![]()
![]()
![]()
P. Quézel, F. Médail, R. Loisel et M. Barbero
Pierre Quézel, Frédéric Médail, Roger Loisel
et Marcel Barbero travaillent auprès de l'Université d'Aix-Marseille III, Faculté des sciences et techniques de Saint-Jérôme, Institut méditerranéen d'écologie et de paléoécologie, Marseille (France).
Evaluation de la biodiversité des forêts méditerranéennes et des défis pour leur conservation.
La diversité végétale des forêts méditerranéennes, beaucoup plus accusée que celle des forêts européennes, s'explique par des facteurs paléogéographiques (Verlaque et al., 1997) et historiques mais aussi par des critères écologiques actuels (Quézel, 1985). Par ailleurs, le monde méditerranéen, plus que toute autre région du monde, offre pour sa flore et pour ses paysages majeurs, d'étroites interrelations avec les activités humaines qui l'ont façonné depuis près de 10 000 ans (Thirgood, 1981; Pons et Quézel, 1985).
En fait, la biodiversité végétale méditerranéenne est le produit, pour beaucoup, d'une utilisation traditionnelle et harmonieuse du milieu par l'homme. Cependant, depuis la fin du XIXe siècle, cet équilibre a été perturbé soit par la surexploitation, soit par la déprise, dont les résultats, sans être identiques, n'en sont pas moins aussi dommageables du point de vue de la conservation des espèces et des habitats. En se focalisant sur les essences forestières majeures ou associées seront ainsi examinées: i) la richesse en espèces ligneuses des deux zones méditerranéennes de l'hémisphère Nord (Californie et bassin méditerranéen); ii) l'origine biogéographique des essences endémiques; et iii) la valeur patrimoniale et les menaces pesant sur les espèces et les forêts de la région méditerranéenne.
Fixation de dunes de sable près de Téhéran, République islamique d'Iran
Peuplement naturel de Quercus suber au Portugal
Richesse de la diversité biologique du maquis naturel à Chypre
RICHESSE EN ESSENCES FORESTIÈRES MÉDITERRANÉENNES: COMPARAISON ENTRE LA CALIFORNIE ET LE BASSIN MÉDITERRANÉEN
Un des caractères majeurs des forêts méditerranéennes, vis-à-vis des forêts européennes, est leur richesse en espèces arborescentes, constitutives ou associées. Si cette particularité a déjà été signalée (Quézel, 1976), aucun bilan précis n'a toutefois été effectué. Sa réalisation pose néanmoins quelques problèmes, car la valeur de diverses nano-phanérophytes, en tant qu'essences forestières, reste discutable. C'est le cas en particulier pour diverses genistées de grande taille (par exemple, Cytisus spp.), certains Tamarix ou Salix, voire les représentants des genres Cotoneaster, Rhus et Withania. Seuls ont été retenus dans cet inventaire les ligneux susceptibles d'atteindre au moins 2 m de haut, et à port arborescent. De même, il n'est pas toujours aisé de décider l'inclusion de certaines espèces, dans les essences préférentielles des forêts de type méditerranéen ou de type européen, d'autant qu'un nombre assez élevé d'arbres existent à la fois dans ces deux types de forêts, sans qu'il soit facile de déterminer leur optimum écologique (par exemple Quercus pubescens, Q. pyrenaica, Q. cerris). Certains taxa ambigus ou de valeur discutée, notamment dans les genres Quercus, Tamarix ou Salix, n'ont pas été considérés, les références restant en priorité Flora Europaea (Tutin et al., 1968-1993) et le Med-Cheklist (Greuter, Burdet et Long, 1984-1989).
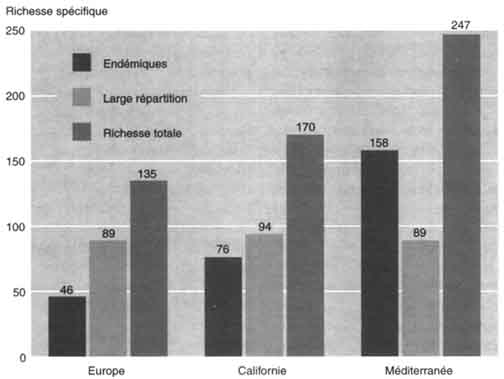
Les forêts de type méditerranéen sont composées par environ le double d'espèces ligneuses par rapport aux forêts de type européen (247 contre 135). Il apparaît que 158 espèces et sous-espèces sont exclusives ou largement préférentielles des forêts méditerranéennes, 46 exclusives ou préférentielles des forêts européennes, et enfin 89 présentes dans ces deux types de forêts. Cette disparité est également perceptible au niveau des genres, puisque 34 sont présents exclusive ment en forêt méditerranéenne, contre sept en Europe. En fait, ce n'est qu'au sein des rosacées, que les forêts européennes offrent un bilan spécifique plus élevé que les forêts méditerranéennes (Pyrus, Malus, Sorbus, Mespilus, Crataegus et Prunus). Les phanérophytes liées aux ripisylves offrent les mêmes particularités, les valeurs affichées étant respectivement de 22 ligneux méditerranéens (dont une quinzaine de Tamarix), sept propres à la région européenne et sept communs aux deux ensembles.
Le bilan des ligneux de Californie (Raven et Axelrod, 1978) a été dressé d'après Hickman (1993), et ce, malgré certaines difficultés liées à la distinction entre arbres et buissons, notamment au sein du genre Arctostaphyllos. Des 170 espèces ligneuses de la région méditerranéenne californienne, 76 sont endémiques. Une comparaison des bilans des deux régions méditerranéennes de l'hémisphère Nord montre que les affinités écologiques, biogénétiques et historiques sont grandes (Mooney et Dunn, 1970; Quézel et Taylor, 1984). En effet, les grands fonds floristiques sont semblables, surtout au niveau des ligneux: sur 58 genres de Californie, 34 sont également présents sur le pourtour méditerranéen. De plus, les perturbations climatiques quaternaires ont entraîné la disparition autour de la Méditerranée de diverses essences encore présentes en Californie telles que Séquoia, Sequoiadendron, Umbellularia, Tsuga, etc. (Roiron, 1992).
Mis à part les différences de superficie entre les zones méditerranéennes en Europe (2 300 000 km2) et en Californie (324 000 km2), il existe aussi une discordance entre les bilans floristiques (respectivement 25 000 contre 4 400 espèces (Quézel, 1985), mais ceux-ci restent très comparables si l'on met en parallèle des superficies analogues (Médail et Quézel, 1997): par exemple, Maroc méditerranéen (300 000 km2 et 4 200 espèces) et Californie méditerranéenne (324 000 km2 et 4 400 espèces). Cependant, la région du pourtour méditerranéen est beaucoup plus riche en éléments arborés, y compris en espèces endémiques, que le reste de l'Europe et la Californie (voir figure), A noter que seuls les genres Dendromecon, Séquoia, Sequoiadendron, et Umbellularia sont endémiques de Californie, contre 14 endémiques ou subendémiques en région circum-méditerranéenne: Argania, Argyrocytisus, Cedrus, Ceratonia, Chamaerops, Fontanesia, Gonocytisus, Hesperolaburnum, Petteria, Podocytisus, Punica, Securinega, Tetraclinis et Warionia.
VALEUR BIOGÉOGRAPHIQUE DES ÉLÉMENTS FORESTIERS ENDÉMIQUES DU BASSIN MÉDITERRANÉEN
L'existence de divers ensembles biogénétiques et biogéographiques majeurs (Quézel, 1985; 1995) constitue un des facteurs essentiels pour expliquer la richesse des essences forestières du pourtour méditerranéen. Un travail a récemment été consacré à cette question (Barbero et al., 1999) et seul un tableau général de la valeur biogéographique des principales essences forestières à vaste répartition (voir tableau p. 25) est fourni ici, tandis que les espèces endémiques sont analysées plus en détail en raison de leur haute valeur biogéographique et des menaces souvent très graves qui pèsent sur plusieurs d'entre elles.
Les espèces endémiques de souche euroasiatique.
Les espèces de souche septentrionale ont donné naissance à quelques espèces endémiques de faible valeur taxinomique notamment: Alnus viridis subsp. suaveolens, en Corse (Gamisans, 1991); Betula aetnensis néoendémique individualisé en Sicile à partir de B. pendula; B. celtiberica en Espagne centro-occidentale et B. fontqueri en Andalousie et sur le Rif, différenciés respectivement à partir de B. pubescens et de B. pendula. On trouve Quercus petraea subsp. huguetiana en Espagne septentrionale, alors que l'on trouve divers autres chênes caducifoliés, à statut taxinomique souvent imprécis, en Sicile, en Italie méridionale, dans la région égéenne et au Proche-Orient. En outre, bon nombre de ligneux associés, individualisés sur l'ensemble du pourtour méditerranéen, se rencontrent en particulier au sein des genres Acer, Cotoneaster, Prunus, Pyrus, Salix et Sorbus.
Les éléments méditerranéens endémique à aire restreinte.
Les espèces endémiques de souche méditerranéenne ou mésoégéenne à aire restreinte, sont assez nombreux et riches de signification biogéographique (Barbero, Loisel et Quézel, 1995). Le cas le plus remarquable est constitué par les sapins (Quézel, 1998) avec plusieurs espèces affines, présentes du détroit de Gibraltar jusqu'au Liban (Abies pinsapo, A. marocana incl. A. tazaotana, A. numidica, A. cephalonica incl., A. borisii-regis et A. cilicica), mais aussi en Anatolie septentrionale pour les sapins pontiques. Chez les autres conifères, on peut citer Cupressus atlantica, résiduel sur le Haut Atlas occidental, et Pinus nigra subsp. dalmatica. Parmi les feuillus, il faut signaler Quercus afares du Maghreb nord-oriental, Q. euboica en Eubée (Grèce), Q. vulcanica en Anatolie centrale (Turquie) et Arbutus pavarii de Cyrénaïque (Jamahirya arabe libyenne). Liquidambar orientalis, remarquable relique préglaciaire, est localisé en Turquie du Sud-Ouest (Akman et al., 1993) et dans les îles du Dodécanèse, tout comme Quercus aucheri, vicariant du chêne vert.
Quelques éléments forestiers endémiques de souche africaine jouent un rôle marginal (Quézel, 1995), sauf dans le sud-ouest marocain où Argania spinosa et Acacia gummifera occupent une place appréciable, associés localement à Olea marocana et à Dracaena draco subsp. ajgal dans les montagnes de l'Anti-Atlas occidental (Benabid et Cuzin, 1997). Les autres espèces endémiques de souche méridionale sont peu nombreuses: Phoenix theophrastii, voisin de P. dactylifera, présent en Crète et en Anatolie sud-occidentale (Yaltirik et Boydak, 1991); Securinega tinctoria se rencontre dans le sud-ouest de la péninsule ibérique, tandis que Argyrocytisus, Hesperolaburnum et Warionia se cantonnent au Maroc. Enfin, le genre Tamarix individualise de nombreuses espèces à répartition restreinte en situation halohygrophile.
Les îles abritent bon nombre d'espèces ligneuses endémiques, notamment en Sicile: Abies nebrodensis (Morandini, Ducci et Menguzzato, 1994) et Rhamnus lojaconoi, localisés sur les monts de la Madonie; Celtis aetnensis très proche de C. tournefortii, sur l'Etna; et Zelkova sicula, espèce endémique relictuelle du Monte Lauro (Di Pascale, Garfi et Quézel, 1992). En Crète, Zelkova abelicea est encore présent sur les trois principaux massifs montagneux, en populations exiguës. Sur les roches vertes de Chypre, Cedrus brevifolia, végète à l'étage mésoméditerranéen (entre 900 et 1 200 m), et Quercus alnifolia structure des peuplements entre 1 000 et 1 900 m.
INSTABILITÉ GÉNÉTIQUE DES ESSENCES CONSTITUTIVES DES FORÊTS MÉDITERRANÉENNES
De nombreuses essences forestières méditerranéennes se caractérisent par une instabilité génétique qui se traduit en général par des incertitudes taxinomiques difficiles à élucider. C'est ainsi que, chez les chênes sclérophylles, le statut précis des complexes Quercus ilex (y compris Q. rotundifolia) ou Q. coccifera, (y compris Q. calliprinos) n'est pas réglé, malgré de nombreuses études biochimiques ou génétiques (Michaud, Lumaret et Romane, 1992). Il en est de même au sein des genres Juniperus, Pinus (Barbero et al., 1998). et Abies (Bergmann et Gregorius. 1992). pour ne citer que les gymnospermes où cette approche est la plus avancée. Cette instabilité est aussi responsable des facilités souvent très grandes qu'offrent beaucoup d'espèces forestières méditerranéennes à l'hybridation ou à l'introgression, en particulier au sein des genres Quercus, Pinus et Abies. Par exemple, il existe d'innombrables hybrides naturels de Quercus tels que Q. streimii (Q, pubescens x Q. sessiliflora) sur le pourtour méditerranéen français.
L'hybridation entre Pinus brutia et P. halepensis a été évoquée par ailleurs, comme celle entre P. sylvestris et P. uncinata (Barbero et al., 1998). Pour les espèces ou les genres à aire disjointe, le problème est encore plus grave et les reboisements en sapins méditerranéens entraînent des phénomènes massifs de pollution génétique avec les types autochtones (Quézel, 1998). Ces phénomènes apparaissent d'ailleurs dans beaucoup d'autres genres (Acer, Fagus, Prunus, Sorbus, etc.), et force est d'en tenir compte lors de l'introduction d'espèces allochtones voisines sur le plan taxinomique, et ce, afin d'éviter d'éventuelles pertes dramatiques du capital biologique - c'est par exemple le cas des dernières populations de pin noir cévenol (Pinus nigra subsp. salzmanii), profondément hybridées à la suite de l'introduction massive des autres types de pin noir.
BIODIVERSITÉ ET VALEUR PATRIMONIALE DES COMMUNAUTÉS FORESTIÈRES MÉDITERRANÉENNES
Les résultats les plus significatifs d'une étude récente (Barbero et al., 1999) - utilisant une approche globale quant à la biodiversité et valeur patrimoniales des communautés forestières méditerranéennes - sont résumés ci-dessous.
Les forêts méditerranéennes, en particulier les formations caducifoliées supraméditerranéennes d'Afrique du Nord (Numidie), de Corse et de Grèce, présentent une importante richesse floristique totale (espèces accidentelles exclues). De nombreuses forêts de conifères montagnards-méditerranéens, et notamment celles de Cedrus spp. et de Pinus nigra comptent plus de 100 espèces. Les seules formations mésoméditerranéennes sclérophylles atteignant ces valeurs sont les structures à Quercus calliprinos du Proche-Orient. Inversement, les niveaux les plus faibles se rencontrent en particulier au sein des communautés forestières des grandes îles méditerranéennes - forêts à Acer sempervirens et Cupressus sempervirens de Crète (25 taxons), et Quercus infectoria subsp. veneris de Chypre (27 taxons) - et en bioclimat aride, par exemple Acacia gummifera du sud-ouest marocain (25 taxons). La richesse en phanérophytes peut être également élevée, notamment au niveau de certaines formations caducifoliées: châtaigneraies corses (29 taxons), chênaies caducifoliées ligures de Fraxinus ornus (29 taxons) et les forêts de Quercus frainetto en Grèce (26 taxons), mais aussi les forêts de cèdres et sapins du Proche-Orient (24 à 5 taxa).
TABLEAU 1. Valeur biogéographique des essences forestières méditerranéennes et des principales phanérophytes associées1
|
ÉLÉMENTS FORESTIERS DE SOUCHE EUROPÉENNE Largements répandus (majorité des espèces): Marges septentrionales ou épars: Macaronésiens: ÉLÉMENTS FORESTIERS DE SOUCHE MÉDITERRANÉENNE Circum-méditerranéens: Arbutus unedo, Celtis australis, Ceratonia siliqua, Cercis siliquastrum, Erica arborea, Euphorbia dendroides, Juniperus phoenicea s.l., J. oxycedrus s.l., Laurus nobilis, Myrtus communis, Nerium, Olea europaea, Phillyrea spp., Pinus pinea, Pistacia lentiscus, P. terebinthus, Punica, Quercus coccifera, Rhamnus alaternus, Rhus oxyacantha, Tamarix spp., Viburnum tinus, Vitex O. Méditerranéens stricts: Chamaerops, Juniperus thurifera s.l., Pinus uncinata, Quercus suber O. méditerranéens préférentiels: Pinus halepensis, Pinus pinaster, Quercus ilex s.l., Q. pubescens s.l. Ibéro-maurétaniens préférentiels: Buxus balearica, Lonicera arborea, Osyris lanceolata, Quercus canariensis, Q. faginea s.l., Q. lusitanica, Q. pyrenaica Maurétaniens: Cedrus atlantica, Fraxinus dimorpha, Pinus nigra subsp. mauretanica, Tetraclinis Ibériques: Pinus nigra subsp. salzmannii Tyrrhéniens: Alnus cordata, Pinus nigra subsp. laricio Sud-Méditerranéens: Acacia raddiana, Pistacia atlantica, Rhus pentaphylla, Ziziphus lotus E. méditerranéens stricts: Arbutus andrachne, Alnus orientalis, Celtis tournefortii, Cupressus sempervirens, Fontanesia, Gonocytisus, Pinus brutia, Platanus, Podocytisus, Quercus infectoria, Q. macrolepis s.l., Q. trojana E. médit. au sens large: Carpinus orientalis. Fraxinus ornus, Ostrya, Pinus nigra subsp. pallasiana, Quercus calliprinos, Q. cerris s.l., Q. frainetto, Styrax Balkaniques: Aesculus, Petteria Italo-Balkaniques: Pinus hetdreichii, P. mugo Gréco-Anatoliens: Acer sempervirens, Juniperus drupacea, Quercus brachyphylla Proche-Orientaux: Acer obtusifolium, Cedrus libani, Juniperus excelsa, J. foetidissima, Lonicera nummulariifolia, Pistacia palaestina, Quercus pubescens subsp. anatolica, Q. vulcanica. |
1
Endémiques exclues; les genres seuls ont été indiqués lorsqu'ils sont monospécifiques en région méditerranéenne.
Les forêts méditerranéennes ne sont généralement pas très riches en espèces endémiques dont le pourcentage ne représente qu'entre 5 et 10 pour cent du nombre total d'espèces. Toutefois, certaines régions ont une vingtaine d'espèces forestières endémiques, et dans les peuplements culminaux de Juniperus foetidissima sur roches vertes du Troghodhos à Chypre, 20 des 31 espèces observées sont endémiques.
ESPÈCES ET COMMUNAUTÉS VÉGÉTALES RARES OU MENACÉES EN RÉGION MÉDITERRANÉENNE
Dans les pays de la partie septentrionale du bassin méditerranéen, l'effondrement du système agrosylvopastoral multiséculaire entraîne de profondes modifications dans la structure et l'architecture des phytocènoses forestières et préforestières, et, d'une manière générale, une maturation des peuplements forestiers (Barbero et al., 1990). En Afrique du Nord, les bouleversements écologiques induits par la déforestation et le surpâturage engendrent des dysfonctionnements graves, et les forêts en haute montagne sont particulièrement touchées. En dépit du fait que les processus écologiques et socioéconomiques varient d'une région à l'autre, les conséquences écologiques qui en résultent sont très similaires: i) dérèglements des cycles naturels de perturbations et déséquilibres écologiques engendrant des catastrophes climatiques se déroulant sur de vastes échelles; ii) homogénéisation des structures et architectures des communautés végétales; iii) perte nette de diversité biologique et banalisation de la flore; et iv) extension des xénophytes à travers l'«artificialisation» des milieux qui font concurrence aux végétaux indigènes en sous-bois.
Divers travaux récents ont cherché à évaluer les risques encourus par l'ensemble de la flore méditerranéenne (Gomez-Campo, 1985; Médail et Quézel, 1997), et l'Alliance mondiale pour la Nature (UICN) a publié un bilan général relatif à la flore circum-méditerranéenne indiquant que 53 pour cent des espèces endémiques sont menacées, soit 1 529 taxons (en excluant la République arabe syrienne, le Liban et la Turquie). Cependant, les données sur les végétaux forestiers menacés en région méditerranéenne sont encore très fragmentaires. Bon nombre d'arbres d'Afrique du Nord (Olea marocana, Pyrus div. spp., Pinus nigra subsp. mauretanica, etc.) ne figurent pas dans la liste des espèces ligneuses menacées sur l'ensemble du globe qui fait partie d'une étude récente effectuée par l'UICN (Oldfield, Lusty et McKinven, 1998), et ils ne figurent pas non plus dans le 1997 IUCN Red list of threatened plants (Walter et Gillett, 1998); ces deux études ne font état que de 45 espèces ligneuses menacées dans le bassin méditerranéen, alors qu'en fait, on en a répertorié 71. Le nombre d'espèces endémiques figurant sur cette liste (48) - nombre d'entre elles étant des espèces rares progénitrices de végétaux cultivés ou qui représentent un pool génique intéressant (Malus, Olea, Phoenix, Prunus et Pyrus) - souligne tout l'enjeu de la conservation de ces arbres.
Les communautés forestières menacées du bassin méditerranéenne n'ont pas fait l'objet, à ce jour, d'une enquête à grande échelle, seuls quelques jalons ayant été posés dans cette direction (Quézel et Barbero, 1990; Quézel, 1991). Les peuplements forestiers peuvent être menacés à cause de: i) leurs exigences écologiques particulières, notamment la nature du substrat, qui en font des systèmes naturellement exigus (cas de certains groupements gypsicoles ou serpentinicoles); ii) leur situation marginale sur le plan chorologique (limites d'aire, isolats), qui s'explique souvent par des compensations bioclimatiques (communautés abyssales) ou des raisons historiques (refuges pléistocènes), comme dans le cas du peuplement de Dracaena drago du sud-ouest marocain (Médail et Quézel, 1999); mais surtout iii) des impacts anthropozoogènes intenses et généralisés que subissent ces communautés, à l'orgine largement répandues.
Dans le nord de la Méditerranée, les peuplements relictuels de Abies nebrodensis et Zelkova sicula (entre 200 et 250 individus) de Sicile sont les plus sérieusement menacés, tandis que Abies pinsapo, Quercus petraea subsp. huguetiana d'Espagne et Pinus nigra subsp. dalmatica de l'ex-Yougoslavie, bien que localisés, semblent un peu moins menacés. Sur le littoral méditerranéen français, les ensembles thermoméditerranéens préforestiers (formations à Ceratonia siliqua et Chamaerops humilis, à Olea et Euphorbia dendroides) et forestiers (chênaies vertes matures à Laurus nobilis ou Cyclamen balearicum et chênaie pubescente à Cyclamen repandum), régressent en raison de l'urbanisation ou de la fragmentation, responsable d'extinctions locales d'espèces ou de l'isolement des populations.
Au sud de la Méditerranée, la plupart des forêts sont menacées, à des degrés divers; au Maghreb (Quézel et Barbero, 1990; Quézel, 1991), la situation est particulièrement préoccupante pour les sapinières à Abies maroccana et A. numidica, les peuplements de Pinus nigra subsp. mauretanica, Cupressus atlantica, Betula pendula subsp. fontqueri, Olea marocana, Laurus azorica, Quercus afares, Q. faginea subsp. tlemcenensis, et même pour certaines formations à Argania spinosa, Cedrus atlantica, Tetraclinis articulata et Juniperus thurifera.
En Méditerranée orientale, la gestion forestière plus stricte (surtout en Turquie) et la participation des populations locales aux bénéfices d'exploitation se sont traduites par un meilleur état de conservation des forêts (Quézel et Barbero, 1990); cependant, certaines communautés méritent un contrôle continu: Abies nordmanniana subsp. equi-trojani, Quercus aucheri, Liquidambar orientalis et Fagus orientalis en Turquie, formations à Cedrus brevifolia et Quercus alnifolia à Chypre, peuplements diffus de Zelkova abelicea et Phoenix theophrasti en Crète, peuplements à Abies borisii-regis, Aesculus hippocastanum et Pinus heldreichii.
Peuplement protégé de Pinus pinea dans le Parc National d'Arabida, au Portugal
CONSERVATION ET GESTION PATRIMONIALE
La conservation des forêts et des végétaux forestiers du bassin méditerranéen constitue un problème complexe du fait de l'hétérogénéité des situations écologiques rencontrées et des multiples usages et pressions anthropiques pratiqués par les diverses entités culturelles. La gestion des divers stades dynamiques des systèmes forestiers nécessite de considérer non seulement les communautés et espèces strictement forestières, mais aussi celles associées aux lisières et clairières. La répartition assez vaste de la plupart des essences forestières conduit à penser que ces végétaux ne subissent que des menaces potentielles réduites, mais il faut tout de même tenir compte de la marginalité chorologique (espèces en limite d'aire de distribution ou en isolats) et/ou écologique (espèces liées à des biotopes rares) de certaines populations (Rameau et Olivier, 1991). De plus, un bon nombre de ligneux représente des espèces clés dont le maintien est essentiel pour la pérennisation des cycles biogéochimiques.
Les multiples régimes de perturbation ont profondément façonné la structure et les successions écologiques des forêts méditerranéennes, et il importe de garantir la persistance, en particulier dans les zones protégées, de telles séquences de perturbation afin d'assurer le maintien d'une mosaïque de biocènoses et d'une biodiversité optimale. Beaucoup d'espèces ligneuses peuvent subsister en populations isolées sur des territoires très restreints pendant plusieurs décennies, voire plus d'une centaine d'années, tant que les principales conditions biotiques persistent. Toutefois, les populations géographiquement isolées et réduites sont plus que toutes autres vulnérables et soumises à l'impact de la stochasticité environnementale. Ainsi, Frankel, Brown et Burdon (1995) avancent que le nombre minimal d'arbres nécessaires pour le maintien d'une population forestière viable sur le long terme est d'environ 500. Par contre, la variabilité génétique semble moins liée à la taille des populations de ligneux. Ainsi, une population extrêmement réduite comme celle de l'Abies nebrodensis (29 adultes et 20 semis selon le récent dénombrement de Morandini, Ducci et Menguzzato, 1994) possède une variabilité génétique semblable à celles analysées sur des populations dynamiques d'Abies alba de Calabre, en situation écologique similaire (Vicario et al., 1995). En fait, la grande précarité de la population relictuelle d'Abies nebrodensis est due au niveau peu élevé de production de semences (lesquelles ont de plus une viabilité très réduite) et à la nature du biotope, défavorable à la régénération du sapin des Nébrodes (Ducci, Proietti et Favre, 1998).
Le problème de la conservation in situ des ressources génétiques des essences forestières méditerranéennes parle biais des réserves naturelles existantes réside dans le fait que ces dernières sont très rarement créées dans cette optique. La désignation et la délimitation de structures foncières de conservation reposent encore le plus souvent sur des opportunités locales ou sur des critères esthétiques subjectif s, plutôt que sur des analyses rigoureuses multiniveaux de la biodiversité d'un territoire donné. Même si un archipel de petites réserves peut contenir plus d'espèces, cette approche n'est pas idéale pour ce qui est de la conservation durable des populations de végétaux forestiers. En effet, certains taxons comme les espèces forestières internes se maintiendront uniquement au sein de zones suffisamment vastes pour que leur cycle de vie se déroule de façon optimale, sans être altéré par les effets de lisière. Seules de vastes réserves peuvent garantir le déroulement des cycles normaux de perturbations, et assurer le maintien des populations de grands vertébrés garants de le persistance des hétérogénéités paysagères et de la dispersion des végétaux. Ainsi, il paraît préférable de mettre en place des systèmes intégrant de vastes réserves intégrales (au moins quelques centaines d'hectares) entourées de zones tampons, en mosaïque avec des parcelles gérées de façon différente (Frankel, Brown et Burdon, 1995). De telles structures sont théoriquement mises en place dans les «Réserves de la biosphère» du Programme l'homme et la biosphère de l'UNESCO.
Une stratégie intégrée de conservation des espèces forestières menacées ou de celles étant d'un intérêt patrimonial nécessite donc avant tout une bonne connaissance de l'écologie et de la biologie des espèces - ce qui est rarement le cas. Une large place doit être accordée aux espèces endémiques, en raison de leur unicité et de leur répartition restreinte, et aux populations marginales. Toutefois, le maintien des capacités adaptatives des espèces par la pression de sélection implique obligatoirement une conservation in situ qui passe par une gestion dynamique des paysages forestiers.
Bibliographie
Akman, Y., Quézel, P., Ketenoglu, O. et Kurt, I. 1993. Analyse syntaxonomique des forêts de Liquidambar orientalis en Turquie. Ecol. medit., 19: 49-57.
Barbero, M., Bonin, G., Loisel, R. et Quézel, P. 1990. Changes and disturbances of forest ecosystems caused by human activities in the western part of the Mediterranean Basin. Vegetatio, 87: 151-173.
Barbero, M., Loisel, R. et Quézel, P. 1995. Les essences arborées des îles méditerranéennes: leur rôle écologique et paysager. Ecol. medit., 20(1/2): 53-69.
Barbero, M., Loisel, R., Médail, F. et Quézel, P. 1999. Signification biogéographique et biodiversité des forêts du bassin méditerranéen. Bocconea. (sous presse).
Barbero, M., Loisel, R., Quézel, P., Richardson D.M. et Romane, F. 1998. Pines of the Mediterranean Basin. In D.M Richardson (ed.), Ecology and biogeography of Pinus. p. 153-170. Cambridge University Press, Cambridge, Royaume-Uni.
Benabid, A. et Cuzin, F. 1997. Populations de dragonnier (Dracaena draco L. subsp. ajgal Benabid et Cuzin) au Maroc: valeurs taxinomique, biogéographique et phyto-sociologique. C. R. Acad. Sci. Paris, Sciences de la vie/Life Sciences, 320: 267-277.
Bergmann, F. et Gregorius, H.R. 1992 Ecogeographical distribution and thermostability of isocitrate deshydrogenase (IDH) alloenzymes in European silver fir (Abies alba). Biochem. Syst. Ecol., 21:597-605.
Di Pascale, G., Garfi, G. et Quézel, P. 1992. Sur la présence d'un Zelkova nouveau en Sicile sud-orientale (Ulmaceae). Biocosme Mésogéen, 8(4)-9, (1): 401-409.
Ducci, F., Proietti, R. et Favre, J.M. 1998. Le genre Abies en Italie: écologie générale, gestion sylvicole et ressources génétiques. Forêt médit., 19(2): 153-164.
Frankel, O.H., Brown, A.H.D. et Burdon, J.J. 1995. The conservation of plant biodiversity. Cambridge University Press, Cambridge, Royaume-Uni. 299 p.
Gamisans, J. 1991. La végétation de la Corse. In D. Jeanmonod. et H.M. Burdet (éds.), Compléments au prodrome de la flore corse, annexe 2. Conserv. et Jard. bot. Genève, Genève. 391 p.
Gomez-Campo, C., éd. 1985. Plant conservation in the Mediterranean area.
Greuter, W., Burdet H.M. et Long. G. 1984-1989. Med-Checklist. 3 vol. Conserv. et Jard. bot. Genève, Genève.
Hickman. J.C., éd. 1993. The Jepson Manual. Higher plants of California. University of California Press, Berkeley et Los Angeles, Etats-Unis.
Médail, F. et Quézel, P. 1997. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean basin. Ann. Missouri Bot. Gard., 84: 112-127.
Médail, F. et Quézel, P. 1999. The phytogeographical significance of S.W. Morocco compared to the Canary Islands. Plant Ecology. (sous presse)
Michaud, H., Lumaret, R. et Romane, F. 1992. Variation in the genetic structure and reproductive biology of holm oak populations. Vegetatio, 99/100: 107-113.
Mooney, H.A. et Dunn, E.L. 1970. Convergent evolution of mediterranean-climate evergreen sclerophyll shrubs. Evolution, 24: 292-303.
Morandini, R., Ducci, F. et Menguzzato, G. 1994. Abies nebrodensis (Lojac.) Mattei, inventario 1992. Ann. Ist. Sperim. Selvic. Arezzo, 22: 5-51.
Olfield, S., Lusty, C. et McKinven, A. 1998. The World list of threatened trees. IUCN Press. Cambridge, Royaume-Uni. 650 p.
Pons, A. et Quézel, P. 1985. The history of the flora and vegetation and past and present human disturbance in the mediterranean région. In: C. Gomez-Campo (éd.). Plant conservation in the Mediterranean area. Geobotany 7, p. 25-43. W. Junk, Dordrecht, Pays-Bas.
Quézel, P. 1976. Les forêts du pourtour méditerranéen. In Forêts et maquis méditerranéens: écologie, conservation et aménagements. Note technique MAB, 2: 9-33. UNESCO, Paris.
Quézel, P. 1985. Definition of the Mediterranean region and the origin of its flora. In C. Gomez-Campo (éd.). Plant conservation in the Mediterranean area. Geobotany 7, p. 9-24. W. Junk, Dordrecht, Pays-Bas.
Quézel, P. 1991. Structures de végétation et flore en Afrique du Nord: leurs incidences sur les problèmes de conservation. In M. Rejdali et V.H. Heywood (éds), Conservation des ressources végétales, p. 19-32. Actes Editions, Institut agronomique et vétérinaire Hassan II, Rabat.
Quézel, P. 1995. La flore du bassin méditerranéen: origine, mise en place, endémisme. Ecol. medit., 20 (1/2): 19-39.
Quézel, P. 1998. Diversité et répartition des sapins sur le pourtour méditerranéen. Forêt médit., 19(2): 93-104.
Quézel, P. et Barbero, M. 1990. Les forêts méditerranéennes. Problèmes posés par leur signification historique, écologique et leur conservation. Acta Bot. Malacitana, 15: 145-178.
Quézel, P. et Taylor, H.C. 1984. Les fruticées sempervirentes des régions méditerranéennes de l'Ancien Monde. Essai comparé d'interprétations des structures biologiques et des données historiques. Bot. Helv., 94: 133-144.
Rameau, J.C. et Olivier, L. 1991. La biodiversité forestière et sa préservation. Intérêt patrimonial de la flore, de la végétation et des paysages forestiers. Rev. For. Fr., 43, num. sp. 1991: 19-27.
Raven, P.H. et Axelrod, D.I. 1978. Origin and relationships of the California flora. Univ. California Public. Bot., 72: 1-134.
Roiron, P. 1992. Flores, végétations et climats du Néogène méditerranéen: apports de macroflores du sud de la France et du nord-est de l'Espagne. Univ. Montpellier II. 296 p. (thèse)
Thirgood, J.V. 1981. Man and the Mediterranean forest. Academic Press, New-York. 194 p.
Tutin, T.G. et al. 1968-1993. Flora Europaea. Vol. 1-5. Cambridge University Press, Cambridge, Royaume-Uni.
Verlaque, R., Médail, F., Quézel, P. et Babinot, J.F. 1997. Endémisme végétal et paléogéographie dans le bassin méditerranéen. Geobios, Mém. sp. 21: 159-166.
Vicario, F., Vendramin, G.G., Rossi, P., Lio, P. et Giannini. R. 1995. Allozyme, chloroplast DNA and RAPD markers for determining genetic relationships between Abies alba and the relict population of A. nebrodensis. Theor. Appl. Genet., 90: 1012-1018.
Walter, K.S., et Gillett, H.J., éds. 1998. 1997 IUCN Red List of threatened plants. Compiled by the World Conservation Monitoring Centre. UICN-Alliance mondiale pour la nature, Gland, Suisse et Cambridge, Royaume-Uni.
Yaltirik F. et Boydak, M. 1991. Distribution and ecology of the palm Phoenix theophrasti (Palmae) in Turkey. Bot. Chron., 10: 869-872.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}