![]()
![]()
![]()
The domestic goat, Capra hircus, is believed to be the first ruminant ever domesticated. Evidence suggests that this took place before 7000 BC in south-west Asia, on the borders of present-day Iran and Iraq, where agriculture was already advanced (Mason 1984). It is believed that Capra hircus is derived from the wild goat, Capra aegagrus, whose horn shape closely resembles the horns of the domestic goats we know today.
Ethiopia has long been recognised as a source of genetic diversity in plants and animals. It appears that Ethiopia has served as a gateway of genetic material from Asia to Africa, and that its diverse ecology served to further diversify and develop the genotypes it received. It is believed that the first goats reached Egypt by 5000 BC and spread south and west thereafter. Remains of a small goat have been found in a site in Sudan thought to date from before 3300 BC and the lop-eared Nubian type has been found in sites dating from 2500 BC (Mason 1984). It is therefore likely that the first wave of goats entered Ethiopia from the north sometime between 2000 and 3000 BC.
Immediately after domestication physical differentiation of goats into types must have begun. Early physical changes affected the ears, horns, colour and hair type. These changes must have arisen from natural mutation and selection by their keepers within the environment in which they were reared. Most type development must have taken place in relative geographical isolation. New blood may have entered populations when man moved during trade, wars and ethnic movements. Looking at the breeds that now exist we can see that there are locations containing populations of goats of relative uniformity and these might be termed 'breeds'. However, the mixing of stock around the boundaries of these areas, through trade or movement, has led to many intermediate, less distinguishable goats which might be referred to as 'types' or 'subtypes'.
In the past the influence of man on livestock populations has, perhaps, been underestimated. However, it is quite clear from the results of this study that man has, in many cases, exerted considerable influence on the populations studied. The Somali goat has clearly been selected for milk production, against twinning (to the point where it is virtually eliminated), and for production and survival in harsh environments. Its uniformly white coat appears also to have been selected for. The prolificacy of the Western Lowland, Western Highland, Keffa and Woyto-Guji goats has clearly been selected for by societies for whom milk is not so important. In addition, the multiple social and economic functions reported during the study demonstrate the social value placed on their flocks by the societies keeping them.
Although Ethiopia's and Eritrea's rugged highland terrains have served to keep their peoples in relative isolation there is a long history of trade with Arab countries across the Red Sea, with Sudan in the west and through pastoral groups on the southern borders. It could be imagined that waves of trading, together with physical movements of people, e.g. the Oromo migrations, must have influenced the genetic make-up of domestic livestock, including goats (Hultin 1987; Mohammed Hassen 1990).
The identification, description, classification and naming of livestock types is the first step in the assessment of a country's livestock resources. The physical description of types can only dimly reveal the genetic relationships between individuals, but is a first step in classifying a diverse population into relatively homogeneous sub-populations. New techniques of genotyping are now better able to reveal the genetic distance between populations and this allows a better understanding of how these populations evolved. However, for the practical exploitation of livestock resources the productive and adaptive performance of identified populations in specified environments is required. It is only in this way that variations in their performance due to genetic effects (additive and non-additive) and non-genetic effects can be elucidated (Fitzhugh 1987). The performance assessment of domestic livestock species must be considered a prerequisite for the sound planning of livestock development in any country.
In addition, the classification and characterisation of livestock types is necessary before homogenous genetic material can be preserved for future generations. Much genetic variability, the raw material of breed improvement, has been lost in developed countries. There is no doubt that preserving genetic diversity, for either immediate or future use, is important.
Goats in Africa have traditionally been divided into three main families - the Dwarf goats of West and central Africa, the Savannah goats of sub-Saharan Africa and the Nubian type goat of northern Africa (Epstein 1971; Wilson 1991). However, the principal components analysis clearly differentiated four major families of goats in Ethiopia - the Somali family (Short-eared Somali, Long-eared Somali and Hararghe Highland), the Nubian family (Nubian and Barka), the smaller Rift Valley family (Abergelle, Worre, Afar, Arsi-Bale and Woyto-Guji) and finally the more heterogeneous Small East African family (West Highland, Keffa, Central Highland and West Lowland).
Lauvergne et al (1993) have developed the concept of an 'index of primarily' in goat populations, reflecting the distance between a population and that originally domesticated. After domestication, mutations arise which are no longer necessarily removed by natural selection. Measuring the incidence of these mutations can indicate the degree of primarily of the population. An index of 0 describes a standardised breed with little variability, while an index of 1 describes an original population close to the original domesticated. In the goat populations they examined in Chad and northern Cameroon, indices of 0.75-0.88 were recorded, indicating that the populations were close to those originally domesticated.
However, the heterogeneity of a population does not just reflect its degree of primitiveness. It must also, to some extent, be a result of mixing and interbreeding of populations combined, possibly, with a relatively low level of selection by owners.
It is hard to definitively identify the order in which the four 'families' entered Ethiopia. The Nubian family is clearly closely related to the goats found in Egypt, Sudan, Syria, Iraq and Iran today. The Rift Valley type is believed to have entered Ethiopia from across the Red Sea and has extended its range along the Rift Valley. The Somali family has been subject to strong selection pressure for some clearly defined characteristics, while the Small East African has been kept by diverse ethnic groups for diverse reasons.
It is evident from the data that goats maintained by pastoralists in relative isolation have more homogenous, clearly defined characteristics than the more heterogenous goat populations kept in the highlands. This heterogeneity may be as a result of human movements mixing previously more homogenous types, combined with the weaker selection pressure that can be exerted in small flocks. The degree of movement thought to have taken place in the 16th and 17th centuries alone is indicated in Figure 6. Ethnic groups must have moved with their livestock which subsequently interbred with stock in the new location. Much of this movement involved the settlement of relatively more hospitable highland areas by groups used to a more pastoral way of life. This inevitably led to a mixing and interbreeding of goats creating the heterogenous highland goat types we observe today.
Figure 6. The great ethnic migrations C 1520-1660.
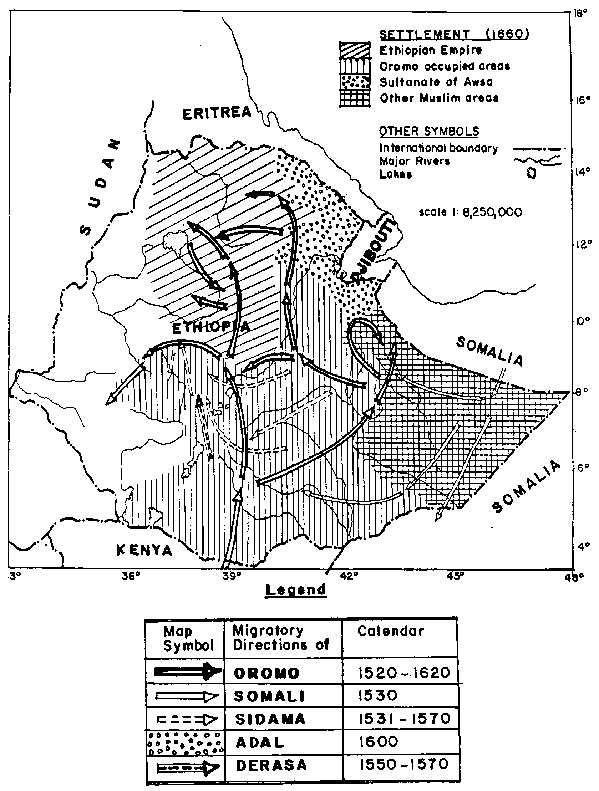
Sources: Adapted from Fage and Verity (1978); Ajayi and Crowder (1985); EMA (1988).
The survey carried out should be the very first step in breed characterisation through differentiating a heterogenous population into relatively homogenous sub-populations which may then be named as breeds, types or subtypes. However, it would be ideal if the relationships of the types identified phenotypically could be confirmed through analysis of their DNA. Kemp (1992) has described the range of techniques currently available to determine the genotype of individual animals. The polymerase chain reaction (PCR) allows the amplification of specific DNA sequences. This can be generated from small amounts of DNA extracted from hair or blood samples. From this the genetic distance between individuals can be determined, or potentially, specific areas of the DNA can be used to identify DNA sequences containing specific traits. If the DNA analysis validates the effectiveness of the field and data analysis method used in this study to identify relatively more homogenous populations, the field method could have much wider applications.
The survey has also clearly shown how widely goats are milked in Ethiopia and Eritrea. Goats are milked in both the lowlands and highlands of both countries with the exception of parts of Gojam, Wellega and Keffa. In all other parts of the country goats are milked as much as the limited milk yields allow. In some parts of the country, most notably Wolayta, milking goats is considered to be a sign of poverty.
Suggestions for future research and development
The limited quantitative and qualitative measurements made during this survey have allowed the preliminary description of the phenotypic characteristics of the goat types of Ethiopia. Once more homogenous populations have been defined what is the next step?
Baker (1992) suggests that once breeds are known there are three levels of research studies required for genetic characterisation of these known breeds:
· Macro-level studies: on-farm surveys to describe the population structure and make a physical description of the breed.· Meta-level studies: for phenotypic characterisation of breeds a more detailed on-farm monitoring is needed. This should be designed to allow comparisons of the breed's mean and variance for physical traits, performance traits and health traits.
· Micro-level studies: more detailed on-station characterisation studies with sufficient dams and sires to allow the estimation of genetic parameters, e.g. heritabilities, and between breed comparisons under the same environment. Genetic characterisation at the molecular level is the surest way to define 'breed' boundaries and to quantify available genetic diversity. Investigation of protein polymorphisms may be used as a step before this.
These studies can assist in developing rational, between-breed, and within-breed, improvement strategies. The most promising goat types could be evaluated 'on station' to try to establish their production potential. On the basis of information obtained from such studies selection programmes could be designed to exploit valuable traits.
In terms of the assessment of these types for the purposes of practical improvement the next step should be the performance testing of the most promising types. The major goat types and their main traits of interest are listed in Table 33.
Table 33. Goat types and some of their promising traits.
|
Goat type |
Trait |
|
Nubian |
Milk |
|
Barka |
Milk |
|
Somali (Short- and Long-eared) |
Milk and survival |
|
Abergelle |
Growth |
|
Western Lowland |
Prolificacy |
|
Keffa |
Prolificacy |
|
Central Highland |
Skin quality |
Suggestions for modifications to field and data analysis methods
The main modifications to the field method suggested are:
· If on-farm phenotypic characterisation is the main purpose of the field survey measurements from adult females only need to be taken. At the same time hair or blood samples can be taken for DNA analysis.· It is hard to make meaningful conclusions from one milk measurement because of the confounding effects of stage of lactation, season, management etc. If possible, enumerators should be employed to record the milk yields of as large a number of lactating females as possible for 3-4 days at each site. Information on parity or age, together with stage of lactation should also be collected wherever possible.
· General management information can most quickly be obtained from a group discussion.
For further data analysis of the existing data set discriminant analysis can be used to create a field identification key. Each cluster should be assumed to group sites where the goat population is considered homogeneous. The interpretation of the knots of the hierarchical tree is indicative of the discriminating factors between clusters. These can be used to produce an aid to the field identification of breed types. For each knot in the hierarchical tree, a stepwise discriminant function (proc MAHAL2 of ADSAS) can be computed (ADDAD 1990). The most discriminating variables at each knot provide indications of the logical path if the intention is to relate an unknown Ethiopian goat population to one of the types described.
The phenotypic characterisation of the goats of Ethiopia and Eritrea described in this report is the first attempt at a systematic description of the goat resources of a nation. It has clearly shown that there are distinct and diverse goat types maintained by their owners in a wide range of environments. It is clear that the more homogenous types are found in the lowlands of the two countries while highland goats show more heterogenous characteristics. There are clearly traits of economic importance the development of which should be considered. There are no populations under threat of extinction but some genetic erosion occurs at the edges of some lowland populations. However, as yet this poses no threat to the continued existence of the types of concern. This unique study has laid the foundation for the future exploitation of the goat breeds described for the benefit of some of the world's most needy livestock keepers.
![]()
![]()
![]()