![]()
![]()
![]()
Camargo, J.[95], Ibrahim, M.[96]
Key words: germination, mortality, population dynamic, silvopastoral system
Frequently in Costa Rica the laurel (Cordia alliodora) appears within grassland due to availability to natural regeneration (Kampen 1996). Its range of adaptation is wide, however it grows better under humid conditions, 1500 to 3000mm of mean annual rainfall, temperatures between 18 and 25 oC and soils with good physical properties and moderate fertility (Greaves and McCarter 1990). The laurel timber has a low price, but the amount of trees profited in pastures and the preference of them by sawmill do laurel promise specie to silvopastoral systems.
Lately CIAT has chosen improving grass species as Brachiarias to recuperate degrade pastures, however these has showed negative effects to establishment trees in grassland (Larb et al 1998). The population dynamic of laurel within pastures is unknown and the study about its early stages growing could to contribute to develop strategies to management and design silvopastoral alternatives.
The objective of this work was to study the populations dynamic in the early phase of establishment of laurel seedling in four different pastures (two improving and two do not improving) in the humid and sub-humid tropic of Costa Rica
Four experiments were conduced between May and August of 1999. Two in one farm localized in the humid zone of Guápiles, with mean annual rainfall of 4000mm, mean temperature 25°C and acid soils (pH 5.6), deep, well drain, with a resistance of penetration of 2.1 Kg m2 and plain slope (3%). The other two experiments in one farm in sub-humid zone of Esparza with mean annual rainfall of 2000mm, mean temperature of 27°C and neuter soils (pH 6), well drain, moderately deep, with a resistance of penetration of 2.1 Kg m2 and steed slope (30%).
The population dynamics of laurel (Cordia alliodora) in the early seedling phase of establishment was studied within pastures currently used by farmers in two zones, Brachiaria (Brachiaria decumbens) and jaragua (Hypaharrenia rufa) in Esparza; and estrella (Cynodon nlemfuensis) and retana (Iscahemun ciliare) in Guápiles. During reproductive stage, the amount of laurel seeds fall to soil in a perimeter of 10m around stem is of 280 seeds m2 (Boshier and Lamp 1997) and these experiments were used 160 seeds m2, because the seeds had been selectioned and they had germination percent between 75 and 85%.
Measurements were taken on germination and mortality during one cycle of 80 days after seed was sown. The experimental design was randomize blocks with split plots and five replicates. Two treatments were made for each grass, with and without grass cover. Variance analysis was done using 5% as significance level and when was found significance differences, test Tukey was used to compare means.
Germination and mortality and their dynamics in the time
In the humid zone of Guápiles, the germination of laurel was higher in C. nlemfuensis swards when grass cover was removed (59 vs 11%), while seedling mortality was in greater when laurel was seeded with grass cover (30 Vs 69%). Within I. Ciliare swards, germination was higher with grass cover seeding (24 Vs 50%), but the mortality was higher in this treatment (85 Vs 42%) (figures 1 and 2).
In the sub-humid zone of Esparza, germination of laurel was higher and mortality lower with seeding with grass cover of B. decumbens (28 Vs 21%; 55 Vs 63% respectively). In H. rufa the germination was higher with grass cover (8 Vs 6%) but mortality was greater in this treatment (83 Vs 76%) (figure 3 and 4).
Relationship between mortality and germination of laurel
Finishing cycle of 80 days was determined the mean number seedlings of laurel living m-2. Within brachiaria and jaragua they were higher with grass cover, although there were not significative differences (P>0.1). In estrella the seedlings living were significantly (p<0.5) higher with grass cover while in retana there were most living with grass cover removed (figure 5).
Species as laurel has soft seeds and they are easily spread by wind, removed by water and lost viability with extreme temperatures (Greaves y Mc Carter 1990; Boshier y Lamp 1997). Is possible so that the germination in Esparza was affected by rainfall, high tempatures and slope. Under Brachiaria its mode of growing improved conditions to germination and decrease mortality with shade and moisture, factors that have showed to help the natural regeneration of laurel (Butterfield 1995). Some tree species have problems in their establishment when there is enough foliage and roots, due to competence by factors productions and their growing decrease (Aide and Cavalier 1993). In Guapiles when the grass cover do not removed within estrella with high density of stem and foliage, there was high moisture and the seeds do not reach the soils, moreover the conditions were very wet and many seedlings died. In retana grass there was high mortality possibility due to alelophatic effect on seedlings, registered in studies to the same zone (Arosemena 1990).
The amount showed a tendency of better conditions in the humid zone (estrella) and both zone there was survivor within improving pastures. Three factors have special influence on the amount of seedling living finishing the evaluation cycle, the pasture and its growing habit the moisture conditions and cover presence.
Aide, T. Y Cavalier, J.1993. Barriers to tropical forest restoration in the Sierra Nevada de Santa Marta, Colombia. Restoration Ecology: 2:219-229.
Arosemena, E. 1990. Determinación de mecanismos de interferencia por alelopatía y requerimientos externos e internos de fósforo en pasto ratana (Ischaemun indicum (Houttt) Merrill). Tesis Msc. CATIE. Turrialba, Costa Rica. 124p
Butterfield, R. 1995. Desarrollo de especies forestales en tierras bajas húmedas de Costa Rica. CATIE. Serie Técnica. Informe Técnico No. 260 41p.
Boshier, A. y Lamp. T. 1997. Cordia alliodora genética y mejoramiento de árboles. Oxford forestry institute Department of plant sciences. University of Oxford. Tropical forestry papers.
Greaves, A., McCarter, P. 1990. Cordia alliodora, A promising tree for tropical agroforestry. Tropical Forestry Papers No. 22. 37p
Kampen, P. 1996. Trees in grassland. The influence of the trees on grass production within sylvopastoral systems of the atlantic zone of Costa Rica. Proyecto REPOSA (CATIE, MAG Wageningen University). Report No. 104. Field report No. 149. 33p
Larb, D.; Lapido, I.; Adekunle, J.; Smith, J. y Jabbar, M. 1998. Multiporpuse tree selection for silvopastoral system on acid Ultisol: The effect of grass competition on early growth of tree and shrub species. Tree crops journal. 9: 213-225.
Camargo, J[97]., Ibrahim, M.[98]
Key words: cattle defoliation, damages, mortality, timber species
The presence of timber species in pastures is a good economic and environmental alternative due to low prices of the animals products and environmental degradation within livestock systems (Pomareda et al 1997). In Cattle farms of Guápiles and Esparza, Costa Rica, the laurel (Cordia alliodora) is widespread by natural regeneration and is a promising species for silvopastoral systems because of little competition with associated grass for light (Kampen 1996).
However, cattle farmers have little knowledge about silvi-cultural management, diseases, cattle defoliation and susceptibility of laurel to weed control practices in the pasture and in general low quality timber of laurel is produced in silvopastoral systems. This study evaluated the quality of laurel in different growth stages of the natural regeneration within pastures in two zones with different ecological and management conditions.
Zone of study
This study was conduced Costa Rica using 60 sites as temporal plots in a nested sampling design. The sites were chosen randomize within 30 farms of the humid zone of Guapiles, with mean annual rainfall of 4000 mm and mean temperature of 24 °C; and 30 farms of the sub-humid zone of Esparza, with mean annual rainfall and temperature of 2000 mm and 24 °C respectively.
Quality evaluation of natural regeneration
Within each sampling plots were measured different stages of natural regeneration of laurel within pastures: seedlings (0.1 m (height < 0.3 m), sapling (0.3 m (height <1.5 m), young trees (1.5 m (height and dbh < 5 cm) and old trees (dbh > 5 cm) of laurel. Data was also taken on damages by cattle defoliation, herbicide burning, cut stems, dead individuals, diseases and shape or form of stems (on young and mature trees only).
Data analysis
Data were collected on farm management and land use dynamics through structured interviews with farmers. Soils were sampled to analyse physical and chemical properties and in each plot the abundance of fruit and timber trees associated with laurel was determined. Relative and absolute frequency of damages, mortality and good shape in four stages of natural regeneration of laurel was calculated. Also the hypothesis of independence of these variables with respect to site was tested using contingence analysis and simple correlation.
The results showed that saplings were more susceptible to damage than other growth stages with 43% of damages while old trees had 13%. Saplings also had a higher percentage of dead individuals(table 1).
Table 1. Damages and dead individuals in natural regeneration of laurel (Cordia alliodora). Guápiles y Esparza, Costa Rica
| STAGE OF GROWTH |
TOTAL POPULATION |
POPULATION WITH DAMAGES (%) |
DEAD INDIVIDUALS(%) |
||||||
| GUAPILES |
ESPARZA |
TOTAL |
GUAPILES |
ESPARZA |
TOTAL |
GUAPILES |
ESPARZA |
TOTAL |
|
| Seedling |
168 |
248 |
416 |
26* |
12.5* |
18 |
0sa |
0.8 |
0.5 |
| Sapling |
303 |
429 |
732 |
50* |
39* |
43 |
1* |
3* |
2.2 |
| Young tree |
261 |
145 |
406 |
18ns |
22ns |
19.5 |
0sa |
1.5sa |
0.5 |
| Old tree |
594 |
179 |
773 |
16* |
3.4* |
13 |
0.7* |
4.5* |
1.5 |
*= Significant differences between zone (p<0.05), ns= not significant, sa= without data
In terms of quality due to shape, old trees had better growth forms in Guápiles than in Esparza (42 vs 27%) (P<0.05). Although, young trees with good shape did not show significant differences between zones, they were more abundant in Guapiles (73 vs 60%) (figure 1). The number of individuals with good shape had high correlation (P<0.01) with the number of individuals without damages, sites with practices of silvicultural management and commercial used timber, while intensive control weed and high cattle density showed unfavourable effect (P<0.05) on growth forms.
For seedling, sapling and young trees the damage caused by cut stem (93.5, 73.5, 81%, respectively) was more frequent (figures 2, 3, 4). The majority of matured trees were damaged by parasite infection (97%) (Table 2). Damages for cut stem showed relationship with manual weed control (use of cutlass), cattle defoliation, and were less when there was a greater population of fruit trees in pastures. For the tree populations, problems for parasites were more important where greater moisture exist and lack of silvicultural practices. In general the damages are different according to growth stage with saplings being more susceptible to damage. Damaged caused by cattle defoliation is more important for saplings and young trees and depends with cow densities. Soil degradation and wet conditions favoured tree damages but better quality of the trees were found in the humid zone.
Table 2. Damages of old trees of laurel. Guápiles and Esparza, Costa Rica.
|
DAMAGES |
GUAPILES (%) |
ESPARZA (%) |
|
Cattle defoliation |
5 |
0sa |
|
Bark (lesions and cut) |
1 |
0sa |
|
Fire burn |
2* |
4* |
|
Stem broken |
0sa |
7 |
|
Diseases (chancro) |
5ns |
6ns |
|
Parasites |
97* |
10* |
*= Significance differences between zone (p<0.05), ns= not significance, sa= without data
Sapling laurel were more susceptible to damage presumably because they are reached easily by cattle and are easily chopped when manual weed control is undertaken, similar results was observed with sapling of Pithecellobium sp y Enterolobium sp within pastures (Hatheway y Baker 1970). The seedling are less defoliated by cattle because of small size. However, some workers found that that cattle trampling was associated with damage of laurel seedlings (Hart y Norton 1988). Young trees are most stable and have fewer problems, because they have surpassed several ecology and anthropoid obstacles. However, they are damaged by cattle defoliation and through indiscriminate weeding. Old trees have fewer damages, although when they present problems these are severed.. The presence of more dead individuals in Esparza, has relation with intensive weed control and burning practices, that additionally cause the death of several species and deteriorate the environmental (Brinkmann y Viera 1971; Lal 1987).
Trees had good shape where silvicultural management was practised and this is important for the marketing of cabinet timber.
Damages in natural regeneration of laurel differed according to stage of growth; saplings were found to more susceptible to different damages measured. Indiscriminate management by farmers regarding weed control and silvicultural practices is one of the main reasons for damages caused to laurel in silvopastoral systems.
Archer, S. 1995. Hervibore mediation of grass-woody Plants interaction. Harry Stobbs Memorial Lecture. Tropical Grassland. 29. 218-235
Brinkmann, W. and Viera, A. 1971. The effect of burning on germination of seeds at different soil depth of various tropical tree species. Turrialba 21: 77-82
Hatheway, W and Baker, H. 1970. Reproductive strategies in Pithecellobium and Enterolobium. Further information. Evolution (24). 253-254
Kampen, P. 1996. Trees in grassland. The influence of the trees on grass production within sylvopastoral systems of the atlantic zone of Costa Rica. Proyecto REPOSA (CATIE, MAG Wageningen University). Report No. 104. Field report No. 149. 33p
Lal, R. 1987. Tropical ecology and physical edaphology. John Wiley y Sons. New York. EUA.328p
Pomareda, C.; Perez, E.; Ganoza, V.; Matamoros, M. y Javier, O. 1997. La ganadería e industrias afines en Honduras: desafios y propuestas para su modernización. 75p.
H. Andrade[99], M. Ibrahim[100]
In silvopastoral systems there are relationships of interference and facilitation between components (Pezo and Ibrahim 1998), one of the most important is the alteration of the incident radiation to pastures and its effect on forage productivity (Tournebize and Sinoquet 1995; Mc Intyre et al. 1996).
The objective of this research was to evaluate the effect of two tree specie on incident radiation, productivity and radiation use efficiency of three improved grasses.
The study was carried out in private farm at Guapiles, Costa Rica (125 masl, average rainfall of 4560 mm year-1, average temperature of 24.6 oC., relative humidity 87%, and global radiation 13 to 17 MJ m-2 day-1).
The treatments were mixtures of two tree species (Acacia mangium and Eucalyptus deglupta) and three improved grasses (Brachiaria decumbens, Brachiaria brizantha y Panicum maximum) that were arranged in a randomized completely block design, with three replications. Trees were sown to 9 x 3 m into grasses. The main plots were divided in six strips between tree lines (Figure 1).
Figure 1. Representation of main
plots and strip plots (A to F).
The incident photosynthetically active radiation (PAR) on grasses was measured with ceptometer, taking 60 instant lectures between 11:00 am and 1:00 pm. Dry matter (DM) production was quantified through a 50 x 50 cm frame. The radiation use efficiency (RUE) was estimated using incident PAR and dry matter production (Eq. 1).
|
RUE = DMS/inPAR |
[Eq. 1] |
Where;
RUE: Radiation use efficiency (g MS MJ-1)
DMP: Dry matter production (g MS m-2 season-1)
InPAR: Incident PAR (MJ m-2 season-1)
InPAR = GR x 0.5 x %RT/100
GR: Global radiation (MJ m-2 season-1)
%RT: Radiation transmission
Incident PAR
Incident PAR on grass swards did not vary significantly (P>0.05) between tree species. However light transmission was significantly (P<0.01) different between tree position. Based on the intensity of light transmission, three zones were determined: full sun (C and D), intermediate shade (E and B) and shade (A and F), with transmissions of 98, 63 and 21%, respectively (Figure 2).
The tree crown of two species is different but apparently strips with high tree density eliminated the individual tree features. The difference between seasons (P<0.01) is due to changes in sun declination.
Forage yield
Forage yield differed significantly (P<0.01) between grass species but there were no significantly effect of tree species on this variable. Forage yield of P. maximum was higher than these of B. brizantha and B. decumbens (Figure 3) and estimated annual yields of these grasses were 34.8, 19.2, and 14.0 t ha-1 year-1, respectively.
Forage yield was greater at full sun (C and D) than in shade places (A and F). B. decumbens was the grass that lowest reduction in production in shade (23%), compared to B. brizantha (30%) and P. maximum (39%). These results show the plasticity of B. decumbens in shaded environments. Others studies have shown different responses of grasses under artificial shade or in silvopastoral systems (Zelada 1996, Bustamante et al. 1998).
Radiation use efficiency (RUE)
Rue was significantly different between grass species with average values of P. maximum (3.6 g DM MJ-1) very higher to these of B. brizantha (1.8 g DM MJ-1) and B. decumbens (1.4 g DM MJ-1). RUE of grasses were higher under shade zones (4.3 g DM MJ-1) compared to sunlight zones (1.3 g DM MJ-1) as shown in Figure 4. P. maximum had high values of RUE under shaded zones may be attributed to morphological and physiological adaptations of this specie under shade environments (Veenendaal et al. 1993).
The tree species had little effect on productivity and radiation use efficiency of the three grasses evaluated. However forage yields and RUE were different between species with Panicum maximum presenting highest values.
Bustamante, J.; Ibrahim, M.; Beer, J. 1998. Evaluación agronómica de ocho gramíneas mejoradas en un sistema silvopastoril con poró (Erythrina poeppigiana) en el trópico húmedo de Turrialba. Agroforestería en las Américas 5(19): 11-16.
McIntyre, B.D.; Riha, S.J.; Ong, C.K. 1996. Ligth interception and evapotranspiration in hedgerow agroforestry systems. Agricultural and forest meteorology 81: 31-40.
Pezo, D.; Ibrahim, M. 1998. Sistemas silvopastoriles. Turrialba, C.R. CATIE. Proyecto Agroforestal CATIE/GTZ. 258 p. Módulo de enseñanza agroforestal No. 2.
Tournebize, R.; Sinoquet, H. 1995. Ligth interception and partitioning in a shrub/grass mixture. Agricultural and forest meteorology 72: 277-294.
Veenendaal, E.M.; Shushu, D.D.; Scurlock, J.M.O. 1993. Response to shading of seedlings of savanna grasses in Botswana. Journal of Tropical Ecology 9: 213-229.
Zelada, E. 1996. Tolerancia a la sombra de especies forrajeras herbáceas en la Zona Atlántica de Costa Rica. M.Sc. thesis. CATIE, Turrialba, Costa Rica. 88 p.
Carlos Mazorra[101],
Ana E. De la Rosa[102], Gerardo
Borges[103],
Liliaysis Tapia[104], Dayamí
Fontes[105], Angela Borroto[106],
Rafael Pérez[107]
Key words: diet training, feeding preferences, food aversion, learning.
The foraging behavior of tropical sheep: sour taste preference (Arnold 1966) and greatness of ingesting shrub species (Favoretto 1990) both enable these animals as excellent consumer of pruning citrus in cut and carry systems (Borroto et al. 1986). However, this behavior constrain the grazing systems, because sheep like to browse the citrus leaf, and depress fruit production (Borroto et al. 1995). Our study look at methods of reducing damage of citrus trees during grazing.
We conducted the experiments between 1995- 2000 in citrus plantation of “Cítrico Ciego” enterprise. We used lambs of Pelibuey breed, which were randomly assigned to three experiments (6 replications per treatment).
Prior consumption of forage
The purpose of this experiment was to determinate whether grasses or legumes ingestion, previous to sheep pasturing, decreases the browsing behaviors.
Treatments
1. 4 hours legume consumption (Clitoria ternatea SN 139) + 4 hours of grazing.
2. 4 hours grass consumption (Dichantium annulatum) + 4 hours of grazing.
3. 8 hours of grazing.
Conditioned food aversion
This study was carried out to determine the effect of aversive conditioning on the sheep-browsing behavior.
Treatments
1. Aversive conditioning.
2. Control.
Dissuasive methods and their own combinations
In this trial we tested different dissuasive methods and their combinations to reduce sheep browsing.
Treatments
1. Aversive conditioning.
2. Mechanical dissuasion with “Bigotera” (Mazorra et al. 1996)
3. Aversion conditioning + Bigotera.
4. Control.
We found a representative vegetation (Borroto 1988) in the experimental areas: Paspalum notatum, Panicum maximun and Dichantium annulatum; in the last trial we observed the legume Terannus labialis cv. Semilla Oscura, too. In all experiment we used a stocking rate of 9 lamb per ha (Borroto, 1988). After grazing the animals were returned to their pens with ad libitum water and minerals. Specifically, in the experiments 2 and 3 we induced gastrointestinal illness (aversion) by administering lithium chloride (LiCl, 225 mg kg-1 body weight to the lambs (Launchbaugh y Provenza 1994), according to method used by Mirza and Provenza (1990). We recorded in all experiments the number and time of browsing per animal per grazing hour using a focal animal sampling procedure (Altman 1974).
The information, in different trials was examined with a simple ANOVA, using SPSS for personal computers (SPSS 1995); if the F-tests in the analyses of variance were significant (P< 0,05) we compared means using Duncan (1955). For the second trial we used paired t-tests to detect differences between treatment means.
The table 1 show the browsing behavior in all experiments.
Table 1. Number of browsing and time (s) employed in each one of them.
| Experiment |
Variable |
Treatment |
ES (±) |
Sign. |
|||
| 1 |
2 |
3 |
4 |
||||
| Prior consumption of forage. |
Browsing number |
0,091b |
0,085b |
0,186a |
- |
0,019 |
*** |
| |
Browsing time |
2,00b |
1,25b |
3,60a |
- |
0,35 |
*** |
| Conditioned food aversion. |
Browsing number |
0,027 |
0,186 |
- |
- |
0,009 |
*** |
| |
Browsing time |
0,24 |
3,60 |
- |
- |
0,19 |
*** |
| Dissuasive methods and own combinations. |
Browsing number |
0,009a |
0,037a |
0,000b |
0,020a |
0,004 |
* |
| |
Browsing time |
0,03a |
0,80a |
0,00b |
0,21a |
0,13 |
* |
** P<0,05; *** P<0,001
Prior consumption of forage
The browsing number and the time employed in each of them were similar in treatment 1 and 2, despite, both differ (P< 0,05) with treatment 3. These results demonstrated that the forage consumption reduced (around 70 %) the sheep browsing to citrus plantation, originated by a lesser grazing time into plantation.
Conditioned food aversion
The results showed in table 1 evidence the effectiveness of aversive conditioning to control the browsing in sheep (93% of the time employed in this activity).
Dissuasive methods and their own combinations
The combination bigotera- aversion refrains the animal browse fully; despite, the Bigotera (treatment 2) enhanced the animal browsing. Our results, in this trial, were very inferiors to the ones reported in the other experiments conducted in natural pasture without legumes. Therefore, the legume presence in the fruit- bearing cover deterrent the browsing to trees; in this sense Provenza (1995) suggest that animals learn which plants or foods to eat and which to avoid through interactions between a food’s flavor (odor, taste and texture) and the post- ingestive consequences of nutrients and toxins. Specifically, T. labialis, Skerman et al. (1991) reported it has a good leaf: stem relationships and high protein contain (around 20 %).
Implications
Table 2 show a preliminary analysis about implication the different methods in fruit production. For his analysis we take the following elements: It’s necessary dispose 2,3 m2 of foliar area to produced 1 kg of orange (Turrel 1969); the leaf foliar area and orange fruit weight are 27,3 cm2 and 192 g, respectively (Borroto 1988); the bite rate (bites per min) in a lamb is 13 leaf of orange (Mazorra, unpublished results). We included too 1 ha of orange plantation, grazing 8 hours all a year with 9 animal per ha of stoking rate.
Table 2. Affectations in fruit production according to different methods.
| Method |
Affectations |
||
| # Leaf |
# Fruit |
Production (kg) |
|
| Grazing in Natural cover |
20498 |
127 |
24,3 |
| Previous consumption of forages |
9253 |
57 |
11,0 |
| Conditioned food aversion in natural cover |
1367 |
8 |
1,6 |
| Grazing in Natural cover with legumes |
1196 |
7 |
1,4 |
| Conditioned food aversion in natural cover with legumes |
171 |
1 |
0.2 |
| Combination bigotera- aversion |
- |
- |
- |
The values observation in the Table 2 evidences that aversion, along or combined, may have potential to prevent the sheep browsing to citrus plantations. Nevertheless, the legume presence in the cover maybe cause reduction in sheep browsing (95 %).
The previous consumption, during 4 hours, either grasses or legumes reduced (around 70 %) the sheep browsing to citrus plantation.
Conditioned aversion has demonstrated to be the effective method to reduces the sheep browsing, independently of cover type.
The single presence of Terannus labialis cv. Semilla Oscura legume in the cover all appearance reduces the sheep browsing.
Altmann J. (1974) Observational study of behavior. Sampling methods, Behavior. 49: 225.
Arnold, G. B. (1960) Selective grazing by sheep of two forage species at different stage of growth. Aust. J. Agric. Res. 11 (6): 1026 - 1033.
Borroto, Angela, Venegas, M.T.; Mesa, M.A. & Hernández, A. (1986) Ceba estabulada utilizando forraje y residuos de podas. ACPA 1: 55 - 56.
Borroto, Angela. (1988) Potencial forrajero de los subproductos citrícolas para la producción de carne. Tesis de C. Dr. en Ciencias. ISACA - ICA. Cuba. 195p.
Borroto, Angela; Mazorra, C; Arencibia, Agueda; Hernández, N; López, M, Pérez, R;
Borroto, A. & Molina, A. (1995) Tecnologías Alternativas Sostenibles para obtener carne Ovina en las Fincas Citrícolas. Seminario Científico Internacional XXX Aniversario ICA. (Mem): 127 - 130.
Duncan, D.B. (1955) Multiple range and multiple F. Test Biometrics. 11: 1.
Favorettto, V. (1990) Pastagens para ovinos. En: Produçao de ovinos. Anais. Jaboticabal. FUNEP. 65 - 80p.
Launchbaugh, K.L & Provenza, F.D. (1994) The effect of favor concentration and toxin dose on the formation and generalization of flavor aversions in lambs. J. Anim. Sci. 72: 10-13.
Mazorra, C; Borroto, Angela; Arencibia, Agueda; Hernández, N; López, M; Rodríguez, Zayda & Pons, J.R. (1996) Dispositivos evaluados para disuadir el ataque de los ovinos a las plantaciones citrícolas. Rev. Automatizada Enlace. 2 (9).
Mirza, S.N. & Provenza, F.D. (1990) Preference of the mother affects selection and avoidance of foods by lambs differing in age. Applied Animal Science, 28: 225- 263.
Provenza, F.D. (1995) Postingestive feedback as an elementary determinant of food preference and intake in ruminants. Journal of Range Management, 48: 2 - 17.
Skerman, P.J.; Cameron, D.G. & Riveros, F. (1991) Leguminosas forrajeras tropicales. Edit. FAO. Roma. 707p.
SPSS. (1995) SPSS para windows. Versión 6.1.3. SPSS inc.
Turrel, F. M. (1969) Growth of the photosynthesis area of citrus. Bot. Gaz. 122: 284 - 298.
Salvador Hernández-Daumás[108], Graham Russell[109], John Arah[110]
Key words: Brachiaria decumbens, Gliricidia sepium, green manure, Leucaena leucocephala, simulation model.
In silvopastoral systems, prunings, leaf litter and root turnover from leguminous trees supply nitrogen for grass production. Pollarding provides green maure (mulch) while minimising competition for nutrients, light and water. This practice allows the synchronisation of nutrient release and nutrient demand. It is difficult to investigate all the possible combinations of treatments using traditional field trials. However, models can be used to extrapolate from a limited set of field observations. The objective of the present work was to develop a model to test the potential of Gliricidia sepium and Leucaena leucocephala as a source of green manure for Brachiaria decumbens pastures. In particular, it was designed to predict the long-term availability of mineral N and thus the sustainability of the system.
Field trials were carried out near Valle Nacional, Oaxaca State, Mexico to provide parameter estimates and data for testing the model. The model coupled two established process-based ecosystem models, the Hurley Pasture (HP) model (Thornley and Verberne, 1989) and the Edinburgh Forest (EF) model (Thornley and Cannell, 1992). Both consist of a Plant and a Soil sub-model driven by photosynthetically-active radiation, temperature and soil water status. The two plant sub-models use a single leaf photosynthesis approach and partition assimilates from photosynthesis, and nitrogen from the soil to the model components. The Soil sub-model consists of litter, soil organic matter (SOM) and mineral pools of N, connected by the soil microbial biomass pool. The animal sub-model, which is part of the HP model, dynamically represents the flows of C and N through the animals and back to the soil. The existing models had to be linked, parameterised for the new species and extended by the introduction of light competition, mulch production and pollarding. The complete model consists of 55 state variables and about 200 other variables and fluxes controlled by nearly 150 parameters. About a quarter of these parameters are species or site specific and had to be estimated from the literature and from the field trials. The model was developed in ModelMaker 3.0.3 (Cherwell Scientific Publishing Ltd, Oxford) and a period of 2500 days was simulated with pasture harvested every 40 days and trees pollarded every 100 days.
Biomass production
Predicted forage production was within the bounds of independent field datasets (Table 1) although seasonal variability was reproduced less accurately. B. decumbens root biomass was underestimated while G. sepium mulch production was 15% more than the highest figures in the literature.
Table 1. Comparison of steady state model outputs, experimental results (Hernández, 2000) and data from the literature.
|
Species |
Model output |
Experiment |
Independent datasets |
| Forage production |
0.85 - 0.98 |
0.54 - 3.45 |
1.09 (Carvalho, 1997) |
| Root biomass |
0.07 - 0.13 |
0.53 - 0.81 |
0.83 - 1.54 (Corrêa et al., 1999, Personal Communication. |
| Mulch production |
0.37 |
0.29 |
0.09 - 0.32 |
|
SOM stabilised protected + unprotected microbial |
0.17 |
- |
0.39 (Schimel et al., 1994) |
|
|
0.56 |
- |
0.57 (ibid.) |
|
|
0.02 |
- |
0.03 (ibid.) |
Soil organic matter
The model predicted that the pool of stabilised SOM would equilibrate at about 2.2 kg C m-2, but that the protected, unprotected, microbial and soluble pools would increase to 3.7, 0.06, 0.14, and 2.0 kg C m-2 respectively at the end of the 2500 days. These estimates agree with Schimel et al. (1994). However, total SOM at the field site ranged from 2.6 to 6.2 kg C m-2, i.e. lower than the model estimates.
When the tree component was removed from the model, an annual application of 150 kg N ha-1 was needed to in order to sustain grass production, in addition to the single application of 150 kg N ha-1 applied to both during the stabilisation period (Fig. 1). Compared with the pasture, litter was predicted to approximately double while below-ground cellulose and lignin increased ten times (Fig. 2). Since this is probably related to the death of tree roots caused by pollarding, too frequent pollarding may prevent the tree roots from populating the deeper parts of the soil. Mulch production from the foliage cut during pollarding was over-estimated in the model. The growth of the stem is limited soon after establishment by clipping the top. This is represented in the model by a partitioning coefficient which distributes the assimilates to stem and branch growth. The overestimated growth rate is thought to be a consequence of inadequate parameterisation.
Fig. 2. Litter fractions in the pasture (left) and the silvopasture (right)
Fig. 3. Tree biomass under a
pollarding regime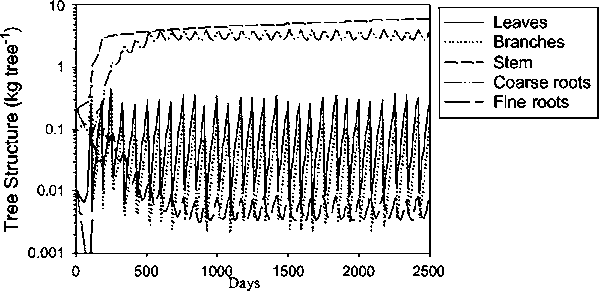
Effect of pollarding frequency
Although longer pollarding intervals produced a higher biomass harvest per cycle from the trees (data not presented), the effect was relatively small. This phenomenon can be explained by the increase in the pool of fine tree roots observed in simulations with longer pruning intervals.
The model results, although compatible with data in the literature, differed in some respects from the data collected in the field trials. In some cases this was thought to be due to poor parameterisation. There were, however, significant differences between the results from the individual field trials suggesting that some processes may have been omitted from the model.
Nevertheless, the results suggest that the model can be used to assess the effect of pollarding frequency on pasture production. It can also be used in the selection of appropriate species. For example, the degree of shading that can be tolerated by a pasture can be investigated and insights can be gained into the characteristics the tree species should have to permit complementarity between the growth of the trees and the grass.
The HP soil submodel (Arah, 1996) provided an effective link between the pasture and tree models.
Future research will involve improved parameterisation, spatial disaggregation of the tree canopy and the soil, and incorporation of the effect of shade on grass temperature.
Arah, J.R.M. 1996. The soil submodel of the ITE (Edinburgh) Forest and Hurley Pasture models. In: Evaluation of Soil Organic Matter Models using Existing Long-term Datasets. D.S. Powlson, P. Smith and J.U. Smith (eds.) pp. 225-230.
Carvalho, M.M. 1997. Asociaciones de pasturas con árboles en la región centro sur de Brasil. Agroforestería en las Américas. 15:5-8.
Corrêa, M.R.; Kanno, T.; Macedo, M.C.M.; Euclides, V.P.B.; Beretta, L.G.; Santos Jr. J.D.G. and Bono, J.A.M. 1999. Produção de raízes en cinco forrageiras tropicais sob pastejo na Região dos Cerrados (umpublished data).
Hernandez, S. 2000. Carbon and Nitrogen Balance in a Tree - Grass Inter - Cropping System in the Humid Tropics of Mexico.PhD Thesis. The University of Edinburgh, UK. 293 pp.
Schimel, D. S.; Braswell, B.H.; Holland, E.A.; McKeown, R.; Ojima, D. S.; Painter, T.H.; Parton, W.J. and Townsend, A.R. 1994. Climatic, edaphic, and biotic controls over storage and turnover of carbon in soils. Global Biogeochemical Cycles 8(3):279-293.
Thornley, J.H.M. and Cannell, M.G.R. 1992. Nitrogen relations in a forest plantation-Soil organic matter ecosystem model. Annals of Botany 70: 137-151.
Thornley, J.H.M. and Verberne, E.L.J. 1989. A model of nitrogen flows in grasslands. Plant, Cell and Environment 12:863-886.
Jean-Michel Harmand[111], Paul Ndonfack[112], Clement Forkong Njiti[113]
Key words: Acacia polyacantha, Eucalyptus camaldulensis, herbaceous dynamics, nitrogen cycling, Senna siamea
Fallows are often used for grazing in sub-Saharan savanna of Africa. The trees present in these systems have a variable influence on the dynamics of the herbaceous stratum according to climatic conditions. In dry conditions (annual rainfall lower than 800 mm), relative to the full sun situation, the reduced evapotranspiration and the greater nutrient availability under the tree canopy can allow a better growth of the understorey stratum (Bernhard-Reversat, 1982). However, in most of the Sudanian conditions with an annual rainfall higher than 800 mm, the growth rate of the herbaceous stratum is lower below the tree canopies than in full sun, as a result of light reduction (Cesar, 1992). Tree species can also play a part in herbaceous dynamics. There has been a growing pressure for the introduction of agroforestry practices in the Sudanian zone of Africa in order to ensure sustainable production. The objective of the present study was to assess the effect of tree species introduced in a fallow on the characteristics of the herbaceous stratum.
The study was carried out near the village of Ngong, located 40 km south of the town of Garoua in the North of Cameroon. The average annual temperature is 28.2°C. Total annual rainfall is approximately 1050 mm, 85% of which occurs within 5 months (May to September). The soil, derived from sandstone, was classified as a ferruginous soil (alfisol). The soil was deep and well drained, poor in organic C (0.35%) and total N, very sandy in the top layer but with an increasing clay content with depth.
In 1977-78, the former woody savanna was clearfelled and burned on-site by the farmer. After this procedure, a cropping system based on a biennial rotation of cotton and peanut was practiced for more than 10 years. In July 1989, the fallow treatments were established in a randomized complete block design with three replications. The tree spacing was 4 x 4 m (625 trees ha-1). Trees and peanut were planted simultaneously in 1989 and intercropping continued in 1990 with cotton. The fallow period lasted from 1991 until 1996. Treatments included three planted tree fallows: Acacia polyacantha, which is a local N fixing species which produces good quality arabic gum; Senna siamea and Eucalyptus camaldulensis, which are exotic species planted by the farmers in order to produce poles and fire wood. In addition, a spontaneous herbaceous fallow was established in 1991. All these treatments were protected against fire and grazing. Furthermore, measurements were made in a natural grazed fallow established in 1987 close to the trial. This was regularly overgrazed and sometimes burned. A herbaceous species inventory was done, at the end of the rainy season, with estimation of a ground cover index for each species and each plot. An average species index was calculated per treatment and transformed into a percentage value of herbaceous ground cover. Herbaceous biomass was estimated, when the development of the herbaceous stratum was maximum. Herbs were collected in each of the planted tree fallows in 10 randomly chosen triangular plots of 2 m²; each plot was representative of the quadrat formed by 4 trees. Nitrogen content was estimated for each of the ten samples.
Throughout the fifth year of fallow, mineralisation and nitrification of soil N was studied via field incubation. In situ incubations of the top 20 cm soil layer were done every four weeks. The results were analyzed by ANOVA followed by F test. In case of significance of the F test at p < 5%, treatment means were compared by Duncan’s multiple range test at the same level of probability.
Herbaceous vegetation characteristics
During the first two years of fallow, weed species of the cropping system were replaced by fallow species such as Pennisetum spp. and Andropogon gayanus (Figure 1a). When shade was not excessive, the protection against grazing induced the progressive development of A. gayanus, a perennial gramineous species, to the detriment of other herbaceous species present initially, especially Pennisetum spp.. In the spontaneous fallow (Figure 1b), which was protected against fire and grazing, A. gayanus provided 70% of ground cover and the greatest biomass (8 t.ha-1). In one particular plot of the trial, the percentage ground cover of A. gayanus was 100% and the biomass 10 t.ha-1 (Figure 1a). In this case the reduction in species diversity of the herbaceous stratum corresponded to an increase in herbaceous biomass. The understorey of E. camaldulensis plots showed the same pattern as the previous treatment (Figure 1d), but as a result of the shade effect, the herbaceous stratum was less dense and its biomass lower (Table 1). This result confirms previous findings indicating that, under these site conditions, the growth rate of the grass was lower below the tree canopy than in full sun. Under A. polyacantha, A. gayanus (heliophilic species) did not grow and the herbaceous stratum consisted mainly of Pennisetum spp., annual gramineous species which are shade tolerant (Figure 1e). Pennisetum polystachyon, which developed to the detriment of Pennisetum pedicelatum, provided 80% of ground cover in the fifth year of fallow, but it was less productive than A. gayanus (Table 1).
Table 1. Biomass, nitrogen content and nitrogen concentration of the herbaceous stratum in fallow treatments in the Sudan Cameroon. In a given range (*column in the case of nitrogen concentration), means followed by the same letter are not significantly different (p<0,05).
|
Year |
Acacia polyacantha |
Senna siamea |
Eucalyptus camaldulensis |
Protected herbaceous fallow |
Grazed herbaceous fallow |
|||||
|
Biomass (kg ha-1) |
||||||||||
|
1993 |
2700 |
c |
2450 |
Cd |
2700 |
c |
4470 |
a |
3150 |
bc |
|
1994 |
660 |
d |
360 |
D |
3190 |
c |
6130 |
a |
4540 |
b |
|
1995 |
2240 |
cd |
180 |
E |
3490 |
bc |
7890 |
a |
4420 |
b |
|
Nitrogen content (kg ha-1) |
||||||||||
|
1993 |
24 |
ab |
13 |
C |
18 |
bc |
26 |
ab |
25 |
ab |
|
1994 |
7 |
d |
3 |
D |
19 |
bc |
30 |
a |
25 |
ab |
|
1995 |
29 |
b |
2 |
C |
25 |
b |
38 |
a |
32 |
ab |
|
Nitrogen concentration (% of dry matter) |
||||||||||
|
1993 |
0,90 |
f* |
0,51 |
|
0,67 |
|
0,59 |
|
0,81 |
|
|
1994 |
1,11 |
g* |
0,75 |
|
0,60 |
|
0,48 |
|
0,55 |
|
|
1995 |
1,29 |
h* |
1,00 |
|
0,72 |
|
0,48 |
|
0,73 |
|
In the natural grazed fallow (Figure 1c), overgrazing led to degraded pastures with annual species such as Andropogon pseudapricus, Schizachyrium exile, Spermacoce stachydea and Spermacoce radiata. This treatment maintained the greatest species richness after more than five years of fallow. Nethertheless, in comparison to the protected spontaneous fallow, the herbaceous biomass was reduced by 50%.
Each year, the ground cover of the grass was often complete in all treatments except in S. siamea where it was lower than 30% despite the relatively discontinuous and open tree canopy. Furthermore, under this species, the herbaceous biomass was very weak, indicating a strong depressive effect of the tree on the herbaceous stratum (Table 1).
Nitrogen cycle
A. polyacantha had the greatest herbaceous N concentration, which increased significantly from one year to the next (Table 1). Nitrogen fixation and a high external recycling of N, characterised by a high soil N mineralisation rate (Table 2), are the processes which contribute to increase the storage of nitrogen in the different compartments of the system, soil included (Harmand and Njiti, 1998). After four years of fallow, A. polyacantha was actually the only species which improved significantly the C and N contents of the 0-20cm soil layer. This increase was mainly due to coarse plant debris (50 µm to 2000 µm size fraction) (Harmand et al., 2000). In the case of the spontaneous fallow, which consisted mainly of A. gayanus, the low soil N mineralisation level associated with a high biomass and the smallest herbaceous N concentration (Table 2) demonstrated the high N use efficiency of this type of herbaceous savanna (Abbadie et al., 1996). Due to their higher biomass, the herbaceous fallows had the greatest amount of N stored in the herbaceous stratum, while the opposite was true for the undergrowth vegetation in S. siamea (Table 1). The lowest soil N mineralisation rate occured under this species and was associated with a decline in tree growth and with some tree decay from the fourth year of fallow onwards. Schroth et al. (1995) have already reported low soil N mineralisation rates under species of the Ceasalpinaceae family. Soil N mineralisation could be inhibited by an allelopathic effect of the tree species (Attiwill and Adams, 1993) resulting in a strong depressive effect on the herbaceous stratum.
Table 2. Soil N mineralisation in the 0-20 cm layer during the fifth year of fallow. In a given range, means followed by the same letter are not significantly different (p<0,05). (Harmand and Njiti, 1998)
|
Fallow treatment |
A. polyacantha |
S. siamea |
E. camaldulensis |
Herbaceous fallow |
||||
|
Kg N ha-1 year-1 |
166 |
A |
25 |
c |
40 |
b |
26 |
c |
|
% soil total N year-1 |
17 % |
A |
3,1% |
d |
6,1 |
b |
3,6% |
c |
The tree species influenced light interception and the N cycle - each in different ways - and thus had different effects on the herbaceous stratum in terms of species composition, biomass and N content. Protection against grazing and the introduction of tree species induced both a reduction in herbaceous species richness and a progressive evolution to a particular species composition. E. camaldulensis provided little shade allowing the growth of A. gayanus, a species readily accepted by cattle, but A. gayanus is so sensitive to grazing that it needs rotational grazing practices to ensure its preservation. As a N fixing species, A. polyacantha provided a gramineous herbaceous stratum consisting of Pennisetum spp. with a higher N concentration than found under the other tree species. Due to its strong depressive effect on the herbaceous stratum, Senna siamea cannot be used in sylvopastoral systems.
Abbadie L Lepage M and Menaut JC (1996) Independence of savanna grasses from soil organic matter for their nitrogen supply. Ecology 73 (2): 608-613
Attiwill PM and Adams MA (1993) Tansley Review N°50: Nutrient cycling in forests. New Phytologist 124(4): 561-582
Bernhard-Reversat F (1982) Biogeochimical cycle of nitrogen in a semi-arid savanna. Oikos 32: 321-332
Cesar J (1992) La production biologique des savanes de Côte-d’Ivoire et son utilisation par l’homme. Biomasse, valeur pastorale et production fourragère, Cirad-IEMVT, France, Maison Alfort, 671 p
Harmand JM and Njiti CF (1998) Effets de jachères agroforestières sur les propriétés d’un sol ferrugineux et sur la production céréalière. Agriculture et Développement, Spécial sols tropicaux, 18: 21-30
Harmand JM, Njiti CF, Bernhard-Reversat F, Feller C, and Oliver R (2000) Variations de stocks de carbone dans le sol au cours du cycle jachère arborée - culture (zone soudanienne du Cameroun). In: Floret C and Pontanier R (eds): vol. I, pp. 000-000, (2000)
Schroth G, Kolbe D, Pity B and Zech W (1995) Searching for criteria for the selection of efficient tree species for fallow improvement, with special reference to carbon and nitrogen. Fertilizer Research 42: 297-314
Figure 1. Change in specific composition of the herbaceous stratum in the different fallows (% of the total ground cover of the herbaceous stratum)
G. Febles, T.E. Ruiz, J. Alonso, J.C. Gutierrez[114]
Key words: Albizia falcataria, Azadirachta indica, Gmelina arborea, silvopastoral systems
The importance of the introduction of silvopastoral systems in Cuba was previously highlighted by Febles, Ruiz and Simón (1995). The thorough research on Leucaena leucocepala studied from an integral point of view in Cuba and other countries as well as the progress in the knowledge of certain genera such as Gliricidia, Morus, Erythrina, Trichantera, Cratylia and Acacia is well known (Rosales et al 1999). The objective of the present study was to introduce tree species in livestock units without interrupting the zoo-technical flux for different purposes such as shade and animal feeding.
Two dairy units with star grass (Cynodon nlemfuensis) as the predominant pasture grass were used at the Institute of Animal Science. All trees were planted by seeds in plastic bags and maintained in a greenhouse and transplanted to the field. In the first experiment Albizia falcataria, Azadirachta indica and Gmelina arborea constituted the plants studied. They were sown along the individual paddocks in strips or bands at 3 m from the border cement fence and at 3 m from the electric fence that protected the plants from the adjacent grazing area. Two other treatments of the same type but at 4 x 4 m and 5 x 5 m were employed. Another variant of this experiment was the sowing at the central corner of the paddocks, but only at the 4 x 4 m distance as was previously discussed. In the second experiment also including Cynodon nlemfuensis, the study was carried out with Brosimum alicastrum, Albizia lebbekoides, Bauhinia galpinii, Bauhinia purpurea and Enterolobium cyclocarpum. In this case the species were planted within the pasture grass in frames of 3 m width and 10 m apart in a well prepared soil. The first three species could not be established and the research was conducted with the other two plants. Survival, height, stem growth and the effect of the animals on the plants in the experimental areas, when the electric fence was eventually withdrawn were measured during two consecutive years. In this last case a score method starting with plants not affected to 5: plants very affected by grazing or other aspects of animal behavior was used.
Previous results were obtained by García Vila (1992) for the variant in which trees were sown in the central corner of the grazing areas. At present other attempts are under study also in relation with the sowing of trees along the paddocks. At this point we would like to emphasize that the experimental area without animals is less than 8% of the total grazing area. Therefore, the negative effect of overgrazing in the Cynodon pasture is not considered. Results indicate that survival was 80 to 89% in Gmelina and Albizia, respectively showing the stability of both species, resistance and an equilibrium with the environment under these competitive conditions in which Cynodon nlemfuensis was present. An increase was observed in plant height, mainly in Albizia and Gmelina compared to the Neem tree. This result also expresses the equilibrium achieved by the former two trees.
Table 1. Effect of different samplings on survival, height and influence of the animals on trees planted along the paddocks
|
Indicators |
Samplings |
SEt |
Sign |
|||
|
1 |
3 |
5 |
7 |
|||
| Survival, % |
81.10ª |
65.3b |
63.8b |
61.1b |
2.2 |
P<0.001 |
| Height, cm |
46.8e |
140.4c |
168.2ab |
1777.7ª |
3.5 |
P<0.001 |
| Animal effect |
|
1.7 |
|
1.9 |
0.1 |
|
abcde Means with different letter within each column differ significantly at P <
0.05 (Duncan 1955)
A more complete information is offered in table 1 indicating survival, height and animal effects on the trees sown along the cement fences. The results were the same as in the other variant. No differences in treatments were found. The information on the effect of animals on the trees was very important since no negative influences were observed. In this sense, only a few secondary branches were broken. A supplementary sampling carried out after the 7 th month and at 2 years from planting suggests a growth and survival of 365.0 cm and 97% for Albizia, respectively and of 242.0 cm and 97% in the case of Gmelina. The Neem tree only survived 33% with a height of 92 cm approximately.
On the other hand, the results of the second experiment underline the stability achieved by the trees measured through indicators and evidently expressed the equilibrium obtained in plant survival under a high competitive stress since the frames within the paddocks were invaded by Cynodon nlemfuensis. Height continuously increased exceeding 200 cm. Stem growth augmented significantly from 1.93 cm to 30.2 cm in Enterolobium and from 1.65 cm to 21.1 cm in Bauhinia. Animal effect was at a lower score of only 1.48 in both trees. It should be stressed that the use of plant diversification based on genera, species and plant families was an essential criterion considered in these two experiments when the introduction of tree plants is required in livestock grazing areas (Ruiz et al 2000 unpublished data). This agrees with the concepts indicated by the first Electronic Conference organized by FAO (Rosales 1999) and by Sánchez (2000). The results of these experiments have shown the possibility of introducing and establishing tree species other than Leucaena leucocephala in commercial livestock areas without altering the management of the unit and for different productive purposes that will be confirmed in the future performance of these areas.
Duncan, D.B. (1955). Multiple range and multiple F tests. Biometrics 11:1.
Febles, G., Ruiz, T.E. & Simón, L. (1995). Consideraciones acerca de la integración de los sistemas silvopastoriles a la ganadería tropical y subtropical. Instituto de Ciencia Animal. Seminario Científico, La Habana, Cuba. Conferencias. p. 391.
García Vila, R. (1992). Algunos índices para la proyección de unidades para la implantación del PRV p. 19.
ICA (1999). Sistemas silvopastoriles. Conceptos y tecnologías desarrolladas en el Instituto de Ciencia Animal de Cuba. Ed. EDICA pp 34.
Rosales, M., Murgueitio, E., Osorio, H., Speedy, A. & Sánchez, M. (1999). Conclusiones y evaluación de la conferencia electrónica. Primera Conferencia Electrónica. FAO, Roma, Italia, p. 492.
Sánchez, M. (1999). Sistemas agroforestales para intensificar de manera sostenible la producción animal en América Latina tropical. Primera Conferencia Electrónica. FAO, Roma, Italia p. 1
Sánchez, M. (2000). Programa de los sistemas agroforestales pecuarios en América Latina. Reunión Internacional sobre sistemas agroforestales pecuarios en América del Sur. Brasil, p. 7
Plevich J O[115], Barotto[116] O, Ohanian[117] A, Pagliaricci H[118], J Garcia[119]
Key words: Eucalyptus viminalis, Medicago sativa, Pinus elliottii, Quercus robur, silvopastoral system
In the last 20 years the investigation of integrated or multiple land use production systems has entered into the area of modeling the diverse physiognomies in order to dedicate them to the production of food for livestock or to the joint production of herbage, wood, firewood, or other products for human consumption or utilization (Olivares, 1992; FAO, 1985; Olivares, 1989; Cabrera, 1993; Ayans 1994).
In the Central-South Region of the province of Cordoba in Argentina the empirical data which exists for these systems is about the behaviour of natural grassland under the influence of the canopy of the genus Pinus sp, finding a prolonged seasonal growth, in relation to areas devoid of trees (Valls, 1993). However, studies do not exist which show the productive behaviour of the principal components of the system over time. In this study the proposed objective is to analyze the behaviour of the woody species of a silvopastoral system during the first year of implantation. In order to do this the effect of the factors plantation design and species on the parameters of height, base diameter, and survival were considered. In the herbaceous species the effect that the accompanying woody species and spatial arrangement utilized have on the survival of plants are analyzed.
The investigation was carried out on land which is used for teaching and research called Pozo del Carril of the Faculty of Agronomy and Veterinary Studies of the National University of Rio IV, Department of Rio Cuarto (Cordoba, Argentina) located at 32ª 51’South Latitude and 64ª 40’ West Longitude and at 550 metros above sea level. To achieve the proposed objective parcels were established where silvopastoral systems were planted with trenches of the woody species (Quercus robur,Ffraxinus americana, Eucalyptus viminalis and Pinus elliottii). The trenches with woody species were placed in double file and in two configurations in two different sizes of squares: 2x2 m and 3x3 m, while the space between trenches was 12m and 21m respectively. In the open space an herbaceous species, Medicago sativa was planted at a density of 4Kg./ha.
From the combination of the two space configurations and the four woody species 8 study trials were formed:
1) MP1E: Plantation design; 3x3m; Eucalyptus viminalis
2) MP2E: Plantation design 2x2m; Eucalyptus viminalis
3) MP1P: Plantation design 3x3m; Pinus elliottii
4) MP2P: Plantation design 2x2m; Pinus elliottii
5) MP1F: Plantation design 3x3m; Fraxinus americana
6) MP2F: Plantation design 2x2m; Fraxinus americana
7) MP1Q: Plantation design 3x3m; Quercus robur
8) MP2Q: Plantation design 2x2m; Quercus robur
These trials were studied using a design of blocks completely randomized. In these parcels provided with three trenches and two open spaces, previously mentioned parameters were evaluated. In the central trench the parameters corresponding to the woody species were evaluated, taking a minimum of 4 plants. In the open spaces on each side of the central trench and at random samples were taken of the herbaceous species. The data was statistically processed using a variance analysis and Duncan test.
In the first stage of the study, the dynamic of the components of a silvopastoral system were analyzed during the first year of implantation. In the woody component the parameters evaluated were: height (cm), base diameter (mm) and survival (% of live plants) and in the herbaceous survival (plants/m2).
The trials correspond to the combination of different spatial arrangements and woody species utilized. The results are shown in Table 1.
Table 1: Effect that the spatial arrangements and woody species utilized have on height, base diameter and survival of the woody and herbaceous species.
| Trials |
Height(cm) |
Parameters evaluated |
Survival(% Live plants) |
Herbaceous species |
| Woody species |
||||
| Base diameter(mm) |
Survival(Nºplants/m2) |
|||
| MP1E |
68,2 ± 5,5 a |
4,7 ± 0,4 b |
76,7 ± 16,4 b |
29,3 ± 11,6 a |
| MP2E |
71,8 ± 6,2 a |
4,9 ± 0,4 b |
63,7 ± 20,4 b |
36,0 ± 8,0 a |
| MP1P |
26,5 ± 1,8 c |
4,4 ± 0,3 b |
65,3 ± 16,9 b |
22,7 ± 1,8 a |
| MP2P |
27,8 ± 2,8 c |
3,9 ± 0,3 b |
71,0 ± 9,3 b |
34,7 ± 4,4 a |
| MP1F |
63,1 ± 2,2 b |
6,8 ± 0,3 a |
82,0 ± 1,3 a |
28,0 ± 2,7 a |
| MP2F |
63,2 ± 2,2 b |
7,1 ± 0,1 a |
88,3 ± 4,4 a |
30,7 ± 4,4 a |
| MP1Q |
51,8 ± 5,8 b |
7,7 ± 1,3 a |
95,0 ± 4,0 a |
29,3 ± 3,6 a |
| MP2Q |
52,6 ± 10 b |
7,2 ± 1,0 a |
91,0 ± 5,3 a |
24,0 ± 2,7 a |
*Different letters in the same column indicate significant differences (p=0.05)
From the partition of the variance and from the study of the behaviour of the woody species it was possible to observe that the differences in the parameters evaluated are principally due to the effect of the species used, since differences due to the plantation design used (spatial arrangement) are not observed, nor the effect of the environment (blocks), nor the effect of the interaction between species x plantation design.
From the analysis of the effect of the species utilized it is observed that the greatest growth in height was obtained by Eucalyptus viminalis which differs significantly from Pinus elliottii, Fraxinus americana and Quercus robur. With regard to the last two species it can be observed that no differences exist between them, but their behaviour was better than Pinus elliottii which presented the smallest amount of growth in terms of height.
When the growth in base diameter of the woody species is analyzed a more homogeneous behaviour is observed with only two groups being formed, that of Fraxinus americana- Quercus robur and Eucalyptus viminalis-Pinus elliottii, of which the first group had the better behaviour.
With regard to survival it is necessary to mention that it was supposed that only one irrigation at the moment of the plantation would be sufficient to pass the threshold of an 80% survival rate. As can be observed in Table 1 this hypothesis was true only for the species of Fraxinus americana - Quercus robur, requiring more supplementary irrigations in the case of Eucalyptus viminalis and Pinus elliottii in future plantations.
When the survival behaviour of Medicago sativa is analyzed, in regard to the plantation design and the woody species utilized for establishing silvopastoral systems no significant differences are observed during the first year of implantation in any of the trials.
During the first year of implantation of a silvopastoral system composed of the woody species: Fraxinus americana, Quercus robur, Eucalyptus viminalis and Pinus elliotti and for the conditions which were manifested in the experiment it is possible to conclude:
The greatest growth in height was expressed in Eucalyptus viminalis, while the greatest growth in diameter corresponded to Quercus robur and Fraxinus americana.
The implementation of only one supplementary irrigation for the woody species during the implantation stage made it possible to reach the 80% rate of survival required by law in order to offer subsidies to forestry activity in the case of Fraxinus americana and Quercus robur; but, this technique must be intensified in Eucalyptus viminalis and Pinus elliottii, where this rate of survival was not obtained.
During the first year of implantation of Medicago sativa planted in the open spaces between trenches of woody species, effects of plantation design were not observed on their rate of survival, nor were any effects observed on the rate of survival of the woody species utilized.
Ayans A, 1994. La dehesa española: origen, tipología, características y gestión. Escuela Técnica Superior de Montes. Fundación Conde del Valle de Salazar. 96 p.
FAO, 1985. Ordenación forestal en los tropicos para uso múltiple e intensivo. Roma. Estudio FAO, Serie Montes (55): 180
Olivares A, 1989. El ecosistema silvipastoral. Avances en Producción Animal 14 (1-2): 3-14.
Olivares B, 1992. Experiencias de la Universidad Austral de Chile en la investigación agroforestal en la decima región: un estudio de caso. IN: Garfias R (De), Seminario de agroforesteria. Potencialidades y restricciones dentro del desarrollo agroforestal chileno. Facultad de Ciencias Agrarias y Forestales, Universidad de Chile. Santiago, Chile. p 13-19.
Sequeira W and Gholz. 1989. Canopy structure, light penetration and tree growth in slash pine (Pinus elliotti) silvopastoral system at stand configurations in florida. Department of forestry. University of florida. Gainesville, Florida.
Valls P, 1993. Introducción al manejo integral de Recursos: pastoreo racional intensivo en un sistema forestal de sierra. Jornadas sobre pastoreo racional intensivo. Sociedad Rural de Río Cuarto, 3 al 4 de junio de 1993. Río Cuarto, Córdoba, República Argentina.
J. Alonso[120], G. Febles[121], T.E. Ruiz[122], J.C. Gutierrez[123]
The utilisation of silvopastoral systems for cattle production is a practical option for land utilisation that can be obtained under adequate management with an economic and ecological balance. The utilisation of trees and shrubs as live fence post is an attractive system used by farmers in tropical Latin America and in other areas of the world. However although the interactions between herbaceous plants and animals are not very important in these cases the system should be focussed on the management and maintenance of the tree components (Otárola 1995 and Montagnini 1994). In this paper, a group of experiments are described to study the establishment of woody species and as live fences in grazing areas. A group of growth components and their performance during the year were considered for a more harmonic equilibrium between plant growth components and their interaction with the environment.
The experiments have been carried out since 1997 at the Institute of Animal Science located at 22 o 53' North latitude and 82º 02'of the Western longitude at 92 m above sea level mainly on a red ferrallitic and greyish brown soils (Anon 1995). Table 1 summarises the experimental design and the treatments used in each experiment. The main measurements assessed were height, number and distribution of regrowth points and survival.
Significant interactions (P < 0.05) were found between species and planting methods for the number of plant regrowths and survival. The use of decorticated stems in piñón florido and piñón de pito species showed the highest values in the number of regrowths (15 and 13, respectively), almácigo planted by the same method (4) and piñón florido planted with injured stems (2) had the lowest number of plant regrowths. Regrowth distribution was not the same for the species studied (graphic 1) indicating a better performance for piñón de pito with values of 83.4% in strata 2 and 3. Piñón florido planted by injured stems showed the lowest survival percentage (8.3%). There were no interactions between planting dates and tree species in any of the measurements studied (table 2). However, main effects showed (P < 0.05) a better performance regarding plant height in May (1.98 m). The best survival (97.5%) was reached by piñón florido not differing from piñón de pito (92.5%). Both species differed (P < 0.05) from ciruela (80.0%). The highest number of regrowths for piñón florido and ciruela grew mainly in the basal part of the stem.
On the other hand, no interaction was found between planting date and the moon phases. Differences were observed (P < 0.01) for the number of plant regrowths with the lowest value during the crescent moon. Significant differences for survival were encountered for the moon phases and the lowest figure (75%) was found during the crescent moon.
The establishment of trees and shrubs, as well as other vegetable species, should consider some indicators that would relate the concepts of growth and development of the plant. However, among these indicators it is important to emphasize the biological characteristics of the species so they allow the settlement of the basis on which the works of establishment of trees and shrubs as living fences in silvopastoral systems rest (Alonso et al 2000).
Re-growth distributions through the stem for these species had the same performance pattern in all the experiments, where the highest percentage of re-growth was found in the strata 1 and 2 (Graphic 1). When it is not possible to protect certain tree species that are introduced as living fences and they are consumed by animals, for example the Gliricidia sepium, we have to take into consideration the part of the stem in which the higher amount of re-growth is reached so to avoid the consumption, being this a biological expression of the plants that changes according to species. When analyzing this topic, it is necessary to take into account that the survival of tree species is closely related to their adaptation capacities to the different agroecological conditions they are developed that are, at the same time, influenced by the great genetic variability of the species
It is concluded that the planting methods used in these experiments showed interactions with the tree species studied finding that the method of normal stem planting and by injured stem are the best methods recommended for almácigo and that decorticated stem for piñón de pito. Regrowth distribution in the stem differs according to species. This characteristic should be taken into consideration to select species to be introduced as live fences in grazing areas. May is the most reliable month for a satisfactory establishment of this species. The moon phases had an important influence on survival
Alonso, J.; Febles, G. y Gutiérrez, J.C. 2000. Métodos de introducir especies arbóreas como cercas vivas en áreas de pastoreos. Rev. Cubana Cienc. Agric. 34 - 157p.
Anon (1995). Nueva versión de la clasificación genética de los suelos de Cuba. Ciudad de la Habana. MINAG 26 P
Montagnini, F. (1994). Boletín Agroecológico. Junio p. 18
Otárola, A. (1995). Cercas vivas de madero negro: Práctica Agroforestal para sitios con estación seca marcada. Agroforestería en las Américas Vol 2 No. 5 enero-marzo
Table 1. Experimental designs and treatments used in the experiments.
|
Experiment |
Experimental design |
Treatments |
Observations |
|
Planting date on grazing areas |
Split plot 2 x 3 with 4 replications |
Main plot |
Grazing areas for milk production. Live fences protected with electric wire. |
|
Methods in grazing areas |
Split plot 3 x 3 with 4 replications Moon phases for G. sepium |
Main plot |
Grazing area with young animals. Live fences not protected from animal access |
|
Moon phase on G. sepium |
Randomized blocks in factorial arrangement Factorial 2 4 with 4 replications |
Factor A: Moon phases |
Areas without animals |
Table 2: Performance of the number of re-growths1 per plant and survival2 in tree species introduced as living fences in grazing areas.
|
|
Number of re-growths per plant |
Survival (%) |
|
Species |
|
|
|
Piñón florido |
3.81 (14) |
1.98a (97.5) |
|
Piñón de pito |
3.42 (12) |
1.96a (92.5) |
|
Ciruela |
3.39 (11) |
1.89b (80.0) |
|
ES± |
0.04 |
0.02* |
|
Plantación Date |
|
|
|
March |
3.49 (12) |
1.97 (95.0) |
|
May |
3.59 (13) |
1.92 (85.0) |
1 Transformed data according to ÖX
2 Transformed values according to arcsine Ö%
() Real values * P<0.05
ab Means with different superscripts differ significantly inside each column (P<0.05)
Idésio Luis Franke[125],
Elias Melo de Miranda[126],
Judson Ferreira Valentim[127],
Felipe Alexandre Vaz[128]
Key words: forages, silvipastoral systems, tree legumes, western Amazon
The State of Acre has a population of 500 thousand inhabitants, with a territorial extension of 152.589 km2, which represents 1,79% of the National Territory and it is located in the Western part of the Brazilian Amazon, occupying 3,16% of this region (IBGE, 1997). In the State of Acre, the predominant ecosystem is the tropical rainforest, with high diversity of species per hectare. In these areas, the main economic activity in the last century has been the extraction of the latex of the rubber trees (Hevea brasiliensis) and the collection of Brazil nut (Bertholletia excelsa)). However, in the last 30 years, the disruption of the extractive system in the “seringais” (native rubber states), the creation of settlement projects and the expansion of cattle ranching activities transformed the economy of the primary sector of Acre (Valentim 1989; Valentim and Moreira 1994).
Tree species generally grow more than forage species, so that when they are established in the same area, the first ones interfere in the passage of light for the herbaceous extract under the tree canopies. In most of the situations, the rate growth of the pastures is lower below the tree canopy than in full sun. However there are differences among the forages, because some are more tolerant than others when submitted to lower light conditions (Pezo and Ibrahim 1998).
There have been growing pressures for reforestation, particularly with multiple use trees in silvipastoral systems which vary according the environmental, social e economic characteristics of the different regions of the world. Silvipastoral systems are land use alternatives that can be ecologically and economically viable in the Amazon. In order to achieve these goals, these systems should be well planned, with the selection of the most appropriate tree species being one of the most important aspects in this process. Successful associations of trees and forage species depend on a careful evaluation of the interactions among the components of the ecosystem.
The objective of this study was to evaluate the effect of shading of two species of native tree legumes on chemical composition of forage produced by Penisetum purpureum.
The study was developed in two farms in the vicinity of the Experimental Station of Embrapa Acre, located in the Highway BR 364, municipal district of Rio Branco, Acre, Brazil. The annual mean precipitation is of 1865 mm, with rains distributed between October and May and dry season going from June to September, with 85% of relative humidity of the air and annual mean temperature of 24,3 oC.
The area was deforested in the beginning of the decade of 80, followed by the establishment of pastures of the grass Panicum maximum and in the sequence Brachiaria decumbens. In 1999 the area was established with the grass P. purpureum.
The experimental design was completely randomized with three repetitions. The tree species were isolated in an area of pasture of P. purpureum with no grazing. The tree species Samanea sp. and Enterolobium maximum, under which the samples of forage of P. purpureum were collected have approximately 15 and 25 years, respectively. The tree species belong to the family Mimosideae, have canopy in the form of an umbrella, possessing from 3 to 5 main trunks with several ramifications, small leaflets, which fall in the beginning of the dry season. The species Samanea sp. remains from two to three months defoliated while E. maximum remains only one month.
The collection of samples took place in April 2000, at the end of the rainy season with the objective of evaluating forage quality under shading conditions provided by the tree canopies in contrast with the conditions of full sun. The grass was cut at 5 cm height from the soil surface and samples were taken and oven dried at 65 oC for 72 hours until constant dry weight was achieved.
The chemical composition of the forage of P. purpureum was analyzed regarding the following quality indicators: crude protein (CP), neutral detergent fiber (NDF), acid detergent fiber (ADF), N, P, K, Ca, Mg, Fe, Cu, Mn and Zn.
The chemical analysis of the forage of P. purpureum was done for total aboveground biomass, leaves and stems. In the three sample components of the dry matter, the following parameters were analyzed: CP) and N using the Kjeldahl method (AOAC; 1965), NDF and ADF using the Van Soest method (1963); P using the calorimeter; K using photometry; and Ca, Mg, Fe, Cu, Mn and Zn using spectrophotometer of atomic absorption (Silva, 1999).
The statistical analysis was accomplished using the SAS Ssystem for analysis of variance and application of the Duncan's test for multiple comparison of the means of the variables.
The total height of the tree species Samanea sp. and E. maximum was 10 and 14 meters, canopy diameter of 10 and 16 m, and diameter at breast height (DBH) of 50 cm and 80 cm, respectively. The mean height of the grass was 2,4 m under the tree canopies (shading) and 2,8 m in conditions of full sun.
The CP content of total forage was higher (P>0,10) in the samples of P. purpureum that were under the tree canopies of E. maximum, when compared to the other treatments. There was no difference (P <0,10) among the treatments in the content of NDF and ADF of the total forage among the treatments. The results of the CP, NDF and ADF content of the stems in the forage samples were not different (P>0,10) (Table 1).
The CP content of the leaves were higher (P <0,10) in the forage of P. purpureum under the tree canopies of Samanea sp. and E. maximum, when compared to the leaves of the forage produced in conditions of full sun. The NDF and ADF content of the leaves if the forage produced under the tree canopies of E. maximum was higher (P <0,10) when compared to the other treatments (Table 1).
Table 1. Effect of shading provided by two tree legume species on crude protein (CP), neutral detergent fiber (NDF) and acid detergent fiber (ADF)contents of leaf, stem and total forage produced by Penisetum purpureum in the State of Acre, western Brazilian Amazon. April,2000.
| Species |
Treatment |
CP |
NDF |
ADF |
| % |
||||
| Samanea sp. |
TFUS |
7.19ab |
77.24a |
51.62a |
| Samanea sp. |
TFIFS |
6.09b |
78.25a |
52.87a |
| E. maximum |
TFUS |
8.46a |
77.27a |
51.09a |
| E. maximum |
TFIFS |
5.79b |
80.77a |
54.18a |
| Samanea sp. |
LUS |
8.86ab |
72.75bc |
47.36ab |
| Samanea sp. |
LIFS |
6.93c |
70.98c |
47.60ab |
| E. maximum |
LUS |
9.86a |
76.67a |
49.49a |
| E. maximum |
LIFS |
7.97bc |
74.87ab |
38.37b |
| Samanea sp. |
SUS |
2.94b |
85.40a |
61.50a |
| Samanea sp. |
SIFS |
2.43b |
79.38a |
62.17a |
| E. maximum |
SUS |
7.46a |
77.44a |
60.59a |
| E. maximum |
SIFS |
3.67b |
80.27a |
55.28a |
*Means followed by the same letter in the same column are not statistically different according the Duncan's test, at the level of 10% of probability.
TFUS-total forage under shading, TFIFS-total forage in full sun, LUS-leaves under shading, LIFS-leaves in full sun, SUS-stem under shading, SIFS- stem in full sun.
The N content of the total forage biomass was higher (P <0,10) in the samples of P. purpureum that were under the canopy of E. maximum, when compared to the other treatments. The total forage biomass of P. purpureum under the canopy of E. maximum presented lower (P <0,10) content of K, compared to the other treatments. The results of the contents of P, Ca, Mg, Fe, Cu, Mn and Zn in the total forage biomass were not different under shading and in full sun conditions (Table 2).
The contents of N and Mn of the leaf biomass were higher (P <0,10) in the forage of P. purpureum produced under the shading of Samanea sp. and E. maximum, when compared to the other treatments. The results of the contents of P, K, Ca, Mg, Fe, Cu and Zn in the leaf biomass were not different (Table 2).
Nitrogen, Fe, Mn and Zn contents in the stem biomass was significantly higher (P <0,10) in the forage of P. purpureum produced under shading of E. maximum, when compared to the other treatments. The stem biomass of P. purpureum that was produced under conditions of full sun, close to Samanea sp. presented lower (P <0,10) content of P compared to the other treatments. The results of the contents of K, Ca, Mg and Cu of stem biomass were not different (Table 2).
Table 2. Effect of shading provided by two tree legume species on chemical composition of leaf, stem and total forage produced by Penisetum purpureum in the State of Acre, western Brazilian Amazon. April, 2000.
| Species |
Treatment |
N |
P |
K |
Ca |
Mg |
Cu |
Fe |
Mn |
Zn |
| % |
g/100g |
|||||||||
| Samanea sp. |
TFUS |
0.83b |
0.07a |
1.3a |
0.58a |
0.28a |
3.94a |
449a |
155a |
45.9a |
| Samanea sp. |
TFIFS |
0.85b |
0.09a |
0.94a |
0.61a |
0.27a |
4.44a |
385a |
154a |
43.8a |
| E. maximum |
TFUS |
1.19a |
0.08a |
0.33b |
0.46a |
0.23a |
0.78a |
423a |
210a |
47.0a |
| E. maximum |
TFIFS |
0.78b |
0.08a |
0.96a |
0.92a |
0.36a |
0.50a |
472a |
183a |
40.6a |
| Samanea sp. |
LUS |
1.28ab |
0.09a |
1.15a |
0.66a |
0.31a |
4.15a |
410a |
213ab |
42.5a |
| Samanea sp. |
LIFS |
1.02c |
0.10a |
1.32a |
0.69a |
0.31a |
2.62a |
287a |
131b |
39.3a |
| E. maximum |
LUS |
1.36a |
0.10a |
1.30a |
0.63a |
0.37a |
0.38a |
309a |
274a |
41.2a |
| E. maximum |
LIFS |
1.11bc |
0.08a |
1.30a |
0.67a |
0.36a |
13.1a |
299a |
125b |
83.7a |
| Samanea sp. |
SUS |
0.43b |
0.05b |
1.08a |
0.45a |
0.29a |
2.82a |
144b |
113b |
61.4b |
| Samanea sp. |
SIFS |
0.36b |
0.10a |
1.10a |
0.23a |
0.26a |
11.68a |
122b |
113b |
41.8b |
| E. maximum |
SUS |
1.12a |
0.06b |
0.37a |
0.36a |
0.32a |
8.62a |
337a |
249a |
107,0a |
| E. maximum |
SIFS |
0.55b |
0.06b |
1.10a |
0.27a |
0.27a |
0.59a |
187ab |
129b |
59.0b |
*Means followed by the same letter in the same column are not statistically different according the Duncan's test, at the level of 10% of probability.
TFUS-total forage under shading, TFIFS-total forage in full sun, LUS-leaves under shading, LIFS-leaves in full sun, SUS-stem under shading, SIFS-stem in full sun.
The results of this study agree with those of Wong and Wilson (1980) cited by Pezo and Ibrahim (1998) who reported that moderate shading stimulates the absorption of nitrogen by the grasses. This factor explains the higher CP content in the forage produced by P. purpureum under the shade of trees of Samanea sp. and E. maximum when compared with that produced under conditions of full sun According to these authors, there are cases in which there is also an increase in the amount of biomass produced by the grasses under shading conditions.
Symbiotic nitrogen fixation by Rhizobium and Brady rhizobium bacteria is a mechanism present in 92-95% of the legume species of the Mimosideae and Papilionoideae (Pezo and Ibrahim, 1998). This can result in higher N content in the tree litter, which associated with a more efficient nutrient cycling in the plant-soil system, could be one of the main factors to explain the higher content of nitrogen and crude protein in the forage produced by the grass under the tree canopies.
The recycling of nutrients through the decomposition of the organic matter from of the leaves and branches of the species of tree legumes is one more advantage for the silvipastoral systems. The deep root system of the trees extract nutrients that were leached deep into the soil profile, out of reach of the grass roots. These nutrients are stored in the tree biomass and when the leaves and branches fall in the soil, they are released after the decomposition of the litter and became available for absorption by the trees and the associated grass.
According Carvalho et al., (1999) the crude protein content of the grasses under the tree canopies is maintained during the dry season. This guarantees gains in animal weight even during the dry season in extensive cattle systems. The same does not happen in grasses grown under full sun conditions, because the gradual decrease in the crude protein content results in a stabilization or even the loss of animal weight during the dry season.
The lack of effect of shading on the fiber fractions of the forage produced by P. purpureum is in agreement with the findings of Pezo and Ibrahim (1998).
Silvipastoral systems consisting of associations of the grass P. purpureum with tree legume species such as Samanea sp. and E. maximum have potential to be used in the process of reclamation of degraded areas of pastures, also contributing to increase productivity, profitability and stability of the livestock production systems in the region.
The establishment of sustainable silvipastoral systems can certainly have significant impacts on reducing deforestation pressures and in the use of fire as a tool in pasture management in the Brazilian Amazon.
AOAC - Association of Official Analytical Chemists. (1965) Official methods of analysis. Washington, DC.
Carvalho MM, Barros JC, Xavier DF, Freitas VP, Aroeira FLJM (1999) Composición química del forraje de Brachiaria decumbens asociada con tres especies de leguminosas arbóreas. CIPAV, Memorias Eletrônicas del VI Seminário Internacional Sobre Sistemas Agropecuarios Sostenibles, 1999. Cali, Colombia
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA (1997) Anuário Estatístico, 1996. IBGE, Rio de Janeiro.
Pezo D, Ibrahim M (1998) Sistemas silvipastoriles. CATIE/GTZ, Módulo de enseñanza nº 2, Turrialba, 258p
Silva FC, Org. (1999) Manual de análises químicas de solos, plantas e fertilizantes. Embrapa Comunicação para Transferência de Tecnologia, Brasília, 370p
Valentim JF (1989) Impacto ambiental da pecuária no Acre. Documento base do Curso de Avaliação do Impacto ambiental da Pecuária no Acre, Embrapa-CPAF-AC/IMAC. Embrapa-CPAF/AC, Rio Branco, Acre, 33p
Valentim JF e Moreira P (1994) Adaptação, produtividade, composição morfológica e distribuição estacional da produção de forragem de ecotipos de Panicum maximum no Acre. Embrapa-CPAF/AC, Boletim de Pesquisa, 11. Embrapa-CPAF/AC, Rio Branco, Acre, 17p
Van Soest PJ (1963) Use of detergents in the analysis of fibrous feed. 1: Preparation of fiber residues of low nitrogen content. Journal of the Association of Agricultural Chemists, 46: 825-882
Del Pozo P.P[129]., Irma Jeréz[130], Lucía Fernández[131], P. Padilla[132], J. Ginoria[133]
The study of the establishment of the Leucaena leucocephala like arboreal component associated with gramineous C4, in banks of proteins or in the total area of the grassland has been a topic which approached by Ruíz and Febles (1987) and more recently by Ruiz et at (1997, 1999) which results have made possible the basic knowledge in the handling of the establishment of this species under our present conditions of exploitation. The functional analysis of the growth has demonstrated to be a valuable tool to characterize in a continuous way the growth of the plants starting from the instantaneous values of the derived physiologic indices of the expressions that represent them (Valero et at 1993, Grove 1994, Poorter and Gainer 1996), through which it can be visualised and defined the mechanisms that explain the restrictive action of any of the factors that affect the growth (Del Pozo 1998).
The objective of the present work consists on analysing the growth and development of the Leucaena cv. Ipil Ipil during their establishment through indicative morpho-physiological derived the models that describe their growth in a silvopastoral ecosystem to leave.
The experimental work was developed during the establishment of 2 leucaena há in areas of a Silvopastoral module which belongs to the group Red Ferrallitic Soils (Hernández et at t 1995), with small slopes and contents of organic matter and phosphorus below 2.5 and 10 PPM, respectively. The crop was carried out in fringes of double furrow with a distribution among them of 8 meters. Each fringe is composed of two arrays with a distance of ridge of 1 m and a distribution of 10 12 plants by lineal meter. Before the sowing Carbonate Calcium was applied at rate of 1 t/há and the seeds were inoculated with the specific rhizobium.
A random block design with 4 replications was used, where the experimental treatments were the age, from 42 up to 294 days of sowing with a frequency of every 42 days. In each treatments the height the yield of total dry matter, leaves, stems and theirs relationships, number of leaves, branches was measured. In each replication 10 plants were harvested (5 from each furrow) distributed at random in each treatment. Each variable was characterized statistically and adjusted to the models: Logistical Growth, Gompertz, Richards, Cubic and Logarithmic Polynomial. For the discrimination among models, statistical and physiological approaches were used. To estimate the parameters the method of Marquart (1963) and Minimum Squared was used for the models non lineal and lineal, respectively. Starting from the expressions that described the behaviour of the studied variables was considered the Rate of Crop Growth (RCG).
The height of the plant and their yield of total dry mass increased (p < 0.001) with the age and they were adjusted to the models of Richards and Logistical, respectively (table 1), with values of superior R2 at 0.98. In both cases problems of correlation in the residuals were not presented. This was corroborated in the analytic test of Durbin Watson (DW height, 1.94 and DW yield of total dry matter, 2.30).
The accumulation of the biomass TCC was slow during the first 60 to 70 days, the moment from which increased, with registrations of up to 2.69 g MS/plant to the 129 days (figure 1). In the height, the maximum speed (inflection point) was reached at 56 days with 3.94 cm/day.
In both cases starting from the 180 days, the height and the half yield of each plant began to be stabilised. Similar behaviour presented the yield of its stems, which didn’t vary (p < 0.001) starting from the 168 days (table 2). The accumulation of the biomass followed a different pattern and probably related to changes presented under the climatic conditions and to the own state of growth reached by the plant starting from the 210 days.
Nevertheless, the proportion of the biomass to foliate diminished gradually with the age of the plants, from 78% to the 42 days up to 21% to the 294 days. We should point out that these changes in the pattern of distribution of the biomass were developed in a quick way and apparently associated with the quick speed of growth that presents the species, since there already exist a constant renovation of the foliage in the plant, the biomass of the stems accumulates in a permanent way. There were no changes (p < 0.001) in the number of branches and leaves for plant starting from the moment where the maximum speed of accumulation of total dry mass is reached in the plant helps to explain this behaviour. Under the studied conditions, starting from the 129 days (4.3 months) it can begin the exploitation of the leucaena in the ecosystem, which presents a height of 197 cm, yield of total dry mass of 103.4 g MS/plants and a foliage percentage in the superior biomass to 40%. The model Richards and Logistical they showed potentialities to characterize the growth in height and yield of total dry mass for plant of the leucaena.
Del Pozo, P.P. 1998. Análisis del crecimiento del pasto (Cynodon nlemfuensis) bajo condiciones de corte y pastoreo. (Tesis de Doctorado). La Habana: ICA, UNAH.
Poorter, H y Gainer, E. 1996.- Plant growth analysis and evaluation of experimental desing and computational methods. Journal of experimental Botany 47.
Ruiz, T.E. y Febles, G. (1987). Leucaena, una opción para la alimentación bovina en el trópico y subtrópico. EDICA. Cuba..
Ruiz, T.E.; Febles, G.; Castillo, E.; Bernal, G. y Díaz, L.E. (1997). Establecimiento de Leucaena leucocephala intercalada en diferentes pastizales de gramíneas.Rev. cubana Cienc. agríc. 31:91.
Ruiz, T.E.; Febles,G.; Castillo, E.; Jordan,H.; Crespo, G.; Valenciaga, N. Y Diaz, H. 1999.- La Experiencia Cubana en la Agronomía y Manejo de Leucaena leucocephala. Primer Congreso Latino Americano de Agroforestería para la Producción Animal Sostenible. Cali, Colombia 28 al 30 de Octubre1999. CDROOM, CIPAV\Agrof99\P-DPozo.htm. ISBN 958-9386-22-9.
Valero, J.J.A.; Martín, F.O.M. y Fabeiro,C.C. 1993.- Estudio comparativo de dos métodos de análisis de crecimiento clásico y funcional en un cultivo de soja sometido a riegos diferenciados. ITEA. 89(3): 166.
Table 1. - Models that describe the growth of the Leucaena leucocephala in the ecosystem.
|
Variable |
Models |
R2 |
Sig. |
|
Height |
210.29/(1 + 4.77e -0.043edad)^3.301 |
0.99 |
*** |
|
Yield |
211.16/(1 + 735.09e - 0.051edad) |
0.98 |
*** |
* * * p < 0.001
Table 2. Behavior of some growth indicators of Leucaena in the ecosystem.
|
Age (days) |
Yield, g MS/plant. |
Number of: |
||
| Leaves |
Stems |
Branches/Plants |
Leaves/Plant |
|
| 42 |
1.00a |
0.41ª |
1.00a |
10.02ª |
| 84 |
15.75b |
18.10ª |
5.82ª |
35.07ª |
| 126 |
30.66c |
51.03b |
15.53b |
103.60b |
| 168 |
31.57c |
162.33c |
21.16b |
130.42b |
| 210 |
55.81d |
166.48c |
16.56b |
126.53b |
| 252 |
38.04c |
160.80c |
22.73b |
139.88b |
| 294 |
31.91c |
178.68c |
22.98b |
130.98b |
| 336 |
57.62d |
170.72c |
21.22b |
137.96b |
| IT IS ( |
4.85*** |
9.60*** |
2.42*** |
13.98*** |
a,b,c,d. Value with non common letters in columns for treatment they differ p < 0.05, Duncan (1955)
* * * p < 0.001.
Humberto González Rodríguez[134] and Israel Cantú Silva[135]
Key words: drought, osmotic potential, soil water, water potential, water stress
The subtropical thornscrub dry woodlands of northern Mexico are dominated by a diverse, dense and spiny shrubs. These woodlands, mixtures of deciduous and broad-leaved evergreen shrubs (McMurtry et al., 1996), are characterized by a wide range of growth patterns, diverse leaf life spans, and growth dynamics with contrasting phenological developments. The farmers of the northeastern region have developed a self-sufficient, subsistence economy in which the thornscrub is exploited for diverse resources such as forage for animals, fuelwood, timber for construction, fences, foodstuffs, herbs, medicine, and revegetation in disturbed sites. Moreover, these plants provide wildlife habitat and cover to prevent soil erosion. In addition, these native plants have evolved various morphological and physiological traits suited for adaptation to adverse environmental constraints, especially on drought-prone sites. Although genetic variation for physiological mechanisms to avoid plant tissue dehydration have been documented in crop plants (Morgan, 1984), to date little evidence is available on plant water relations for shrub species of the northeastern region of Mexico. Therefore, this region provides an opportunity to investigate the physiology of native shrub species and their responses to changes in resource availability, particularly soil moisture content. The present study was undertaken in ten native and dominant shrub species to quantify how diurnal and seasonal plant water potentials are related to soil water availability and to describe if osmotic adjustment occurs among the species as a potential mechanism of adaptation to drought stress.
Study area
This study was carried out at the Experimental Research Station of the Faculty of Forest Sciences of the Autonomous University of Nuevo Leon (24o47’ N; 99o32’ W, at an altitude of 350 m a.s.l.) located 8 km south of Linares county, in Nuevo Leon state of Mexico. The climate is typically subtropical and semi-arid with a warm summer. Mean monthly air temperature ranges from 14.7oC in January to 22.3oC in August. Average annual precipitation is 805 mm.
Plant material, leaf water and osmotic potential measurements
To achieve the proposed objectives, ten shrub species were evaluated in two different studies. In the fist study, named herein as the pure stand experiment, the following plant species were chosen: Acacia berlandieri Benth. (Fabaceae), Pithecellobium ebano (Berl.) Muller (Fabaceae), Codria boissieri DC. (Boraginaceae) and Helietta parvifolia (Gray) Benth. (Rutaceae). Each shrub species was growing as a pure stand in already-established field plots. The study plot of each shrub species was laid out in 8 by 8 m, with a spacing between plants of 2 m. In the second study, named herein as the mixed stand experiment, six co-existing shrubs (Pithecellobium pallens (Benth.) Standl. (Fabaceae), Acacia rigidula Benth. (Fabaceae), Eysenhardtia polystachya (Ort.) Sarg. (Fabaceae), Diospyros texana Scheele (Ebenaceae), Randia rhagocarpa Standl. (Rubiaceae), and Bernardia myricaefolia (Scheele) Watts. (Euphorbiaceae) were selected from a 20 by 20 m natural, representative and undisturbed thornscrub plot.
In both studies, determinations of leaf water potentials were at 14 days intervals. At each sample period, four (replications) representative, permanent and tagged plants were randomly chosen. On each sampling date, leaf water potential (Yw) measurements were taken from terminal twigs and monitored at 06:00 (predawn) and 14:00 (midday) h local time for each tagged shrub during the experimental period. Yw was estimated using a Scholander pressure bomb. Simultaneously, at the time of leaf excision to measure Yw, four mature leaf discs (0.19 cm2 each) were sampled from each of the four plants to determine leaf osmotic potential (Yp). Enviromental variables such as air temperature (oC), relative humidity (%), precipitation (mm), air saturation deficit (KPa), and gravimetric soil water content were also registered.
Seasonal variability in leaf water potential
Seasonal patterns of predawn leaf water potentials for shrub species evaluated at the pure and mixed stand experiments are shown in Figure 1. During the experimental period of the pure stand experiment, two dry down periods were registered; the first occurred between July 2 and September 26, 1997 and the second, and more severe one, was registered between December 17, 1997 and August 13, 1998, both of which caused a drop in dawn water potential (Figure 1A). Averaged predawn water potentials during the wettest period (from April 23 to June 18, 1997) ranged between -0.18 and -0.68 MPa for C. boissieri and P. ebano, respectively, while during the extended and unusual driest period (from February to mid July, 1998) averaged predawn water potentials ranged from -5.0 to -7.3 MPa for A. berlandieri and H. parvifolia, respectively (Figure 1A). These results were associated to the gradual depletion of soil moisture content as the study progressed, rainfall events of low intensities, and high ambient air temperatures (data not shown). Similarly, during the course of the mixed stand experiment, two dry periods were observed: the first occurred between November 12, 1998 and April 29, 1999 and the second one from July 27 to September 3, 1999, both of which caused a drop in dawn water potential (Figure 1B). Although both dry periods are related to low precipitation, the first one is associated to low temperatures whereas, the second to high temperatures. Maximum (-0.10 MPa) and minimum (-0.27 MPa) predawn leaf water potentials values during the wettest period were observed in R. rhagocarpa and A. rigidula, respectively. In contrast, during the driest period, maximum (-4.9 MPa) predawn leaf water potentials were experienced in A. rigidula whereas, minimum values ranged between -6.3 and -6.7 MPa for D. texana and R. rhagocarpa, respectively (Figure 1B).
During the two dry periods registered in the mixed stand experiment, B. myricaefolia and A. rigidula were the most drought sensitive and tolerant shrub species, respectively, to soil water depletion. In fact, as the soil water stress develops, there is a sharp leaf water potential decline in B. myricaefolia compared to the other five shrubs. Moreover, while A. rigidula attained predawn leaf water potentials values in the range of -4.0 to -5.5 MPa, leaf water potentials in B. myricaefolia could not be measurable since its values were far below of the limit of calibration of the pressure bomb, which in this study was -7.3 MPa. Thus, this sensitive shrub species experienced the greatest seasonal fluctuation in dawn water potentials.
Figure 1. Seasonal predawn
leaf water potentials (yw) of shrub species
monitored at the pure stand (A; between April 23, 1997 and September 22,
1998)
Figure 1. Seasonal predawn leaf water potentials
(yw) of shrub species monitored at the mixed stand
(B; from September 10, 1998 to September 11, 1999) experiments at the research
site.
The seasonal fluctuations in predawn leaf water potentials in both studies showed a considerable response to precipitation, which in fact determined the soil water availability pattern. Other factors associated to variations in dawn water potentials are the physical properties of the soil. For instance, during the driest season of the pure stand experiment, plant water stress was greatest in H. parvifolia since its shallow root system (according to field observations) was confined in the first 30 to 40 cm soil depth profile, and because this shrub species was growing in the finest textured soil (data not shown). In contrast, the other three species had relatively higher water potentials, presumably due to coarser textured and deeper soils observed. Another possible factor related to high plant water potentials during dry periods, (as was observed, for example, in A. berlandieri, P. ebano, P. pallens, and A. rigidula) is probably due to the presence of deep taproots of these species, which apparently their root system would allow access to a greater volume of soil water as has been documented by Monson and Smith (1982). As in the present study, similar predawn leaf water potential responses have been observed in co-existing shrubs such as Larrea tridentata (Montaña et al., 1995), Prospis galndulosa (Montaña et al., 1995; McMurtry et al., 1996), Berberis trifoliata, Diospyros texana, and Zanthoxylum fagara (McMurtry et al., 1996), subjected to soil moisture stress. Two basic physiological processes might be considered in response to low water potential: a) it could be proposed that the rigid structure of the leaves would prevent dehydration-induced cell collapse leading to the conclusion that osmotic adjustment (passive and active) may have occurred at least to a value of leaf water potential for the maintenance of a positive turgor potential, and b) perhaps an increase in the elastic properties of the leaf cell walls (i.e., indicating a decrease in the bulk tissue elastic modulus) could have contributed to turgor maintenance more than changes in solute potential, which in some cases seems to be a high change, between -3.0 and -4.0 MPa. In this regard, it has been suggested that these two mechanisms have the same effect in different plant species, to avoid the mechanical stress on the protoplasm layer and especially the cell membrane (Monson and Smith, 1982; Meier et al., 1992).
Leaf osmotic potential
Decreases in the leaf osmotic potential through accumulation of solutes under water stress have been documented as a physiological criterion for adaptation to drought (Montaña et al., 1995). Changes in predawn leaf osmotic potential at the mixed stand experiment were evaluated under high (0.238) and low (0.138) soil water content (kg kg-1). Under high soil moisture content, maximum (-1.16 MPa) and minimum (-2.09 MPa) osmotic potential values were observed in B. myricaefolia and A. rigidula, respectively, whereas under low soil water content, osmotic potential values ranged from -2.74 (E. polystachya) to -4.04 MPa (D. texana and B. rhagocarpa). If it is assumed that leaf water potential values under adequate and limited soil moisture content represent a true equilibrium with soil water potential (not measured in this study), the above results suggest that the capacity for osmoregulation, expressed through a total variation in leaf osmotic potential changes between the wet and dry period, ranged from -2.65 (B. myricaefolia) to -1.11 MPa (P. pallens). The seasonal adjustment in leaf osmotic potential measured in this study is within the range of values reported for studies on native plant species from mesic (Montaña et al., 1995) and desert habitats (Monson and Smith, 1985).
Soil-Plant water relations
The relationship between leaf water potential and soil water content for the six co-existing shrub species studied at the mixed stand experiment is showed in Figure 2.
Water potential values were positively and curvilinearly related to the average soil water content in the 0-50 cm soil depth. The regression analysis showed that the best seasonal relationship between predawn water potential as a function of average soil water content was described by a power mathematical function (data not shown). At the pure stand experiment, average soil water content in the 0 to 50 cm soil depth profile explained between 65 and 77% of the variation in leaf water potential, in contrast, at the mixed stand experiment, this relationship ranged between 66 and 87%. Moreover, it was found that gravimetric soil water content values above 0.17 (pure stand experiment) and 0.15 kg kg-1 (mixed stand experiment) predawn water potential values were high and roughly constant; below these threshold values, water potentials declined gradually.
Figure 2. Relationship between predawn leaf water potentials
(?w) of six co-existing shrub species and average soil water content monitored
at the mixed stand experiment. Data are for the period September 10, 1998 to
September 11, 1999 at the research site.
On a seasonal basis, all shrub species showed basically the same response of predawn leaf water potential to changes in soil moisture.
Shrub species fall into three main groups. i) highly responsive (Helietta parvifolia, Cordia boissieri, Randia rhagocarpa, and Bernardia myricaefolia), ii) moderately responsive (Pithecellobium pallens, Eysenhardtia polystachya, and Diospyros texana), and iii) slightly responsive (Acacia berlandieri, Pithecellobium ebano, and Acacia rigidula) to changes in soil water content.
Seasonal patterns in plant water potentials could be explained by the soil water availability in a range from 65 to 87%, which in turn would depend on the precipitation trend.
Predawn water potential values between the wettest and driest period ranged from -0.5 to -7.3 MPa for the ten shrub species examined.
Since osmotic adjustment is related to water stress tolerance, this physiological mechanism was observed in shrub species which experienced gradual soil water depletion. Osmotic adjustment values ranged between -1.11 and -2.65 MPa.
Under high soil water content, differences among shrub species for diurnal changes in leaf water potential might be controlled, presumably, by the capacity of the leaf tissue to control the rate of water loss through the transpirational flux, whereas under low soil water content, differences could be related to the capacity of the root system to extract water from deeper soil layers.
This type of studies may allow us to identify patterns of water uptake and partitioning between woody plant species, to understand plant successions processes, and to chose tolerant species to drought which could be used in silvopastoral systems.
This research was funded by the International Foundation for Science (IFS, grant D/2610-1). Valuable field technical assistance provided by Elsa González Serna is gratefully acknowledged.
Hesla BI, Tieszen LL and Boutton TW (1985) Seasonal water relations of savanna shrubs and grasses in Kenya, East Africa. Journal of Arid Environments 8: 15-31
McMurtry CR, Barnes PW, Nelson JA and Archer SR (1996) Physiological responses of woody vegetation to irrigation in a Texas subtropical savanna. La Copita Research Area: 1996 Consolidated Progress Report, pp 33-37. Texas Agricultural Experiment Station - Corpus Christi, Texas A&M University System, College Station, Texas, USA
Meier CE, Newton RJ, Puryear JD and Sen S (1992) Physiological responses of loblolly pine (Pinus taeda L.) seedlings to drought stress: osmotic adjustment and tissue elasticity. Journal of Plant Physiology 140: 754-760
Monson RK and Smith SD (1982) Seasonal water potential components of Sonoran Desert plants. Ecology 63: 113-123
Montaña C, Cavagnaro B and Briones O (1995) Soil water use by co-existing shrubs and grasses in the southern Chihuahuan Desert. Journal of Arid Environments 31: 1-13
Morgan JM (1984) Osmoregulation and water stress in higher plants. Annual Review of Plant Physiology 35: 299-319
Raúl Erick Mederos, José Ortiz, Verena Torres,
Gustavo Crespo, Idalmis Rodrígues
Key words: computers, nutrients, programming, recycling
Among nutrients that are given more importance in the studies of soil-plant-animal ecosystem are Nitrogen (N), Phosphorus (P) and Potassium (K) because they actively influence on nutrient recycling. Many studies aim at quantifying the amount of such elements input or output of the system following a systemic approach. Knowing the balance of nutrients is of a great importance to carry out an adequate management of ecosystem resources (Dijk et al, 1196, Shiyomi et al., 1992) for farmers could develop strategies to guarantee nutrients flow balance at the level of each particular component and the system in general.
For these reasons, the current study was developed, having as objective to create a software that were able to predict annual nutrients balance both at system level and at component level for Cuban cattle pasture conditions. This was carried out, based on the simulation of the most relevant biological processes that take place within the ecosystem. To develop this software we used Windows 95 as operating system and Borland Delphi version 5.0 as programming language.
From the methodology described by (Ortiz, 2000) we created a simulation model of N, P, K balance in cattle pasture ecosystem. It was necessary to make an abstraction of the real system and take into consideration all affecting processes and factors.
A lgorithm design permits the adaptability of the application to different productive conditions, independently of the management technology, productive aims, available resources, herd characteristics and other factors.
For the creation of the software we followed the Analysis and Design Oriented to Object of Information System (ADOOSI) because object-oriented programming offers a great potential due to the code re-utilization. As operating system Windows 95 (or a superior version) was defined for facilities it offers from a graphic perspective and for the friendliness of its applications (Senn, 1998). Borland Delphi 5.0 was used as programming language because this system supports object-oriented programming and is highly potent for the development of friendly interfaces and the work with a great amount of objects as lists, databases, among others.
A software able of representing estimations of inputs and outputs was obtained, considering the main sources of N, P, and K in the ecosystem. In our conditions, it is possible to forecast their contributions for a year period. The conceptual structure of nutrient recycling defined considers the three components of the ecosystem: Soil, Plant and Animal and also different ways of nutrients input and output.
Users can evaluated up to12 possible alternatives of a case study, making variations to some factors that intervene in the model. Some of these factors are the number of animals in the herd, live weight gain or loss, amount of milk liters, number of births and animal removals of the system (sales and deaths) and also pasture areas, including surface occupied by pastures, could be changed by the user, according to the percent of grass and legume utilization and of pasture species that occupies such area. Factors like excretions percent in pasture areas, supplementation consumed by animals and soil fertilization can also be modified by users.
To carry out the already mentioned operations, the system has a menu with five main options:
Files
Edit
Balance
Databases
Help
In the Files option, Windows standard operations like creating, opening, closing, saving, and printing a project, or exiting the application are carried out. In the Edit option, alternatives can be inserted or eliminated, general project data can be changed, among other functions. In the Balance option, nutrients balance is executed. In the Databases option, fifteen databases included in the system are manipulated, among such bases are: nutrients, pastures and fertilizers. These databases can be modified by the user according to his needs. In the Help option, user is given the necessary information to operate the system.
The result of this system is a N, P, K balance of each component and of the system in general, where inputs and outputs of nutrients are analyzed separately and in details and the deficit or excess of some of them are specified, as observed in table 1.
Table 1. Result of the balance of nutrients, analyzing its components.
|
SOIL |
|
Inputs: |
N |
P |
K |
|
Rain |
15.9 |
0.00 |
0.00 |
|
Litter |
1.0 |
0.1 |
1.0 |
|
Excretions (in pasture areas)- N volatilized |
89.3 |
15.4 |
104.9 |
|
N biological fixation |
20.0 |
0.00 |
0.00 |
|
Fertilizers |
--- |
--- |
--- |
|
Inputs total |
126.4 |
15.6 |
105.9 |
|
Outputs: |
|
|
|
|
1. Pasture Extraction (10% N for legumes) |
161.1 |
29.8 |
169.9 |
|
Outputs total |
161.1 |
29.8 |
169.9 |
|
¿Deficit in the Soil? |
-34.7 |
-14.2 |
-63.9 |
|
PLANT |
|
Inputs: |
N |
P |
K |
|
1. Soil Extraction (only 10% N for legumes) |
161.1 |
29.8 |
169.9 |
|
Inputs total |
161.1 |
29.8 |
169.9 |
|
Outputs: |
|
|
|
|
1. Animal consumption |
112.7 |
20.9 |
118.9 |
|
2. Litter |
1.0 |
0.1 |
1.0 |
|
Outputs total |
113.7 |
21.0 |
119.9 |
|
¿ Deficit in the Plant? |
47.7 |
8.8 |
50 |
|
ANIMAL |
|
Inputs: |
N |
P |
K |
|
1. Pasture consumption |
112.7 |
20.9 |
118.9 |
|
2. Supplement consumption |
0.6 |
0.1 |
0.00 |
|
Inputs total |
113.3 |
30.0 |
118.9 |
|
Inputs: |
|
|
|
|
1. Animal product: milk, LW gain and births |
11.1 |
3.9 |
2.4 |
|
2. Excretions |
102.2 |
17.1 |
116.5 |
|
3. Death or Sale |
- |
- |
- |
|
Outputs total |
113.3 |
30.0 |
118.9 |
|
¿Deficit in the Animal? |
0 |
0 |
0 |
|
SYSTEM |
|
Inputs: |
N |
P |
K |
|
1. Rain |
15.9 |
0.00 |
0.00 |
|
2. N biological fixation |
20.0 |
0.00 |
0.00 |
|
3. Supplementation |
0.6 |
0.1 |
0.00 |
|
Inputs total |
36.5 |
0.1 |
0.00 |
|
Inputs: |
|
|
|
|
1. Animal product: milk, LW gain and births |
11.1 |
3.9 |
2.4 |
|
2. N volatilization of deposited excretions in the pasture areas |
2.6 |
0.00 |
0.00 |
|
3. Excretions out of the pasture area |
10.2 |
1.7 |
11.6 |
|
4. Death or Sale |
- |
- |
- |
|
Inputs total |
23.3 |
5.6 |
14 |
|
¿Deficit in the System? |
13.2 |
-5.5 |
-14.0 |
The algorithm of simulation allows to increase accuracy in estimations and save time.
The software has a friendly interface and requires little computers knowledge for its use.
By means of the software we could predict ecosystems performance and create alternatives for maintaining balance.
The software possesses databases of conventional and non-conventional feed.
Authors would like to thank the collaboration of Lic. Pedro J. Padilla in the translation and revision of this work.
Cárdenas S, 1997. Metodología de Análisis y Diseño Orientada a Objetos de Sistemas Informáticos. ISPJAE
Crespo, G., Rodriguez, Idalmis, Torres, Verena, Ortiz, J. y otros. 1998. Estudio del reciclado de los nutrientes en sistemas de producción vacuna. Informe final de proyecto. ICA.
Dijk, J.; Leneman, H. and Marianne van der Veen. 1996. The nutriment flow model for Dutch Agriculture: a tool for environmental policy evaluation. Journal of Environmental Management. 46:43-55.
Ortiz, J. 2000. Modelación y Simulación del reciclaje de N, P, K en sistemas de pastoreo vacuno. ICA
Senn, J.A. 1998. Analysis & design of information systems. McGraw-Hill, Inc., USA.
Shiyomi, M; Kirita, H. and Takahashi, S. 1992. Energy, nitrogen, phosphorus and carbon budgts at plant, animal and ecosystem levels in grazings grasslands in the Nishinasuno Area, Japan. Ecological Processes in Agro-Ecosystems. NIAES. Series No. 1: 173-188.
Benjamin Name[136], José Villarreal
Acacia mangium (Willd.) is a robust leguminous tree, easily established in plantations. In fertile soils, it has high survival and good initial growth, although inferior to other fast growing trees. However, in degraded, compacted, acid (pH lower than 5.5) soils with high aluminum contents it has overcome other species (CATIE, 1986). Experimental data of IDIAP, in acid Ultisols, classify it with better behavior, when comparing with other native and exotic species, such as Tectona grandis, Eucalyptus camandulensis, Genipa americana (Name, Cordero and Bernal, 1987). The grass Brachiaria humidicola was introduced and selected as one of those of better adaptability under conditions of acid soils, deficient in macro and micronutrient (Urriola et. al. 1985). The objective of the present work, was to determine the effect of the association Acacia mangium-Brachiaria humidicola on an acid ultisol of Panama by means of the quantification of the variability and behavior in the soil parameters: quantity of organic matter, roots, variation in the number of soil organisms, infiltration, density, natural fertility and foliar content of elements of the plants in the subsystems under evaluation.
The experimental area is located (8º 15’ N 81º 5’ W) at an elevation of 100 meters above sea level, with an average precipitation of 2500 mm/yr and yearly average temperature of 27 ºC. The rainy period runs between May to December. The soil is a Typic Plinthudult, fine, mixed, isohypertermic, deep, acid, of subangular blocks structure, of low cationic exchange capacity (14 cmol/kg; average of the profile).
To measure the different parameters plots planted sequentially in different years (1990-1996) with A. mangium associated with natural grasses or Brachiaria humidicola were selected, under different management conditions. The soil sampling to determine organisms was carried out at 10cm. of depth. For every planting year and management, four soil samples were taken for analysis and organism count. For the case of soil fertility, all the analyses of the association A. mangium - B. humidicola (16 samples) were grouped and the averages were compared with those of the natural association. For soil profile root determination, a square frame of 0.25 m2 was used (soil volume of 0.025 m3). The parcels of A. mangium planted in 1990 had 3 meters between plants and eight meters between rows in an area of about 5000 m2. Soil samples were submerged in a 110 liters water tank where the thick roots were separated by decantation and later on filtered the finest ones. After air drying, the root dry matter weight was reported.
For the foliar analyses, the 4th,, 5th and 6th leaf of the branches of the unpruned superior third of the crown, in parcels of A. mangium of 2 to 5 years with different managements were selected at random. In the case of Curatella americana, the same leaves in the branches over 1.5 meter high were selected.
Bulk density determination was based on two replicas. Infiltration was measured by means of the double cylinder method with two field readings to report the speed of infiltration and the sheet infiltrated. The soil organic matter was determined using the methodology proposed by Walkley-Black. Variance analysis, for the foliar content of elements and for the quantity of soil organisms were made with the SAS statistical package.
The quantity of soil organisms detected in the association A. mangium - B. humidicola was affected by the time of planting of A. mangium; the older the association, the higher the organism content, especially earth worms (TabIe 1).
Under these conditions, the lime and phosphoric rock application, and the grazing seemed to not affect the quantity of soil organisms. The areas under native grass and Curatella americana (chumico) present significantively smaller contents of soil organisms (Test of Duncan).
The A. mangium planted between 1990 and 1994 have a higher content than the ones planted after that time, or those that stayed with the native association. A possible explanation is that with time, the soil with A. mangium improves, especially due to the great leaf acumulation, meaning a bigger organic matter content and humidity retention, ideal microclima for the soil organisms, specially the common earth worm (Lumbricus terrestris).
When comparing the land planted with Acacia mangiun (two years age) and that of natural vegetation, 59% increments were detected in the accumulated infiltration sheet after three hours and an increase in the mean speed of infiltration (Table 2).
In the land with. A. mangium + B. humidicola and different management, after only two years of the association, the bulk density decreases iby 10%, when compared with the land in the natural vegetation (Table 3). In the fourth year, the land with the association under study, but with cattle grazing, shows an increment in the density with regard to the two year old association, effect of compaction due to trampling by the animals. When comparing the fourth year of association, with the natural parcel, a tendency to decrease the density is observed.
Table 1. Soil organisms in Acacia mangium plantations under different management.
|
Planting year of A.mangium and treatment |
Soil organisms in 0.025 m3. |
|
A. mangium associated with B. humidicola, 1990 |
10.8 A |
|
A. mangium associated with B. humidicola, 1992, with lime. |
10.5 A |
|
A. mangium associated with B. humidicola., 1994, Under grazing. |
9.5 A B |
|
A. mangium associated with native grass, 1993 + 600 kg/ha F.R. |
6.5 A B C |
|
A. mangium associated with native grass, 1993 +0 kg/ha oC FR |
3.5 B C |
|
A. mangium associated with B. humidicol., 1995. |
2.7 C |
|
Area under native grass and Curatella americana |
2.3 C |
VC = 51.38 R2 =0.64; means of four repetitions
The main quantifiable effect of the association A. mangium + B. humidicola, is reflected with the organic matter analysis. In the first 15 cm of depth, 4.3 and 3.8% were detected for the association planted in 1990 and 1993, respectively (Table 4). This is highly significant if we compare the means with the organic matter content of areas with natural vegetation of chumico (Curatela americana and Andropogon bicornis). The soil organic matter can act as a liberator or accumulator of atmospheric carbon, depending on the management that is provided to the system.
When associating the Tables 3 and 4, it can be seen that the decrease of the bulk density is associated to the increase of the soil organic matter. No relationship between the content of soil organic matter and lime application was detected.
Table 2. Water infiltration under different management.
|
Treatment |
Time of infiltration (minutes) |
Infiltrated sheet (cm) |
Mean speed infiltration (cm/hr) |
|
Land planted with A. mangium for two years. |
180 |
12.4 |
4.1 |
|
Land under natural vegetation. |
165 |
7.8 |
2.8 |
Table 3. Soil bulk density under different management.
|
Depth (cm) |
Land planted with A. mangium for two years. |
Land planted with A. mangium four years and grazing. |
Land under natural vegetation. |
|
0 - 20 |
1.09 |
1.17 |
1.20 |
Table 4. Soil organic matter content in associations of Acacia mangium Brachiaria humidicola planted in different years and management.
| Depth (cm) |
1990 |
1993 |
Natural Association |
Lime Effect |
|
| With |
Without |
||||
| 0-15 |
4.3 |
3.8 |
2.7 |
4.4 |
4.1 |
| 15-30 |
3.2 |
2.2 |
2.3 |
2.8 |
2.2 |
Roots procedure were detected to 70 cm. depth in the A. mangium association versus 30 cm. in the natural association (Table 5). The root weight in the first 10 cm is higher by more than 250% in the A. mangium - B. humidicola association.
Table 5. Root weight at different depths under two associations in a soil volume of 0.025 m3.Calabacito 1996.
|
Depth (cm) |
Association A. mangium + B. humidicola (g) |
Native association C. americana + natural pastures (g) |
|
0-10 |
1136.4 |
454 |
|
10-20 |
85.1 |
56.7 |
|
20-30 |
42.5 |
28.4 |
|
30-40 |
70.9 |
Non quantifiable |
|
40-50 |
28.4 |
Non quantifiable |
|
50-60 |
28.4 |
Non quantifiable |
|
60-70 |
28.4 |
Non quantifiable |
The soil fertility analysis of the association A. mangium + B. humidicola and the natural association of the area only detected differences in the content of organic matter (Table 7). The levels of P, K, Ca and Mg were deficient in both systems, and practically the same. A darker soil color and the development of a granular structure in the first soil centimeter of depth was observed in the field. The actual conventional methodology of soil analysis is probably not sufficiently precise to detect changes in the chemical parameters in the period in which the study was developed. According to Sánchez (1995), precise methodologies have still not been developed to evaluate the capacity of systems to recover. This well-known concept, the soil resilence or the agroforestal system resilience, should be implemented in economic, social and ecological aspects.
Table 6. Soil fertility change
| Depth (cm) |
Association |
P |
K |
Ca |
Mg |
M.O |
Mn |
Fe |
Zn |
Cu |
| mg 1-1 |
cmol kg-1 |
% |
mg 1-1 |
|||||||
| 0-15 |
B. humidicola + A. mangium |
1 |
20 |
Tr |
tr |
4.8 |
38 |
26 |
Tr |
5 |
| 0-15 |
Natural association association |
1 |
28 |
Tr |
.1 |
2.7 |
29 |
13 |
Tr |
2 |
When comparing the foliar mineral content in leaves of A. mangium of different ages with that of the species C. americana, as the leaf of A. mangium ages, the levels of N decreases; but this content excedes significantly the levels found in the species C. americana. (Table 7) The contribution of K in the leaf of C. americana is significantly bigger that in the A. mangium. In the case of Ca, the foliar content of limed A. mangium is higher than those not limed but in general it is similar to the content in C. americana.
Table 7. Foliar analysis of leaves of A. mangium and C. americana.
|
|
Content in% |
||||
|
|
N |
P |
K |
Ca |
Mg |
|
Leaves of A. mangium 2 year old |
1.4 A |
002 C |
19 B |
83 C |
49 BC |
|
Leaves of A. mangium 3 years + lime |
1.2 B |
060 B |
19 B |
2.3 A |
76 A |
|
Leaves of A. mangium 3 years - lime |
1.2 B |
120 A |
25 B |
1.4 B |
56 B |
|
Leaves of A. mangium 5 years old |
0.9 C |
103 A |
20 B |
1.4 B |
46 BC |
|
Leaves of C. americanaNatural association |
0.6 D |
118 A |
59 A |
1.7 B |
39 C |
* Means followed by same letter are not significantly different at 5 % level.
When changes in parameters such as soil organism content, organic matter content, quantity of roots, infiltration, soil color and the structural arrangement are evaluated, improvements in the natural fertility of the soil can be detected. The agroforestal systems can play a role of importance in the accumulation of biomass and carbon in an ecosystem. The increase in the infiltration speed improves the hydrological efficiency of the watershed. This parameter indirectly indicates a better use of the effective rain that could increase the hydrological yield of a watershed (decreased flood risk, increase base flow in the dry season). It is recommended to adapt and standardize an analysis methodology to detect the improvement of the fertility of the land (physical and chemical aspects). It is also recommended to design an integral system of land evaluation that the economic efficiency of the hydrological, forest, silvículture, soil fertility and ecological components.
CATIE. 1986. Forestry of promissory species for the firewood production in Central America. Technical report Nº 86.
NAME, B.; CORDERO, A.; BERNAL, J. 1987. Alternatives for use and handling of acids in Panama. In: I Compendio de Resultados de Investigaciones IDIAP, Region Central.
SANCHEZ, P. A. 1995. Science in Agroforestry. Agroforestry Systems 30: 5-55, 1995. Kluwer Academic Publisher.
URRIOLA, D.M.; ARGEL, P.J. AND ORTEGA, C.M. 1985. Estudio agronómico de 21 ecotipos de Brachiarias en el período de establecimiento. Gualaca. En: Resúmenes análíticos de la investigación pecuaria en Panamá. (1968 - 1985). IDIAP.
Pekka Nygren[137],
Frank Berninger[138], Eero Nikinmaa[139],
Risto Sievänen[140] and Pablo
Cruz[141]
Key words: biomass production, Gliricidia sepium, nitrogen fixation, nitrogen yield, pruning
Tree fodder is harvested by means of periodic prunings. The cutting interval of fodder trees is usually quite short, because the proportion of nitrogen-rich foliage out of total harvest is high in the young resprouts (Catchpoole and Blair, 1990), and the nutritive value of the young leaves is high (Mochiutti, 1995). Short cutting interval also reduces forage loss in litterfall. The tree regrowth following the pruning depends on the reserves of non-structural carbohydrates (Erdmann et al., 1993; Tschaplinski and Blake, 1994), and the pruning interval must be long enough to allow for the regeneration of these reserves. Pruning of dinitrogen fixing legume trees, which are common in tree fodder production, causes turnover of nodules and subsequent cessation of N2 fixation (Nygren and Ramírez, 1995). The dieback and renodulation rate seem to depend on pruning intensity (Nygren and Cruz, 1998). In this paper, we used a process-based tree growth model, which takes into account the reserve carbohydrate and N2 fixation dynamics (Berninger et al., 2000), for evaluating the effects of four pruning intensities on 10-year productivity and N yield of Gliricidia sepium (Jacq.) Kunth ex Walp., a legume tree common in silvopastoral systems throughout the tropics (Simons and Stewart, 1994).
The model applied in this study simulates C and N substrate (mobile sugars and free amino acids, respectively) pools and flows, and the conversion of the substrates to structural dry matter. A complete account of the model is given by Berninger et al. (2000). Photosynthesis is the C source, and N assimilation is divided into NH4+ and NO3¯ uptake and N2 fixation. The C and N substrate transport between tree sources and sinks is proportional to the concentration differences between them, according to the transport-resistance approach (Thornley, 1972; 1991). The C substrate sinks are maintenance and growth respiration, accumulation of reserves and biomass growth. N substrate is consumed in growth. Senescence drains both C and N from the plant while retranslocation of N from old leaves is a N substrate source. Biomass is divided into six compartments: leaves, branches, stem, coarse roots, fine roots and nodules. Differing from the original transport-resistance approach (Thornley 1972), each plant compartment draws its C and N needs from an aboveground or a belowground pool of C and N substrate or C reserve. The pipe model approach (Shinozaki et al., 1964; Mäkelä, 1986) is used to allocate the available aboveground C and N substrate pools for the growth of non-productive (stem, branches) and productive (leaves) tissues. The present model version is parameterised for G. sepium, but the model structure is applicable with minor modifications for other N2-fixing trees as well.
The model parameters were set to correspond observed growth of G. sepium in a cut-and-carry forage production system in the experimental farm of the French National Institute for Agronomic Research (INRA) in Godet (16°20’N, 61°30’W, 30 m a.s.l.), Guadeloupe. The tree/grass association was established in May 1989 by planting G. sepium cuttings at 0.3 (2 m spacing in a natural Dichantium aristatum (Poir) C.E. Hubbard grassland. The climate is subhumid tropical, with mild hydrologic deficit from January through May. The site is described in detail by Tournebize and Sinoquet (1995) and Nygren et al. (2000). Most of the data for model parameterisation was collected from April 1996 to March 1997. The pruning regime was partial (ca. 50%) foliage removal every two months (P-60) and grass was harvested every 4 to 6 weeks.
The model was first run for the P-60 regime that is practised in the field site. The initial biomass values, and N availability and average irradiance parameters reported by Berninger et al. (2000) were adjusted to correspond the field site using a Monte Carlo type iteration. When the simulation results by the end of the 10-year-period corresponded to the observed values in 1997, three other pruning regimes were simulated with the same parameter values. The regimes simulated were partial pruning every 45, 90 and 120 days (P-45, P-90 and P-120, respectively). The model simulates single tree growth. The hectare-based values were obtained by multiplying the single tree values by 13,000, which is the actual tree density in the reference field site.
The simulation results for the P-60 pruning regime indicated decreasing foliage biomass production at the beginning of the 10-year-period (Fig. 1A). The production was stabilised in about four years, and thereafter a sustainable production was observed. Fine root biomass decreased after the first simulation year. Root and shoot reserve C concentration was strongly affected by prunings at the beginning of the simulation period, but only a slight effect was observed after five years, mainly in the shoot starch concentration (Fig. 1B). The stabilisation of starch concentration is probably caused by the growth of reserve organs - stem and coarse roots -
Table 1. Fodder production and N2 fixation under four partial pruning intensities of G. sepium according to simulations over ten years.
|
Pruning regime |
Foliage biomass at last
harvest |
Cumulative fodder yield |
Nodule biomass at last
harvest |
Average N2 fixation % |
|
P-45 |
N.A.* |
16.7 |
N.A.* |
57% |
|
P-60 |
2,036 |
65.8 |
11.0 |
60% |
|
P-90 |
5,017 |
80.4 |
21.0 |
44% |
|
P-120 |
5,915 |
83.1 |
21.9 |
40% |
* The trees died in 4.5 years under the P-45 pruning regime and consequent increase in total C reserves. García et al. (2001) have shown that high total starch reserves are more important than a high concentration for regrowth rate of G. sepium.
The simulated foliage biomass before pruning at the end of the ten-year-period was 2,036 kg ha-1 under the P-60 regime (Table 1). This is close to the biomass of 1,800 - 1,900 kg ha-1 observed in the Godet field site by Nygren et al. (2000). Cruz (2001) measured foliage biomass range of 3,900 - 5,200 kg ha-1 in Godet during the fourth year after establishment of the experiment. These values correspond to the initial high productivity simulated by the model. Thus, the simulated decreasing pattern in foliage biomass production is supported by field data, but the observed decrease occurred later than in our simulations (Fig. 1A). Both the simulated nodule biomass at the end of the ten-year simulation period, 11.0 kg ha-1 (Table 1), and the average N2 fixation percentage during the last simulation year, 80% (Fig. 2) were close to the values observed in the field, 8.6 kg ha-1 and 76%, respectively (Nygren et al., 2000).
The highest simulated harvest intensity, P-45, caused turnover of G. sepium in 4.5 yrs, while the P-45 regime has been highly productive in short-term studies (Barnes, 1998), but it seems unsustainable in the medium-term. The cumulative fodder harvest during the ten-year-period was almost the same under P-90 and P-120 regimes (Table 1). Although the latter regime allowed better foliage production between the prunings, the higher litterfall reduced the forage yield. The proportion of N2 fixation out of total foliage N yield was higher under the P-60 regime, while the total nodule mass was higher under the lower pruning intensities that produced more N consuming foliage. There was an increasing trend in the N2 fixation percentage over time under all harvest intensities (Fig. 2). Although more C is needed in the N2 fixation process than in N uptake, the higher C consumption in fine root growth may turn the N benefit to C consumption ratio in favour of N2 fixation in big trees that need extensive root systems for N uptake. Under the serious C restrictions caused by the higher pruning intensities, the regrowth of fine roots may require so much more C compared to renodulation that the total C balance favours N2 fixation.
Figure 2. Simulated yearly average of N2 fixation
percentage in Gliricidia sepium under four partial pruning regimes during
a ten-year-period.
Under various C availability options, generated by fodder harvesting, the model seemed to converge to a functional balance. The simulation results for P-60 regime agreed with field data. Simulations over ten years indicated an optimal combination of fodder yield and sustainability under the P-90 regime. The decrease in foliage production during the initial four years under all pruning regimes suggests caution for interpreting short-term forage production data. Somewhat surprisingly, C limitation by pruning appeared to favour N2 fixation over mineral N uptake.
Barnes P (1998) Fodder production of some shrubs and trees under two harvest intervals in subhumid southern Ghana. Agrofor Syst 42: 139-147
Berninger F, Nikinmaa E, Sievänen R and Nygren P (2000) Modelling of reserve carbohydrate dynamics, regrowth and nodulation in a N2-fixing tree managed by periodic prunings. Plant Cell Environ 23: 1025-1040.
Catchpoole DW and Blair GJ (1990) Forage tree legumes I. Productivity and N economy of leucaena, gliricidia, calliandra and sesbania and tree/green panic mixtures. Aust J Agric Res 41: 521-530
Cruz P 2001. Annual dry matter and nutrient yields in a Dichanthim sward with or without Gliricidia shrubs. Proc XIX Int Grassl Congress, 12-23 Feb, São Pedro, Brazil, in print.
Erdmann TK, Nair PKR and Kang BT (1993) Effects of cutting frequency and cutting height on reserve carbohydrates in Gliricidia sepium (Jacq.). Walp. For. Ecol. Manag. 57: 45-60.
García H, Nygren P and Desfontaines L (2001) Dynamics of non-structural carbohydrates and biomass yield in a fodder legume tree under different harvest intensities. Tree Physiol 21, in print.
Mochiutti S (1995) Comportamiento agronómico y calidad nutritiva de Gliricidia sepium (Jacq.) Walp. bajo defoliación manual y pastoreo en el trópico húmedo. M.Sc. thesis, CATIE, Turrialba, Costa Rica, 145 pp
Mäkelä A (1986) Implications of the pipe model theory on dry matter partitioning and height growth in trees. J Theor Biol 123: 103-120
Nygren P and Cruz P (1998) Biomass allocation and nodulation of Gliricidia sepium under two cut-and-carry forage production regimes. Agrofor Syst 41: 277-292.
Nygren P, Cruz P, Domenach AM, Vaillant V and Sierra J (2000) Influence of forage harvesting regimes on dynamics of biological dinitrogen fixation of a tropical woody legume. Tree Physiol 20: 41-48
Nygren P and Ramírez C (1995) Production and turnover of N2 fixing nodules in relation to foliage development in periodically pruned Erythrina poeppigiana (Leguminosae) trees. For Ecol Manage 73: 59-73
Shinozaki K, Yoda K, Hozumi K and Kira T (1964) A quantitative analysis of plant form - the pipe model theory. I: Basic analyses. Jpn J Ecol 14: 97-105
Simons AJ and Stewart JL (1994) Gliricidia sepium - a multipurpose forage tree legume. In: Gutteridge RC and Shelton HM (eds) Forage tree legumes in tropical agriculture, pp 30 - 48. Wallingford, UK, CAB International
Thornley JHM (1972) A balanced quantitative model for root:shoot ratios in vegetative plants. Ann Bot 36: 431-441.
Thornley JMH (1991) A transport resistance model of forest growth and partitioning. Ann Bot 68: 211-226.
Tschaplinski TJ and Blake TJ (1994) Carbohydrate mobilization following shoot defoliation and decapitation in hybrid poplar. Tree Physiol 14: 141-151.
A.T. Omokanye[142], R.O. Balogun[143]
Key words: browse plants, early growth, Nigeria, phosphorus
Browse plants are important sources of feed for livestock especially during the dry season of drier tropical areas. These plants remain green longer into the dry season and fluctuate less in quality than herbaceous legumes and grasses. Studies have been conducted to evaluate the forage potential and uses of leguminous shrub and tree species in northern Nigeria (Asare et al., 1984; Omokanye et al., 2001). However, one of the major factors limiting the usefulness of some of these forage species is their slow growth rate during establishment. Slow establishment growth may be related to deficiencies of essential nutrients in the soil (Cobbina et al., 1989).
The Rhizobium requirements for effective N2 fixation of tropical browse species have been examined by various authors. Studies have further shown that a low level of phosphorus is among the main chemical constraints to successful establishment of legume trees on tropical soils (Sanginga, 1985) and that nodulation and nitrogen fixation in tree legumes is closely associated with the availability of P (Andrew, 1982; Sanginga, 1985). The objective of this study was to determine the seedling growth rates of introduced and native browse species and their ability to nodulate effectively with or without P fertilisation on an upland soil of the subhumid ecological zone of Nigeria.
A potted trial was conducted between February and May 1995 at the National Animal Production Research Institute at Shika (11o15’N, 7 o 32’E; altitude 610m) in northern Nigeria. The Shika soil is a well-drained sandy loam soil with a clay fraction consisting mainly of kaolinite and small quantities of illite, and is deficient in N and P (Kowal, 1968). Shika has a subhumid tropical climate with a mean annual precipitation of 800-1300mm (average 1100mm) most of which falls from June - September.
Soil was obtained from an upland site dominated by Digitaria smutsii. The soil was ground and passed through a 10mm sieve to remove stones and plant parts, and then transferred into plastic pots of 20cm in height and 18cm top diameter. The pots were watered to full capacity and left for a day before planting. The pots were arranged in a split-plot design with 3 replications. Eight leguminous browse species consisting of 4 introduced and 4 native species formed the main treatments (Table 1) and 6 phosphorus (P) levels [0 (control), 20, 40, 60, 80 and 100 kg/ha] constituted the sub-treatments.
Basal fertiliser was applied to all pots at the rates of 10 kg/ha of N and 10 kg/ha of K in the form of urea and muriate of potash, respectively. The P treatments and basal fertiliser were applied at planting. Micronutrients were applied via Agroliser® (with no NPK), a commercial product. Five seeds were planted per pot, then thinned to 2 plants per pot 14 days after germination. Prior to planting, seeds of Leucaena, Sesbania, Adenodolichos and Desmodium sp. were scarified with concentrated sulphuric acid for 22, 4, 7 and 5 minutes, respectively. The seeds of G. sepium and C. argentea do not require scarification and as such were not scarified. With the exception of A. paniculatus and D. velutinum, the species were inoculated with appropriate Rhizobia at planting. Plants were harvested at 12 weeks post planting, (WPP), soil was gently washed from the roots of the plants and nodules were detached and counted. Nodulation index (NI), which accounts for the effect of plant size and nodule mass, was calculated for each plant. The following parameters were also recorded: plant height, nodule number, shoot DM, root DM and nodule DM. Data were statistically analysed using SAS procedures (SAS, 1988). Only the means pooled across either the main or sub-treatments are presented.
Plant height
For the various browse species, plant height varied significantly (P<0.05) from 19cm for D velutinum to 46cm for native G. sepium (Table 1). The influence of P application on plant height was pronounced, with plant height increasing from 21cm for P0 to 44cm for P80 (Table 2). Phosphorus application above 80 kg/ha caused a dramatic decrease in height. No significant species x P interactions for plant height occurred. Both introduced and native G. sepium and A. paniculatus grew faster with increasing levels of P.
Table 1. Mean seedling growth parameters of eight browse species, 84 days planting in subhumid Nigeria.
|
|
|
Plant |
Nodule |
Nodule |
Shoot |
Root |
Root:Shoot |
Nodulation |
|
|
|
height |
number |
DM |
DM |
DM |
(by weight) |
index |
|
Browse species |
Acc1 |
cm |
no. |
---g/plant---- |
|
& |
||
|
Leuceana pseudotrichoides |
ILCA14192 |
27 |
28 |
0.11 |
2.2 |
1.7 |
0.79 |
5.1 |
|
Leuceana leucocephala2 |
Shika3 |
21 |
13 |
0.10 |
1.8 |
1.2 |
0.66 |
5.6 |
|
Sesbania sesban |
ILCA1193 |
38 |
26 |
0.18 |
2.9 |
2.1 |
0.71 |
6.2 |
|
Cratylia argentea |
n.a. |
32 |
37 |
0.28 |
3.1 |
2.5 |
0.81 |
9.2 |
|
Gliricidia sepium |
ILCA 14984 |
42 |
46 |
0.33 |
3.5 |
3.1 |
0.88 |
9.2 |
|
Gliricidia sepium |
Shika |
46 |
51 |
0.37 |
3.4 |
2.8 |
0.85 |
11.0 |
|
Adenodolichos paniculatus |
Shika |
30 |
48 |
0.39 |
3.1 |
2.6 |
0.86 |
12.7 |
|
Desmodium velutinum |
Shika |
19 |
8 |
0.06 |
1.3 |
1.5 |
1.17 |
5.0 |
|
LSD (P=0.05) |
|
5 |
11 |
0.05 |
0.21 |
0.28 |
0.45 |
2.74 |
1 Accession, 2 Naturalised in many regions
Dry weight and ratio
Root and shoot DM were higher with G. sepium ILCA 14984, while A. paniculatus had better nodule DM than did other species. Shoot, root and nodule DM of the browse species respectively varied (P<0.05) from 1.3 to 3.5, 1.2 to 3.1 and 0.06 to 0.39 g/plant (Table 1). Phosphorus application significantly (P<0.05) improved seedling DM production. The DM production increased with increase in P application in this order: P 80>P60>P40>P20>P0 (Table 2). The highest values recorded were 0.324, 3.46 and 2.76g/plant respectively for nodule, shoot and root DM (Table 2).
At 12WPP, the mean root: shoot ratio was not significantly affected (P>0.05), and this was highest (0.913) for D. velutinum and lowest (0.79) for L. pseudotrichoides ILCA 14192. Surprisingly, root: shoot ratios for P rates over all species were not significantly (P>0.05) affected (Table 2).
Table 2. Mean seedling growth parameters of eight browse species, subjected to six phosphorus rates in subhumid Nigeria.
|
|
Plant |
Nodule |
Nodule |
Shoot |
Root |
Root:Shoot |
Nodulation |
|
|
height |
number |
DM |
DM |
DM |
(by weight) |
index |
|
Phosphorus rates (KgP2O5/ha) |
cm |
no. |
---g/plant---- |
|
% |
||
|
0 |
21 |
7 |
0.12 |
1.4 |
1.2 |
0.83 |
8.2 |
|
20 |
27 |
13 |
0.13 |
1.9 |
1.6 |
0.83 |
6.9 |
|
40 |
32 |
24 |
0.21 |
2.7 |
2.2 |
0.83 |
7.8 |
|
60 |
38 |
40 |
0.28 |
3.2 |
2.6 |
0.80 |
8.9 |
|
80 |
44 |
57 |
0.32 |
3.5 |
2.7 |
0.78 |
9.4 |
|
100 |
30 |
53 |
0.30 |
3.3 |
2.7 |
0.83 |
9.0 |
|
LSD (P=0.05) |
7 |
9 |
0.05 |
0.15 |
0.13 |
0.02 |
1.03 |
Nodulation - nodule weight and number
The two G. sepium, and A. paniculatus produced significantly higher (P<0.05) nodule number and nodule DM (>40 nodules/plant) at 12WPP than did most other species. Number of nodules was highest for native G. sepium (51 nodules/plant) and lowest for D. velutinum (13 nodules plant-1) (Table 1). With respect to P application, nodule count was least (7 nodules/plant) for P0 and this increased by about 77, 85, 89 and 88% with the application of P40, P60, P80 and P100, respectively. In general, the two species of Gliricidia and Leucaena, and C. argentea produced numerous, but smaller nodules. Nodulation index of both G. sepium accessions, C. argentea and A. paniculatus were high (>9.0%) compared to the other 4 species.
Correlation of nodulation was high with root length and number of nodules (r=0.693 and r= 0.620), moderately with shoot DM (r=0.579) and fair with nodule DM (r=0.405). On the average, the two nodulation parameters (number and weight of nodules) correlated significantly (r=0.527).
The two G. sepium accessions established rapidly, indicating the adaptation of this species to acid-infertile soils. In contrast, the two Leucaena species were slow to establish confirming earlier reports by Wandera et al. (1991) and Veasey et al. (1997). Although C. argentea showed slow early growth, observations have shown that, once established, C. argentea has good attributes such as vigorous growth after defoliation, profuse flowering and drought tolerance (Omokanye et al., 1998), with a dense canopy and impressive green foliage far into the dry season. The native A. paniculatus and D. velutinum in their natural habitat were not as fast as the other browse species in initial plant growth. Earlier observations show the above-ground part (shoot) dies back gradually immediately after seeding (December and late January-early February), thus giving way for a very much faster and impressive regrowth (especially with A. paniculatus) at the beginning of the following raining season.
Root: shoot ratios, recorded 84 day post sowing showed that only D. velutinum allocated greater energy to below-ground biomass in the early growth period. However, some such as G. sepium, A. paniculatus, S. sesban also have high preference for below-ground energy supply. Leucaena leucocephala allocates slightly more energy to the above- ground biomass. These observations could be attributed to the level of nodulation.
The results show that nodulation occurred freely for the control (P0) of the two G. sepium, Leucaena leucocephala and A. paniculatus. The free nodulation was expected for G. sepium (Kang and Mulongoy, 1987). Adenodolichos paniculatus is native to the subhumid environment of Nigeria and this probably explains why nodulation was better for it even without inoculation.
Nodulation (nodule number and nodule DM) was greatly inhibited (except for L. The application of 80kg/ha P significantly favoured the responses of all parameters (excluding root DM) and this could probably be attributed to better nodulation. This may in turn, explain why plant height grew faster at 80kg/ha P. All these were due to increase the availability of P in the soil, a function of the P added to the soil. The drop off in growth at 100 kg/ha P is difficult to explain.
Andrew, C.S. (1982) Influence of nutrition on nitrogen fixation and growth of legumes. In: A Review of Nitrogen in the Tropics, with Particular Reference to Pasture. Commonwealth Bureau of Pastures and Field Crops, Farnham, UK. Bulletin, 46, 130-146.
Asare, E.O., Shehu, Y. and Agishi, E.C. (1984) Preliminary studies on indigenous species for dry season grazing in the Northern Guinea Savanna Zone of Nigeria. Tropical Grasslands, 18, 148-152.
Cobbina, J., Kang, B.T. and Atta-Krah, A.N. (1989) Effect of soil fertility on early growth of Leucaena and Gliricidia in alley farms. Agroforestry Systems, 8, 157-164.
Kang, B.T. and Mulongoy, K. (1987) Gliricidia sepium as a source of green manure in an alley cropping system. In: Withington, D., Glover, N. and Brewbaker, J.L. (eds) Gliricidia sepium (Jacq.) Walp.: Management and Improvement. Special NFTA Publication (87-01). (NFTA: Hawaii).
Kowal, J. (1968) Some physical properties of soils at Samaru, Zaria, Nigeria: Storage of water and its use by Crops. I. Physical status of soil. Nigerian Agricultural Journal, 5, 13-20.
Omokanye, A.T., Dung, D.D., Balogun, R.O., Awemu, E., Akinpelumi, O.P., Onifade,O.S. and Kallah, M.S. (1998) Degradability and nutritive indices of Brachiaria decumbens hay mixed with Cratylia argentia. Proceedings of the Silver Anniversary Conference of the Nigerian Society for Animal Production and West African Society for Animal Production Inaugural Conference, Abeokuta, Nigeria,1998. pp. 1-2.
Omokanye, A.T., Onifade, O.S., Amodu, J.T. and Kallah, M.S. (2001) The performance of forage germplasm in a screening trial at Shika, Nigeria. Tropical Grasslands (in press).
Sanginga, N. (1985) Nodulation of Leucaena leucocephala (Lam) De Wit and its contribution to nitrogen status of soils. Ph.D. Thesis. Facilte des Sciences Agronomiques, Yangambi, Zaire.
SAS (STATISTICAL ANALYSIS SYSTEMS) (1988) SAS/STAT User’s Guide, Release 6.03. (SAS Institute Inc., Cary, North Carolina, USA).
Veasey, E.A., GhisI, M.A.A.,Valarini, M.J., Otsuk, I.P., Cardelli, M.A., Sanchez, M.J.F. and Beisman, D.A. (1997) Early growth and native nodulation of legumes shrub and tree species in Brazil. Tropical Grasslands, 31, 40-48.
WanderA, F.P., Dzowela, B.H. and KarachI, M.K. (1991) Production and nutritive value of browse species in semi-arid Kenya. Tropical Grasslands, 25, 349-355.
Gertrudis Pentón Fernández[144]
Key words: botanical composition, chemical composition, grass, trees,
The projection of shade by Albizia lebbeck’s tree is a particular case in studies about the effects of the reduction of light in the associated grass. A. lebbeck is a leguminous fast growing tree which shed its leaves in the dry season (November- January) and this makes a dynamic of the light regimen all year round for the pasture management. For this reason, the propose of this investigation is to know the level of shade projected by this species in semi-natural condition, and its effects on the botanical and chemical composition of the pastures in silvopastoril condition.
This investigation was carried out at Indio Hatuey Experimental Station of Pastures and Forages, Matanzas, Cuba. In a Red Moisturized Soil Ferralitic with an average value of 6,3% of organic material, pH slightly acid and low content of phosphorous. The experimental area was used with 12 young bovine, 5/8 Holtein x 3/8 Cebú; with 8-10 month average age of exploitation and 110 Kg average liveweight. The study area consisted 1.72 ha of silvopastoril system which was characterized by natural population of A. lebbeck (algarrobo de olor). The plant density was 1500 trees and grass yield was between 9600 and 10200 Kg dry matter (DM)/ha/yr. The experimental design was in blocks at random with 6 repetitions. The treatments were the shade area (with trees) and light area (without trees). The variables studied were: projected shade of the trees, botanical and chemical composition of forages (% DM, % protein, PB; % fiber, FB; % ash, Ce; % calcium, Ca and % phosphorus, P.
In table 1, we can observe that the percentage of Panicum maximum under shade was greater than that of open sunlight for both years. This in an indicates that P. maximum have a greater ability to grow in shaded tree environments of A. lebbeck than other grasses. Dicanthium contrarily, expresses a great increase of their population on the treatment with shade. About this, we could observe that this combined action of the shade of the trees and the presence of the P. maximum produced the resession of Dicanthium with R= 0.95 (equation 1). Which can be explained if we consider that D. annulatum and D. caricosum species have among their ecology requirement the pickup of high intensities of light. In this condition the D. annulatum and D. caricosum are highly competitive. On the other hand, the plant groups with big leaves have a superior behavior on a treatment with trees in 1994. However, in 1997 decreased in all system and don’t show evidences of affectation caused for the shade; which permit to infer that there were other factor inciting over population. The herbage leguminous developed better under light treatment. About this behavior Hernández (1984) said that the level of the competition of herbage leguminous in the shade’s condition is determinate for their capacity to obtain preferential access to the radiation by means of their high. Which was very limited for the high level of the competition existing in this system. Moreover, it was demonstrated that the herbage leguminous inhibits their square-root sing development in a soil with high nitrogen content, which affects the capacity of expansion (Cuesta and López, 1984).
Table 2 presents a variation at sun and shade of the chemistry composition of herbage plants. In it we can observe that in the rain period as well as in the dry period contain of FB was bigger a treatment of light. Which confirms the results historically reported on the world literature, referred to the stimulation that sun full light and high temperature cause in the structure carbohydrates. In respect to contain of MS their significant reduction under the shade of a trees during the rain period stage, when A. lebbeck projected 51% of a vertical shade calls the attention. This phenomenon coincides with Páez, González and Pereira statement (1994), referred to the fact that the increase of a shade into the pastures provoqued a reduce of dry material content, as result of a decrease of the concentration of the increase in the contain of humidity in the grass (Capote and Shishchenko, 1974). However, the significance increase of the dry material concentration under the shade and during a dry period as result of a shade reduction of the trees to 36.87%, indicates that a projection of moderated levels of shade and diffused sun radiation brings about decrease of the light saturation and the increase of the efficiency on the effects of the decrease of light referred by Alberta (1965) and Capote and Shishchenko (1974). A chemistry component that didn’t show difference between a treatment was phosphorous, which has been reported by some authors as an not so much variable element under the light action (Guevara, Ruiz, Curbelo, Canino y Jimenez, 199).
The significant increase of calcium contain in spring under a shade treatment, confirms the results of Belsky (1992, 1994), that finded great increase in Ca under the Acacia tortilis and Adansonia digitata. Contain of gross protein and total ash in a grass under the shade was higher widely than value of this under full sun. This confirms that under canopy of trees, a shade, humidity and contain of the organic matter, among other aspects, favor the process of mineralization and supply of nutrient in form of ions of Ca, K, Mg, P and nitrates. Which favor an increase of the protein in the plants for concept of increase in mineral content in the soil and extraction of nutrient for a root (Anon, 1990). In this research it was observed a high relation of dependence of grass protein contain in respect to percent of ash, the presence of P. maximun, the density of trees and the projected shade (equation 2).
From all of this it was concluded that A. lebbeck in its condition of caducifolia plant caused a great season variation of shade. Among the pasture species present in the system P. maximun showed high associative potential with trees and was a controlator factor of the plants with a low nutritional value. Under studies conditions, the Dichantium complex moves back notably in the presence of the increase of shade level and the competition of P. maximun. Under the shade of trees, it occurred a significant increase in the protein and ash content; which depended in great measurement on botanical composition, that was favored by shade.
Alberda, T. 1965. The influence of temperature light intensity and nitrate concentration on dry matter production and chemical composition of Lolium perenne L. Neth. J. Agric. Sci. 13:335
Alvarez, F. 1994. La tierra viva: Manual de agricultura ecológica. Universidad Metropolitana, Madrid, España
Anon. 1990. Programa AM: Pasture management and livestock production. CSIRO. Division of Tropical Crops and Pastures. Annual Report. p. 10
Belsky, A. J. 1994. Influences of tress on savanna productivity: Tests of shade, nutrients, and tree-grass competition. Ecology, 75 (4) Pp. 922-932
Belsky, A.J. 1992. Effects of trees on nutritional quality of understorey grasmineous forage in tropical savannas. Tropical Grassland 26:12-20
Capote, S. & Shishchenko, S.V. 1974. Investigation of physiological processes in alfalfa, clover and Rhodes grass in Cuba, with respect to conditions of lighting and root feeding. In: Biological and physiological aspects of the intensification of grassland utilization. Proc. XII Int. Grassld. Congr., Moscow. p. 62
Cuesta, A. & Lopez, M. 1984. Resúmenes II Jornada Científico-Técnica Universitaria de la Isla de la Juventud. p. 91
Guevara, A.; Ruiz, R.; Curbelo, L.; Jimenez, A. & Canino, E. 1994. Efecto de la sombra del algarrobo común (Samanea saman) sobre el comportamiento productivo del pastizal. Resúmenes. Taller Internacional “Sistemas Silvopastoriles en la Producción Ganadera”. EEPF “Indio Hatuey”. Matanzas, Cuba. p. 54
Hernández, R. 1984. Evaluación agronómica de gramíneas en regiones ganaderas bajo diferentes ambientes. Tesis presentada en opción al grado de Candidato a Doctor en Ciencias. Instituto Superior de Ciencias Agropecuarias de la Habana, Cuba. p. 37
Sinclair, J.; Shiraiwa, T & Hammer, S. 1992. Variation in crop radiation use efficiency with increased diffuse radiation. Crop Science 32: 1281-1284
Table 1. Variation of botanical composition due to shade effect.
| Treatments/botanical components (%) |
1994 |
1997 |
||||
| Shade |
Full light |
S. E. |
Shade |
Full light |
S. E. |
|
| P. maximum |
34.81(a) |
10.18(b) |
3.555 |
63.54(a) |
32.92(b) |
1.661 |
| B. decumbens |
0 |
1.45 |
0.572 |
1.34(b) |
9.51(a) |
1.370 |
| Complex Dichanthium |
9.57(b) |
39.86(a) |
0.978 |
1.52(b) |
24.37(a) |
0.503 |
| P. notatum |
2.32 |
5.87 |
1.240 |
1.28 |
2.89 |
0.683 |
| Other grasses |
22.28 |
26.62 |
1.030 |
0.79 |
2.67 |
0.616 |
| Broadleaves |
25.04(a) |
8.47(b) |
1.970 |
2.38 |
2.03 |
0.285 |
| prostrate legumes |
0.76 |
2.81 |
0.571 |
1.50(b) |
4.18(a) |
0.506 |
Table 2. Variation of the chemistry composition due to shade effect.
|
Treatments/Indicators |
FB (%) |
MS (%) |
PB (%) |
Ca (%) |
P (%) |
Ce (%) |
|
Dry period |
||||||
|
Shade |
31.14(b) |
28.88(a) |
12.61(a) |
0.94 |
0.14 |
10.11(a) |
|
Full light |
34.94(a) |
26.20(b) |
9.52(b) |
0.86 |
0.12 |
9.50(b) |
|
S. E |
0.690 |
0.330 |
0.640 |
0.027 |
0.007 |
0.146 |
|
Rain period |
||||||
|
Shade |
30.05(b) |
18.79(b) |
15.16(a) |
0.98(a) |
0.15 |
10.89(a) |
|
Full light |
33.58(a) |
19.95(a) |
8.52(b) |
0.76(b) |
0.14 |
9.28(b) |
|
S. E |
0.500 |
0.140 |
0.600 |
0.026 |
0.008 |
0.189 |
(a, b): Commune superindites to values: P? 0.05
|
Equations of Regression Lineal |
|
|
I. Y= 32.7-0.31X1-0.27X2 |
|
|
X1= % of shade |
Est. Residual Desv. = 4.53 |
|
X2=% of P. máximun |
R **2= 0.90 |
|
|
Multiple R = 0.95 |
|
II. Y=-3.748+1.319X1+0.048X2+0.017X3+0.001X4 |
|
|
X1= % of ash |
Est. Residual Desv.= 0.879 |
|
X2= % of P. maximun |
R**2= 0.953 |
|
X3= % of shade |
Multiple R = 0.976 |
|
X4= Trees density |
|
Perla, H., Kass, D., Ibrahim, M., Jiménez, F
Species and varieties within species have been shown to vary with respect to their ability to access different pools of phosphorus in soils. Since total P in soils is often adequate for plant growth but available pools vary widely, it is essential to utilize germplasm which through their own peculiarities (exudation of solubilizing elements and high P efficiency) or their ability to form associations with fungi (mycorrhizae) are capable of accessing more pools of soil P. Fallow species with high P contents can make more P available to succeeding crops (Buresh, 1999).
An experiment was set up in randomized complete blocks on a soil classified as a Typic Dystrudept, at 10º 09’ N, 83º 38’ W Germania association, in Pococi, Costa Rica. The site had an elevation of 125 m above sea level, an annual precipitation of 4360 mm, an average temperature of 24.6 C and a relative humidity of 87%. Global solar radiation varies from 13 to 17 MJ m-2 day-1 being highest in the driest months (February, March, April, May and September). Acacia mangium tress were planted at the site in September, 1997 at a spacing of 3m X 9m. In January of 2000, the area was weeded manually and paraquat applied and propagules of Arachis pintoi planted at a spacing of 0.5 X 0.5
m. Each block consisted of six lines of trees, 33m long, oriented at 40º East-South. Within each 9m X 3m plot, the following treatments were established:
T0: A. mangium without A. pintoi
T1: A. mangium + A. pintoi CIAT 22155
T2: A. mangium + A. pintoi CIAT 22157
T3: A.mangium + A. pintoi CIAT 22159
T4. A. mangium + A. pintoi CIAT 22148
T5: A. mangium + A. pintoi CIAT 22150
T6: A. mangium + A. pintoi CIAT 17434
T7: A. mangium + A. pintoi CIAT 18744
Accessions were selected according to indications in the literature and from a preliminary count of mycorhizal spores. Since, for certain variables, plots were sampled at two dates and at three distances from the trees (1.5 m, 3.0 m, and 4.5 m), the resulting design was a split split plot experiment with repeated observations in time and sampling distances considered a variable applied in strips since it could not be randomized in each plot. The following variables were evaluated: total soil P, inorganic P extracted by a resin, inorganic P extracted by NaHCO3, organic P extracted by NaHCO3, presence of mycorrhizal spores, biomass production.
At bimonthly intervals (3 determinations during the experiment), foliar nutrient levels at the end of the experiment, and specific root length of A. pintoi at three distances from the trees and three soil depths. Data were taken from January to July, 2000.
Total phosphorus and P fractions were not signficantly different among treatments but fractions of inorganic P extracted with resins (membrane Pi) as well as organic phosphorus extracted with sodium carbonate (NaHCO3 Po) were lower in concentration at the end of the study (six months after planting the A. pintoi) in all treatments with the exception of the line CIAT 22155. Significant differences (p< 0.05) were found with respect to the levels of foliar P, Mg and Zn with accession CIAT 18744 showing the highest levels of foliar Mg (8866.67 mg kg-1) and P (2400 mg kg-1) and accession CIAT 22150 showing the highest levels of P (2400 mg kg-1) and Zn (53.58 mg kg-1).
The results of the spore count and colonization of roots by VAM did not show significantly differences (p <0.05) but all accessions showed relatively high colonization rates (80%). These data are summarized in Table 1. Significant differences in dry matter production were observed. Dry matter varied significantly (p<0.05) with distance from the tree row (1.5, 3.0 and 4.5
m). Dry matter increased from 1.5 m to 3.0 m from the tree row. Accessions CIAT 22157 and CIAT 18744 showed a linear response to distance from tree over all three distances while CIAT 17434 showed a quadratic response. There was also a significant response to sampling dates with the highest dry matter production being obtained in the sixth month when CIAT 18744 presented the highest dry matter yield (2010 kg ha-1).
Specific root length (SRL) showed significant difference with CIAT 174324 presenting the highest value (741.3 mm mg-1). Distance from the trees did not significantly affect specific root length. It was concluded that proper selection of Arachis pintoi lines could significantly increase P cycling in this silvopastoral system, well adapted to infertile soils of the tropics.
Different accessions of Arachis pintoi show sufficient variability in dry matter production and ability to accumulate P and other nutrients in soils to justify the selection of strains with a greater ability to accumulate P and dry matter in agroforestry and silvopastoral systems. The best strains used in the present studied compared well to species such as Tithonia diversifolia (Helmsley) A. Gray, often recommended for their ability to accumulate P (Palm et al., 1999), having the threshhold level of 2400 mg kg-1 There are also signficant differences in accessions with respect to their ability to grow in association with Acacia mangium.
Buresh, R., 1999. Phosphorus management in tropical agroforestry: current knowledge and research challenges. Agroforestry Forum 9 (4): 61-66.
Palm, C., Nziguheba, G., Gachengo, C., Gacheru, E. and Rao, M.R. 1999. Organic materials as sources of phosphorus. Agroforestry Forum 9(4): 30-33
Table 1. Changes in levels of soil P fractions (kg ha-1), accumulation of P in above ground biomass (kg ha-1), foliar P concentration (mg kg - 1), and fraction of roots colonized by vesicular arboreal mycorrhizae (MVA) of different accessions of Arachis pintoi grown in association with Acacia mangium for six months in a Typic Dystrudept of Costa Rica (from Perla, 2000).
|
Treatment |
Change in membrane Pi |
Change in bicarbonate extracted Pi |
Change in bicarbonate extracted Po |
P accumulated in above ground tissue in six months |
Foliar P (mg kg-1) |
Fraction of roots colonized by mycorrhizae |
|
A.mangium alone |
-0.01 |
+5.28 a |
-4.17 |
|
|
|
|
A. mangium + A. pintoi CIAT 22155 |
0.00 |
+2.31 ab |
-6.21 |
4.30 |
2167ab |
0.87 |
|
A.mangium + A. pintoi CIAT 22157 |
-0.32 |
-3.02 b |
-6.98 |
4.24 |
2100ab |
0.84 |
|
A. mangium + A. pintoi CIAT 22159 |
-0.51 |
+2.18 ab |
-1.83 |
4.48 |
2133ab |
0.80 |
|
A. mangium + A. pintoi CIAT 22148 |
-0.26 |
+3.20 ab |
-6.84 |
4.39 |
2033b |
0.91 |
|
A. mangium + A. pintoi CIAT 22150 |
-0.25 |
+1.77 ab |
-2.64 |
3.43 |
2400a |
0.96 |
|
A. mangium + A. pintoi CIAT 17434 |
-0.06 |
+3.46 ab |
-2.73 |
4.15 |
2200ab |
0.87 |
|
A. mangium + A. pintoi CIAT 18744 |
-0.38 |
-0.14 ab |
-10.41 |
5.97 |
2400a |
0.84 |
Values followed by the same letter do not differ significantl by the Tukey test at p < 0.05.
Plevich, J.O.[145], Demaestri, M. [146] Viale, S.N[147], Cantero J.J. [148], Nuñez C.O[149]
Key words: grassland, Pinus elliotti, thinning
The development of herbaceous species is affected when they grow under the canopy of woody species. Usually the productivity of these grasses increases when the competition with the woody species for light, water, and nutrients decreases. In plantations of Pinus elliottii present in the mountain range of Comechingones in the Argentinian province of Cordoba, scarce growth of natural grassland is observed, however, this grassland regenerates easily in open spaces of the forest. Valls (1993), for the same geomorphologic entity in pine plantations, found production superior to 1500 kg m.s./ha. Because of that, it was hypothesized that by reducing the density of the Pinus elliotti plantations, it would be possible to regenerate part of the floristic combinations of the natural pasture that existed at the time of its implantation and obtain levels of production compatible to the establishment of integrated systems for the production of forage and wood.
The study was carried out between October of 1996 and April of 1998 in a plantation 18 years old with a density of 1600 plants/ha and a homogeneous sector from a geologic, soil, and climatic point of view. The trials corresponded to four intensities of thinning: 25%, 37%, 50% and 62% of the trees and a control parcel. In order to evaluate the dry material accumulated in the grassland a design completely randomized was used. In each trial 16 repetitions distributed on a grill of 400M2 were realized. In each one of the 16 points of the grill a sample was taken of 1/10 M2. The plants collected in the samples were classified in grass and other families, later they were dried in an oven (100ª C) and weighed. The data of quantity of the dry material was statistically processed using a variance analysis and Duncan test.
In the herbaceous strata the dominant grass were Piptochaetium lasianthum, Piptochaetium stipoides, Stipa neesiana, Setaria sp., Bothriocloa sp., Briza subaristata and in the other families: Rubus ulmifolius, lepechinia floribunda, Hypochoeris chillensis, Viola odorata y Carduus acanthoides. The variance analysis detected significant differences in the production of dry material of pasture (p>0.05) when the trials reached intensities of thinning of 50% and 62% of the trees, with a production of 181.25 and 223.12 kg ms/ha produced by the grass and the independent variable (x) to the basimetric area of the plantation. The parameters obtained in the function were the following: y = 406.71 - 6.79* x, with a R2 = 0.82.
In accordance with the proposed working hypothesis it is possible to conclude the following: 1) When the density of the plantation of Pinus elliotti is reduced below the basimetric areas of 36m2/ha (thinning intensity of 50%), part of the floristic combinations of natural pasture which existed at the time of their implantation are regenerated 2)The low levels of physical production of natural grassland do not make it possible to affirm that integrated systems can be established for the production of forage and wood.
Gomez k and Gomez A, 1983. Statistical procedures for agricultural research. Seda edición. Editorial: Wiley J. and Sons. 679 p.
Grelen H, Witaker L and Lohrey R, 1972. Herbage response to precommercial thinning in direct seeded slash pine. Journal of range managment, 25 (6):435-437.
Montoya J M, 1982. Efectos del arbolado de las dehesas sobre el sistema pastoral. Criterios de ordenación, INIA. Anales serie forestal (5): 31-41
Olivares A, Cornejo R y Gandara J, 1983. Influencia de la estrata arbustiva (Acacia caven, Mol. Hook Et Arn.) en el crecimiento de la estrata herbacea. Avances en Producción Animal 8 (1-2): 19-28.
Pyke D and Zamora B, 1982. Relationships between overstory structure and understory production in the grand fir/myrtle boxwood habitat type Northcentral Idaho. Range manage 35: 769-773.
Valls P, 1993. Introducción al manejo integral de Recursos: pastoreo racional intensivo en un sistema forestal de sierra. Jornadas sobre pastoreo racional intensivo. Sociedad Rural de Río Cuarto, 3 al 4 de junio de 1993. Río Cuarto, Córdoba, República Argentina.
Jorge Ribaski[150], Jarbas Y. Shimizu[151], Mario T. Inoue[152]
Key words: Cenchrus ciliaris, chlorophyll, photosynthesis, Prosopis juliflora, soil fertility.
Livestock grazing has been a traditional activity in the Brazilian semi-arid region. However, production is low, mainly due to low quality or even total lack of pasture during the dry season. In order to overcome the nutrient deficiency, local farmers started to grow mesquite trees (Prosopis juliflora (SW) DC) mainly for pod production as a supplemental fodder.
The objective of this study was to study the influence of mesquite tree on several aspects of a silvopastoral system for the region involving buffel grass (Cenchrus ciliaris var. Biloela).
The experiment was established at Petrolina, PE, at latitude 09o09’S and longitude 40o22’W, at 365 m elevation above sea level. The study was based on observations made during the period of August 1997 to April 1998, in a silvopastoral system which is 15 years in operation. The experiment was set up in a randomized block design with ten replications of treatments consisting of two sampling points under the tree canopy and a third located 20 m away from each selected tree, representing the conditions of a monoculture pasture.
With a “LI-COR” portable photosynthesis measurement device, the following variables were measured: photosynthetically active radiation (PAR), net photosynthesis, photosynthetic efficiency, air temperature, relative humidity, and buffel grass leaf temperature. After harvesting the grass, the following data were collected: a) above ground dry matter yield; b) content of dry matter in the forage; c) content of chlorophyll a and b in the leaves; d) specific leaf area (SLA); e) nutritional value (crude protein, fiber and digestibility), and f) mineral composition of the forage (N, P, K, Ca and Mg). Soil data included contents of moisture, organic matter and nutrients (N, P, K, Ca, and Mg).
Mesquite trees had, on average, 7.4 m total height; 1.8 m commercial stem height; 5.6 m crown height; 10.4 m crown diameter, and 25.7 cm stem base diameter. PAR measured under the tree canopy was 50.4% during the dry season and 43.8% in the rainy season, in relation to full light conditions. On average, there was a reduction of 1.5oC in air temperature and 2.7oC in grass leaf temperature.
The presence mesquite trees in buffel grass pasture improved soil fertility by increasing the contents of organic matter, nitrogen and phosphorus and decreasing soil pH. Grass dry matter yield decreased under tree canopies. However, the forage produced under such condition (reduced light) had higher nutrient value. There was a significant increase in crude protein content. In spite of that, there was no statistically significant difference between full light and shaded conditions on the average content of neutral detergent fiber (NDF) or on “in vitro” dry matter digestibility (IVDMD), except during the rainy season.
The chemical composition of buffel grass forage was differently affected by the presence of mesquite tree. There was an increase in the concentration of some macronutrients (N and Mg) and decrease in others (P, K and Ca).
Under reduced light, buffel grass produced larger specific leaf area (SLA) and higher contents of chlorophyll, of both a and b types than under full light. Also, buffel grass grown in shaded environment compensated the low level of PAR with a higher photosynthetic efficiency.
Mesquite leaf abscission occurs year all round, but with higher intensity during the dry season. Sprouting of new leaves occurs mostly during the rainy season, from December to May (Lima, 1994). The shading by mesquite trees contributes to the conservation of soil moisture, thereby improving the environmental conditions for the pasture. Increases in SLA and in chlorophyll contents under shaded conditions were, also, reported in other species elsewhere (Friendship-Keller et al., 1987; Igboanugo, 1989; Valenzuela et al., 1991; Morita et al., 1994).
Mesquite tree is considered a potential species for the restoration of fertility and productivity of degraded soils. Garg (1998) and Bhojvaid and Timmer (1998) showed this species ability to reduce electrical conductivity and to increase both the organic carbon content and the nutrient availability in the soil. The effect of trees on the soil, in different silvopastoral systems is, usually, in the form of improved fertility due to increase in nitrogen content (Sanchez and Palm, 1996; Bhojvaid and Timmer, 1998; Buresh and Tian, 1998; Botero and Russo, 2000).
Lower dry matter yield but with higher nutrient value in buffel grass observed under this silvopastoral system was similar to other experiences involving different grass species, although, in some cases, dry matter yield has been greater in shaded environment (Botero and Russo, 2000; Hernández et al., 2000). According to Giraldo et al. (1995) and Daniel and Couto (2000), forage production, normally, decreases as tree canopy cover increases, with higher intensity with more than 50% cover.
Except for phosphorus, macronutrient contents in buffel grass under mesquite trees meet the nutrient requirements for cattle (National Research Council, 1984), even with the slight decrease in potassium and calcium contents as observed in this study.
The compensation with higher photosynthetic efficiency under low PAR observed in this experiment was similar to the case reported by Cruz (1997). In his study, Dichanthium aristatum had a higher rate of CO2 assimilation and a higher RUE (radiation use efficiency) under Leucaena leucocephala canopy than under full light.
Therefore, silvopastoral systems made up with buffel grass and mesquite tree are potentially viable for the semi-arid region in Brazil, given the importance of this leguminous tree as a source of timber as well as pods for supplemental fodder during the dry season.
Bhojvaid PP and Timmer VR (1998) Soil dynamics in age sequence of Prosopis juliflora planted for sodic soil restoration in India. Forest Ecology and Management, 106: 181-193
Botero R and Russo RO (2000) Utilización de árboles y arbustos fijadores de nitrógeno en sistemas sostenibles de producción animal en suelos ácidos tropicales. Memorias: Conferencia Electrónica: I Conferencia Latinoamericana sobre Agroforestería para la Producción Animal Tropical. URL: http://www. cipav.org.co/cipav/confr/index/htm
Buresh RJ and Tian G (1998) Soil improvement by in sub-Saharan Africa. Agroforestry Systems, 38: 51-76
Cruz P (1997) Effect of shade on the growth and mineral nutrition of a C4 perennial grass under field conditions. Plant and Soil, 188: 227-237
Daniel O and Couto L (2000) Una visión general de sistemas silvopastoriles y agrosilvopastoriles con eucalipto en Brasil. Memorias: Conferencia Electrónica: I Conferencia Latinoamericana sobre Agroforestería para la Producción Animal Tropical. URL: http://www. cipav.org.co/cipav/confr/index/htm
Friendship-Keller RA, Tsujita MJ, Ormrod DP (1987) Light acclimatization effects on japanese maple for interior use. HortScience, Alexandria, 22: 929-931
Garg VK (1998) Interaction of tree crops with a sodic soil environment: potential for rehabilitation of degraded environments. Land Degradation and Development, 9: 81-93
Giraldo LA, Botero J, Saldarrieaga J and David P (1995) Efecto de tres densidades de árboles en el potencial forrajero de un sistema silvopastoril natural, en la región atlántica de Colombia. Agroforestería en las Américas, 8: 14-19
Hernández I, Milera M, Simón L, Hernández D, Iglesias J, Lamela L, Toral O, Matías C and Francisco G (2000) Avances en las investigaciones en sistemas silvopastoriles en Cuba. Memorias: Conferencia Electrónica: I Conferencia Latinoamericana Sobre Agroforestería para la Producción Animal Tropical. URL: http://www. cipav.org.co/cipav/confr/index/htm
Igboanugo ABI (1989) Adaptation to changes in ilumination of chloroplast structure, chlorophyll content and light transmission of mature leaves of some deciduos tree seedings. Botanical Bulletin of Academia Sinica, Taipei, 30: 133-142
Lima PCF (1994) Comportamento silvicultural de espécies de Prosopis, em Petrolina-PE, região semi-árida brasileira. Curitiba,Paraná, Brasil, 110pp Tese (Doutorado) Universidade Federal do Paraná
Morita O, Goto M and Ehara H (1994) Growth and dry matter production of pasture plants grown under reduced light conditions of summer season. Bulletin of the Faculty of Bioresources, Mie University, 12: 11-20
National Research Council (1984) Nutrient requirements of beef cattle. National Academy Press, 6th edition. Washington DC, 90 pp
Sanchez P and Palm C (1996) Nitrogen and phosphorus in African soils: what role for agroforestry? Agroforestry Today, 8: 14-16
Valenzuela HC, O’hair SK and Schaffer B (1991) Shading, growth and dry matter partitioning of cocoyam [Xanthosoma sagittifolium (L.) Schott]. Journal of American Society of Horticultural Science, East Melbourne, 116: 1117-1121
Michaela Schaller[153], Götz Schroth[154], John Beer[155], Francisco Jiménez[156]
Key words: root barriers, root competition, root management
The sensitivity of trees to root competition from grasses for nutrients and water, which often results in lower survival and growth rates, is a common problem in horticulture and forestry as well as for the establishment of trees in degraded pastures. On the other hand, grass barriers could be used to limit the lateral extension of tree root systems; e.g., grass root systems can restrict the lateral and increase the vertical extension of tree root systems (Atkinson et al., 1978). Increased water and nutrient uptake of the trees from the subsoil could reduce competition with adjacent crops. An increased rooting depth of the trees could also have a stabilizing effect against soil shear. This is an example of one of the methods designed to optimize root interactions in agroforestry, which have collectively been termed “root management” (Schroth, 1999).
This study focused on the effect of three competitive grasses, Panicum maximum (guinea grass), Brachiaria brizantha (brachiaria) and Saccharum spp. (sugarcane), planted in three different widths (one, two or three rows), on the lateral extension of the root system of Eucalyptus deglupta, a fast-growing timber tree with a superficial root system (Schaller et al., 1999) that is increasingly used in agroforestry associations and reforestation projects in Central America and other parts of the humid tropics (CATIE, 1994).
In October 1998, a field trial was established in the experimental area “La Montaña” of CATIE in Turrialba, Costa Rica (9°53’ N.; 83°38’ W.; 602 m.a.s.l.; 2684 mm annual rainfall, usually only one dry month (March); average annual temperature 21.7°C; Aquandic Dystrudept; 0% slope). Ten treatments were established in a randomized complete block design with four repetitions: lines of four E. deglupta trees were planted with a 3 m within-row spacing between one, two or three rows (at 90, 110 and 130 cm from the trees, respectively) of P. maximum, B. brizantha or Saccharum, and in a control treatment without barriers. B. brizantha and P. maximum were established from seeds and Saccharum from stem cuttings. The trees were planted from three-month-old, bagged nursery seedlings three weeks after planting the grasses. The barriers were cut regularly to prevent shading of the trees.
Tree height and stem diameter were measured three times: in May (at 10 cm from the soil surface) and September 1999 and March 2000 (dbh). Soil cores (0-20 and 20-40 cm depth) were taken in February 2000 for the determination of grass and tree root length densities within the grass strips and at half-distance between these and the tree line. Root profiles were cut at 1.50 m distance from the trees in all four directions (October 1999 until March 2000). The profile walls behind the grass barriers (“barrier profiles”) had a width of 3 m and a depth of 0.8 m. The profiles between the trees (“between-tree profiles”) measured approximately 1.5 m in width and 0.8 m in depth. The roots were exposed with a fork or a screw-driver and the tree roots were mapped on graph paper with the help of a 10 x 10 cm grid, distinguishing between root diameter classes of <2 mm, 2-5 mm and >5 mm; the grass roots were only qualitatively evaluated in the profiles. The resistance of the top soil (0-10 cm) in the barrier profiles was measured with a penotrometer.
Analysis of variance for a two-factorial (3x3) design with three grass species and three strip widths, plus the control (without width-subtreatments) was initially used. As strip width did not prove to be a significant factor, analyses of variance for a randomized complete block design with ten individual treatments, and contrast analyses, were used to compare groups of treatments. In case of significance of the F-test at p<0.05, treatment means were compared with least significance tests at the same level of probability.
Tree growth was clearly limited by the grass strips, with the strongest reduction occurring in the P. maximum treatments (p=0.015 for contrast analysis), but the effect decreased with time (Table 1). The total number of tree roots in the P. maximum treatments was 40% less than in the no-grass control. Hence, reduced initial tree growth was probably caused by limited tree root development as is indicated by significant correlations between total tree roots and tree diameters (dbh) of 0.90 and 0.80 for the first and second block, respectively (but only 0.48 for the third block).
Table 1. Stem diameters (cm) of Eucalyptus deglupta trees planted between strips of Panicum maximum, Brachiaria brizantha and Saccharum spp., and without grass strips (control), in May 1999 (D1, measured at 10 cm from the soil surface), September 1999 (D2, measured at 130 cm from the soil surface, dbh) and March 2000 (D3, dbh).
|
Treatment |
D1 |
D2 |
D3 |
|
P. maximum |
2.5 (0.1)*) |
3.1 (0.3) |
6.6 (0.3) |
|
B. brizantha |
3.3 (0.2) |
4.0 (0.2) |
7.2 (0.3) |
|
Saccharum |
3.0 (0.2) |
4.5 (0.3) |
7.8 (0.3) |
|
Control |
3.9 (0.4) |
4.8 (0.8) |
8.5 (1.1) |
*) standard errors in brackets
The tree roots were significantly (p=0.008) more superficial in the barrier profiles than in the between-tree profiles. In the profiles beyond the barriers, 83% of total tree roots (s.d.=0.15) were in 0-10 cm in contrast to 76% (s.d.=0.13) in the profiles between the trees (for 0-30 cm, 96% and 94%, respectively). Tree roots were more superficial in the more restrictive barrier treatments (p=0.0001). Thus, none of the grasses had forced the E. deglupta roots to grow at greater depths; on the contrary, it seems as if the opposite occurred. The more competitive grasses caused, however, an asymmetry in the development of the tree root systems, resulting in an accumulation of tree roots between the trees due to their lateral restriction by the grass barriers.
At 0-30 cm, counts of tree fine roots (d<2 mm; 92% of total roots; s.d.=4; Table 2a) in the profile beyond the barriers were significantly lower in the P. maximum treatments than in the control (p=0.01 for contrast analysis), which corresponds with the reduction in total tree roots in the P. maximum treatments (see above). However, root ratios, (i.e., the number of roots in the barrier profile divided by total roots), were also significantly lower in the P. maximum treatments than in the control (p=0.004 for contrast analysis) suggesting that the reduced number of tree roots beyond the barriers was not just due to a reduction in the total number of tree roots but also due to a redistribution of rooting zones. In the B. brizantha treatments, the lateral extension of these fine roots was also significantly reduced in comparison to the control treatment (p=0.026 for contrast analysis) but the Saccharum barriers, in contrast, did not restrict lateral tree root extension. The root barrier effect of the grasses seems to be related to grass root densities, as P. maximum had significantly higher (p=0.005) root length densities than the other grasses, and Saccharum had the lowest root length densities (11.2 vs. 3.4 cm cm-3 for 0-20 cm in the grass strip, respectively). Grass strip width did not have a significant effect on tree root development.
Table 2. Number of Eucalyptus deglupta roots in the barrier profile (No. dm-2) and root ratios at 0-30 cm (number of E. deglupta roots in the barrier profile divided by total roots); P=Panicum maximum; B=Brachiaria brizantha; S=Saccharum spp.; C=Control; 1-3=strip intensities.
a) Roots with d<2 mm diameter (p=0.034 and p=0.08 for root numbers and root ratios, respectively)
|
Treatment |
P1 |
P2 |
P3 |
B1 |
B2 |
B3 |
S1 |
S2 |
S3 |
C |
|
Number of roots |
3.2 |
2.7 |
3.1 |
5.7 |
4.1 |
3.8 |
8.2 |
5.4 |
9.0 |
6.0 |
|
Root ratio |
0.247 |
0.263 |
0.260 |
0.385 |
0.273 |
0.257 |
0.425 |
0.357 |
0.375 |
0.403 |
b) Roots with d>5 mm diameter (p=0.30 and p=0.81 for root numbers and root ratios, respectively)
|
Treatment |
P1 |
P2 |
P3 |
B1 |
B2 |
B3 |
S1 |
S2 |
S3 |
C |
|
Number of roots |
0.037 |
0.037 |
0.033 |
0.034 |
0.045 |
0.011 |
0.084 |
0.018 |
0.078 |
0.045 |
|
Root ratio |
0.72 |
0.49 |
0.84 |
0.54 |
0.68 |
0.42 |
0.57 |
0.28 |
0.46 |
0.73 |
*) standard errors in brackets (n=3 except for B1, S1 and S3, where n=2)
The tree roots with a diameter between 2 and 5 mm (approximately 8% of total roots; s.d.=4) responded similarly to the different barrier treatments as did the fine roots (<2 mm), with the exception that their lateral extension was more restricted as indicated by slightly lower root ratios (data not shown). Lateral restriction of larger tree roots (> 5 mm;<1% of total roots; Table 2b) was, in contrast, not detected. Their root ratios were, in general, approximately twice as high as for the smaller root diameter classes except for Saccharum. The P. maximum treatments had relatively high root ratios for this root class in comparison to the other treatments. This might indicate a strategy of the Eucalyptus tree root system to reach beyond the area of influence of the barriers and may be a result of intra-tree competition within the tree line.
There was a significant time effect, e.g., effect of tree age (p=0.0001), with an increase of total tree roots accompanied by a decrease in the efficiency of the grass strips to limit the lateral tree root extension - especially for the treatments that were initially more efficient in restricting lateral tree root extension. This correlated with an increase in tree growth in the respective treatments (see above). Thus tree roots may only be temporarily inhibited from passing the grass strips; i.e., their growth through the grass strips may only be delayed, depending on the competitiveness and extension of the barrier. The intraspecific competition between the trees will increase with tree age (except where there is natural or artificial thinning of the trees), whereas the intensity of the competition exerted by the grass strips will be approximately constant or even decrease due to shading and increasing competition from the trees reducing grass vigor. However, in plots with a compacted top-soil, hardly any tree roots passed the grass strips at the time of the evaluation. This suggests that the efficiency of the grass root barriers is increased if there is some other limitation on tree root development; e.g., low contents of nutrients and/or water.
The extension of lateral roots of 12 to 16 month old E. deglupta was restricted when trees were planted between strips of the competitive grasses P. maximum and B. brizantha, in comparison to the control treatment without grasses and Saccharum strips. Limited tree root development beyond grass strips should reduce tree root competition with associated crops, and the accumulation of the tree roots between the barriers should contribute to the soil stabilization within the grass strips. Barrier strip width did not significantly affect lateral tree root extension and there was some evidence of a decrease in the efficiency of the barriers to restrict lateral tree root extension with time; i.e., with increasing tree development and intraspecific competition between the tree component. This needs to be studied in more detail. Management practices, such as planting distances of the grass strips and frequency and degree of grass clipping, probably have an important influence on the performance of the grass root barriers. It is also possible that under conditions of less precipitation or less fertile soils, (i.e., under conditions of increased competition for restricted soil resources) that the efficiency of the barriers in restricting lateral tree root extension would be altered.
This work was funded by the “Tropical Ecology Support Program” of the Deutsche Gesellschaft fuer Technische Zusammenarbeit (GTZ) under project number 96.2151.7-00.107.
Atkinson D, Johnson MG, Mattam D and Mercer ER (1978) The effect of orchard soil management on the uptake of nitrogen by established apple trees. J Sci Food Agric 30: 129-135
CATIE (1994) Deglupta. Eucalyptus deglupta Blume, Especie de árbol de uso múltiple en América Central. Turrialba, Costa Rica, CATIE. 43 p. (Serie Técnica. Informe Técnico No. 240)
Schaller M, Schroth G, Beer J and Jiménez F (1999) Control de crecimiento lateral de las raíces de especies maderables de rápido crecimiento utilizando gramíneas como barreras biológicas. Agroforestería en las Américas 6 (23): 36-38
Schroth G (1999) A review of belowground interactions in agroforestry, focussing on mechanisms and management options. Agrofor Syst 43 (1-3): 5-34
M. Dulormne[157], J. Sierra[158]*, P. Nygren[159], P. Cruz[160]
Key words: carbon sequestration, Gliricidia sepium, nitrogen mineralization, 15N natural abundance, pruning, tree roots
The association of leguminous trees and forage grasses has received increasing attention in tropical agroforestry. This system offers at least two significant advantages: i- an increase in land-use efficiency (Benavides et al., 1989; Cruz et al., 1993), and ii- a balanced diet with legume browse as the protein source and grass as the energy source (Archimède et al., 2001). Another advantage observed by several authors is the increase in soil N content mainly associated with the recycling of fixed nitrogen via leaf litter and/or animal excreta (e.g. Sadeghian et al., 1998). The extent of this effect depends upon how the system is managed; e.g. the presence of animals and the pruning regime of the tree (Kass et al., 1997). Because N export from fodder production systems may be high, especially if forage trees and grasses are grown in the same plot (Nygren and Cruz, 1998), there are concerns about the sustainability of cut-and-carry systems. If N2 fixation of trees is impeded by the pruning regime, and nutrient export is high, rapid degradation of soil nutrients is possible. Thus, understanding the dynamics of N2 fixation by legume trees and the turnover of the fixed N is important for optimal development and management of these systems.
This paper summarizes the results of several studies dealing with the N dynamics in a tropical cut-and-carry system. The objectives of these studies were: i- to analyze the effects of season and pruning regime on N2 fixation of the woody legume (Gliricidia sepium (Jacq.) Kunth ex Walp.); ii- to estimate the recycling rate of the fixed N in the soil; and iii- to quantify the mineralization potential of the recycled N.
Study site
The study was conducted at the experimental station of the Institut National de la Recherche Agronomique (INRA) in Godet (16°20’N, 61°30’W), Guadeloupe (French Antilles). The soil is a Vertisol (pH 7.8) and the climate is warm and subhumid (annual rainfall 1300 mm) with a dry season from February to June. The plot was established in 1989 by planting G. sepium cuttings at 0.3 m (2 m spacing in natural grassland of Dichanthium aristatum (Poir) C.E. Hubbard. The plot was managed as a cut-and-carry system where all the tree prunings and cut grass were removed. No N fertilizer was applied.
Plant measurements
Trees were completely pruned prior to the beginning of the N dynamics experiment; thereafter, three pruning regimes were initiated (Nygren et al., 2000a): total removal of all leaves and branches above 0.5 m once a year (T12), or every six months (T6), and partial pruning every two months (P2; defoliation intensity 45%). The standing biomass of the tree and its N concentration was measured every two months during one year. Nodule biomass in the 0-0.2 m soil layer was sampled twice, two and six months after the pruning corresponding to the T6 regime. For this, undisturbed soil blocks of 1 dm3 were removed at 0.25 m and 0.75 m from the hedgerows on both sides of the tree rows.
Nodules were cut and then classified as fixing (pink or red interiors) and non-fixing (Nygren et al., 2000a). The litter bag technique was used to determine the nodule disappearance and nitrogen release rate (Nygren et al., 2000b). Dinitrogen fixation by G. sepium was estimated by the 15N natural abundance method using Gmelina arborea Roxb. as the non-N2-fixing reference plant (Nygren et al., 2000a). The measurements were carried out six times during the experiment and corresponded to the pruning dates of the P2 regime. At each sampling date, the youngest fully developed leaves were collected for analysis of total N content (Elemental Analyzer) and 15N/14N ratio (Mass Spectrometer). The d15N value of G. sepium in N-free medium (-2.07) was determined in other experiment (Nygren et al., 2000c). Above-ground grass biomass and its N concentration were determined every 40-50 days for two years.
Soil measurements
Soil organic N in the 0-0.1 m and in the 0.1-0.2 m layers was determined in 1999 on samples taken at several locations between and within tree rows. Additional measurements were carried out on soil samples taken on a transect through a 20 m wide subplot kept under natural grassland since 1989 (open grassland). The open grassland was situated in the same plot between two silvopastoral subplots. The subplots were not trenched.
Nitrogen mineralization rates were estimated on soil samples taken in the open grassland and in an adjacent silvopastoral subplot. Samples were aerobically incubated (Sierra and Marbán, 2000) for 30 days in an experiment with a factorial design of temperature (20°C, 30°C and 40°C) (soil moisture (-1500 kPa, -300 kPa and -30 kPa). Soil temperature and moisture were monitored in both subplots for two years in order to estimate in-situ N mineralization.
Dinitrogen fixation
The pruning regimes had little effect on the proportion of the above-ground N coming from N2 fixation; however, in absolute terms, the T6 regime produced the highest amount of fixed N (Table 1). Fixed N comprised 54-87, 54-92, and 60-87% in the T12, T6 and P2 regimes, respectively, of the total N in aboveground tree biomass, depending on sampling date (Table 1). Total annual accumulation of N in harvestable aboveground biomass was highest in trees in the T6 regime. In all treatments, about 70% of the N exported per year from the plot in the fodder harvest came from N2 fixation. The highest percentages of N2 fixation observed corresponded to periods of high foliage biomass production and periods when trees had high nodule:foliage biomass ratio. Flowering decreased foliage biomass production and total N2 fixation of trees in the T12 regime, which is unsuitable for forage production because of low foliage harvest. Foliage harvest in the P2 regime was satisfactory and well distributed throughout the year, but the total biomass production and amount of N2 fixation was partially affected by pruning. The results showed that renodulation and onset of N2 fixation following pruning occurred rapidly in G. sepium.
Table 1. Total N increment and N coming from N2 fixation in aboveground biomass of Gliricidia sepium during a 2-month period coinciding with the interval between pruning events in the P2 regime (T12=total pruning every 12 months, T6=total pruning every 6 months and P2=50% pruning every 2 months). The negative values (foliage loss) are considered as zero for the calculation of the totals.
|
Pruning date |
Weeks of regrowth |
Total N increment (kg ha-1) |
Total fixed (kg ha-1) |
||||
|
|
|
P2 |
T6 |
T12 |
P2 |
T6 |
T12 |
|
May 30 |
8 |
38.8 |
30.2 |
22.6 |
26.8 |
24.1 |
16.0 |
|
Jul 25 |
16 |
55.4 |
52.7 |
49.1 |
48.2 |
28.5 |
26.5 |
|
Sep 19 |
24 |
33.7 |
33.9 |
35.4 |
27.6 |
31.2 |
30.8 |
|
Nov 18 |
33 |
48.7 |
36.6 |
86.5 |
29.2 |
23.4 |
53.6 |
|
Jan 9 |
40 |
-2.3 |
44.7 |
-30.6 |
- |
31.7 |
- |
|
Mar 10 |
49 |
17.9 |
114.9 |
29.6 |
15.4 |
65.5 |
18.9 |
|
Total |
|
194.5 |
312.9 |
223.2 |
147.2 |
204.4 |
146.0 |
Soil nitrogen balance and nitrogen mineralization
Significant differences (P < 0.05) for soil organic N were found between subplots and within the silvopastoral subplot. For the 0-0.2 m soil layer, organic N decreased in the following order: tree row (2.9±0.1 g kg-1) > inter-row (2.7±0.2 g kg-1) > open grassland (2.2±0.2 g kg-1). Organic N in 1989, prior to the installation of the silvopastoral system, was 2.0 g kg-1. These results corresponded to an increase of approximately 180 kg N ha-1 yr-1 in the soil under the silvopastoral system and 60 kg N ha-1 yr-1 under the open grassland. Organic carbon and N increased in the same proportion and therefore the C/N ratio of the soil (»11) did not vary in the silvopastoral subplot or in the open grassland. Therefore, soil carbon sequestration averaged 2 Mg [C] ha-1 yr-1. In the open grassland, organic N diminished as a function of distance from the tree row, and the increase was slight in the middle of the subplot (Figure 1).
Figure 1. Soil organic N in the open grassland as a function
of distance from the legume tree in Guadeloupe (French Antilles). Vertical bars
indicate the experimental error.
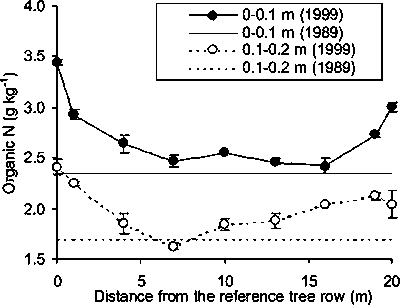
This suggests that the increase in organic N was directly associated with the turnover of the N fixed by the tree (Dulormne et al., 2001). The N release rate from the nodules was high (half-life 3-5 days) but the total amount of N released was small (»10 kg N ha-1 yr-1; Nygren et al., 2000b) due to the low nodule biomass. As litterfall was small (Figure 2) and animals were excluded in our system, it seems clear that most of the N recycled came from the turnover of tree roots. Therefore, the gradient of organic N observed in the open grassland could reflect the gradient of tree root density below this subplot.
Figure 2. Annual N balance in the silvopastoral system in Guadeloupe (French Antilles).
Nitrogen export from the silvopastoral system was about 500 kg [N] ha-1 yr-1 (300 kg [N] ha-1 yr-1 from tree pruning in the T6 regime and 200 kg [N] ha-1 yr-1 from grass cutting; Figure 2). Independently of the source of N exported from the system (from soil N or from biological fixation), these values may be used to estimate N fixation by G. sepium: N2 fixation = N exports by grass cutting + N exports by tree pruning + Increase in soil organic N. Thereby, N2 fixation averaged 680 kg N ha-1 yr-1. This value is within the large range of values reported by Peoples and Herridge (1990) for several woody legumes in the Tropics.
Mineralization rates were higher in the soil of the silvopastoral system (e.g. 3.7 times at 40°C and 1.6 times at 20°C). Soil moisture had no effect on N mineralization in the range of water content tested in this study. In spite of a higher soil temperature (2-3°C) in the open grassland, the estimate of in-situ N mineralization was 20% higher in the silvopastoral subplot because of its higher substrate content. Grass production in the silvopastoral subplot averaged 13 Mg ha-1 yr-1 (15 Mg ha-1 yr-1 in the open grassland), 20% of which was associated to the higher soil N availability (Dulormne, 2001).
Dinitrogen fixation makes an important contribution to the N economy of the G. sepium-D. aristatum association studied in this work and indicates its suitability for sustainable forage production. Our results showed that G. sepium trees subjected to periodic pruning actively fixed atmospheric N2. This characteristic and the ability of the system to increase soil nitrogen level and to favor C sequestration contributed to the high productivity of the G. sepium-D. aristatum association over ten years of intensive harvesting. The recycled N fixed by the tree is an important compartment of the organic soil N and supplies significant amount of N to the grass. Further effort is necessary to understand the effects of tree roots and their temporal dynamics on the turnover of the fixed N.
Archimède H, Dulormne M, Tournebize R, Saminadin G, Periacarpin F and Xandé A (2001) The effects of Gliricidia sepium supplementation on intake and digestion of Digitaria decumbens hay by black belly sheep. J Agric Sci (Cambridge) (in press)
Benavides J, Rodríguez R and Borel R (1989) Producción y calidad del pasto King grass (Penisetum purpureum (P. thypoides) y poró (Erythrina poeppigiana) sembrados en asociación. In: Xandé A and Alexandre G (eds) Pâturages et alimentation des ruminants en zone tropicales humide, pp 367-376. INRA, Paris, France
Cruz P, Munier-Jolian NM, Tournebize R and Sinoquet H (1993) Growth and mineral nutrition of a Dichanthium aristatum sward shaded by trees. In: Proc XVII International Grassland Congress, pp 2056-2057. Rockhampton, Australia
Dulormne M (2001) Analyse de l’effet ombrage dans un système agroforestier légumineuse arbustive - herbe. Ph. D. thesis, University Paris Sud, France, 120 pp
Dulormne M, Sierra J, Sophie SA and Solvar F (2001) Capacidad de secuestración del carbono y del nitrógeno en un sistema agroforestal a base de Gliricidia sepium en clima tropical sub-húmedo (Guadalupe, Antillas Francesas). In: Sánchez MD and Rosales M (eds) Proc Segunda Conferencia electrónica de la FAO sobre Agroforestería para la producción animal en Latinoamérica. FAO, Rome, Italy (in press).
Kass DCL, Sylvester-Bradley R and Nygren P (1997) The role of nitrogen fixation and nutrient supply in some agroforestry systems of the Americas. Soil Biol Biochem 29:775-785
Nygren P and Cruz P (1998) Biomass allocation and nodulation of Gliricidia sepium under two cut-and-carry forage production regimes. Agrofor Syst 41: 277-292
Nygren P, Cruz P, Domenach AM, Vaillant V and Sierra J (2000a) Influence of forage harvesting regimes on dynamics of biological dinitrogen fixation of a tropical woody legume. Tree Physiol 20: 41-48
Nygren P, Lorenzo A and Cruz P (2000b) Decomposition of woody legume nodules in two tree/grass associations under contrasting environmental conditions. Agrofor Syst 48: 229-244
Nygren P, Vaillant V, Desfontaines L, Cruz P and Domenach AM (2000c) Effects of nitrogen source and defoliation on growth and biological dinitrogen fixation of Gliricidia sepium seedlings. Tree Physiol 20: 23-30
Peoples MB and Herridge DF (1990) Nitrogen fixation by legumes in tropical and subtropical agriculture. Adv Agron 44: 155-223
Sadeghian S, Rivera JM and Gómez ME (1998) Impacto de sistemas de ganadería sobre las características físicas, químicas y biológicas de suelos en los Andes de Colombia. In: Sánchez MD and Rosales M (eds) Proc Conferencia electrónica de la FAO sobre Agroforestería para la producción animal en Latinoamérica, pp 77-95. FAO, Rome, Italy
Sierra J and Marban L (2000) Nitrogen Mineralization Pattern of an Oxisol of Guadeloupe (French West Indies). Soil Sci Soc Am J 64: 2002-2010
Alejandra E. Vilela[161], Damián A. Ravetta[162]
Management of any system is a matter of making decisions and taking actions that are intended to have particular effects. Management actions taken without the ability to predict the way the system is likely to react are mostly guesswork. To predict the results of any particular action, the manager must understand how the system works (Landsberg & Gower, l997) and this is what ecology and ecophysiology have to offer to agroforestry research. When a silvopastoral system is going to be established many questions arise such as: Which species? How long will it take for the system to get established and be productive? Which products? What levels of production can we obtain, etc. The answer to some of these questions can be found with the aid of ecophysiological research. Physiology can provide the tools to determine the mechanisms through which the systems react to a set of abiotic and biotic conditions, and predict responses under different management conditions.
A simple example of a management action in nurseries is the practice of shading seedlings. Seedlings are shaded to protected them from above-optimal radiation intensity which can result in the plants dead, but, at least in some species, at the cost of reductions in plant growth. The balance between potential benefits and costs of shading in nurseries is species - dependent.
The objective of this paper is to show that the knowledge about species physiological responses under certain environmental conditions could help focusing efforts to design proper management practices, saving time, money, and man-power. Two ecophysilogical studies on the genus Prosopis are given as examples of the utility of the approach and in a third example, preliminary observations and data are given on the influence of a biotic factor on product yield as well as questions that need to be answered to understand the underlying mechanisms.
First example
Seedlings were grown in a greenhouse at 14/38°C (night-day). Plants were assigned to one of three light environments in a randomized complete block design (three blocks and 10 reps/block): 1) direct solar radiation (full-sun); 2)47% reduction in direct solar radiation (53%-sun) and 3) 62% reduction in direct solar radiation (38%-sun). At the end of the experiment (60 days after seeded) plants were harvested, and leaves, stems and roots separated and weighed (10 plants per species and light treatment). Total non-structural carbohydrates (TNC) were extracted and determined by the anthrone method (Yemm & Wills, 1954). Protein content was determined by Kjeldahl. The C:N ratio was determined by computing the amount of TNC (C) and nitrogen content (N) respectively.
Second example
Seedlings were grown in a greenhouse at 20/30°C (night-day). A factorial experiment with two factors (water and nitrogen) was carried out. Water availability had two levels: Low water availability (W0, irrigation at 37 to 59 day intervals, depending on the species; water was applied when significant differences were found in CO2 assimilation between this plants and plants under W1) and high water availability (W1,irrigation every other day). Nitrogen availability had four levels: N0 (no nitrogen); N50 (addition of 50 mg N seedling-1); N100 (addition of 100 mg N seedling-1) and N200 (addition of 200 mg N seedling-1), applied throughout a 5 mo period.. Plants were randomly assigned to one of these eight treatments (15 reps/treatment). TNC and N were determined as in the first example.
Third example
Pods from 5 populations of Prosopis chilensis and P. flexuosa (5 individual per population, 10 pods per tree) were harvested. Total number or seeds per pod, number of bruchid-attacked seeds, pod and seed dry weight, and exo-mesocarp gum content were recorded. Gum content was determined according to Kalman (2000).
First example
The effect of radiation availability on seedling growth and physiology. Differences in radiation availability resulted in significant differences in biomass accumulation (p<0.01). Total biomass was reduced, all species averaged, 24% and 38% for plants under 52%-sun and 38%-sun respectively (Fig.1). The shoot/root ratio significantly increased with reductions in radiation availability (p<0.05). TNC content in roots decreased with reductions in light availability. There was a clear reduction in C:N ratio with reduced radiation availability. Plant survival in Prosopis proved to be positively related to total biomass and root TNC and inversely related to shoot:root ratio (Vilela & Ravetta, in press). In Prosopis the occurrence of C-based secondary metabolites (gums) has been proposed as a defense mechanism against invading microrganisms (Espejel, l981; Kalman, 2000). Therefore, the reduction in C:N ratio found in our experiment should decrease the defense capability of plants grown under reduced radiation. Although shading is a common practice in Prosopis nurseries, under certain environmental conditions it reduces growth and alters certain physiological variables that should lessen the chances of Prosopis survival in the field, since vigour and survival of seedlings are directly related to size, biomass allocation and the amount of stored carbohydrates (Puttonen,1986)
Figure 1. Total dry weight (g plant-1) of 60-days
plants of Prosopis chilensis, P. flexuosa, P. glandulosa and P. alba.
Mean +- S.E. for 10 plants

Second example
The effect of nitrogen and water availability on seedling growth and physiology. Plants under low water availability (W0) did not respond to fertilization by increasing total biomass. Low-resource species (P. strombulifera and P. flexuosa) under W1did not increase total biomass with nutrient addition, while P. alba, a higher-resource species, did (Fig.2). All species, except for P. strombulifera, increased biomass 2-4 times when grown under W1. Fertilization did not change dry matter partitioning or the shoot/root ratio.
Increasing N availability decreased root TNC content and C:N ratio. Low-resource species resisted 44-59 days without irrigation, without showing a decline in CO2 assimilation, while P. alba after 37 days showed significant differences in CO2 assimilation between plants under W1 and W0. Although fertilization usually promotes seedlings growth, the plasticity in the response of Prosopis species to nutrient availability seemed to be related to the resource availability in each species’ native habitat, as hypothesized by Chapin (1980). Growth and survival-related variables in Prosopis species seem to be more responsive to water availability than to nitrogen supply.
Figure 2. Total dry weight (g plant-1) of five
month plants of Prosopis alba, P. flexuosa-tree, P.flexuosa-shrub and
P.strombulifera. Mean +- S.E. for 10 plants. a) water stressed plants (W0);
b) non-water stressed plants.
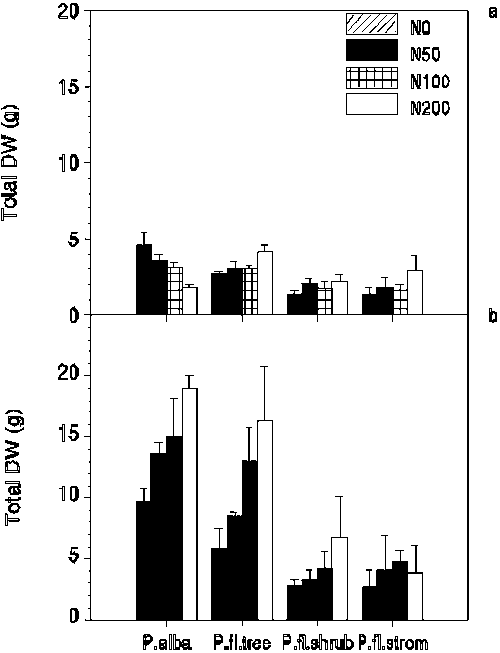
Third example
The effect of insect attack on pods’ gum content. Gums are a constitutive component of South-American Prosopis species pods' epi-mesocarp. Constitutive gum contents range from 0.6-17.1% depending on the species. Prosopis pods are attacked by beetles (Bruchidae) that can completely destroy a years' seed-crop. Insect attack seems to promote gum production in the epi-mesocarp (Kalman, 2000). For Prosopis flexuosa, a positive relationship between insect attack and epi-mesocarp gum content was found (r2=0.71). On the other hand, a negative relationship was found between pod dry weight (pod size) and gum content (r2=0.42). A trade-off between pod production (kg plant.-1) and pod gum content was found. The extent of insect attack is very difficult to manage in natural environments, but are there other ways of increasing gum accumulation? May accumulation be promoted managing resource supply? Is pod gum content mostly a defense reaction against insect attack or the result of the interaction of biotic and abiotic factors (nutrients, water, temperature direct and indirect effects, etc.)? Most of the gum is produced in the exo-mesocarp fraction of the pod; How can the proportion of this fraction in the fruit be maximized? Does resource availability modify the fruit's partition? These and other questions need to be answered to understand how pod-gum regulates seed attack by bruchids, a process essential to Prosopis natural o naturalized stands and an important mechanism for gum production.
The development of production alternatives in the tropics and in arid lands is still a challenge. Agroforestry and silvopastoral systems are proposed as sustainable practices for these areas. A great amount of the research on these systems goes into the description of each particular species potentials, how it can be used economically and its yields under different management conditions. The results obtained cannot be extrapolated to other areas since very rarely is there information available on the mechanisms behind the observed responses, which are basic for the prediction of general response patterns. Agroforestry research is carried out at a number of levels; we propose that ecophysiological studies should be among the very first steps in silvopastoral/agroforestry research since they can provide the ability to predict the way a system will respond, at a low research cost and avoiding the trial and error approach. Even if our particular plant species/system combination does not make it into the field we will have produced information that can be used in other fields of knowledge.
Espejel, I.1981. Técnicas para muestrear y medir la producción de goma de mezquite. Inventarios de recursos de tierras áridas. USDA. Forest Service General Technical Report WO-28. 451-454.
Kalman, D. 2000. Variabilidad interespecífica e interpoblacional en el contenido de goma en frutos de Prosopis L. (Mimosaceae) y su relación con el ataque de insectos. Tesis para optar al título de Ingeniero Agrónomo de la Universidad de Buenos Aires.
Landsberg.J.J. & Gower, S.T. l997. Applications of Physiological Ecology to Forest Management. Ed. Academic Press, San Diego. pp 354
Puttonen, P. l986. Carbohydrate reserves in Pinus silvestris seedling needles as an attribute of seedling vigor. Scandinavian Journal of Forestry Resources 1:181-183
Vilela, A. & Ravetta, D. in press. The effect of seed scarification and soil-media on germination, growth, storage and survival of seedlings of five species of Prosopis L (Mimosaceae). Journal of Arid Environments
Yemm, E.W. & Villies, A.J. l954. The estimation of carbohydrates in plant extracts by anthrone. Biochemistry Journal 57:508-514
|
[95] Research-professor
Environmental Sciences Faculty, Universidad Tecnológica de Pereira,
Colombia e-mail: [email protected] [96] Research-professor, Area of Watersheds and Agroforestry Systems, CATIE, Turrialba, Costa Rica. e-mail: [email protected] [97] Research-professor Environmental Sciences Faculty, Universidad Tecnológica de Pereira, Colombia e-mail: [email protected], 2 Research professor, Area of Agroforestry, CATIE [98] Research-professor, Area of Watersheds and Agroforestry Systems, CATIE, Turrialba, Costa Rica. e-mail: [email protected] [99] Researcher, CATIE, Turrialba, Costa Rica, [email protected] [100] Researcher Professor, CATIE, Turrialba, Costa Rica, [email protected] [101] Universidad de Ciego de Avila. [102] Universidad de Ciego de Avila. [103] Universidad de Ciego de Avila. [104] Universidad de Ciego de Avila. [105] Universidad de Ciego de Avila. [106] Universidad de Ciego de Avila. [107] Centro de Investigaciones en Bioalimento. [108] El Colegio de la Frontera Sur, Unidad Villahermosa, México. [109] Institute of Ecology and Resource Management, University of Edinburgh, UK [110] Centre for Ecology and Hydrology, Penicuik, UK. [111] CIRAD-Forêt/CATIE, Apdo 31, 7170 Turrialba, Costa Rica [112] IRAD (Institut de la Recherche Agricole pour le Développement), BP 415 Garoua, Cameroon [113] IRAD (Institut de la Recherche Agricole pour le Développement), BP 415 Garoua, Cameroon [114] Instituto de Ciencia Animal, P.O.Box 24, La Habana, Cuba. [115] Dept. of Plant Production, Faculty of Agronomy and Veterinary, National University of Rio IV, Argentina [116] Dept. of Plant Production, Faculty of Agronomy and Veterinary, National University of Rio IV, Argentina [117] Dept. of Plant Production, Faculty of Agronomy and Veterinary, National University of Rio IV, Argentina [118] Dept. of Animal Production, Faculty of Agronomy and Veterinary, National University of Rio IV, Argentina [119] Dept. of Plant Production, Faculty of Agronomy and Veterinary, National University of Rio IV, Argentina [120] Instituto de Ciencia Animal, P.O.Box 24, San José de las Lajas, La Habana, Cuba. [121] Instituto de Ciencia Animal, P.O.Box 24, San José de las Lajas, La Habana, Cuba. [122] Instituto de Ciencia Animal, P.O.Box 24, San José de las Lajas, La Habana, Cuba. [123] Instituto de Ciencia Animal, P.O.Box 24, San José de las Lajas, La Habana, Cuba. [124] This study was partially financed by the Project for Use and Conservation of the Brazilian Biological Diversity - PROBIO, and the Alternatives to Slash and Burn Project - ASB [125] Researcher at Embrapa Acre, Caixa Postal 392, CEP 69908-970, Rio Branco, AC. Email: [email protected]; [email protected]; [email protected] [126] Researcher at Embrapa Acre, Caixa Postal 392, CEP 69908-970, Rio Branco, AC. Email: [email protected]; [email protected]; [email protected] [127] Researcher at Embrapa Acre, Caixa Postal 392, CEP 69908-970, Rio Branco, AC. Email: [email protected]; [email protected]; [email protected] [128] Researcher/Scholarship from Embrapa Acre/CNPq, Email: [email protected] [129] Agrarian University of the Havana. Post Office box 18, Cuba. e-mail: e-mail: [email protected] [130] Agrarian University of the Havana. Post Office box 18, Cuba. e-mail: e-mail: [email protected] [131] Agrarian University of the Havana. Post Office box 18, Cuba. e-mail: e-mail: [email protected] [132] Agrarian University of the Havana. Post Office box 18, Cuba. e-mail: e-mail: [email protected] [133] Agrarian University of the Havana. Post Office box 18, Cuba. e-mail: e-mail: [email protected] [134] Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León. Apartado Postal 41, Linares, Nuevo León 67700 México. E-mail: [email protected] [135] Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León. Apartado Postal 41, Linares, Nuevo León 67700 México. E-mail: [email protected] [136] E-mail: [email protected]. Urbanización Verdum. Santiago, Veraguas, Panamá. [137] Center for Agroforestry, 203 A-B Natural Resources Bldg., University of Missouri, Columbia, MO 65211, USA [138] Department of Forest Ecology, PO Box 27, 00014 University of Helsinki, Finland [139] Department of Forest Ecology, PO Box 27, 00014 University of Helsinki, Finland [140] Vantaa Research Centre, Finnish Forest Research Institute, PO Box 18, 01301 Vantaa, Finland [141] INRA, Centre Toulouse, Station d’Agronomie, BP 27, 31326 Castanet Tolosan Cédex, France [142] National Animal Production Research Institute, Ahmadu Bello University, P.M.B. 1096, Shika, Zaria, Nigeria.: (Corespondence: A. T. Omokanye, Centre for Farming Systems Reseach, University of Western Sydney, Hawkesbury, Locked Bag #1, Richmond NSW 2753, Australia. e-mail: [email protected]). [143] National Animal Production Research Institute, Ahmadu Bello University, P.M.B. 1096, Shika, Zaria, Nigeria.: Department of Animal Science, University of New England, Armidale, NSW, Australia. [144] 1 Experimental Station of Grass and Forages “Indio Hatuey” Central “España Republicana”, CP 44280. Matanzas, Cuba. Tel: 37 7510/37 7284. E-mail: [email protected]. [145] Faculty of the Agronomy and Veterinary. University of Río Cuarto, Pcia de Córdoba, Argentina. [146] Faculty of the Agronomy and Veterinary. University of Río Cuarto, Pcia de Córdoba, Argentina. [147] Faculty of the Agronomy and Veterinary. University of Río Cuarto, Pcia de Córdoba, Argentina. [148] Faculty of the Agronomy and Veterinary. University of Río Cuarto, Pcia de Córdoba, Argentina. [149] Faculty of the Agronomy and Veterinary. University of Río Cuarto, Pcia de Córdoba, Argentina. [150] Embrapa Florestas - Caixa Postal 319, CEP 83411-000, Colombo, PR. Brazil. E-mail: [email protected] [151] Embrapa Florestas - Caixa Postal 319, CEP 83411-000, Colombo, PR. Brazil. E-mail: [email protected] [152] Universidade Federal do Paraná. Rua São Tomé, 141, CEP 81540-320, Curitiba, PR. Brazil. E-mail: [email protected] [153] Centro Agronómico Tropical de Investigación y Enseñanza (CATIE), 7170 Turrialba, Costa Rica, Fax: (506) 556 7766, email: [email protected]; [email protected]; [email protected] [154] University of Hamburg, c/o Embrapa Amazônia Ocidental, C.P. 319, 69011-970 Manaus-AM, Brazil, Fax: (55) 92-622 1100, email: [email protected] [155] Centro Agronómico Tropical de Investigación y Enseñanza (CATIE), 7170 Turrialba, Costa Rica, Fax: (506) 556 7766, email: [email protected]; [email protected]; [email protected] [156] Centro Agronómico Tropical de Investigación y Enseñanza (CATIE), 7170 Turrialba, Costa Rica, Fax: (506) 556 7766, email: [email protected]; [email protected]; [email protected] [157] INRA, Unité Agropédoclimatique de la Zone Caraïbe, Domaine Duclos, 97170 Petit-Bourg, Guadeloupe, France. [158] INRA, Unité Agropédoclimatique de la Zone Caraïbe, Domaine Duclos, 97170 Petit-Bourg, Guadeloupe, France. [159] Center for Agroforestry, 203S Anheuser Busch Natural Resources Building, University of Missouri, Columbia, MO 65211, USA. [160] INRA, Station d’Agronomie de Toulouse, BP 27, 31326 Castanet-Tolosan, France. *Author for presentation and correspondence: [email protected] [161] IFEVA and Facultad de Agronomía, Universidad de Buenos Aires, Av. San Martín 4453 (1417) Buenos Aires, Argentina. E-mail: [email protected] [162] IFEVA and Facultad de Agronomía, Universidad de Buenos Aires, Av. San Martín 4453 (1417) Buenos Aires, Argentina. E-mail: [email protected] |
![]()
![]()
![]()
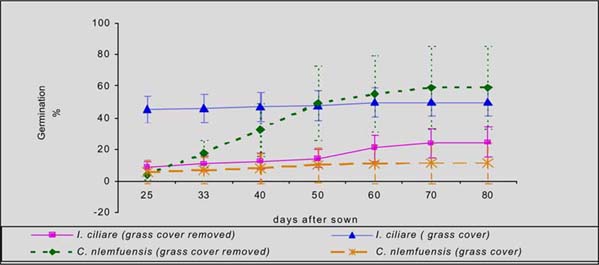{kind=link}
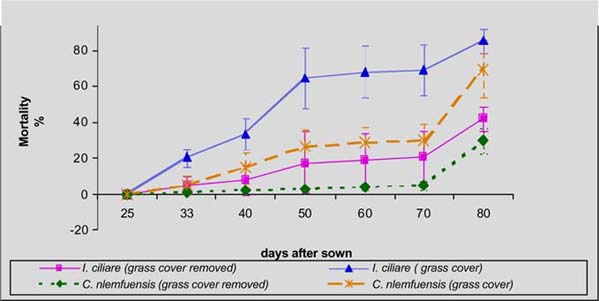{kind=link}
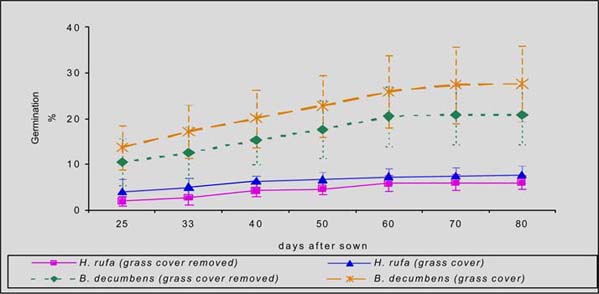{kind=link}
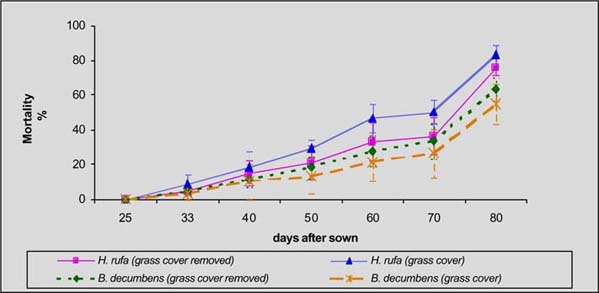{kind=link}
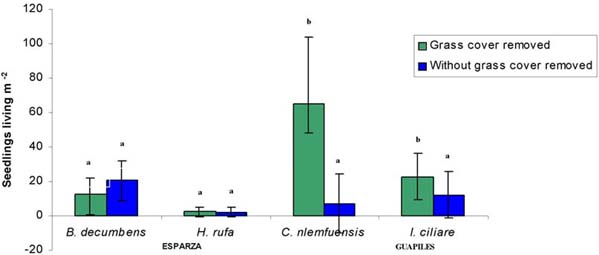{kind=link}
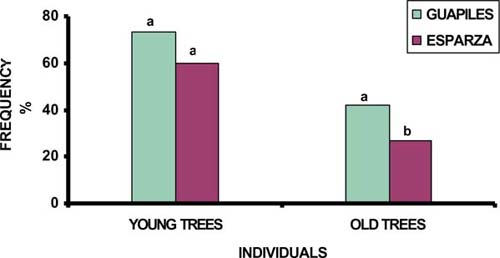{kind=link}
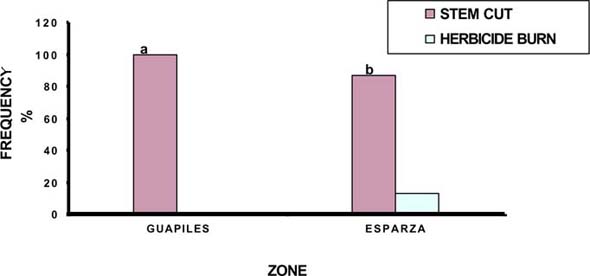{kind=link}
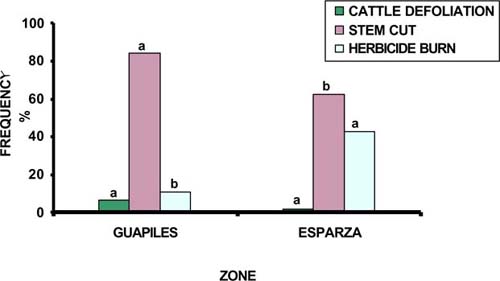{kind=link}
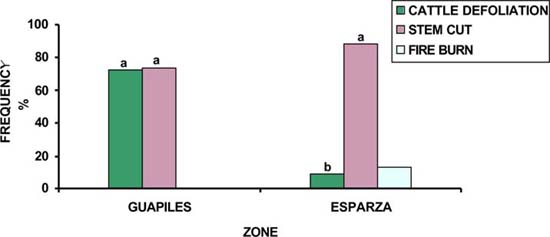{kind=link}
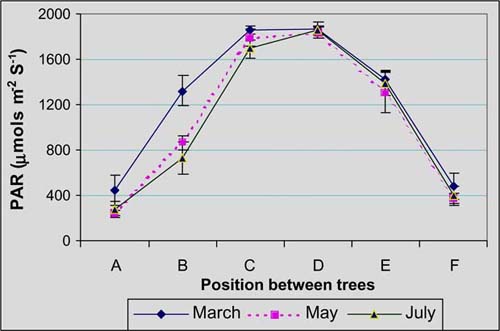{kind=link}
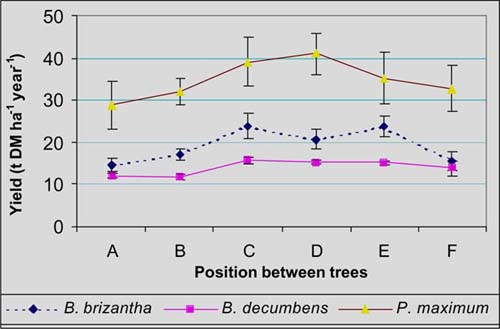{kind=link}
{kind=link}
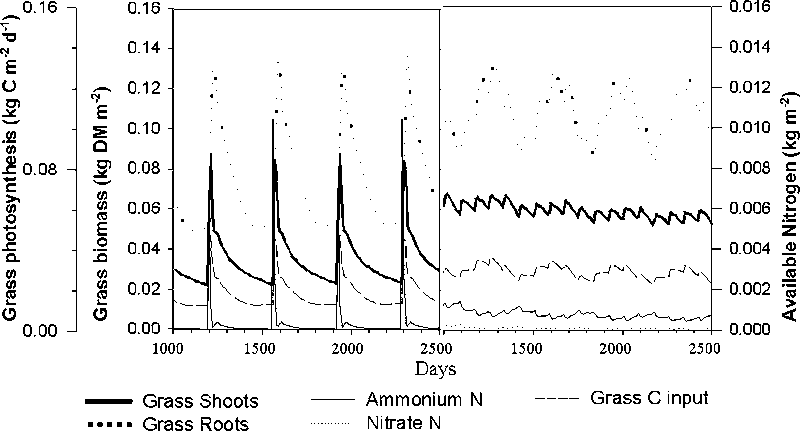{kind=link}
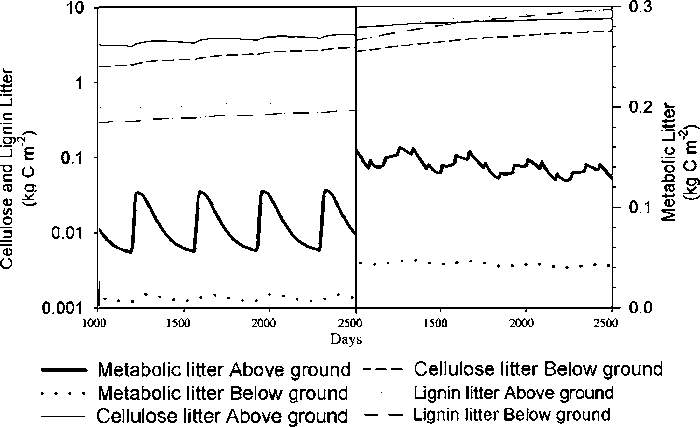{kind=link}
{kind=link}
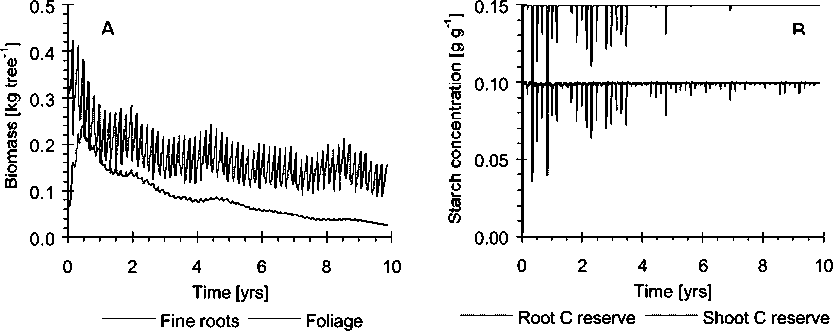{kind=link}
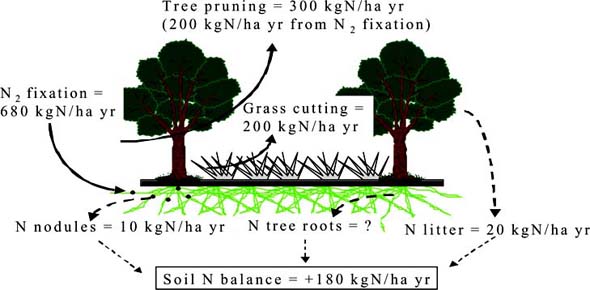{kind=link}