop. The former necessarily carries nutrients to the leaching prone zone of soil, whereas the latter converts mineral nitrogen into organic forms, of slow release rate, that are deposited, hopefully, near the soil surface.
Although pollarding is primarily an above ground agricultural practice, it has an effect on processes at root system level. As to litter production, pruning causes fine roots to die, whether this response occurs at a rate proportional to the fraction lopped, is an issue that requires clarification. In the longer term, mulching is expected to generate a higher soil nutrient status, thus modifying the requirements of new roots. It is also unclear whether or not roots translocate nutrient before natural death, although it is likely that translocation is very limited when death occurs as a consequence of above ground pollarding.
The rationale behind this hypothesis is that this particular modality of root turnover occurs as a means of self-balance of plant parts. On the one hand, in the absence of leaves, the supply of photosynthates to maintain the root system suddenly collapses, causing the massive and more or less immediate death of fine roots. On the other hand, the demand of mineral nutrients in the canopy for the build up of new tissue is temporarily interrupted, creating a virtual surplus of nutrients at root level, minimising the need for retaining-translocating the stock.
In order to provide a minimum of nitrogen to maintain a good growth rate in the pasture (60 kg ha-1 yr-1 in amendments, Arosemena et al., 1996) different volumes of mulch should be applied depending on its nitrogen content. Nitrogen rich plant material must be produced through a fast growing tree, to allow frequent pruning of the stand, maximising light incidence at the sward level. At 4500 trees ha-1 and prunings every 100 days, it is possible to produce such volume of nitrogen. There are reports of densities higher than 4500 trees ha-1 where grass grew satisfactorily, provided a programme of pruning and fertilisation is maintained (Nygren and Cruz, 1998; Catchpoole and Blair, 1990). The ideal mulch for B decumbens silvopastures should be one of C:N ratio lower than 20 (Frankenberger and Abdelmagid, 1985) but with a great deal of its nitrogen attached to the cell wall. This would retard pro rata the release of this element, thus maintaining its supply according to crop demands. Such a paradigm should be valid for most plant nutrients.
The amount and quality of plant litter is determinant for the content of organic matter in soil and for the rate at which nutrients become available for crops and associated plant populations. Agricultural practices, particularly the pruning of leaves for mulching, causes an acceleration of nutrient cycling by preventing translocation of nutrients from old to new tissues. The quantity of nitrogen held in soil organic matter greatly exceeds annual inputs and outputs (Porter, 1975). The key issue in manipulating the recycling of nitrogen in tropical pastures is knowledge of the fractions of organic matter with different mineralisation rates and the interactions between the turnover of nutrients and the plant uptake (Powlson and Jenkinson, 1990; Heal and Harrison, 1990).
Mulch fractions decompose unevenly in different times of the year. Whilst cell contents fall to 35 and 50 % of its original value in the wet and dry season respectively, cellulose risen more than 30 and 20 % in the wet and dry seasons respectively and lignin increased 300 and 220%. Such different features stress the importance of taking into account the proper selection of species for agroforestry systems where tree prunings are to be utilised as a source of green manure. Species of rapid degradability can be as valuable for fast growing, seasonal crops, as species of slow degradability can be for permanent crops, such as tropical pastures.
The use of trees as inter-crops in pastures increases the complexity of the system and reduces its predictability even when each component is well understood. An important cause of the complexity of such systems is the management practices adopted, which differ from the ones used for sole crops. Establishment of silvopastoral demonstration plots, for local farmers to examine and discuss, requires many variables to be controlled and several years for the trees to grow and the system to be profitable. Although this long term, expensive process is indispensable, it is impossible to test every feasible change in the variables, for reasons of cost. Models can provide insights into the ability of trees to introduce nitrogen into the system and the amount of this nutrient that will be available for utilisation by the grass crop. In addition to representing the tree and grass components, the model has to take into account the interactions between them, which lead the two populations towards either resource competition or complementarity.
The silvopastoral model was designed to represent such a system by assessing the potential of prunings and dead tissues on supplying organic matter and specially nitrogen to soil. Main model components respond to the following premises: Nitrogen fixing trees can contribute to the restoration of soil fertility, although external inputs can be eventually required to amend soil fertility. Inter-specific competition for soil nitrogen is based on root biomass and root activity. Pollarding the tree canopy accelerates nitrogen cycling, such action produces both mulch and dying roots and nodules, all of them high in readily decomposable organic matter. Additionally, it allows more sun light to reach the grass canopy. Soil organic matter decomposition depends on the quality of the mulch and litter produced and on the natural abundance of soil microbial biomass.
Since the silvopastoral model was build up from two parent models of tree and grass mono-crops respectively, several mayor modifications were done in order to represent processes arising from the interaction of species. Namely light competition from tree to grass, pollarding and mulch production from trees and combined sources and flows in the soil sub-model.
Modelling should not be conceived instead but in addition to field trials, in which a cycle of modelling, parameterisation, testing and back to improving the model is a constant assignment of research teams. Field trials are necessary for parameterising the model equations so as to adequate the model to the new species and environmental conditions. Future advances in the applicability of the silvopastoral model rely on its validation with independent datasets and testing of model solutions for treatments in particular circumstances in farmers fields. Hopefully this shall lead to new improvements to the model.
Concentrating in the pure physiological aspects like those this paper deals with can result in the limited applicability of research findings. Social aspects differ between sites and so do the outcomes of interactions. Naive assumptions as to the use of communal land, water sources and forest, for example, could set in jeopardy the entire base line of the theories we aim to develop. Hardin (1968) portraits, in his ‘Tragedy of Commons’ the irremediable degradation of common resources, since the paradigm of maximisation of benefits still rules the course of actions among farmers. The following paradox is a nice example of the sentence. “Adding one more animal to my herd shall bring me, on the one hand, the profit of this animal being sold at the end of the season, and on the other hand, additional overgrazing to the grassland. Since subtracting the negative component (some degree of degradation of the communal land) from utilities causes little harm to my utilities this year, the only rational conclusion for me is to actually add that animal to my herd, and then another, and another...” till desirable grass species are gone because of overgrazing.
Moreover, this author suggest that education could hardly be put forward as a solution in the misuse of commons as it is self-eliminating, i.e. people that restrains from overexploiting a commons ‘for the general good’ will profit less, ergo, will become weaker and compete in disadvantage against the maximisation-of-benefit ones. Whether or not one is in agreement with this author, the fact is the uneasy to decipher web of interactions to be considered as the analyses of agricultural systems scales up to aboard morality.
At this point you may ask what is the relation of maximisation of benefits and interactions in silvopastoral systems. My answer is, agroforestry is a kind of juvenile science that is increasingly being requested to solve big leagues problems such as soil degradation and home economics sustainability. If agroforesters do not consider interactions between the household system and a minimum set of key components of the supra-system, we may well arrive to the disastrous conclusion that agroforestry was not as promising as our funding bodies have been told.
Tree - grass - soil interaction at low tree density (< 900 trees ha-1) may not be beneficial for the pasture in the short term, but it is not detrimental when taken over a full year. Meanwhile, trees provide shade for the animals, retain nutrients for recycling (both in plant and soil organic matter) and incorporate environmental services, such as soil stability and reduced chemical fertilisation and act as niche for birds and other fauna. Care must be taken of the nutritional status of silvopastures in the humid tropics, as it is unlikely that trees develop deep rooting system, thus high competition with grass should be expected. The main reasons for shallow rooting of trees could be: pruning, short periods of hydric stress; infertile forest soils, thus nutrient availability depending on a) mineralisation of new soil organic matter, b) biological nitrogen fixation and c) chemical fertilisation, which are inherent to the top layer of soil. Bio-physic as well as socio-economic factors will alter the suitability of different tree-grass combination and must therefore be considered when formulating new research projects.
Arosemena, E.; Pezo, D.A.; Kass, D.L. and Argel, P.J. 1996. Requerimientos externos de fósforo en pasto ratana (Ischaemum indicum (Houtt.) Merrill) y Brachiaria brizantha (A. Rich.) Stapf. Pasturas Tropicales 18:34-40.
Cadisch, G.; Schunke, R.M. and Giller, K.E. 1994. Nitrogen cycling in a pure grass pasture and a grass-legume mixture on a red latosol in Brazil. Tropical Grasslands 28:43-52.
Catchpoole, D.W. and Blair,G.J. 1990. Forage tree legumes. II Investigation of nitrogen transfer to an associated grass using a split-root technique. Australian Journal of Agricultural Research 41:531-537.
Cruz, P. 1997. Effect of shade on the growth and mineral nutrition of a C4 perennial grass under field conditions. Plant and Soil 188:227-237.
Frankenberger, W.T. and Abdelmagid, H.M. 1985. Kinetic parameters of nitrogen mineralization rates of leguminous crops incorporated into soil. Plant and Soil 87:257-271.
Hairiah, K.;Van Noordwijk, M.;Santoso, B. & Syekhfani, M. 1992. Biomass production and root distribution on eight trees and their potential for hedgerow inter-cropping on an ultisol in southern Sumatra. Agrivita 15:54-68.
Handayanto, E.; Cadish, G. and Giller, K. E. 1995. Manipulation of quality and mineralization of tropical legume tree prunings by varying nitrogen supply. Plant and Soil 176:149-160.
Harding, G. 1968. The tragedy of commons. Science 162:1243-1248.
Heal, O.W. and Harrison, A.F. 1990. Turnover of nutrients: a technological challenge. In: Nutrient Cycling in Terrestrial Ecosystems. Field Methods, Application and Interpretation. A.F. Harrison, P. Ineson and O.W. Heal (eds.). Elsevier Applied Science, London. pp. 170-178.
Jonsson, K., Fidjeland, L., Maghembe, J. A. and Högberg, P. 1988. The Vertical Distribution of Fine Roots of Five Tree Species and Maize in Morogoro, Tanzania. Agroforestry Systems 6:63-69.
Killham, K. 1994. Soil Ecology. Cambridge University Press. 242p.
Ludlow, M.M. 1978. Light Relations of Pasture Plants. In J.R. Wilson (ed), Plant Relations in Pastures. CSIRO, East Melbourne, Australia. p. 35-49.
Myers, R.J.K. and Robbins, G.B. 1991. Sustaining productive pasture in the tropics. 5. Maintaining productive sown grass pastures. Tropical Grasslands 25:104-110.
Norton, B.W.; Wilson, J.R.; Shelton, H.M. and Hill, K.D. 1991. The effect of shade on forage quality. In: H.M. Shelton and W.W. Stür (eds.). Forages for Plantation Crops. Proceedings of a Workshop held in Sanur Beach, Bali, Indonesia. 27-29 June, 1990. ACIAR Proceedings No. 32. ACIAR, Canberra, A.C.T. Australia. p.83-88.
Nygren, P. and Cruz, P. 1998. Biomass allocation and nodulation of Gliricidia sepium under two cut-and-carry forage production regimes. Agroforestry Systems 41:277-292.
Porter, L.K. 1975. Nitrogen transfer in ecosystems. In: Soil Biochemistry. Vol. 4. E.A. Paul and A.D. McLaren (eds.). Marcel Dekker, NY. pp. 1-30.
Powlson, D.S. and Jenkinson, D.S. 1990. Quantifying inputs of non-fertiliser nitrogen in agro-ecosystems. In: Nutrient Cycling in Terrestrial Ecosystems. Field Methods, Application and Interpretation. A.F. Harrison, P. Ineson and O.W. Heal (eds.). Elsevier Applied Science, London. pp. 56-78.
Rao, M.R.; Muraya, P. and Huxley, P.A. 1993. Observations of some tree root systems in agroforestry intercrop situations, and their graphical representation. Experimental Agriculture 29:183-194.
Van Noordwijk, M.; Lawson, G.; Soumaré, A.;Groot, J.J.R. and Hairiah, K. 1996. Root distribution of trees and crops: Competition and/or complementarity. In: C. Ong and P. Huxley (eds.). Tree-Crop Interactions, a Physiological Approach. CAB International. Wallingford, UK pp.319-364.
Vidhana Arachichi, L.P. and Liyanage, M. de S. 1998. Soil physical conditions and root growth in coconut plantations interplanted with nitrogen fixing trees in Sri Lanka. Agroforestry Systems 39:305-318.
Wilson, J.R. & Ludlow, M.M. 1991. The environment and potential growth of herbage under plantations. In: Shelton, H.M. and Stür, W.W. (eds.). Forages for Plantation Crops. Proceedings of a Workshop held in Sanur Beach, Bali, Indonesia. 27-29 June, 1990. ACIAR Proceedings No. 32. ACIAR, Canberra, A.C.T. Australia pp. 10-24.
Wilson, J.R.; Hill, K.; Cameron, D.M. and Shelton, H.M. 1990. The growth of Paspalum notatum under the shade of Eucalyptus grandis plantation canopy or in full sun. Tropical Grasslands 24:24-28.
P. Balandier[89], J.E. Berges[90], M. Etienne[91]
ALWAYS (Alternative Land-use With Agroforestry Systems) is a plot-based silvopastoral model derived from a European contract grouping Mediterranean and temperate countries (Auclair, 1996). It describes the biophysical functioning of a silvopastoral system at a plot and daily scale (Bergez et al., 1999). In its current version, it simulates the functioning of a "broadleaved tree - natural pasture - sheep" association. However it is possible to parametrize it for other associations if proper parameters are available.
A silvopastoral plot is defined as part of an agricultural system where trees, pasture, and animals co-exist on the same unit of land. Therefore, silvopastoral systems are complex biological systems with multiple interactions (Sibbald, 1996) and modelling the evolution of such a system needs to take these interactions into account. At the plot level, interactions occur among the different components in terms of space occupation (vertical and horizontal stratification above- and below-ground) and resource allocation (light, water, nutrients) (Anderson and Sinclair, 1993). The modelling of the temporal dynamics of this association is crucial as it determines the time course of the interactions. It must permit to test management options based on tree density, grazing calendar and harvest schedule in order to give to the farmer a rapid way to evaluate the productivity trend of his plot and to define the most efficient management strategy.
ALWAYS is defined as a management-oriented model as it specifically takes into account the management decision in the model linked with robust biophysical equations describing the system functionning (Li et al., 2000 ; Bergez et al., 2001). Such a model is between what is called a «knowledge model» which tries to summarize all the knowledge for a given process but does not integrate management and a more «economic model», which quite often uses large empirism to describe the biophysical process and evaluates management decision for their costs.
The ALWAYS model is described, calibration and evaluation processes are presented and some example of potential uses of the model and type of outputs are given.
Description of the model
A detailed description of the model structure and of the underlying growth processes is given in Bergez et al. (1999).
ALWAYS simulates the temporal biophysical behaviour of a silvopastoral system and the impact of management practices on the system. In ALWAYS, five main components are identified: climate, soil, trees, sward and animals. Climate and soil generate resources shared by the other three components (Bergez and Msika, 1996).
The spatial disaggregation scale of ALWAYS is the plot unit for animal, soil, sward and climate processes, but a tree where tree processes are concerned. Average values are given on a plot basis (mean microclimate, mean water budget). Processes are on a daily basis except for the tree growth, which is annual. However, tree leaf area development is dealt with on a daily basis to allow calculation of the daily microclimate (radiation regime, PET, wind, etc). Daily processes follow the sequence: i) microclimate is computed; ii) grazing calendar is applied; iii) sward growth is computed; iv) animal growth is computed; v) soil nitrogen and water contents are computed. At the end of the year, the trees grow.
The following management practices are implemented in ALWAYS:
- Tree management : planting density (distance between trees in the row and distance between the rows) and the weeding around the trees (how many years, weeding every year or not; radius of the weeded area) can be described.
- Pruning: either all the trees are pruned at the same height, or individual pruning can be performed using a percentage of the stem length. A regular timestep can be used to prune the trees.
- Thinning: each tree is individualised with its own growth curve according to a random variability coefficient. The percentage of trees to be felled, the year of thinning and the method of thinning are provided by the user.
- Timber: final harvest may be performed according to age, DBH or volume of the trees.
- Sward management: sward production is modified by weeding and fertilisation. The user specifies the date of fertiliser application. At present only nitrogen fertilisation is currently taken into account.
- Rearing system: four choices are proposed to manage the pasture ressource: i) do nothing; ii) harvest hay on the plot at a given date or when a certain amount of dry matter has accumulated; iii) graze the meadow; iv) adjust the stocking rate to a predetermined sward growth curve. The flock is described in categories of animals grazing (empty, pregnant or lactating ewes, lambs, rams) and number of animals per category. At present only sheep are considered.
ALWAYS was developed using an Object Oriented Design (Booch, 1994), linked with a Graphic User Interface (GUI). Several users were targeted: scientists (research tool), students and teachers (educational tool), and agricultural advisers (demonstration tool). Outputs from ALWAYS are of three types: i) physiological information such as tree growth curves, grass production and animal performance; ii) time course of the interactions between the components of the system (water stress, rainfall interception, light reduction, etc) and iii) economic information related for example to timber production, forage harvest and animal stocking density.
ALWAYS can run at three different levels: a) interactively using (or not) default values (taken from the literature and from different experiments) to test the model and create scenarios and so learn how the model works; b) automatically using parameter files and decision rules to perform automatic runs and test scenarios; c) sensitivity analysis automatically incrementing the value of a given parameter to test the sensitivity of certain parameters. For b) and c) the process is carried out from planting to tree harvest without any key input from the user. Management and decision rules are read from an ASCII file. To help understand and analyse the results, the model creates output files holding the daily or yearly results of the simulation. All these files are ASCII formated values and may be read under any spreadsheet to produce graphs and carry out analyses. Under interactive functioning, graphs are available to visualise the state of the plot at any given time.
Data used for calibration and evaluation processes
The simulations are based on real data from two contrasted agroforestry plots in France: Lamartine (Massif Central, Centre of France) corresponding to a wet mountaneaous climate and Cessous (Southern France) corresponding to a hot and dry mediterranean climate. Both stands were established in 1989. In both plots, wild cherry trees (Prunus avium L.) were planted at 200 or 400 stems ha-1. They were weeded in a 0.6 m radius around the trees for the first five years. They were pruned every year to form a 3 to 4 m high straight bole of quality wood. Both plots are grazed by sheep.
Tree and sward growths, and animal allocations have been recorded every year on these both plots.
As it is sometimes difficult to have long climatic series for the climatic data required by ALWAYS (total PAR radiation, total diffuse radiation, mean and minimum air temperatures, mean soil temperature, wind speed, relative humidity, rainfall), ALWAYS allows us to create a random frequential series of climatic years from a limited number of selected years (chosen for their representativeness) and their frequency of occurrence.
The indirect validation of the model results was performed as follow :
1- Tree and sward growths were calibrated using available data of the plots ;
2- Then, the description of the flock and the allocation dates recorded for some years in both plots were fed into the model ;
3- Then we analysed the consistency between sward availability and animal allocations according to time all along the year (i.e. the eventual forage deficits). As sward growth is linked with tree growth, if the forage production predicted by the model allows the different animal allocations recorded in the field, this leads to conclude that the different outputs of the model are consistent as a whole.
Data used to compare different management practices
Initial data from the first 10 years of growth of the site of Lamartine (see previous section) have been used to simulate and to compare different management practices : plantation densities, thinning patterns, animal rearing systems ...
Some examples of how ALWAYS can help study and simulate the management of a silvopastoral plot are given.
The results of the calibration and evaluation processes in both temperate mountaineous and mediteranean climates showed that simulated tree growth (height and DBH) is well in accordance with recorded tree growth in different plots in France (Balandier and Dupraz, 1998; Cabanettes et al., 1998). In the same way, total annual production of sward, intra-annual seasonal variations and inter-annual variations of sward growth are in good agreement with recorded data. The simulated production of forage allowed to support the different recorded animal allocations in the field, which shows that the model is consistent as a whole.
Different management practices and their consequences on tree and sward growths, animal production, rearing system were simulated and compared. In particular different planting densities, thinning strategies and animal rearing systems were evaluated and results discussed in terms of possible managements for the farmer, their constraints and advantages.
The model was parametrised and some processes have been validated. However, the model does not show the same accuracy for all the processes. For instance results showed a mild decrease in sward production when a given type of thinning was used because the canopy closure remained low. This is an interesting result but the model does not predict how the sward botanical composition will change with canopy closure, even with some slight modifications. Such changes could lead to important variations in sward quality and dry matter production. The importance of taking into account climatic variability in the prediction of tree and sward growth is underlined. ALWAYS enables us to integrate it by reducing tree growth with water deficit. The tests performed showed that this process can lead to strong variations in tree and sward production. Consequently the competition for water between tree and sward, which is only coarsely taken into account in ALWAYS, needs further investigations.
Even with some uncertainties in the processes and some inaccuracies in the results, running the model raises some questions. The importance of particular parameters or practices can be identified. Results of simulations on the type of thinning and initial tree density show that it might be worthwhile testing such thinning and planting density in the field.
Finally, ALWAYS appears sufficiently consistent to i) account for the complexity of silvopastoral systems; ii) compare different scenarios, management practices and decision rules; iii) highlight the gap of fundamental knowledge and identify the needs for future research to overcome weaknesses in the model; iv) design new experiments to test some of the results of the model based on the most interesting management scenarios. Futhermore, ALWAYS allows us to assemble different levels and types of knowledge (multidisciplinarity), and to test the consistency of the whole system and the appropriateness of mathematical models accounting for physiological processes such as sward and tree growth curves. Ecological models such as interactions among the different components of the system need some improvements to fit reality more closely. More sensitivity analyses must also be carried out to cover a wider range of conditions and to test other aspects besides the pedo-climate. So far, ALWAYS is a plot-based model and results from a single plot cannot be generalised directly on a commercial farm. Integrating silvopastoral plots into a whole farming system is a major step to aim for in the near future if the model is to fit the reality more closely and provide decision support.
The authors thank Daniel Auclair (INRA, France) and Alan Sibbald (MLURI, UK) for their help during work. Many thanks also to Myriam Dauzat (INRA, France), André Marquier and Fabrice Landré (Cemagref, France) for the field measurements. This study was partly funded by the European Commission (contract AIR-CT 92 0134).
Anderson LS and Sinclair FL (1993) Ecological interactions in agroforestry systems. Agrofor Abst 6(2): 57-91.
Auclair D (1996) Scientific and technical basis for silvopastoral systems in Europe. In: Etienne M (ed) estern European Silvopastoral Systems, pp 195-206. INRA editions, Paris, France.
Balandier P and Dupraz C (1998) The juvenile growth of widely spaced forest trees: a case study from recent temperate agroforestry plantations in France. Agrofor Syst 43 (1-3): 151-167.
Bergez JE and Msika B (1996) A silvopastoral model for the E.U. In: Etienne M (ed) Western European Silvopastoral Systems, pp 207-220. INRA editions, Paris, France.
Bergez JE, Etienne M, Balandier P (1999) ALWAYS: a plot-based silvopastoral system model. Ecol Model 115: 1-17.
Bergez JE, Debaeke P, Deumier JM, Lacroix B, Leenhardt D, Leroy P and Wallach D (2001) MODERATO: an object-oriented decision tool for designing maize irrigation schedules. Ecol Model 137 (1): 43-60.
Booch G (1994) Object-Oriented analysis and design with applications. The Benjamin/Cummings Publishing Company, Inc., 589 pp.
Cabanettes A, Auclair D and Imam W (1998) Diameter and height growth curves for widely-spaced trees in European agroforestry. Agrofor Syst 43 (1-3): 169-181.
Li M and Yost RS (2000) Management-oriented modeling : optimizing nitrogen management with artificial inelligence. Agric Sys 65: 1-27.
Sibbald AR (1996) Silvopastoral systems on temperate sown pastures: a personal perspective. In: Etienne M (ed), Western European Silvopastoral Systems, pp 23-36. INRA editions, Paris, France.
Camargo, J[92]., Ibrahim, M[93]., Finegan, B.[94]
Key words: cattle, damages, defoliation, germination, mortality, population dynamic, silvopastoral system, timber species
In Latin America, more than 90 million ha of land is under pasture, mostly as a result of forest conversion to cattle ranching (FAO 1999). Over the last years timber production from cattle farming systems, has increased because of good markets (Howard 1995). Livestock farmers in Central America region recognize the potential of valuable timber trees in their pastures to generate income to buffer low animal product prices on the national and international markets and their contribution to recuperate degraded areas (Pomareda 2000). Thus the presence of timber species in pastures is not only a good economic alternative, but also one possibility to improve environmental conditions within livestock systems.
The management natural regeneration of trees, can be a cost effective way of producing timber. The success of natural regeneration of trees in pastures depends on production and dissemination of high quality seeds, germination on compacted eroded soils, long and deep root system and possibility to recover after cattle defoliation (Archer 1995). Frequently in Costa Rica species such as Cordia alliodora, Cedrela odorata and Tabebuia rosea are very common in traditional silvopastoral systems because of good natural regeneration (Kampen 1996, Camargo et al., 2000 and Souza 2000). In spite of the abundance of natural regeneration of some trees within pastures, silvicultural management, diseases, cattle defoliation, susceptibility to weed control practices and to agressive grass species and population dynamics in the early stages of growth are generally unknown.
To contribute with strategies to management and design silvopastoral alternatives, in this work were identified timber species more common within pastures in Costa Rica and was studied the effect of ecological and socio-economic factors on natural regeneration of C. alliodora within pastures.
Increase timber price due to decrease offer from natural forest, available low cost technologies to establish trees in pastures using abundant resources as land and the acceptance of quality timber arising from cattle systems, have contributed with a reconversion and modernisation process of cattle systems (Barrios 1998). The compatibility between trees and cattle, mainly depend of their relationship and management of pasture. The cattle grazing can cause damages on trees due to cattle defoliation, treading and compaction of soils. Animals usually graze selectively which affects the species composition in pastures and competition between grasses and trees. Spatially, selectivity and treading change, grazing is heterogeneity distributed according to spatial diversity of the vegetation, topography, distances to watering places, supplementation and shade (Hart et al., 1993).
Natural regeneration is a good method to establish trees in pastures. Nevertheless there are different factors considered as critical to it such as low available of propagation source (lack seedbed trees); poor efficacy to natural spread by cattle due to low proportion of feasible seeds and concentration spread seeds in preferable places to depredates; high mortality of plants due to germination in periods with low humid (most vulnerability of seeds) (Janzen 1977); soil degradation; grass competence (Nepstad et al., 1991); and damages cause by cattle (Hatheway and Baker 1970). To identify timber species with abundant natural regeneration in pastures and to study the effect of environment conditions on them might contribute with strategies to surpass critical factors and to do compatible establishing trees and grazing within cattle systems.
Most farming systems in Central America include some agroforestry practices that contribute partially to their economic and biotic sustainability. Agrosilvopastoral systems provide forage, shade for animals and other benefits. Valuable timber species, such as C. alliodora (laurel), C. odorata (Cedar), Pithecellobium saman and Albiza spp., retained at low tree densities (4 to 15 trees ha-1), are common in the tropical lowland pastures (Barrios et al., 1999; Camargo et al., 2000, Souza et al., 2000) and in the highland regions, where intensive dairying is practised, there are good examples of planted and natural regeneration of Alnus acuminata in pastures (Russo 1990) and Cupressus lusitanica wind breaks (Harvey and Haber 1999).
Two studies conducted within farms in low lands of Costa Rica, show 25 timber species with natural regeneration in pastures of the humid (La Fortuna and Guápiles) and sub-humid zone (Esparza); where C. alliodora resulted most common (Table 1).
Table 1. Relative frequency of farms with natural regeneration of timber species within pastures in Costa Rica
|
SPECIE |
RELATIVE FREQUENCY (%) |
n |
ZONE |
|
Cordia alliodora |
100 |
60 |
Guápiles and Esparza |
|
Cordia alliodora |
100 |
35 |
La Fortuna |
|
Cedrela odorata |
94 |
35 |
La Fortuna |
|
Cedrela odorata |
26 |
30 |
Guápiles |
|
Terminalia oblonga |
54 |
35 |
San Carlos |
|
Terminalia oblonga |
10 |
30 |
Guálipes |
|
Tabebuia rosea |
30 |
30 |
Esparza |
|
Guazuma ulmifolia |
43 |
30 |
Esparza |
|
Pentaclethra macroloba |
23 |
35 |
La Fortuna |
|
Pentaclethra macroloba |
16 |
30 |
Guápiles |
|
Tabebuia ochracea |
17 |
30 |
Esparza |
|
Tabebuia ochracea |
11 |
35 |
La Fortuna |
|
Cochlospermun vitifolium |
17 |
30 |
Esparza |
|
Zanthoxylum belizense |
14 |
35 |
La Fortuna |
|
EnteroLlobium cyclocarpum |
13 |
30 |
Esparza |
|
Brossimun allicastrum |
11 |
35 |
La Fortuna |
|
Brossimun allicastrum |
10 |
30 |
Esparza |
|
Carapa guianensis |
11 |
35 |
La Fortuna |
|
Ceiba pentandra |
8 |
35 |
La Fortuna |
Source: data from Guápiles and Esparza Camargo 1999; and data from La Fortuna Souza et., 2000.
Materials and Methods
Zone of study
This study was conduced Costa Rica, using 60 sites chosen randomise within two contrasting ecological zones. The humid zone of Guapiles, with mean rainfall annual of 4000 mm and mean temperature of 24°C; and the sub-humid zone of Esparza, with mean rainfall and temperature annual of 2000mm and 24°C respectively. Simultaneously, four experiments were conduced. Two in one farm localised in Guápiles, with acid soils (pH 5.6), deep, well drain, with a resistance of penetration of 2.1 Kg m-2 and plain slope (3%). The other two experiments in one farm in Esparza with neuter soils (pH 6), well drain, moderately deep, with a resistance of penetration of 2.1 Kg m-2 and steed slope (30%).
Evaluation of natural regeneration and data analysis
To identify the patterns of natural regeneration of laurel, temporal plots were established in a nested sampling design and was accounted population in different growth stages. Seedlings (0.1 m (height < 0.3 m in plots with 50 m2), sapling (0.3 m (height <1.5 m in plots with 200 m2), young trees (1.5 m (height ³1.5 m and dhb < 5 cm in plots with 450 m2) and old trees (dhb > 5 cm in plots with 700 m2). Also, was registered information on soils, topography, vegetal composition, abundance of fruits and timber trees associated with laurel and with interviews to farmers were evaluated socio-economic aspects as dynamic land use and management farm.
Was done one analysis of principal components and more weight variables were extracted of them to adjust multiple regression models to each growth stage of laurel. Also, these variables were used in cluster analysis to classified sites according their characteristics. Next, a discriminate canonical analysis determined variables with most influence on cluster separation.
To determine the quality of laurel in different growth stages of the natural regeneration, variables as damages by cattle defoliation, burn by herbicides, cut stem, dead individuals, diseases and shape stem (only in young and mature trees), were evaluated in the different stages of natural regeneration of laurel . Relative and absolute frequency of damages, mortality and good shape in four stages of natural regeneration of laurel was calculated. Also the hypothesis of independence of these variables with respect site was tested with contingence analysis and simple correlation.
The population dynamics of laurel (Cordia alliodora) in the early seedling phase of establishment was studied within pastures currently used by farmers in two zones, Brachiaria (Brachiaria decumbens) and jaragua (Hypaharrenia rufa) in Esparza; and estrella (Cynodon nlemfuensis) and retana (Iscahemun ciliare) in Guápiles. Measurements were taken on germination and mortality during one cycle of 80 days after seed was sown. The experimental design was randomise blocks with split plots and five replications. Two treatments were made for each grass, with and without grass cover. In order to analyse data from experiment variance analysis was done.
Abundance and patters of natural regeneration
The adjusted regression models explained 58% of variability for seedling, 77% for sapling, 65% for young trees and 65% for trees. The soil type, neighbouring vegetation, management of pastures and previous land use were variables, which explained variations of seedling and tree populations. Soil variables were more important for seedling and sapling stages whereas socio-economic variables had more weight for mature stages of laurel populations .
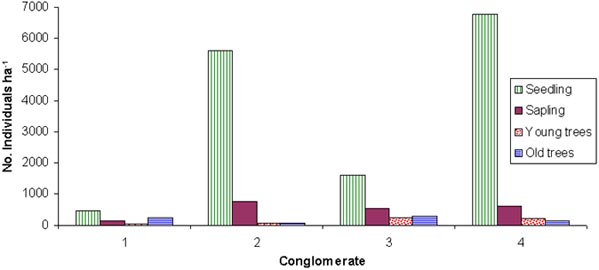
The cluster analysis identified four groups or conglomerates with different patterns of natural regeneration (two by zone). Discriminate analysis showed that the variables with more weight to separated conglomerates, were soil characteristics, represented by canonical variable 1 (with better conditions to conglomerates 2 and 4 in Esparza); and economic dependence of the farm, management of pastures, previous use of land and practices to establishment of pastures, regard canonical variable 2 (with better result to conglomerates 2 in Esparza and 3 in Guápiles) (Figure 1). Natural regeneration of laurel showed the better behaviour within conglomerate 3 (Figure 2).
Figure 1. Relation between conglomerates and canonical variables 1 and 2. (Numbers represents conglomerates, 1 and 3 located in Guápiles; and 2 and 4 in Esparza). Costa Rica
Figure 2. Natural regeneration of laurel by conglomerates. Guápiles and Esparza, Costa Rica
One balance between seedling and young trees (% young trees regard actual population of seedlings), showed that in Guápiles natural regeneration is significantly (P< 0.001) more effective than Esparza (31% vs. 8%, respectively). The same tendency was observed between conglomerates where the 3 was significantly (P< 0.001) the most effective with 37%.
Quality of natural regeneration
The results showed that saplings were more susceptible to damage with 43% of damages, while old trees had 13%. Also the presence of dead individuals was higher with saplings (Table 2).
Table 2. Damages and dead individuals in natural regeneration of laurel (Cordia alliodora). Guápiles y Esparza, Costa Rica
|
STAGE OF GROWTH |
POPULATION WITH DAMAGES (%) |
DEAD INDIVIDUALS (%) |
||
|
GUAPILES |
ESPARZA |
GUAPILES |
ESPARZA |
|
|
Seedling |
26a |
12.5b |
0a |
0.8b |
|
Sapling |
50a |
39b |
1a |
3b |
|
Young tree |
18a |
22a |
0a |
1.5b |
|
Old tree |
16a |
3.4b |
0.7a |
4.5b |
Different letters between zone are significance differences (p<0.05)
In terms of quality due to shape, old trees had better growth forms (P<0.05) in Guápiles than in Esparza (42 vs. 27%) (P<0.05). Although, young trees with good shape do not showed significance differences between zones, were more abundant in Guapiles (73 vs. 60%). Number of individuals with good shape had high correlation (P<0.01) with individuals without damages, sites with practices of silvicultural management and commercial used timber; while intensive control weed and high cow density showed unfavourable effect (P<0.05).
Populations dynamics in the early seedling phase of establishment
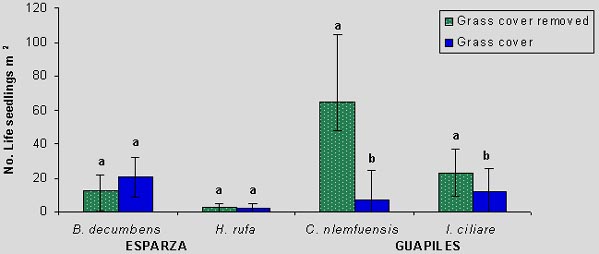
In Guápiles, the germination of laurel was higher in C. nlemfuensis swards when grass cover was removed (59 vs. 11%), while seedling mortality was in greater when laurel was seeded with grass cover (30 vs. 69%). Within I. Ciliare swards, germination was higher with grass cover seeding (24 vs. 50%), but the mortality was higher in this treatment (85 vs. 42%). In Esparza, germination of laurel was higher and mortality lower with seeding with grass cover of B. decumbens (28 vs. 21%; 55 vs. 63% respectively). In H. rufa the germination was higher with grass cover (8 vs. 6%) but mortality was greater in this treatment (83 vs. 76%). Finishing cycle of 80 days was determined the average density (individuals m-2) of seedling of laurel mean number seedlings of laurel living m-2. Within brachiaria and jaragua they were higher with grass cover, although there were not significative differences (P>0.01). In estrella the seedlings living were significantly (P<0.05) higher with grass cover while in retana there were most living with grass cover removed (Figure 3).
Models show first growth stages of laurel affected by specific characteristics of site (mainly soils and moisture), due to in this phase individuals have more susceptibility to environmental conditions (Janzen 1977). Thus variables such as the shade and soil fertility proportioned by associate vegetation could contribute to improve environmental conditions to seedling and sapling guarantying their growth (Uhl and Jordan 1984). Natural regeneration depend of the management practices intensity and trees are the result of the different treatments or conditions through the time (Concklin 1987). Young and old trees of laurel have a response in the models with more weight to socio-economic variables. In this case the time has a important role, because individuals have accumulated the effect of management and they are the final result.
The greater susceptibility to damages of sapling laurel can have relation with their size, because they are reached easily for the cattle and weed control, similar results was founded with sapling of Pithecellobium and Enterolobium within pastures (Hatheway and Baker 1970). The seedling are less defoliated by cattle, because their size is smaller than pastures. However, they could have important damages by treading (Hart and Norton 1988). Young trees are most stables and they have fewer problems, because they have surpassed several ecology and anthropoid obstacles. However, they are defoliates by cattle and cut by farmers due to their size. Young trees are very important to sure natural regeneration, because if they have gotten survivor will be old trees (Archer 1995). Old trees have fewer damages, although when they present problems these are sever. Theirs damages have relation with the more time of exposition to different factors. The presence of more dead individuals in Esparza, has relation with intensive weed control and burn practices, that additionally cause the death of several species and deteriorate of environmental conditions (Brinkmann and Viera 1971; Lal 1987). The good shape of the trees has relation with silvicultral practices and commercial activity to sell timber, because the market conditions require specific characteristics of the stem, only got them with adequate management of trees in the pastures.
Species as laurel has soft seeds and they are easily spread by wind, removed by water and lost viability with extreme temperatures (Greaves and Mc Carter 1990; Boshier and Lamp 1997). Is possible so that the germination in Esparza was affected by rainfall, high temperatures and slope. Under Brachiaria its mode of growing improved conditions to germination and decrease mortality with shade and moisture, factors that have showed to help the natural regeneration of laurel (Butterfield 1995). Some tree species have problems in their establishment when there is enough foliage and roots, due to competence by factors productions and their growing decrease (Aide and Cavalier 1993). In Guapiles when the grass cover do not removed within estrella with high density of stem and foliage, there was high moisture and the seeds do not reach the soils, moreover the conditions were very wet and many seedlings died. In retana grass there was high mortality possibility due to alelophatic effect on seedlings, registered in studies to the same zone (Arosemena 1990).
The soil characteristics and the neighbouring vegetation were the biophysical variables more important for explaining natural regeneration of laurel. The socio-economic variables were important in all models including the management of the pastures and the last use systems. Theses also represent the decision of farmers to permit the trees within cattle farm. In Guápiles and Esparza, there are different patterns of natural regeneration of laurel with influence of particularities of each zone.
Damages in natural regeneration of laurel differ according to stage of growth and the saplings are the more susceptible. The farmers have been the main cause of damages of trees within pastures due to weed control and lack management silvicultural practices.
The amount showed a tendency of better conditions in the humid zone (estrella) and both zones there was survivor within improving pastures. Three factors have special influence on the amount of seedling living finishing the evaluation cycle, the pasture and its growing habit the moisture conditions and cover presence.
Natural regeneration of trees in pastures more abundant and individuals have less damages by cattle defoliation when there is more diversity of others timber and fruits species. Therefore, is important to maintain these species associates. High intensity of manual weed control, herbicide and burn, has a harmful effect on natural regeneration of trees. However, these can be management by farmers reducing intensity, applying them on specifically on weeds or applying by lanes. Burns should not be used never because they cause severe damages.
Usually the farmers have calves or light animals isolates or in different lots. This situation could be a good opportunity to begin a natural regeneration process, because the cattle have most severe effect through of treading on first growth stage. Thus weight animals would be in lots with young and old trees, where damages by treading are less important.
There are general conditions with effects on natural regeneration of trees in pastures, nevertheless, the management should be according to dynamic of each growth stage. For seedling management practices should maintain moderate shade and moisture, soil fertility and grazing with light animals. For sapling and young trees, grazing regulation, weed control practices located, presence of fruits trees species that are ate by cattle, are necessaries. For young and old trees, all practices mentioned are very important because they reveal management process, additionally silvicultural practices such as thinning and pruning improve their quality and to reduce problems with diseases and parasites.
Within improving pastures there are good results to natural regeneration, further their quality nutritional is one advantage. However, is necessary to know growth patter because these could generate competence and to suppress the trees. So is important to remove cover of grass when growth is much aggressive.
There is a strong correlations between economic dependence of farm and abundant natural regeneration, showing that have had one decision to maintain trees in pastures. These farmers could be candidates to try strategies of natural regeneration management.
Some estimations show that is possible to have incomes by U$ 3000 ha year -1 or more (depending timber price) equivalent to one timber volume of 52 m-3 ha-1 arise from 54 trees. That Beginning with 265 young trees ha-1 of laurel (Average density registered in the conglomerate 3) and if practices recommended previously are conducted.
Aide, T. and Cavalier, J.1993. Barriers to tropical forest restoration in the Sierra Nevada de Santa Marta, Colombia. Restoration Ecology: 2:219-229.
Archer, S. 1995. Hervibore mediation of grass-woody Plants interaction. Harry Stobbs Memorial Lecture. Tropical Grassland. 29 . 218-235
Arosemena, E. 1990. Determinación de mecanismos de interferencia por alelopatía y requerimientos externos e internos de fósforo en pasto ratana (Ischaemun indicum (Houttt) Merrill). Tesis Msc. CATIE. Turrialba , Costa Rica. 124p
Barrios, C., Beer, J. and Ibrahim M., 1999. Pastoreo regulado y bostas del ganado para la protección de plántulas de Pithecolobium saman en potreros. Agroforestería en las Américas, vol. 6, no. 23, pp. 63 - 68.
Barrios, C. 1998. Pastoreo regulado y bostas del ganado como herramientas forestales para la protección de arbolitos en potreros. Tesis Msc. CATIE, Turrialba, Costa rica. 93p
Boshier, A. y Lamp. T. 1997. Cordia alliodora genética y mejoramiento de árboles. Oxford forestry institute Department of plant sciences. University of Oxford. Tropical forestry papers.
Brinkmann, W. and Viera, A. 1971. The effect of burning on germination of seeds at different soil depth of various tropical tree species. Turrialba 21: 77-82
Butterfield, R. 1995. Desarrollo de especies forestales en tierras bajas húmedas de Costa Rica. CATIE. Serie Técnica. Informe Técnico No. 260 41p.
Camargo, J. 1999. Factores ecológicos y socioeconómicos que influyen en la regeneración natural de Cordia alliodora en sistemas silvopastoriles del trópico húmedo y subhúmedo de Costa Rica. Tesis Msc. CATIE. Turrialba , Costa Rica. 127p
Camargo, C., Ibrahim, M., Somarriba, E., Finegan, B. and Current, D., 2000. Factores ecológicos y socioeconómicos que influyen en la regeneración natural de laurel (Cordia alliodora) en sistemas silvopastoriles del trópico húmedo y sub-húmedo de Costa Rica. Agroforestería en las Américas, vol 7 no. 26: 46 - 52.
Concklin, N. 1987. The potencial nutritional value to cattle to some tropcal browse species for Guanacaste, Costa Rica. Thesis PhD. Cornell University. 124p.
FAO (Food and Agriculture Organisation of the United Nations), 1999. Production yearbook, FAO, Rome.
Greaves, A., McCarter, P. 1990. Cordia alliodora, A promising tree for tropical agroforestry. Tropical Forestry Papers No. 22. 37p
Hart., R. and Norton, B. 1988. Grazing management and vegetation responce. in handbook of vegetation science. vegetation science application for rangelands analisys and management. Paul Tueller (ed). Kluwer Academic Publisher. Dordretch, Netherlands. Pp 493-525.
Hart., R., Bissio, H., Samuel, M. and Waggoner, J. 1993. Grazing systems, pasture size and cattle grazing behaviour, distribution and gains. Journal for range management, 46 (1).
Harvey, C. and Haber W., 1999. Remnant trees and the conservation of biodiversity in Costa Rican pastures. Agroforestry Systems, vol. 44, pp. 37 - 68.
Hatheway, W and Baker, H. 1970. Reproductive strategies in Pithecellobium and Enterolobium. Further information. Evolution (24). 253-254
Howard, A. 1995. Price trends for stumpage and selected agricultural products in Costa Rica. Forest Ecology and Management (26) 101-110
Janzen, D. 1977. Intensity of predation on Pithecelobium saman (leguminoseae) seed by Merobruchus colombinus and Stator limbatus (Bruchidae) in Costa Rican deciduos forest. Tropical ecology (18): 162-176.
Kampen, P. 1996. Trees in grassland. The influence of the trees on grass production within sylvopastoral systems of the atlantic zone of Costa Rica. Proyecto REPOSA (CATIE, MAG Wageningen University). Report No. 104. Field report No. 149. 33p
Lal, R. 1987. Tropical ecology and physical edaphology. John Wiley y Sons. New York. EUA.328p
Nepstad, D.; Uhl, C. y Serrao, E. 1991. Recuperation of degraded amazonian landscape: forest recovery and agricultural restoration. Ambio(6):248-255.
Pomareda, C., 2000. Perspectivas en los mercados y oportunidades para la inversión en ganadería. In: Pomareda C. and Steinfeld H. (eds) Intensificación de la Ganadería en Centroamérica: Beneficios Económicos y Ambientales. pp 55-72. CATIE-FAO-SIDE, Costa Rica.
Russo, R.O. 1990. Evaluating Alnus acuminata as a component in agroforestry systems. Agroforestry Systems 10, pp. 241-252
Souza de Abreu., Ibrahim, M., Harvey, C. and Jiménez, F., 2000. Caracterización del componente arbóreo en los sistemas ganaderos de La Fortuna de San Carlos, Costa Rica. Agroforestería en las Américas, vol 7 no 26: 53 - 56.
Uhl, C.; Jordan, C. 1984. Succesison and nutrient dyanmics following forest cutting and burning in Amazonía. Ecology, 65(5): 1476-1490.
|
[87] EL Colegio de la
Frontera Sur, Unidad Villahermosa, Mexico [88] Institute of Ecology and Resource Management, The University of Edinburgh, Scotland. [89] CEMAGREF, Groupement de Clermont-Ferrand, Team of Applied Ecology of Woodlands, 24 av. des Landais, BP 50085, F-63172 Aubière Cedex, France, tel. + 33 4 73 44 06 23, Fax + 33 4 73 44 06 98, Email: [email protected] [90] INRA, Station d'agronomie, BP 27, F-31326 Castanet Tolosan cedex, France [91] INRA, Unité d’Ecodeveloppement, Site Agroparc, F-84914 Avignon Cedex 9, France [92] Professor Environmental Sciences Faculty, Universidad Tecnológica de Pereira, Colombia e-mail: [email protected] [93] Research-professor, Watersheds and Agroforestry Systems, CATIE, Turrialba, Costa Rica. e-mail: [email protected], [email protected] [94] Research-professor, Watersheds and Agroforestry Systems, CATIE, Turrialba, Costa Rica. e-mail: [email protected], [email protected] |
![]()
![]()
![]()
{kind=link}
{kind=link}