森林、生物多样性与人类

下载本出版物
EPUB
MOBI
PDF
森林、生物多样性与人类
本旗舰出版物系联合国粮食及农业组织“世界之状况”系列之一。
引用格式要求:
粮农组织和环境署。2020。《2020年世界森林状况:森林、生物多样性与人类》。罗马。
DOI: https://doi.org/10.4060/ca8642zh
本信息产品中使用的名称和介绍的材料,并不意味着联合国粮食及农业组织(粮农组织)或联合国环境规划署(环境署)对任何国家、领地、城市、地区或其当局的法律或发展状态、或对其国界或边界的划分表示任何意见。地图中使用的名称和介绍的材料,并不意味着粮农组织或环境署对任何国家、领土或海区的法定或构成地位或其边界的划分表示任何意见。提及具体公司或厂商产品,无论是否含有专利,并不意味着这些公司或产品得到粮农组织或环境署的认可或推荐,优于未提及的其它类似公司或产品。
本信息产品中陈述的观点是作者的观点,不一定反映粮农组织或环境署的观点或政策。
ISBN 978-92-5-132424-0
ISSN 1020-5748 [印刷]
ISSN 2663-8037 [在线]
© 粮农组织 2020年

保留部分权利。本作品根据署名-非商业性使用-相同方式共享3.0政府间组织许可(CC BY-NCSA 3.0 IGO; https://creativecommons.org/licenses/by-nc-sa/3.0/igo/deed.zh)公开。
根据该许可条款,本作品可被复制、再次传播和改编,以用于非商业目的,但必须恰当引用。使用本作品时不应暗示粮农组织认可任何具体的组织、产品或服务。不允许使用粮农组织标识。如对本作品进行改编,则必须获得相同或等效的知识共享许可。如翻译本作品,必须包含所要求的引用和下述免责声明:“该译文并非由联合国粮食及农业组织(粮农组织)生成。粮农组织不对本翻译的内容或准确性负责。原[语言]版本应为权威版本。”
任何与在此许可下出现的纠纷有关的调解,均应根据现行《联合国国际贸易法委员会仲裁规则》进行。
第三方材料。欲再利用本作品中属于第三方的材料(如表格、图形或图片)的用户,需自行判断再利用是否需要许可,并自行向版权持有者申请许可。对任何第三方所有的材料侵权而导致的索赔风险完全由用户承担。
销售、权利和授权。粮农组织信息产品可在粮农组织网站(www.fao.org/publications)获得,也可通过publications-sales@fao.org购买。商业性使用的申请应递交至www.fao.org/
封面图片:©Ricky Martin/CIFOR
印度尼西亚:一名当地人在Gede Pangrango的森林湖里捕鱼以满足其日常生活的需要。
3. 截至2019年12月,世界自然保护联盟红色名录中森林动植物和真菌的脆弱性状况
9. 2015年按斑块大小等级和全球生态区划分的森林面积构成比例
12. 2015年按森林密度等级和全球生态区划分的森林面积构成比例
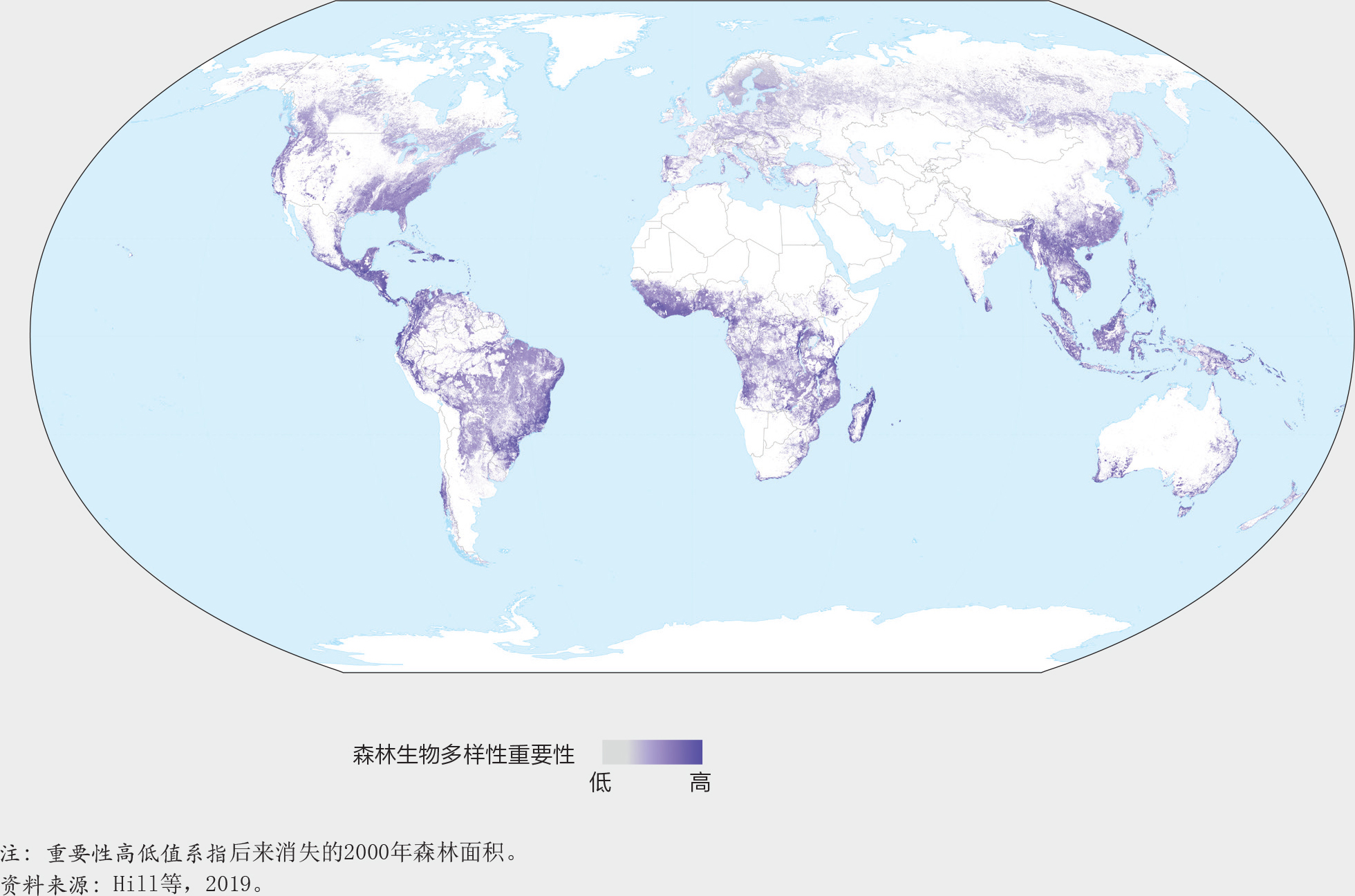
20. 森林生物多样性对2000-2018年间森林面积减少的重要性
22. 2018年森林生物群落内森林生物多样性重要性和完整性的双变量图
23. 2018年森林生物群落内森林生物多样性重要性和完整性双变量图的详细信息
24. 1970-2014年间,基于268种仅存于森林的脊椎动物(455个种群)的指数总体下降
29. 按区域划分的2000-2010年间毁林和森林退化驱动因素
30. 资源利用的进程、政策和驱动力之间的互动影响当地对森林保护的应对和结果
31. 毁林和森林退化的复杂驱动因素:来自赞比亚的分析逻辑树
32. 已确定的31个国家REDD+战略和行动计划中减少森林砍伐和退化的优先行动领域
33. 按区域分的2000-2015年间处于退化状态的土地比例(%)
35. 2000-2019年按区域和恢复类型计的通过森林恢复、再造林和造林活动而增加的森林面积
39. 1992-2015年按森林类型划分的保护区内森林面积的增加
40. 1992-2015年按全球生态区划分的保护区内森林的增加
42. 1990-2020年按区域计的主要指定用于生物多样性保护的森林面积趋势
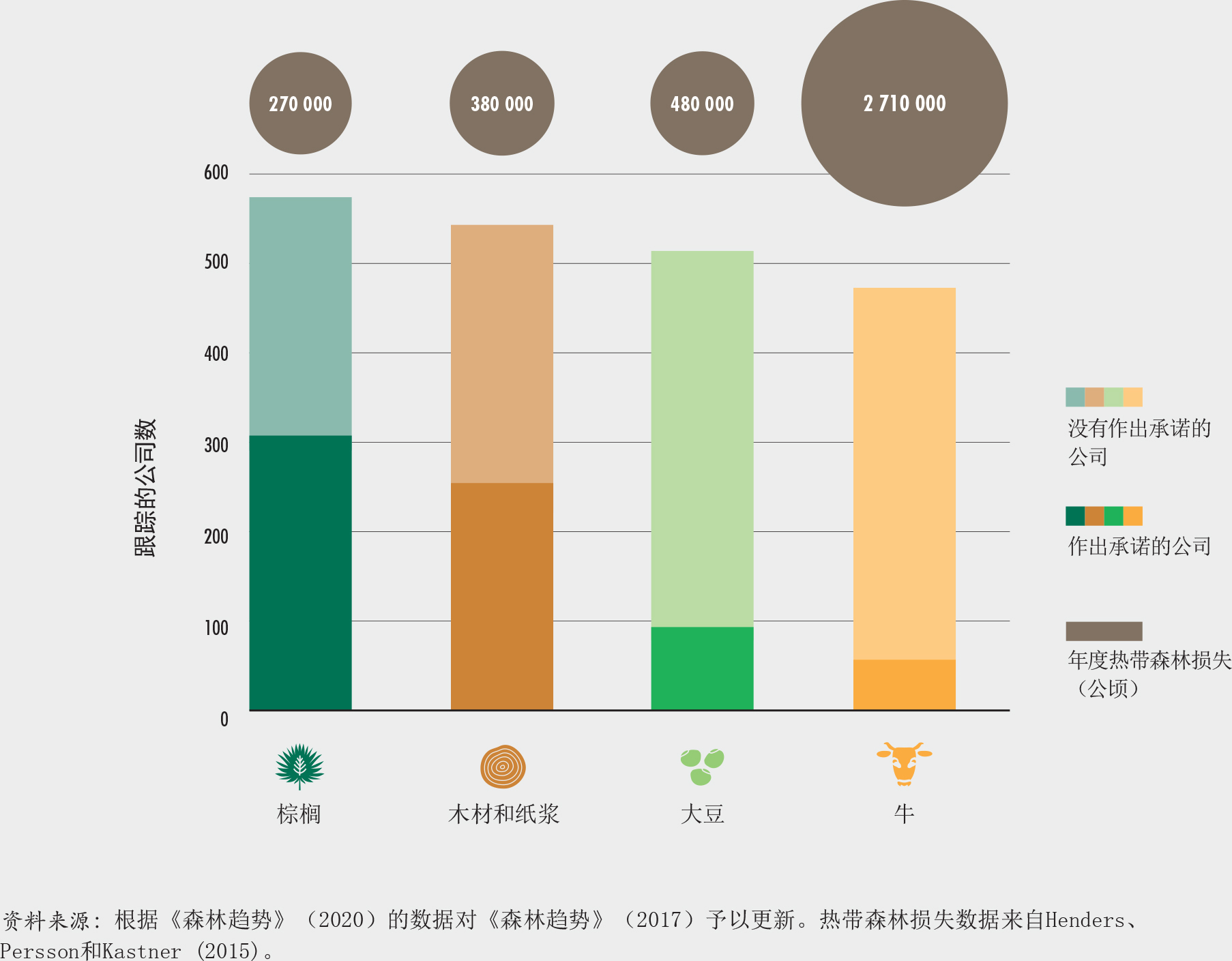
43. 按商品划分的2020年已作出以及尚未作出与毁林相关承诺的公司数量
4. 保护和利用与森林相关生物多样性的国际文书以及相关目标和具体目标
15. 与保护森林物种和遗传资源有关的关键目标、具体目标和指标
32. 森林是中国哈尼水稻梯田应对气候变化能力和保护农业生物多样性的关键要素
35. 《联合国气候变化框架公约》和《巴黎协定》下的REDD+
45. 与保护区和其他基于区域的保护措施有关的关键目标、具体目标和指标
正值我们对《2020年世界森林状况》最终润色之际,全世界突然迎面遭遇了2019冠状病毒病(COVID-19)疫情这一前所未有的挑战。尽管全球当务之急是应对这一突发公共卫生紧急情况,但我们的长期对策必须是解决此类疫病的根本原因。森林的退化和丧失就是其中一个驱动因素,对自然平衡造成破坏,加剧了人类感染人畜共患病的暴露风险。了解并跟进世界森林状况,变得从未如此重要。
今年标志着“联合国生物多样性十年”以及《2011-2020年生物多样性战略计划》实施工作的结束。各国正共同努力,审议该战略计划五项战略目标和20个爱知生物多样性目标的进展状况,从而形成2020年后全球生物多样性框架。
该框架必须以证据为基础:有关世界生物多样性现状和近期趋势的证据;生物多样性与可持续发展之间联系的证据;以及为保护和可持续利用世界生物多样性提供的众多产品和服务以支持粮食安全和人类福祉而采取的成功行动的证据。
绝大多数陆地生物多样性都存在于世界森林之中 — 从极北的北方针叶林到热带雨林。总体而言,这些森林共蕴含了6万多不同树种,为80%的两栖动物、75%的鸟类和68%的哺乳动物提供了栖息地。热带森林中有约60%的维管植物。红树林为众多鱼类和贝类提供了繁殖地和抚育地,并有助于吸收沉积物,否则这些沉积物可能对海草床和珊瑚礁,也就是海洋生物的栖息地,产生不利影响。
因此,世界大部分生物多样性的保护完全取决于我们如何与世界森林互动,以及我们利用森林的方式。
本版《世界森林状况》探讨了森林以及利用和管理森林的人类对保护和可持续利用生物多样性所做的贡献。本书评估了迄今在实现与森林生物多样性有关的全球目标方面取得的进展,并通过创新做法和双赢解决方案的案例研究,说明了保护和可持续发展方面政策、行动和方法的有效性。
本出版物目的并不是要成为关于森林生物多样性的全面论述,而是介绍其当前发展状况,并概述其对人类的重要意义。本书旨在补充联合国粮食及农业组织(粮农组织)粮食和农业遗传资源委员会于2019年发布的《世界粮食和农业生物多样性状况》、以及去年政府间生物多样性和生态系统服务科学政策平台(IPBES)发布的《生物多样性和生态系统服务全球评估报告》和《生物多样性公约》(CBD)发布的第五版《全球生物多样性展望》。
本版《世界森林状况》首次由两家联合国机构 — 粮农组织和联合国环境规划署(环境署)— 共同合作完成。基于现有合作及各自比较优势,我们把粮农组织《2020年全球森林资源评估》生成的最新信息与联合国环境规划署世界自然保护监测中心(UNEP-WCMC)对受保护森林的状况和代表性开展的分析融汇在一起。
《2020年世界森林状况》证实,毁林和森林退化仍以惊人的速度在继续,大大加剧了生物多样性的丧失。农业扩张仍然是主要驱动力之一,而同时人类粮食系统的复原力及其适应未来变化的能力恰恰取决于生物多样性。
《2020年世界森林状况》同时还看到了希望迹象。全球森林损失率正在下降,而且确实存在能够在保护与可持续利用森林生物多样性之间实现平衡的解决方案。为扭转毁林和生物多样性丧失的趋势,我们迫切需要看到这些解决方案得到推广,同时还要改变我们生产和消费食物的方式。我们还需要通过综合景观方法保护和管理森林和树木,并通过森林恢复工作来修复已经造成的破坏。
变革的关键是有效治理、各部门和行政级别之间的政策协调、有保障的土地权属、对当地社区和土著人民权利和知识的尊重、加强监测生物多样性成果的能力,以及创新的筹融资方式。
最后,我们必须要与自然建立起一种新型关系,一起努力,就能加以实现。《2020年世界森林状况》有助于实现这一愿景。我们希望本出版物内容有趣、富有价值并能带来启发。
|
粮农组织总干事 |
环境规划署执行主任 |
《2020年世界森林状况》由粮农组织林业政策及资源司与联合国环境规划署世界保护监测中心(UNEP-WCMC)合作编写。
报告的编写工作在由粮农组织和世界保护监测中心的五名资深专家组成的核心小组的指导之下完成。粮农组织林业政策及资源司司长领导该核心小组,并负责全面协调。
对实现与森林及其生物多样性有关的目标和具体目标进展状况的评估,主要基于现有文献和专门研究。同时还汇编了一系列案例研究,提供世界各地保护和可持续利用森林生物多样性方面的实例。
本版《世界森林状况》借鉴了粮农组织《2020年全球森林资源评估》(FRA 2020)的结果,后者也将于2020年发布。
《2020年全球森林资源评估》对1990-2020年间236个国家和地区的60多个涉及森林的程度、特征、状况、管理和用途的变量进行了现状和趋势研究。
《2020年全球森林资源评估》的基石是官方数据,这些数据由各国官方指定的通讯员提供,报告流程为一个具有透明度且可追溯的整合系统。标准化报告方法的应用可以监测变量随时间的发展变化,也可以对数据进行区域和全球的汇总。
《2020年世界森林状况》仅采用了与森林生物多样性有关的数据。大多数数据是全球层面的,并基于不久之前发布的《2020年森林资源评估关键发现》。读者可以在即将发布的《2020年全球森林资源评估》(粮农组织,2020)中找到更多区域和国家层面的详细信息。《2020年全球森林资源评估》中使用的术语和定义可以在http://www.fao.org/3/I8661EN/i8661en.pdf上找到。
为筹备编写《2020年世界森林状况》,专门委托进行了三项新研究:
世界保护监测中心对1992-2015年间每年土地覆盖数据的分析,提供了树木覆盖面积每年如何显著变化的最新信息。随后又对此做了进一步探讨,将粮农组织的全球生态区地图、世界关键生物多样性区域数据库(WDKBA)和世界保护区数据库(WDPA)联系起来,产生了关于保护区代表性和森林保护状况随时间变化的新发现。
欧洲委员会联合研究中心与美国林务局合作,将现有的用于分析森林空间分布的方法应用于2015年全球哥白尼土地覆盖地图之上,再与粮农组织的全球生态区图叠加。该研究提供了按广泛的森林类型划分的有关森林完整性和破碎化状况的新数据。
世界银行对森林与贫困之间的关系进行了研究。该研究主要基于文献综述以及将森林地图与世界银行持有的贫困数据相叠加。
所有章节都得益于工作人员和顾问专家在数据收集和/或编写方面的支持。最终出版物由一位高级顾问专家整理和编辑而成。
来自粮农组织和环境署不同部门的内部同行评审专家以及外部同行评审专家对初稿提供了许多宝贵的意见和建议。
《2020年世界森林状况》在Mette L. Wilkie的总体协调之下完成。由其领导的核心小组成员包括来自粮农组织的Anssi Pekkarinen、Ewald Rametsteiner、Andrew Taber和Sheila Wertz-Kanounnikoff,以及来自联合国环境规划署世界保护监测中心的Will Simonson。Andrea Perlis为核心小组在编写和编辑上提供了协助。其他提供贡献的编撰人员和审阅人如下:
粮农组织:
撰写专家:Hitofumi Abe、Safia Aggarwal、Astrid Agostini、Damien Bertrand、Simone Borelli、Marco Boscolo、Pierre Bouillon、Amanda Bradley、Anne Branthomme、Vito Brito、Lyndall Bull、Malgorzata Buszko-Briggs、Benjamin Caldwell、Laura Cerioni、Michela Conigliaro、Jose Diaz Diaz、Yoshihide Endo、Aurelie Fernandez、Serena Fortuna、Julian Fox、Sarah Fumey、Monica Garzuglia、Emma Gibbs、Marta Gruca、Abdel Hamied Hamid、Daphne Hewitt、Sooyeon Jin、Örjan Jonsson、Adolfo Kindgard、Jarkko Koskela、Arvvdas Lebedys、Thais Linhares Juvenal、Erik Lindquist、Yuka Makino、Peter Moore、Giulia Muir、Azdad Mustapha、Scott Newman、Maria Isabel Ochoa、Chiara Patriarca、Peter Pechaek、Clelia Maria Puzzo、Sara
Casallas Ramirez、Kristina Rodina、Moctar Sacande、Shiroma Sathyapala、Kenichi Shono、Bianca Sipala、Simona Sorrenti、Elaine Springgay、Ashley Steel、Tiina Vähänen、Martina Venturi、Pedro Vivar、Anni Vuohelainen、Sven Walter、夏祖璋和张道卫。
审阅专家:Julie Belanger、Lorenzo Bellu、Nora Berrahmouni、Jeffrey Campbell、Frederic Castell、Ana Paula De la Ocampos、Michael Euler, Adriana Ignaciuk、Lourdes Orlando、Dafydd Pilling、Eran Raizman、Selvaraju Ramasamy、Kostas Stamoulis和Carlos Vaquero。
环境规划署和环境规划署世界保护监测中心:
撰写专家:Andy Arnell、Abigail Burns、Lauren Coad、Alexander Gangur、Joe Gosling、Samantha Hill、Lisa Ingwall-King、Valerie Kapos、Steven King、Edward Lewis、Calum Maney、Emma Martin、Ana Paula de la O Campos、Barbara Pollini、Marieke Sassen、Emma Scott、Arnout van Soesbergen和James Vause。
审阅专家:Abdelkader Bensader、Neil Burgess、Katherine Despot-Belmonte、Satu Glaser、Kelly Malsch和Susan Mutebi-Richards。
欧洲委员会联合研究中心(关于森林破碎化的研究):
Peter Vogt。
美国林务局(关于森林破碎化的研究):
Kurt Ritters。
世界银行(关于森林与贫困的研究):
撰写专家:Shun Chonabayashi(Yulin Chen、Shanjun Li、Luming Tan和Ziye Zhang提供支持)。
审阅专家:Benoît Blarel、Timothy H. Brown、Susmita Dasgupta、Martin Heger、Minh Cong Nguyen。
案例研究和插文:
案例研究和插文由粮农组织和环境规划署世界保护监测中心的工作人员以及如下外部贡献者提供:
关于约旦达纳生物圈保护区的案例研究:Qamar Almini、Nashat Hamidan和Amer Rfou’(约旦皇家自然保护协会),以及Mohammad Alnsour(约旦流域和发展计划)。
关于北美野生动物保护的案例研究:Shane Patrick Mahoney(Conservation Visions总裁)。
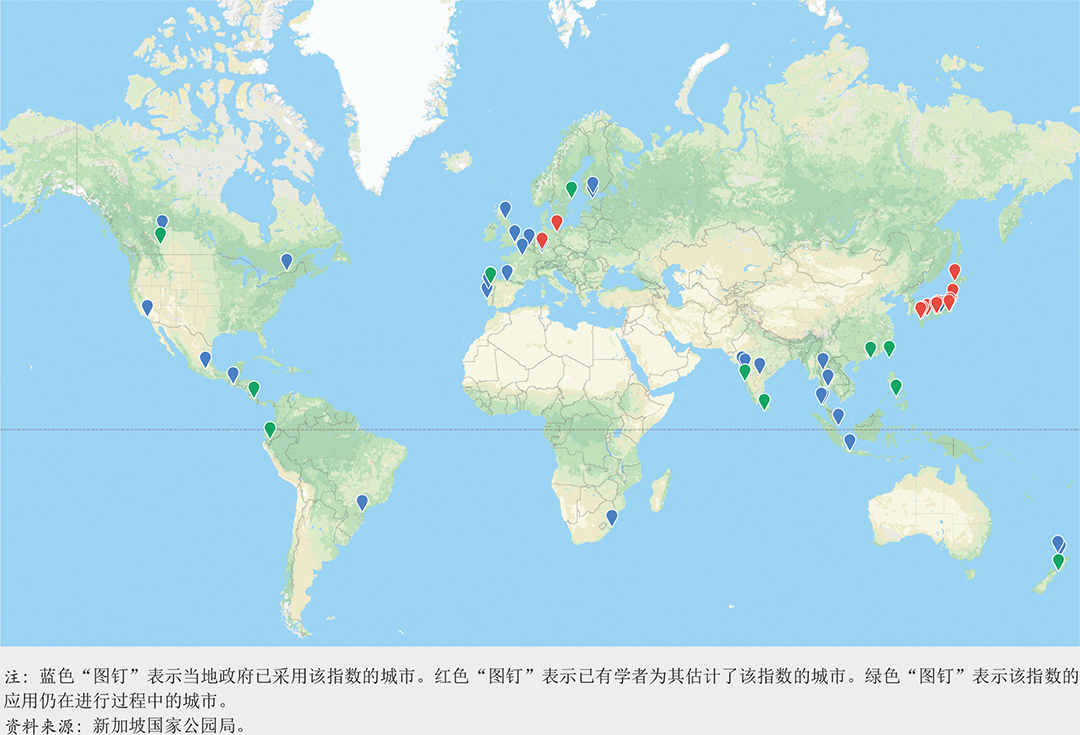
关于新加坡城市生物多样性指数的案例研究:Lena Chan(新加坡国家公园局)。
关于欧洲保护森林遗传资源区域战略的插文:Michele Bozzano(欧洲森林研究所森林遗传资源计划)。
关于评估威胁布基纳法索食用树种遗传资源保护因素的插文:Hannes Gaisberger和Barbara Vinceti(国际生物多样性中心)。
《2020年世界森林状况》还得益于David Cooper和Lisa Janishevski(《生物多样性公约》秘书处)、Christel Palmberg-Lerche(粮农组织前雇员)和Fred Stolle(世界资源研究所)的外部审评意见,以及许多粮农组织内部其他技术部门同事对个别章节的评论意见。
粮农组织会议规划及文件处提供了翻译及印刷服务。粮农组织全组织交流办公室出版处为所有六种官方语言版本提供编辑支持、设计和排版以及制作协调。
AAD
防治荒漠化行动
ABS
获取和利益分享
ADB
非洲开发银行
AU
非洲联盟
BESNet
生物多样性和生态系统服务网络
BGCI
国际植物园保护联盟
CAFI
中非森林倡议
CATIE
热带农业研究与高等教育中心
CBD
生物多样性公约
CBI
城市生物多样性指数
CBNRM
基于社区的自然资源管理
CEPF
关键生态系统合作基金
CFS
世界粮食安全委员会
CGRFA
粮食和农业遗传资源委员会
CIFOR
国际林业研究中心
CIRAD
国际发展农业研究中心
CITES
濒危野生动植物种国际贸易公约
COMIFAC
中非森林委员会
CONAFOR
墨西哥国家林业委员会
CONAP
危地马拉国家保护区委员会
CPF
森林合作伙伴关系
CPW
野生动物可持续管理合作伙伴关系
CRITFC
哥伦比亚河部落间鱼委会
DBR
达纳生物圈保护区
DFSC
达尼达森林种子中心
EC
欧盟委员会
ESA
欧洲航天局
ESA CCI
欧洲航天局气候变化倡议
EU
欧洲联盟
EUFGIS
欧洲森林遗传资源信息系统
EUFORGEN
欧洲森林遗传资源计划
FAO
联合国粮食及农业组织
FAOSTAT
粮农组织统计数据库
FCPF
森林碳伙伴基金
FERI
森林生态系统恢复倡议
FLD
丹麦森林与景观
FLEGT
森林执法、治理和贸易行动计划
FONAFIFO
哥斯达黎加国家林业融资基金
FPIC
自由、事先和知情同意
FRA
全球森林资源评估
FSC
森林管理委员会
GBP
英镑
GCF
绿色气候基金
GDP
国内生产总值
GEF
全球环境基金
GEZ
全球生态区
GPFLR
全球森林与景观恢复伙伴关系
HLPE
世界粮食安全委员会粮食安全和营养问题高级别专家小组
HWC
人与野生动物的冲突
ICCA
土著人民和地方社区保护的领土或地区
IDS
英国苏塞克斯大学发展研究所
IFAD
国际农业发展基金
IFPRI
国际粮食政策研究所
IIED
国际环境与发展研究所
ILO
国际劳工组织
IMF
国际货币基金组织
INAB
危地马拉国家森林研究所
INBAR
国际竹藤组织
INTERPOL
国际刑警组织
IPBES
生物多样性和生态系统服务政府间科学政策平台
IPCC
国际植物遗传资源研究所
IPGRI
政府间气候变化专门委员会
ITC
国际贸易中心
ITTO
国际热带木材组织
IUCN
世界自然保护联盟
IUCN WCPA
世界自然保护联盟世界保护区委员会
JRC
欧盟委员会联合研究中心
KBA
关键生物多样性区域
MAP
药用芳香植物
MEA
千年生态系统评估
MEF
印尼环境与林业部
MERECP
埃尔贡山地区生态系统保护计划
MINEF
喀麦隆森林与环境部
MINEPDED
喀麦隆环境、自然保护与可持续发展部
MINFOF
喀麦隆森林与野生动物部
MIPAAF
意大利农业、粮食和林业政策部
MNRT
坦桑尼亚联合共和国自然资源和旅游部
MoE
约旦环境部
MoP
约旦计划与国际合作部
MPP
山区合作伙伴产品计划
NACSO
纳米比亚社区自然资源管理支持组织协会
NCED
国家地役权保护数据库
NDC
国家自主贡献
NGO
非政府组织
NGS
国家地理学会
NWFP
非木质林产品
NYDF
纽约森林宣言
OECD
经济合作与发展组织
OECM
其他有效的基于区域的保护措施
OIE
世界动物卫生组织
PES
生态系统服务付费补偿
PFM
参与式森林经营
PNAS
美国国家科学院院报
PREDICTS
不断变化的陆地系统中生态多样性的预测反应
REDD+
减少毁林和森林退化所致排放以及发展中国家的森林保护、可持续管理和增加森林碳储量的作用
RNZ
新西兰电台
RRI
权利与资源倡议
RSCN
约旦皇家自然保护协会
SADC
南部非洲发展共同体
SDG
可持续发展目标
SEEA
环境经济核算体系
SEGeF
监测生产林中的野生动植物管理
SEPAL
土地监测地球观测数据获取、处理和分析系统
SI
新加坡城市生物多样性指数
SMFE
中小型森林企业
SOFO
世界森林状况
SPDA
秘鲁环境法学会
SVLK
印度尼西亚木材合法性验证系统
TFCA
美国热带森林保护法
UAESPNN
国家自然公园系统特别行政单位
UN
联合国
UN-REDD
联合国减少毁林和森林退化所致排放计划
UNCCD
联合国防治荒漠化公约
UNCTAD
联合国贸易和发展会议
UNDESA
联合国经济和社会事务部
UNDP
联合国开发计划署
UNEP
联合国环境规划署
UNEP-WCMC
联合国环境规划署世界保护监测中心
UNESCO
联合国教育、科学及文化组织
UNFCCC
联合国气候变化框架公约
UNICEF
联合国儿童基金会
UNODC
联合国毒品和犯罪问题办公室
USAID
美国国际开发署
USD
美元
USDA
美国农业部
US/ICOMOS
国际古迹遗址理事会美国委员会
VFR
村镇森林保护区
WCMC
世界自然保护监测中心
WCPA
世界保护区委员会
WCS
野生动物保护协会
WDPA
世界保护区数据库
WHO
世界卫生组织
WRI
世界资源研究所
WWF
世界自然基金会
ZSL
伦敦动物学会
联合国生物多样性十年(2011-2020)即将到期,各国政府正着手准备采用2020年后全球生物多样性框架。值此之际,本版《世界森林状况》(SOFO)考察了森林以及利用和管理森林的人类对保护和可持续利用生物多样性所做的贡献。本报告旨在补充联合国粮食及农业组织(粮农组织)于2019年2月发布的《世界粮食和农业生物多样性状况》;生物多样性与生态系统服务政府间科学政策平台(IPBES)发布的《生物多样性和生态系统服务全球评估报告》(初稿发布于2019年);以及《生物多样性公约》即将发布的第五版《全球生物多样性展望》。
森林为地球大部分陆地生物多样性提供了庇护之所。因此,世界生物多样性的保护完全取决于我们与世界森林如何互动,以及我们如何利用森林。森林为80%的两栖动物、75%的鸟类和68%的哺乳动物提供了栖息地。热带森林中可发现约60%的维管植物。红树林为无数鱼类和贝类提供了繁殖地和抚育地,吸收了可能对海草床和珊瑚礁产生不利影响的沉积物,而这些正是更多海洋物种的栖息地。
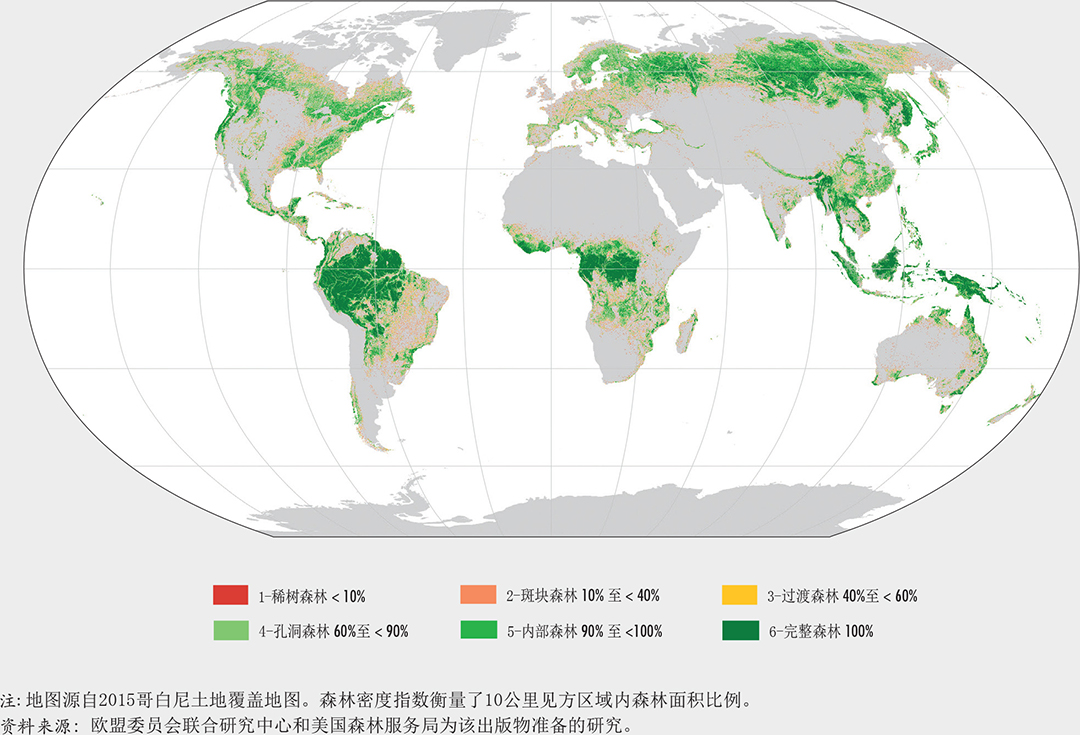
森林占全球土地面积的31%,但并非均匀分布于地球各地。大约一半的森林相对完整,超过三分之一的森林是原始森林。一半以上的世界森林仅分布在五个国家(巴西、加拿大、中国、俄罗斯联邦和美国)。大约一半的森林面积(49%)相对完整,而9%的森林则是严重破碎的,连通性较差或几乎没有连通性。热带雨林和北方针叶林破碎化最低,而亚热带干旱林和温带阔叶森林具有最高的破碎度。全世界约80%的森林斑块规模超过100万公顷。其余20%的森林遍布世界3400万个森林斑块之中,绝大多数斑块规模小于1000公顷。
超过三分之一(34%)的世界森林是原始森林,定义为原生树种的天然再生林,没有明显的人类活动迹象,生态过程也没有受到明显干扰。
毁林和森林退化仍以惊人的速度不断发生,导致了生物多样性的持续显著减少。尽管毁林速度在过去的30年间已有所降低,但自1990年以来,据估计森林面积通过转为其他用地丧失了4.2亿公顷。在2015-2020年间,毁林速度据估计为每年1000万公顷,低于上世纪90年代的每年1600万公顷。世界范围内的原始森林从1990年起已经减少了超过8000万公顷。超过1亿公顷的森林受到了火灾、病虫害、入侵物种、干旱和灾害性天气事件的不利影响。
农业扩张仍然是毁林和森林退化以及森林生物多样性丧失的主要驱动因素。2000-2010年间,大规模商业化农业生产(主要是养殖牛、种植大豆和油棕)导致了热带地区毁林的40%,当地的自给农业导致了另外的33%。具有讽刺意味的是,人类粮食系统的适应力及其应对未来变化的能力恰恰取决于这些生物多样性,包括有助于防治荒漠化的适应干旱地区的灌木和树木,为农作物授粉的森林、昆虫、蝙蝠和鸟类,有利于防止土壤侵蚀的山岳生态系统内根系发达的树木,以及可以提高防洪能力的红树林。随着气候变化日益加剧了粮食系统的风险,森林对农业部门来说在固碳以及缓解气候变化中的作用也变得越来越重要。
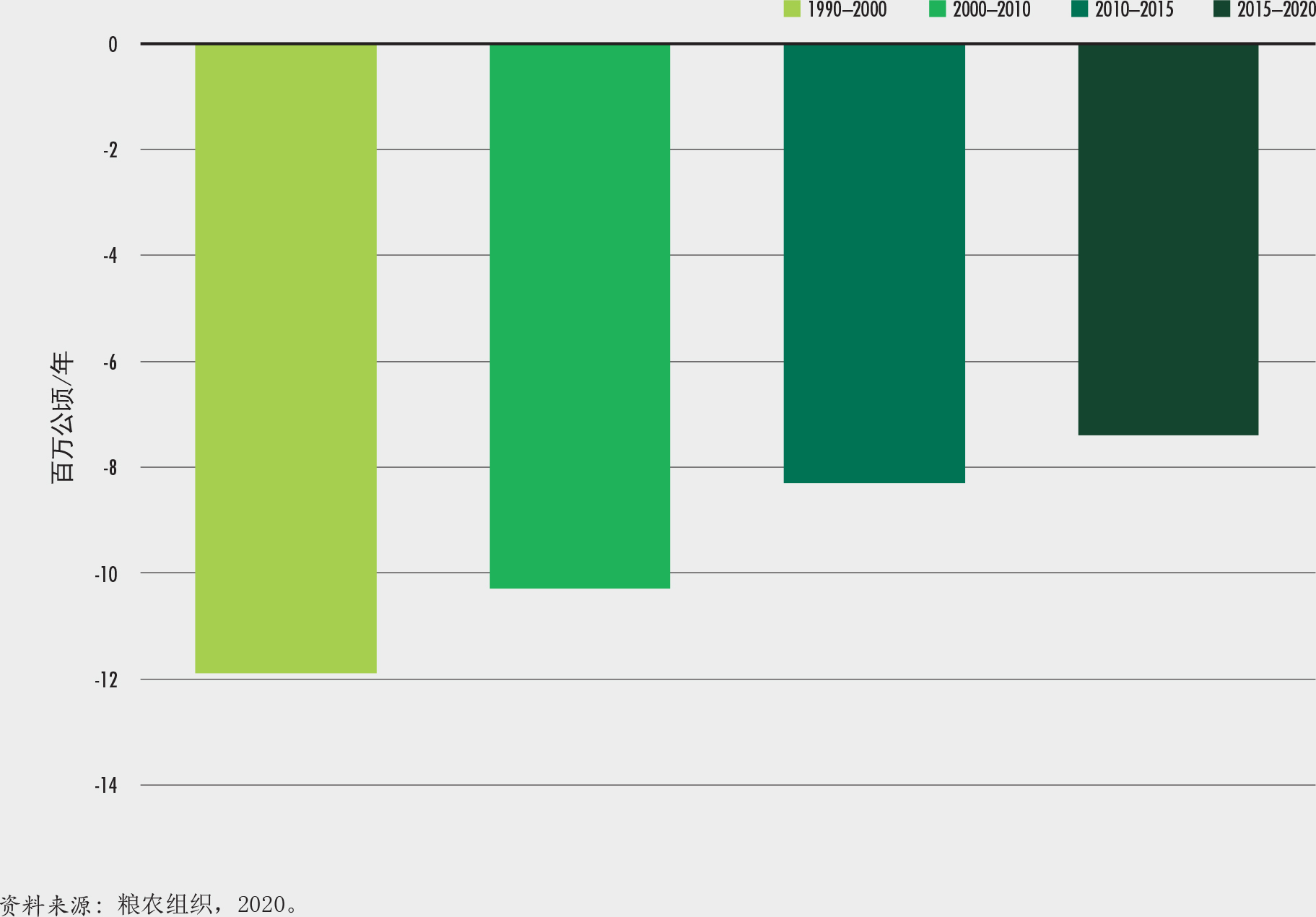
森林面积净损失由上世纪90年代的每年780万公顷降低至2010-2020年间的每年470万公顷。尽管毁林在一些地区仍在继续,然而在另一些地区以自然扩张和人工辅助的方式新建的森林逐渐增加。从绝对数值来看,全球森林面积在1990-2020年间共减少了1.78亿公顷,规模大致与利比亚国家面积相当。
除了受人类的利用影响之外,森林的生物多样性还因森林类型、地理条件、气候和土壤等因素而存在极大的差别。温带地区的大多数森林生境蕴含的动物物种和树种较少,且这些物种的地理分布较广。非洲、南美和东南亚的山地森林以及澳大利亚、巴西沿海岸、加勒比海岛屿、中美洲和东南亚岛屿的低地森林则具有许多物种,地理分布较集中。人口和农业用地密集的地方(如欧洲、孟加拉国局部地区、中国、印度和北美等)从生物多样性角度来说完整性较差。北非、澳大利亚南部、巴西沿海、马达加斯加和南非也被认定为是生物多样性的完整性遭受重大损失的地区。
在防止已知受威胁物种灭绝和改善其保护情况方面的进展较为缓慢。人类已知60000个不同树种,其中超过20000个树种已被列入世界自然保护联盟(IUCN)的濒危物种红色名录,超过8000个树种被评定为全球范围受胁的(极危、濒危、易危)。多于1400个树种被评为极危,亟需采取保护行动。约8%被评估的森林植物、5%的森林动物、以及5%的森林真菌目前被列为极危物种。
仅存于森林的物种指数的构建基于被监测的268种森林哺乳动物、两栖动物、爬行动物和鸟类的455个种群。该指数在1970-2014年间降低了53%,年降幅为1.7%。这也凸显了这些物种面临灭绝的危险性。
值得欣慰的是,122个缔约方通过了《关于获取遗传资源和公正和公平分享其利用所产生惠益的名古屋议定书》(比2016年增长74%),146个缔约方通过了《粮食和农业植物遗传资源国际条约》。
所有人都依赖森林及其生物多样性,而一些人比另一些人依赖性更强一些。森林为超过8600万人提供了绿色工作,支持了更多人的生计。全球据估计有8.8亿人要花一部分时间用于采集薪柴或生产木炭,其中许多为妇女。在低收入国家森林覆盖高、生物多样性丰富的地区,人口一般较少,但是这些地区的贫困率也通常较高。约有2.52亿居住在森林和稀树草原的人每天收入少于1.25美元。
墨西哥
每年有数以百万计的帝王蝶(Danaus plexippus)从加拿大迁移到墨西哥,在那里的森林中过冬。
满足人类粮食需求和可持续利用生态系统具有互补性且为互相依赖的目标。森林提供水,减缓气候变化,为许多对可持续粮食生产起重要作用的授粉者提供栖息地。据估计世界75%最主要的粮食农作物(占全球35%粮食生产)得益于动物在水果、蔬菜或种子生产中的授粉作用。
在全球范围内,大约有10亿人在某种程度上依赖野生食物,例如野味肉、食用昆虫、食用植物产品、蘑菇和鱼类,它们通常内含大量关键的微量元素。森林食物作为营养资源的价值并不仅限于中低收入国家;欧盟超过1亿民众经常性食用野生食物。24亿人口(城市和农村)用木质燃料烹煮食物。
人类健康和生计福祉都与森林息息相关。目前超过28000种植物据记载有药物作用,其中许多被发现存在于森林生态系统之中。接触自然环境对人们的身心健康都有积极影响,许多人还与森林有着深厚的精神联系。然而,森林也对健康产生风险。与森林相关的疾病包括疟疾、查加斯病(又称美洲锥虫病)、非洲锥虫病(嗜睡病)、利什曼病和莱姆病、艾滋病和埃博拉病。绝大多数影响人类的新疾病,包括导致当前2019冠状病毒大流行的新型冠状病毒(SARS-CoV2),属于人畜共患病,它们的出现也许与森林面积改变和人类活动向森林延申而引发的栖息地丧失有关,这些变化增加人类与野生动物接触的机会。
森林生物多样性保护和可持续利用之间取得平衡的解决方法是关键,也是可行的。人类对生物多样性的影响并非所有都是负面的,本版报告展示了许多最近在管理、保护、恢复和可持续利用森林生物多样性方面成功的项目计划案例。
遏止毁林和非法采伐在过去十年中加快了步伐,国际协定和基于结果的付费补偿也同样取得长足发展。到目前为止,七个国家向《联合国气候变化框架公约》(UNFCCC)报告其毁林有所减少,各国现在可以从绿色气候基金和其他类似机制获得基于结果的支付奖励。消费国的贸易法规带动了应对非法采伐的行动,这些法规要求进口商证明木材是合法采伐的。许多热带木材生产国正在做出相应努力,以加强对合法性法规的遵守和取得木材合法性验证。其中的15个国家正在建立国家体系,实现确保在欧盟森林执法治理和贸易机制之下木材运营的合法性。作为该机制的一部分,要求各国采取措施以防止非法狩猎。
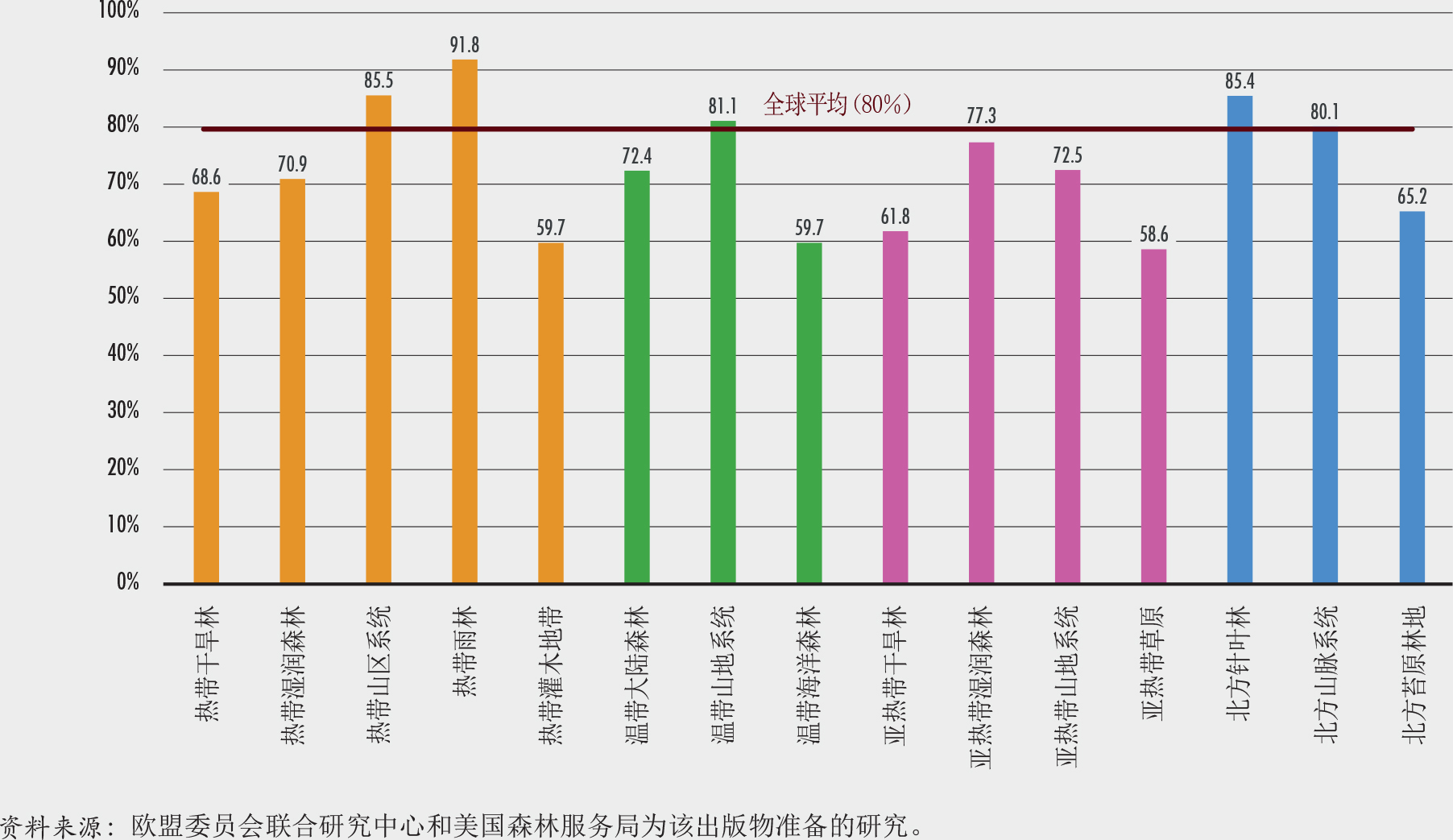
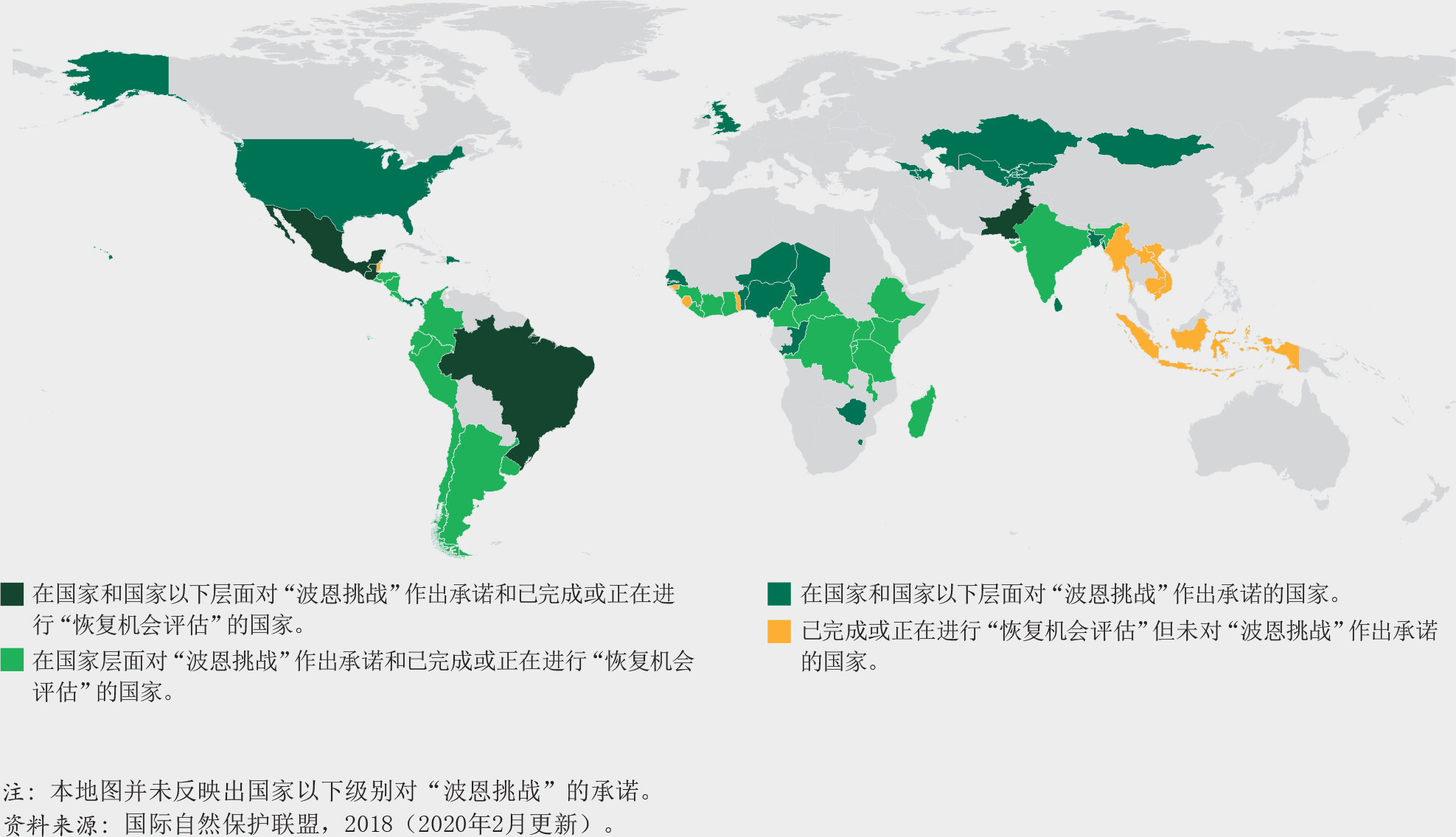
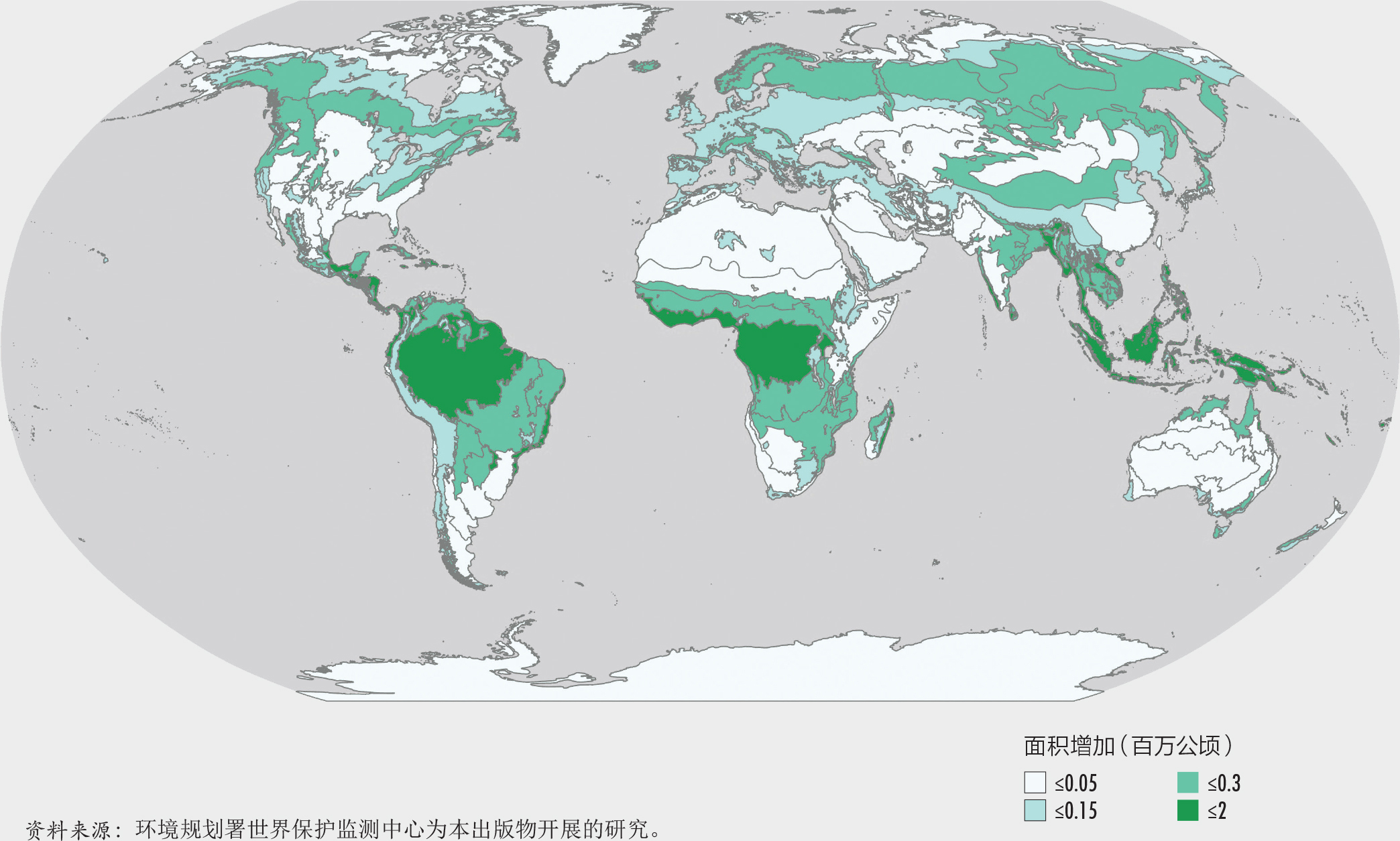
森林生态系统作为一个整体已经超越了爱知生物多样性目标11(到2020年保护至少17%的陆地面积)。然而,仅有保护区对保护生物多样性来说是不够的。从全球来看,世界森林面积的18%(超过7亿公顷)处于如国家公园、保护区和野生动物保护区等(国际自然资源保护联盟第I到第IV类保护区)依法设立的保护区范围内。然而,这些保护区尚未完全代表森林生态系统的多样性。专门为本报告进行的一项探讨1992-2015年间按全球生态区(GEZs)划分的森林保护区趋势的研究表明,超过30%的热带雨林、亚热带干旱林和温带海洋林在2015年已处于法律保护的地区(第I到第VI类保护区)。该研究还发现,亚热带湿润森林、温带草原和北方针叶林应当在未来的决策中予以优先考虑,因为这些地区目前只有少于10%的森林处于被保护的状态。其他在生物多样性和完整性上有价值的地区也应当高度重视,包括安第斯山脉北部和中美洲、巴西东南部、刚果盆地的部分地区、日本南部、喜马拉雅山、东南亚和新几内亚的部分地区。
到目前为止,在设定其他有效的基于区域的保护措施方面取得的进展甚为有限,但是有关这类保护的指导性文件正在准备之中,这类措施在保护森林方面很有潜力。
森林还未达到爱知生物多样性目标7的要求(到2020年,对农业、水产养殖业和林业用地实现可持续管理,确保生物多样性保护),然而世界森林管理水平正在逐步改善。长期管理计划之下的森林面积在过去30年显著上升,增长到2020年的大约20.5亿公顷,相当于全球森林面积的54%。
当前生物多样性和生态系统的不利趋势将会有损于实现可持续发展目标(SDGs)的进展。世界生物多样性是地球生命的基础,目前尽管有一些积极的趋势,但是生物多样性仍在以很快的速度继续丧失。我们亟需变革自己如何管理森林及其生物多样性、生产和消费食物的方式,以及与自然互动的模式。我们必须将环境恶化和不可持续的资源利用与经济增长及相关的生产和消费方式分离开来,而且在进行土地使用决策时必须考虑到森林的真正价值。
确保有益于生物多样性和人类的积极成果,需要在保护目标和因生计而对资源的需求之间达成现实的平衡。迫切需要将生物多样性纳入所有类型森林经营管理的主流。为此,必须在保护目标和为了满足生计、粮食安全和人类福祉对资源产生的需求之间取得平衡。这需要各部门之间和行政层级之间政策协调一致;有保障的土地所有权;对当地社区和土著居民的权利和知识的尊重;增强监测生物多样性结果的能力。同时也需要创新型筹资融资方法。
我们需要改变我们的粮食系统来遏止毁林和生物多样性的丧失。在这方面,需要做出最大改变的就是我们生产和消费粮食的方式。鉴于农业扩张是毁林的主要驱动力,我们生产和消费粮食的方式需要进行颠覆性的变革。我们必须摆脱目前因粮食需求而致的不恰当的农业模式,这些模式促使森林被大规模转为农业生产,与此同时也造成了与森林有关的生物多样性的丧失。采取混农林业和可持续生产方式,恢复退化农业用地的生产力,采用来自可持续粮食体系的更健康的饮食,减少粮食损失和浪费,这些都是迫切需要在更大范围和规模上采取的行动。农业企业必须履行其对零毁林商品链的承诺,而尚未做出零毁林承诺的企业也应当这样做。商品投资者应采用对环境和社会负责的商业模式。在许多情况下,这些行动的实施意味着需要调整当前的政策(特别是财政政策)和监管框架。
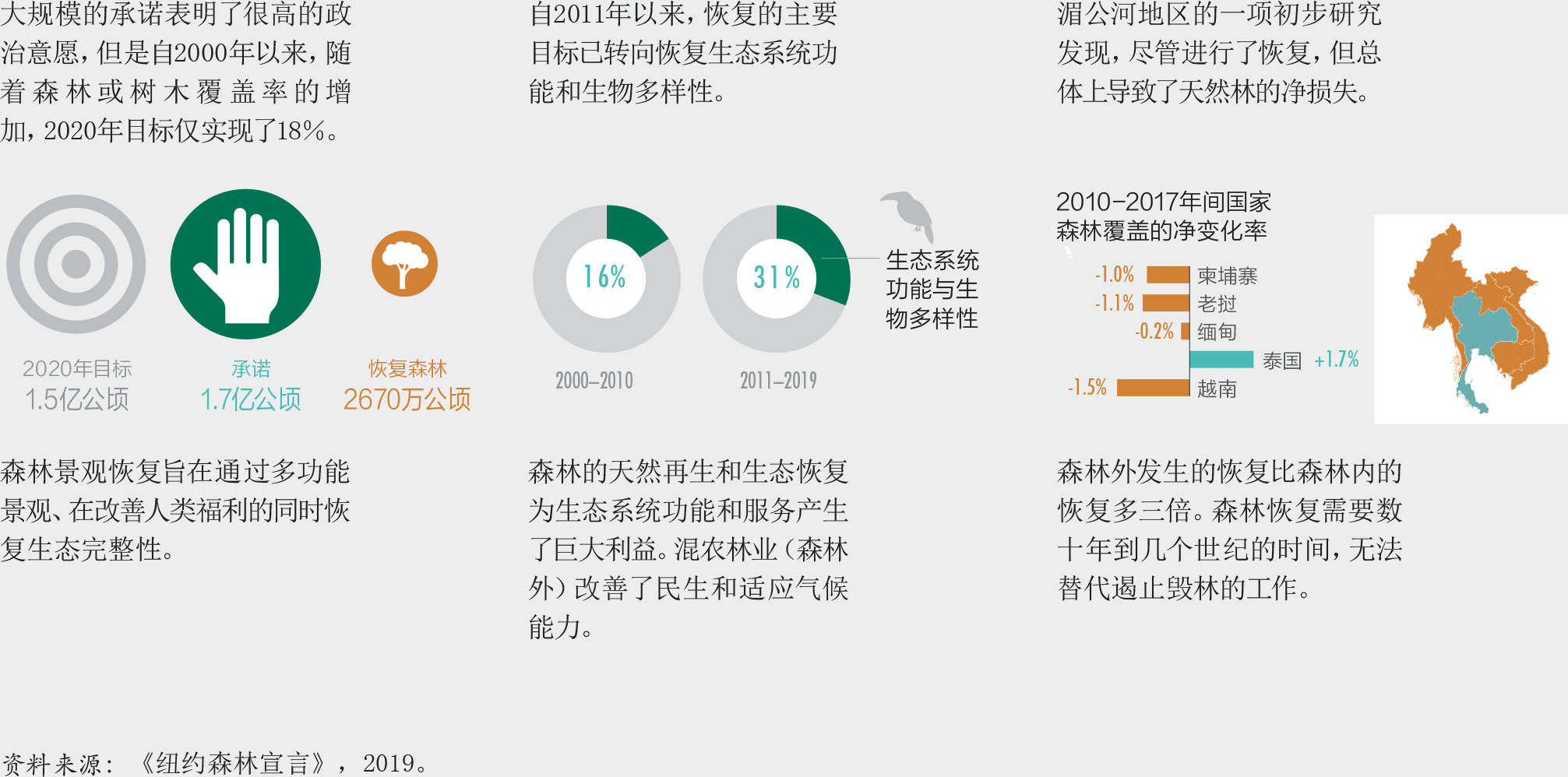
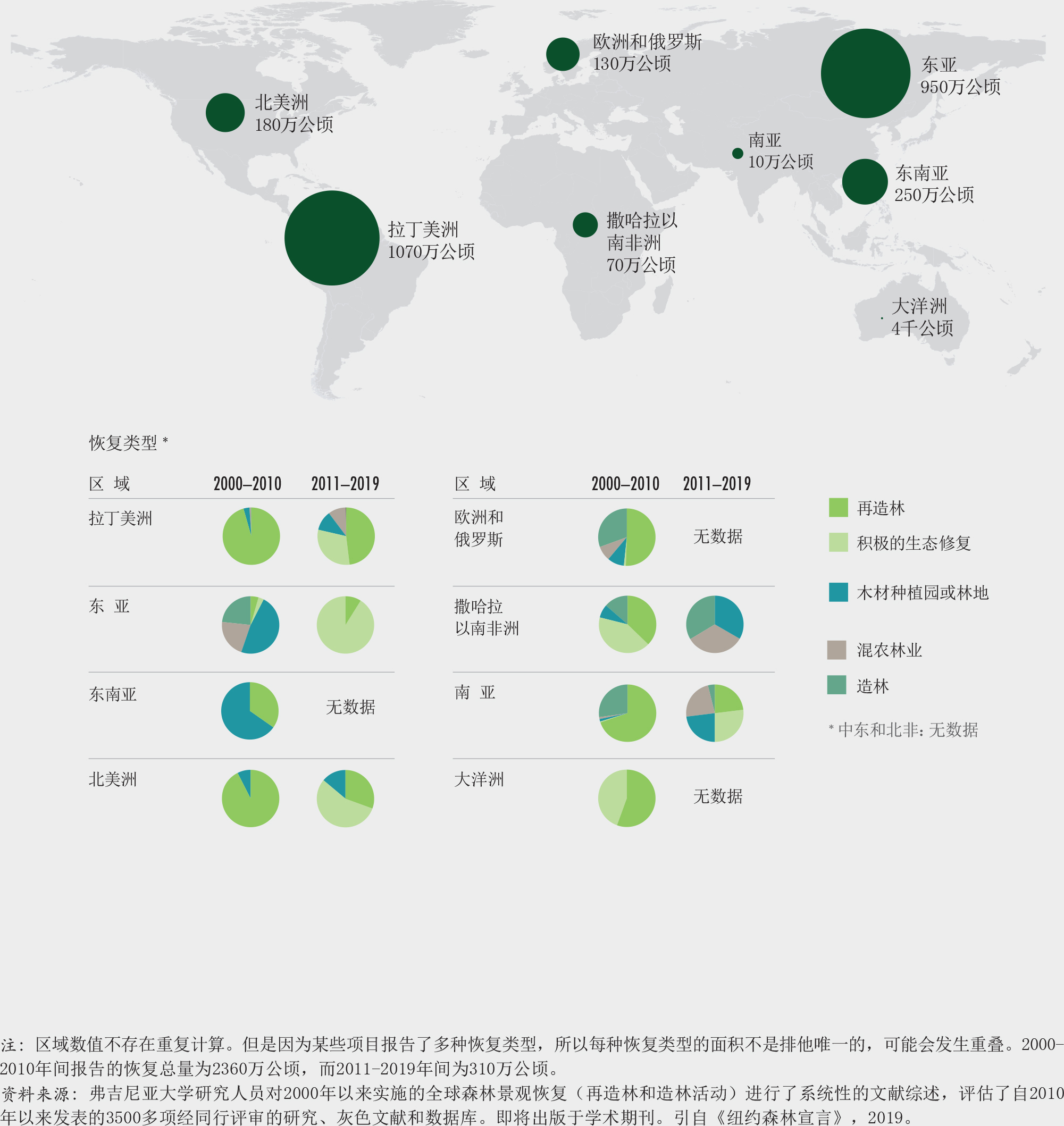
需要进行大规模的森林恢复来实现可持续发展目标,防止、遏制和扭转生物多样性的丧失。尽管已有61个国家在“波恩挑战”下做出承诺恢复总计1.70亿公顷的退化土地,然而到目前为止进展颇为缓慢。如果执行得当,森林恢复有助于恢复栖息地和生态系统、创造就业和收入,是一个有效的应对气候变化的基于自然的解决办法。于2019年3月宣布的“2021-2030联合国生态系统恢复十年”,旨在加快实现全球生态系统恢复的进程。
森林是一种基于自然的可以应对诸多可持续发展挑战的解决方案,这一观点日益得到认可,具体体现在政治意愿强化和一系列减少毁林速度和恢复退化森林生态系统的承诺之上。我们必须把握这一机会,采取果断有力行动,来防止、遏制和扭转森林及其生物多样性的丧失,造福今世后代。
联合国生物多样性十年(2011-2020)即将到期,各国政府正着手准备采用2020年后全球生物多样性框架。值此之际,本版《世界森林状况》(SOFO)考察了森林以及利用和管理森林的人类对保护和可持续利用生物多样性所做贡献(插文1)。通过聚焦森林及其生物多样性,本报告旨在补充粮农组织2019年2月发布的《世界粮食和农业生物多样性状况》(粮农组织,2019a)(插文2);生物多样性与生态系统服务政府间科学政策平台发布的《生物多样性和生态系统服务全球评估报告》(初稿发布于2019年);以及《生物多样性公约》即将发布的第五版《全球生物多样性展望》。
森林生物多样性是一个广义术语,系指森林内发现的所有生物及其在生态系统中担当的角色。因此,森林生物多样性不仅包括树木,还包括居住在森林内的各种植物、动物和微生物及其具备的遗传多样性。
森林生物多样性可以有不同层面上的含义,包括生态系统、景观、物种、种群和遗传基因。这些层面之内和之间可能产生复杂的交错关系。在生物多样性良好的森林中,这些复杂关系使生物能够适应不断变化的环境条件,并维持生态系统功能。
在第II/9号决定的附件中(《生物多样性公约》,日期不详),《生物多样性公约》缔约方大会认识到:
“森林生物多样性是数千年甚至数百万年进化过程的结果,而进化过程本身是由气候、火灾、竞争和干扰等生态因素所驱动。此外,森林生态系统的多样性(包括物理和生物学特征)导致了高度适应性,这也正是作为生物多样性一部分的森林生态系统的重要特征。在特定的森林生态系统中,生态过程的维持取决于其生物多样性的维持。”
资料来源:《生物多样性公约》,日期不详。
《世界粮食和农业生物多样性状况》(粮农组织,2019a)对与粮食和农业有关的生物多样性的所有组成部分(农作物和牲畜生产、林业、渔业和水产养殖业)的状况进行了全球评估。它有益地补充了历年来在粮食和农业遗传资源委员会的指导之下对森林、植物(农作物)、动物(牲畜)和水生物种(国家范围内的养殖物种及其野生亲缘种)遗传资源进行的全球评估(粮农组织,1997;2007;2010a;2014a;2015a;2019b)。为实现这一目的,它特别关注这些报告中未详细涉及的生物多样性类别,包括在生产系统内部及其周遭维持和调控生态系统服务的无脊椎动物、微生物和其他物种,以及作为野生食物来源的野生物种。它还侧重于生物多样性不同组成部分之间的相互作用。该出版物借鉴了91份国家报告、27个国际组织的报告、若干特别委托开展的专题研究以及更广泛的全球文献。报告概述了生物多样性对粮食和农业的各种贡献、生物多样性相关组成部分的现状和趋势以及影响其变化的驱动因素。该报告还讨论了针对粮食和农业生物多样性可持续利用和保护的做法和战略,以及相关政策、法律和体制框架的实施状况。
森林为地球大部分陆地生物多样性提供了庇护之所(MEA,2005),为80%的两栖动物、75%的鸟类和68%的哺乳动物提供了栖息地(Vié、Hilton-Taylor和Stuart,2009)。GlobalTreeSearch数据库(BGCI,2019)记录了超过60000个树种,其中20000多个树种已被列入世界自然保护联盟濒危物种红色名录,近8000个被评估为全球性受威胁物种(IUCN, 2019a)。约60%的维管植物生长于热带森林之中(见第3章)。生长于热带海岸的红树林为无数的鱼类和贝类提供了繁殖地和抚育地,吸收了可能对海草床和珊瑚礁产生不利影响的沉积物,而它们正是更多海洋物种的栖息地。
在所有气候区和所有收入水平的国家中,生活在森林里的社会群体都最直接依赖于森林生物多样性谋求生存和生计。然而,今天我们几乎所有人都与森林和/或其生物多样性的物产有着一定的联系,我们每个人都通过碳循环、水循环和营养物质循环以及与粮食生产的联系得益于生物多样性。
人类与森林及其蕴含的生物多样性之间的深厚关系由来已久,体现了人类在森林和稀树大草原中的渊源(Roberts,2019)。化石记录显示,人类使用植物至少可以追溯到大约6万年前的旧石器时代中期(Solecki,1975)。几千年来,森林内种类繁多的动植物为食物、饲料、建筑、服装、手工艺品、药品和其他日常需求提供了重要的原材料来源(Camara-Leret和Denney,2019)。至少可以追溯到查尔斯·达尔文的众多学者们,已经认识到有林地区的生态特征以及森林生物多样性对人类社会的本质、人类的分布以及人类文明史的影响。许多林产品的采伐和贸易支持了(在某些情况下甚至驱动了)人类社会在全球范围内的扩散:例如,产于南美东部沿海的巴西红木(Paubrasilia echinata)木材以及昂贵的萃取物红色染料的贸易,以及产于印尼的肉豆蔻(Myristica fragrans)贸易,均从十五世纪起对欧洲的殖民活动产生了重大的影响。
考古学和人类植物学证据均表明,人类活动对森林生态系统及其生物多样性的影响由来已久(Roosevelt等,1996;Peters,2000)(插文3)。即使在某些最偏远的森林中也是如此,例如在亚马逊河的心脏地带,某些物种的多样性和分布反映了人类引种驯化植物的悠久历史(Kareiva等,2007;Dourojeanni,2017;Levis等,2017)。例如红木(Swietenia spp.)一类珍贵的木材物种在整个热带地区的分布部分与数百年前消失的古代群落产生的生态影响相关(Vlam等,2017)。果树和其他森林食物也是如此。
塞尔瓦玛雅是伯利兹、危地马拉和墨西哥交界处低地热带雨林的一片广袤地区。其面积超过420万公顷,是一个生物多样性极其丰富的地区。除了其生物资源外,该地区还具有丰富的考古和文化价值。它是世界上最伟大的古代文明之一 — 玛雅文化 — 的摇篮。玛雅人在公元前2000年和公元900年间修建了主要城市,如蒂卡尔、埃尔米拉多尔、奇琴伊察奇琴伊察和艾克巴兰。在古典时代晚期(公元650-800年)的鼎盛时期,该地区的人口可能介于700万至1100万之间(Canuto等,2018)。
尽管拥有丰富的生物和文化资源,这些森林如今却面临着严重威胁。估计显示,在过去的25年间,仅塞尔瓦玛雅的危地马拉部分就丧失了约38%的森林,在1991-2016年间森林覆盖面积由262万公顷降至163万公顷(INAB,2019)。这主要是由于人口迅速增长、农业(种植和养殖)扩张、非法采伐和森林火灾(Blackman,2015)。这些森林损失造成了严重的环境和经济后果,包括依赖森林生存的群体和人们生计受损、水资源短缺、濒危物种的栖息地遭到破坏以及温室气体排放增加,这些都加剧了气候变化。
然而,塞尔瓦玛雅人过去曾经有过许多森林受损但又得以恢复的经历。科学证据表明,玛雅文明在“古典终结期”(公元830-950年)的衰落与气候变干有关。农业的扩张可能加剧了这一变化,导致森林覆盖的下降,进而减少了水资源的供给(Cook等,2012;Evans等,2018)。尽管由此引起的环境变化并不是造成玛雅文明衰落的唯一原因,但它似乎是一个重要因素(Turner和Sabloff,2012)。就这一点而言,一千年前发生的事情与今天正在发生的事情有着惊人的相似之处。
古代历史上的这一教训应该为当今的自然资源管理方法和政策提供启示。重要的是要平衡保护森林及其生物多样性与利用资源来改善依赖森林的当地社区和土著居民的生计这两者之间的关系。处于同一地区的危地马拉玛雅生物圈保护地的社区森林特许经营权的成功经验证明了取得这一平衡的可能性(参阅第118页的案例研究3)。在保护地中授予社区森林特许经营权的尝试提供了有力的证据,证明在条件合适的前提之下(例如适当的法规系统、强大的社区组织、技术援助、市场准入、机构支持和其他激励措施),有可能在改善民生、促进发展的同时也实现保护自然资源、维持森林覆盖和养护生物多样性。
今天,森林生物多样性仍然面临严峻挑战,一方面来自森林的过度开发,但最重要的是由于农业扩张,后者是导致毁林和森林破碎化以及相关森林生物多样性丧失的最主要的驱动力。具有讽刺意味的是,人类粮食系统的复原力及其应对未来变化的能力恰恰取决于这些生物多样性。仅举几个例子,譬如有助于防治荒漠化的适应干旱地区的灌木和树木、为农作物授粉的森林蜂类、有利于防止土壤侵蚀和沉积的山岳生态系统内根系发达的树木、以及可以提高防洪能力的红树林。森林作为粮食和药用作物的基因库在维持生物多样性方面发挥着至关重要的作用。随着气候变化对粮食系统风险的加剧,森林在固碳以及缓解气候变化中的作用日益变得举足轻重。
然而,人类对生物多样性的影响并非所有都是负面的,正如本报告展示的许多成功实例所表明的那样,这些举措是管理、保护、恢复和可持续利用森林生物多样性的成功典范。
本版《世界森林状况》并非旨在形成一部全面论述森林生物多样性的专著,而是对其现状进行更新,并就其对人类的重要意义做出总结。本报告评估了迄今为止在实现全球目标和具体目标方面取得的进展(插文4),并通过一系列旨在寻找创新做法、成功因素和双赢方案的案例研究,展示政策、行动和方法在保护和可持续发展上的有效性。
1992年生效通过的《生物多样性公约》(联合国,1992a)旨在保护生物多样性(包括森林生物多样性),可持续利用其组成部分,以及公平公正地分享基因资源带来的惠益。《2011-2020年生物多样性战略计划》(CBD,2010a)包括了到2020年要实现的20个有期限、可衡量的目标:爱知生物多样性目标。其中一些目标与森林生态系统有关。预期在2020年10月举办的《生物多样性公约》缔约方第十五次会议上将达成对新目标的共识。作为对2010年通过的《生物多样性公约》的补充,《生物多样性公约关于获取遗传资源和公正和公平分享其利用所产生惠益的名古屋议定书》(CBD,2011)对于森林和依赖森林的群体也具有重大意义。
根据《联合国气候变化框架公约》(联合国,1992b),森林在减少温室气体排放和缓解气候变化方面具有关键作用。2016年签署的《巴黎协定》(联合国,2015)第5条,通过基于成果的支付和减少毁林和森林退化所致排放以及在发展中国家保护、可持续管理森林及增强森林碳储量(REDD+)的机制,为保护包括森林在内的碳储库制定了框架。《联合国气候变化框架公约》(2011年)明确指出,增加森林碳储量的行动应“与保护天然林和生物多样性相一致”,并“用于激励天然林及其生态系统服务的保护和养护,增强其他社会和环境惠益”。作为国家自主贡献(NDC)的一部分,许多国家向《联合国气候变化框架公约》承诺将采取行动以减少因毁林和森林退化所致排放,增加森林面积,加大碳汇。
《联合国防治荒漠化公约》(UNCCD)于1992年通过(联合国,1992c)。其《2018-2030年战略框架》(UNCCD,2018)为所有利益相关者提供了实现零土地退化的框架。尽管该框架未明确提及森林生物多样性,但如预期影响4.1(可持续土地管理和防治荒漠化/土地退化有助于生物多样性的保护和可持续利用)所示,与《生物多样性公约》和《联合国气候变化框架公约》的协同增效被列为工作重点。包括再造林在内的景观恢复显然是实现这一目标的手段之一。
2015年通过的《联合国2030年可持续发展议程》和可持续发展目标(联合国大会,2015a)为2015-2030年间动员诸方努力来消除贫困、减少不平等和应对气候变化提供了框架。可持续发展目标15(“陆地生物”)与森林及其生物多样性的保护和可持续管理直接相关。
1973年签署的《濒危野生动植物种国际贸易公约》(CITES)(CITES,1983)在其附录中列出了许多依赖树木和森林的物种,对这些物种的国际贸易实行不同程度的控制。《公约》要求其183个缔约方确保所列物种的国际贸易不危害该野生物种的生存,并且确保贸易是合法、可持续和可追溯的。
于2011年12月生效的《2006国际热带木材协定》(贸发会议,2006)是一个确保被出口的非《濒危野生动植物种国际贸易公约》所列物种热带木材和木材产品来源于可持续森林的协定。
《国际重要湿地特别是水禽栖息地公约》(《拉姆萨尔公约》)(教科文组织,1971)指定将如红树林和泥炭地森林这样的森林生态系统列入保护湿地名录。《公约》还支持恢复倡议,并于2002年通过了恢复湿地的原则和准则。
第一份《2017-2030年联合国森林战略计划》(联合国, 2017a)在联合国森林论坛的主持之下产生,并于2017年经联合国大会通过。该战略计划包括了在2030年前自愿和普遍实现的六项全球森林目标和26项相关的具体目标。
《纽约森林宣言》(联合国,2017b)呼吁采取行动阻止全球森林损失,并拟定了十个与森林保护和恢复有关的目标。《宣言》于2014年在联合国气候峰会上首次签署通过,现在已有200多个背书方,包括国家政府、公司、土著和地方组织以及非政府组织(联合国,2017b)。
《森林遗传资源养护、可持续利用和开发全球行动计划》由粮食和农业遗传资源委员会在2013年(粮农组织,2014b)同意通过,确定了27个行动战略重点。
《国际植物保护公约》(粮农组织,2011a)是一项国际条约,旨在确保采取协调有效的行动,来预防和控制植物和植物产品有害生物的传入和扩散,这也正是森林健康的关键。其《2020-2030年战略框架》的启动与“2020国际植物健康年”相呼应。
《保护野生动物迁徙物种公约》(环境署,1979)为保护和可持续利用迁徙动物及其栖息地提供了一个全球平台,将迁徙动物经过的成员国汇集在一起,为在迁徙范围内采取国际协调保护措施奠定了法律基础。
随后两章论述了森林生物多样性的生物物理状况 — 生态系统(第2章)以及物种和遗传多样性(第3章)。第4章着眼于森林及其生物多样性对人类及其生计和福利的重要性。该章探讨了贫困与森林生物多样性之间的关系,以及森林资源在支持生计、粮食安全、营养和人类健康方面的社会经济作用。第5章和第6章讨论了可以确保森林能继续为地球及其所有居民的健康和福祉做出贡献的行动。第5章探讨了扭转森林流失的方法。首先回顾了毁林和森林退化的根本原因和驱动因素,然后介绍了一些致力于森林恢复的成功经验和实践。第6章着重于森林资源和生物多样性的保护和可持续利用。着眼于保护区和其他有效的基于区域的保护措施的作用;还探讨了其他鼓励可持续森林利用,支持森林地区人民的生计的管理制度。第7章强调了以整合和创新方式将这些行动结合在一起的重要性。认识到的一点是,以保护森林和社会经济发展为目的对森林进行经营管理时需要权衡取舍,而且在监测结果和采取必要后续行动时存在一定的难度。尽管存在这些挑战,但是该章还是表明协同效应是可能的,并且汇总了一系列有效的干预措施。
本章介绍有关森林生态系统状况的最新数据。这些数据来自粮农组织的《2020年全球森林资源评估》(FRA 2020)以及欧盟委员会联合研究中心(JRC)和环境规划署世界保护监测中心(UNEP-WCMC)利用卫星图像为《2020年世界森林状况》所进行的两项最新分析。本章专注于全球范围和广泛的生物群落(全球生态区)。粮农组织(2020)提供了有关区域和国家层面更多的详细信息。
由于许多森林比其他生态系统的生物多样性更为丰富,所以森林生态系统是世界生物多样性的一个关键组成部分。因此,森林覆盖的土地面积也是可持续发展目标15“陆地上的生命”的指标之一。
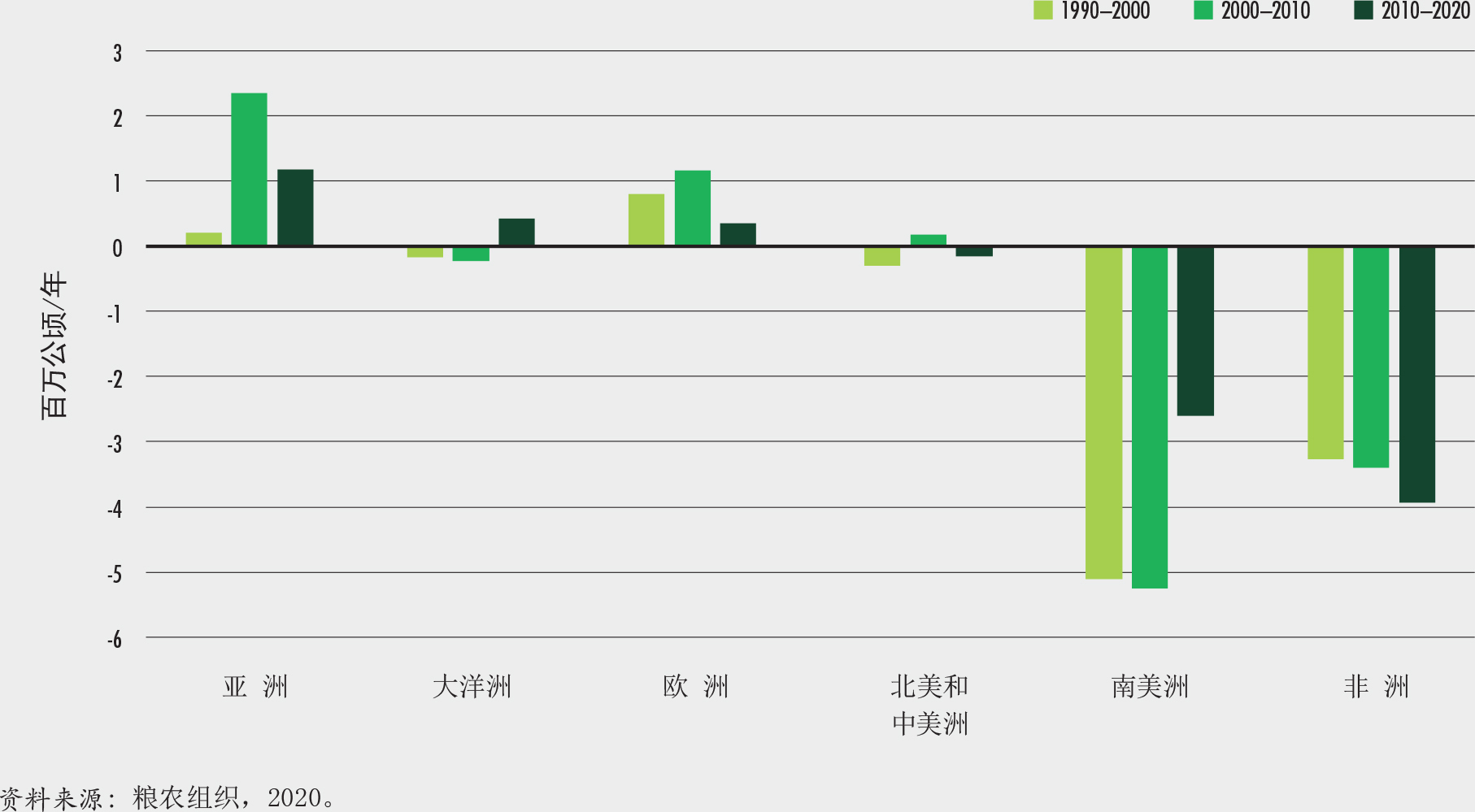
根据《2020年全球森林资源评估》,森林目前占全球土地面积的30.8%(粮农组织,2020)。森林总面积为40.6亿公顷,人均约0.5公顷,但森林并非均匀分布于全球。五个国家(俄罗斯、巴西、加拿大、美国和中国)拥有世界一半以上的森林,十个国家拥有全球三分之二(66%)的森林(图1)。
作为可持发指标15.1.1(插文5),森林占土地总面积的比例在1990-2020年间的三个十年中,从32.5%下降至30.8%。这意味着森林净损失了1.78亿公顷,约相当于利比亚的面积。然而,平均森林净损失率由1990-2000年间的每年减少784万公顷下降约40%,到2010-2020年间,每年减少474万公顷。这是森林面积在某些国家减少而在其他一些国家增加的结果(表1)(粮农组织,2020)。森林损失主要是由农业扩张引起的,而森林面积的增加则是由于森林的自然扩张(例如在废弃的农田上)或者是通过再造林(包括通过辅助自然再生)或造林。这些自然和人为的变化对森林生物多样性产生了不同的影响
▸ 可持续发展目标15.1:到2020年,根据国际协议规定的义务,保护、恢复和可持续利用陆地和内陆的淡水生态系统及其服务,特别是森林、湿地、山麓和旱地。
— SDG 15.1.1:森林面积占土地总面积的比例。
▸ 爱知生物多样性目标5:到2020年,包括森林在内的所有自然栖息地的丧失速度至少减半,并且在可行的情况下降低到接近零,同时大幅度减少退化和破碎化状况。
▸ 联合国森林战略计划目标1:通过包括保护、恢复、造林和再造林在内的森林可持续管理,扭转全球森林覆盖下降的趋势,并加大努力防止森林退化,为应对气候变化的全球努力做出贡献。
— 具体目标1.1:到2030年,全球森林面积增加3%。
▸ 《纽约森林宣言》目标1:到2020年至少使全球天然林的丧失速度减半,并力争到2030年消除天然林的丧失。
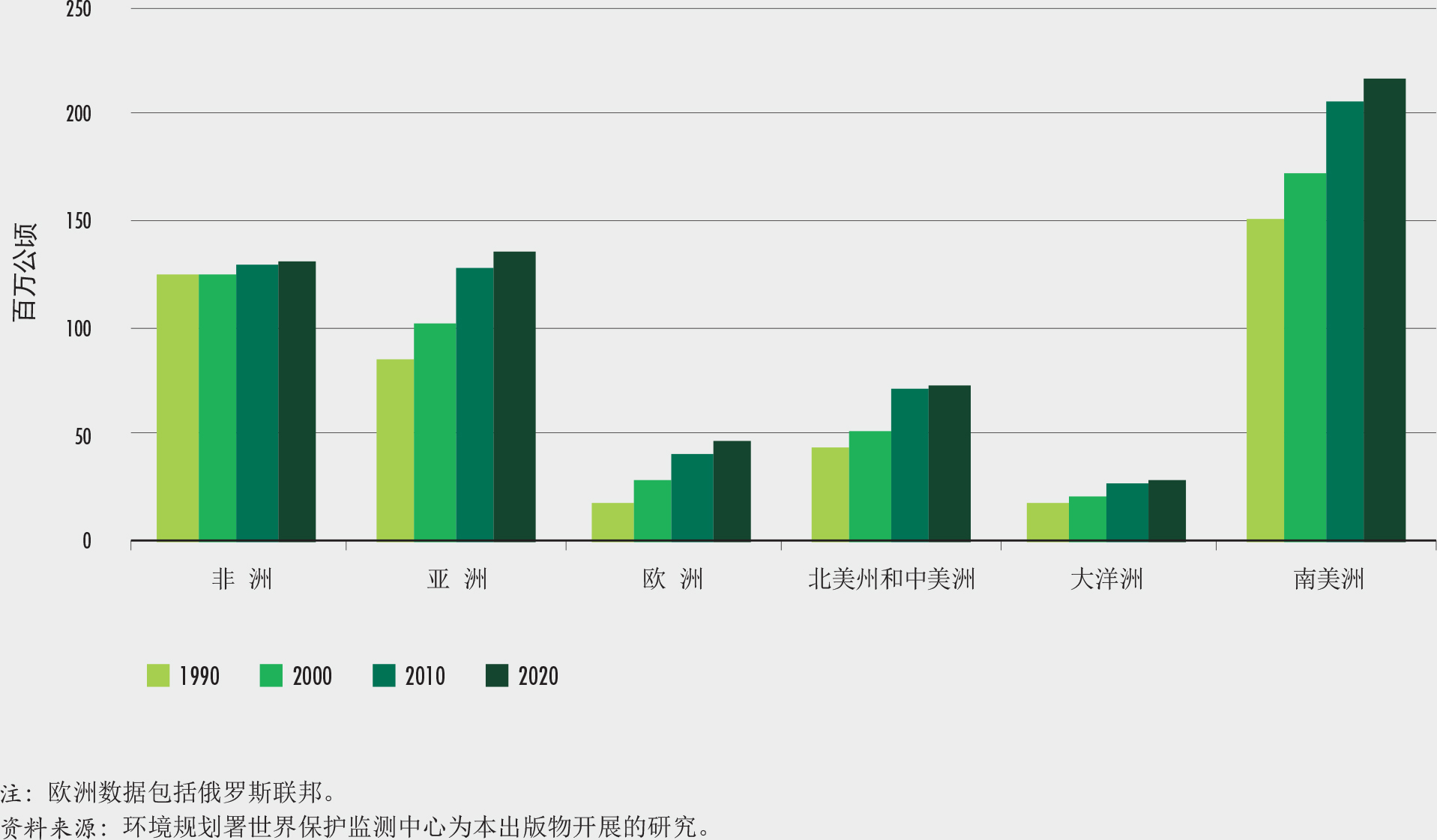
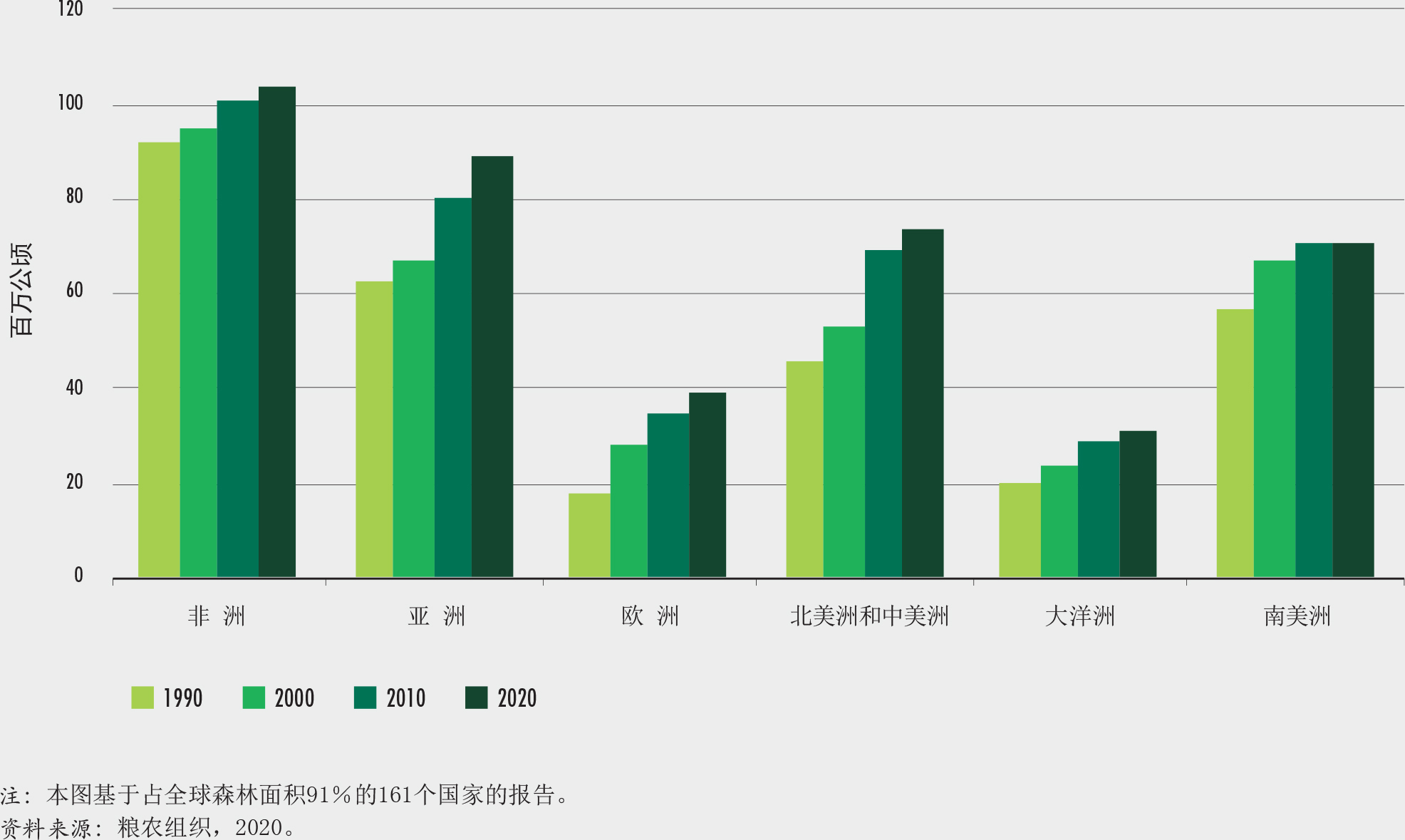
非洲在2010-2020年间的森林面积净损失最高,每年损失394万公顷。其次是南美,每年损失260万公顷(图2)。自1990年以来,非洲净损失率增加了,而南美的森林损失却大大减少了,自2010年以来的森林损失与前十年相比减少了一半以上。
2010-2020年间,亚洲森林面积净增幅最高,其次是大洋洲和欧洲。自1990年以来的每个十年,欧洲和亚洲都呈现森林面积净增,然而这两个区域自2010年以来都呈现出增幅的明显放慢。
作为《2020年全球森林资源评估》报告的一部分,要求各国报告“其他有树木覆盖的土地”,定义为“其他(意即没有被划分为森林、其他林地或内陆水域的土地)面积超过0.5公顷的土地,且树木的林冠覆盖率达到10%,并在成熟时高度能达到5米”(见插文6)。“其他有树木覆盖的土地”分为五类(表2)。少于一半的国家能够报告此参数,较少的国家能够提供其趋势走向。然而,从收到的数据来看,世界上至少有1.62亿公顷不列为森林的有树木覆盖的土地,如果参照数据之间的差别的话,有可能多达3亿公顷。唯一没有随时间增加的类别是城市中的树木。
本版《世界森林状况》报告的全球森林面积数据与其他机构所报告的数据不同,主要是因为获取信息的方法上有所不同,以及对森林的定义上存在差异。粮农组织将森林定义为树木覆盖和土地使用的结合,而其他机构则仅根据树木的覆盖来定义森林(即包括森林和《2020年全球森林资源评估》定义为“其他有树木覆盖的土地”)。仅基于中低分辨率遥感信息的数据无法区分农业生产系统(如果园、油棕种植园、咖啡种植园)中的树木覆盖和农业或城市土地使用之外土地上的树木覆盖。这意味着这些来源报告的树木覆盖总面积通常大于森林总面积。此外,根据粮农组织定义,因计划内采伐或者因受自然因素干扰而出现暂时树木缺失的土地仍被视为森林,而基于树木覆盖的遥感分析则会将这些状况解释为森林的损失。反之,如果仅基于遥感信息,有木本农作物增加的地方会被视作森林面积的增加。此外,幼林也不易被卫星检测到。报告年份也有所不同,但即使将这一点考虑在内,仅基于遥感数据的树木覆盖面积的年净变化率仍可能与森林面积的净变化有很大不同,因为后者是参考了其他辅助数据的,包括土地使用数据。
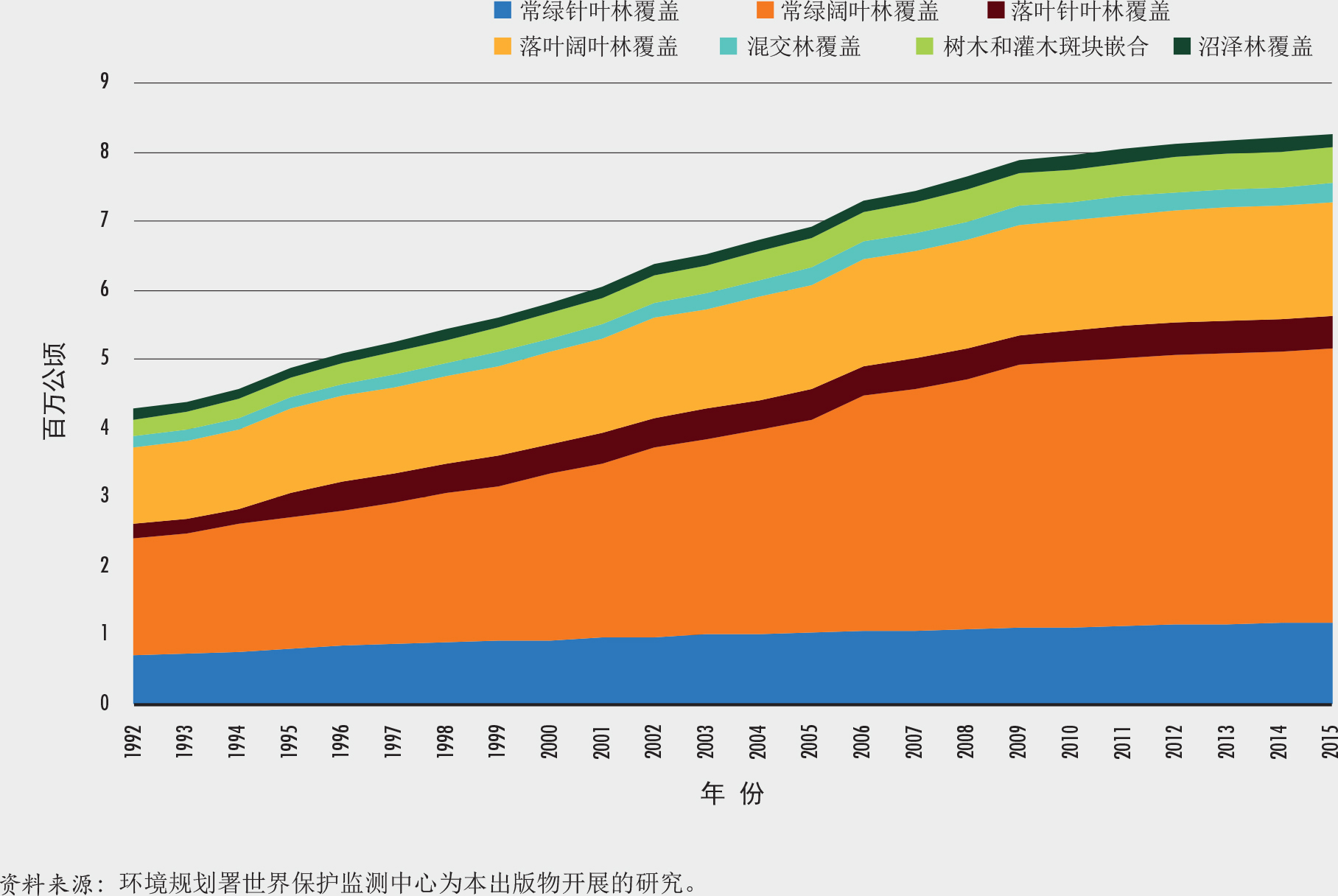
因此,尽管以上《2020年全球森林资源评估》的结果显示全球森林面积净损失率稳步下降,但《纽约森林宣言》(NYDF,2019)报告自2000年以来全球树木覆盖率下降,指的是所有类型树木的总损失(不包括一个时间段内树木覆盖的任何增加)。Song等(2018)研究了两个时间点之间面积上的差异(因而是基于净变化),得出全球树木覆盖率在1982-2016年间有所增加的结论。相反,世界保护监测中心为本版《世界森林状况》所做的一项研究(见图3)表明,在1992-2015年间总树木覆盖有所下降。
本报告力图将涉及森林覆盖的结果和涉及树木覆盖的结果区别开来。
世界保护监测中心对欧洲航天局1992-2015年分辨率为300米的年度土地面积数据进行的分析(Bontemps等,2013)表明,全球树木(包括棕榈和木本农作物)的覆盖面积在1992年约为44.2亿公顷,但到2015年已降至43.7亿公顷,减少了约5000万公顷。然而,各年份树木覆盖面积差异变化显著(图3)。树木覆盖的净变化率和规模在国家之间和森林类型之间也存在很大差异。虽然该研究中的全球树木覆盖面积与《2020年全球森林资源评估》中的森林面积和“其他有树木覆盖的土地”面积之和非常吻合,但平均净损失却要低得多,究其原因一部分是由于此期间“其他有树木覆盖的土地”的增加,另一部分则是由于评估方法不同。
《2020年全球森林资源评估》首次要求各国不仅报告不同时间点的森林总面积,并使用这些数据报告森林面积净变化,而且还要求其提供有关毁林速度的信息,即森林转换为其他土地用途或树冠覆盖率永久性降至10%这一用于定义森林的最低阈值。自1990年以来,据估计,森林面积因毁林损失了4.2亿公顷,但是自1990-2000年以来,毁林速度已显著降低。2015-2020年间,毁林速度据估计为每年1000万公顷,低于上世纪90年代每年1600万公顷。图4显示了年均毁林速度和森林扩张率,二者结合起来反映了森林面积净变化。
《2020年全球森林资源评估》将森林分为天然林(进一步细分为原始森林和次生林)和人工林(进一步细分为人工造林和其他人工林)。在全球范围内,天然林占世界森林的93%。其余7%由人工林组成(图5)。
原始森林。粮农组织将原始森林定义为原生树种的天然再生林,没有明显人类活动的迹象,生态过程也没有受到明显干扰,有时也被称为老熟林。这些森林因其丰富的生物多样性、碳储存和其他生态系统服务(包括文化和遗产价值)而具有不可替代的价值。现在,这些森林大多在热带和亚寒带地区。在《生物多样性公约》2020年后全球生物多样性框架下,对它们进行协调一致的保护是工作重点,但是需要建立在对其现状切实了解的坚实基础之上。
森林生态系统为全球大多数陆地生物多样性提供了庇护之所,尤其是原始森林,是这些生态系统特有的一些物种的家园。在亚马逊地区,一项对原始森林、次生林(此处指林龄约为14至16年的天然林)和人工林的物种丰富度和群落相似性的研究表明,这些物种中有25%是原始森林所独有,约60%的树种和藤本植物仅存在于原始森林中(Barlow等,2007)。在更加碎片化的景观中,即便物种可以在幼生林和人工林中短期生存,原始森林斑块仍然在确保物种长期生存方面起着关键作用(Watson等,2018)(插文7)。
金头狮面狨(Leontopithecus chrysomelas)仅生活在巴西巴伊亚州的大西洋雨林中。由于该地区的原始森林历经数十年的毁林后非常零散,因此根据世界自然保护联盟红色名录(IUCN,2019a),这种绢毛猴处于濒危状态,野生种群总数估计为6000至15000只。该物种可以利用保留一些老树的再生林和橡胶园;但仍需要原始森林来栖息,从而得以生存(World Land Trust, 日期不详)。
北方斑点鸮(Strix occidentalis caurina)是北美西部原始森林的标志。其森林栖息地特征为树冠茂盛、树干茂密、有残立木以及有顶端断枝的活树。尽管斑点鸮在其他类型栖息地中也筑巢、栖息和觅食,尤其是在其栖息地的南部,但它们主要依靠较老的(150至200年林龄)的多层林分,具备一定的开放空间以便在树冠下飞行(俄勒冈州鱼类和野生动物局,日期不详)。
根据《2020年全球森林资源评估》,大约三分之一(34%)的世界森林是原始森林(粮农组织,2020)。三个国家(巴西、加拿大和俄罗斯)拥有其中的一半以上(占61%)。
原始森林在全球范围内继续减少。自1990年以来,全世界原始森林减少了8100万公顷,但在过去十年中年损失率降低了一半以上。然而,由于对原始森林的测量、监测和报告存在明显的挑战(见插文8),所以这些现状和趋势的估计是基于不完整数据的。1990-2020年间,只有137个国家报告了完整的时间序列数据,这些国家合起来仅占全球森林面积的一半以上(57%)。显然还需要采取进一步工作来改善全球和国家的估算值。
粮农组织(2018a)将原始森林定义为“原生树种的天然再生林,其中没有明显可见的人类活动迹象,生态过程也没有受到明显干扰。”《生物多样性公约》(2006)使用了类似的定义:“一个不论林龄、从未被砍伐过的、在大自然的干扰和自然过程影响之下成长的森林,…也包括被土著和当地社区使用的森林,其传统生活方式与保护和可持续利用生物多样性息息相关。”这两个定义都体现了原始森林的定性特征,但没有提供可衡量的指标,为各国提供可以量化并监测其变化的手段。
由于缺乏可操作的定义和一致、易于比较的指标,当前《2020年全球森林资源评估》的国家报告存在一些不一致和偏差(Bernier等,2017)。大多数国家使用基于土地使用和/或土地面积的替代指标来推断原始森林的数据,并且这些指标不一而同。十个国家占2020年总报告的全球原始森林面积的91%,但每个国家都使用了不同的替代指标和变量,例如保护区中的森林;没有明显人工迹象的森林;基于森林地图的地理信息系统分析,没有交通网络、城市、其他可检测到的干扰现象的森林;以及基于图片的观察。一些国家报告的原始森林面积近些年来有所增加(特别是在温带和亚寒带国家),通常是由于使用了新定义或采用了新方法(粮农组织,2020)。
“完整森林景观”目前是最常用于识别原始森林的指标。Potapov等(2017)将完整的森林景观定义为“森林和相关的无树自然生态系统的无缝拼接,没有任何人类活动或栖息地破碎的迹象,并且足够维持所有本地生物多样性,物种种群规模大且丰富。”实际操作中,他们根据森林斑块的大小和构造(面积至少500平方公里,宽度至少10公里,廊道至少2公里宽)来识别此类景观,没有由于农业、木材采伐或矿物开采引起的任何改变或管理,与道路和输电线等任何基础设施之间缓冲区超过1公里。尽管这些标准也可能不适用于所有森林生物群落(另参阅第25页有关森林完整度和破碎度的讨论)。
如果仅使用遥感技术来检测完整森林景观,就有可能会漏掉某些干扰类型(如择伐),这些森林不应归为原始森林(Bernier等,2017)。一些结合了遥感、参与式制图和其他方法的监测原始森林的新兴方法和技术,可以衡量人类的改造活动和空间完整性,而这正是识别原始森林的两个最基本且可量化的特征。森林斑块的大小、空间加权平均森林密度、森林连通性是可以较容易地进行测量,并用来量化森林空间完整性的一些指标(Kapos、Lysenko和Lesslie,2002)(参阅第25页关于森林完整度和破碎度的内容)。除了这些指标之外,多维度的变化指标还可以包括某些特定的导致森林变化的人类活动,例如定居点和基础设施的开发。由于这些因素通常不一而同,因此最好开发一种区域性指标来考虑当地实际状况,但在全球范围内仍保持一致性和可比性,而不是只用单个指标或全球定义的指标(Bernier等,2017)。
粮农组织与包括《生物多样性公约》、世界保护监测中心和一些拥有大量原始森林的国家在内的合作伙伴共同协作,开展了许多旨在改进对原始森林面积及其变化进行报告的工作。
原始森林中毁林的成因不尽相同,但包括不可持续的工业用木材采伐、农业扩张以及与基础设施建设和伐木场地建设有关的林火(Potapov等,2017)。参阅第5章中更多有关导致毁林的因素相关信息。
人工林。自1990年以来,人工林面积增加了1.23亿公顷,现在覆盖达到2.94亿公顷,但自2010年以来增长速度有所放缓。大约45%的人工林(或所有森林的3%)是人工种植林,即集约经营的森林,主要由一种或两种树龄相当、本地或外来树种组成,以规则的间隔种植,并以木材生产为主要目的。其余55%的人工林,即“其他人工林”,是指林分成熟时与天然林相似的森林,包括为恢复生态系统以及保护土壤和水而造的森林。南美的人工林占所有人工林比例最大(人工林面积的99%,或森林总面积的2%);欧洲的份额最小(人工林占6%,占森林总面积的0.4%)。
在全球范围内,有44%的人工林由引进树种组成,区域差异很大(图6)。在南美洲,97%的人工林是由引进物种组成的,而在北美和中美洲只有4%的人工林是引进物种。
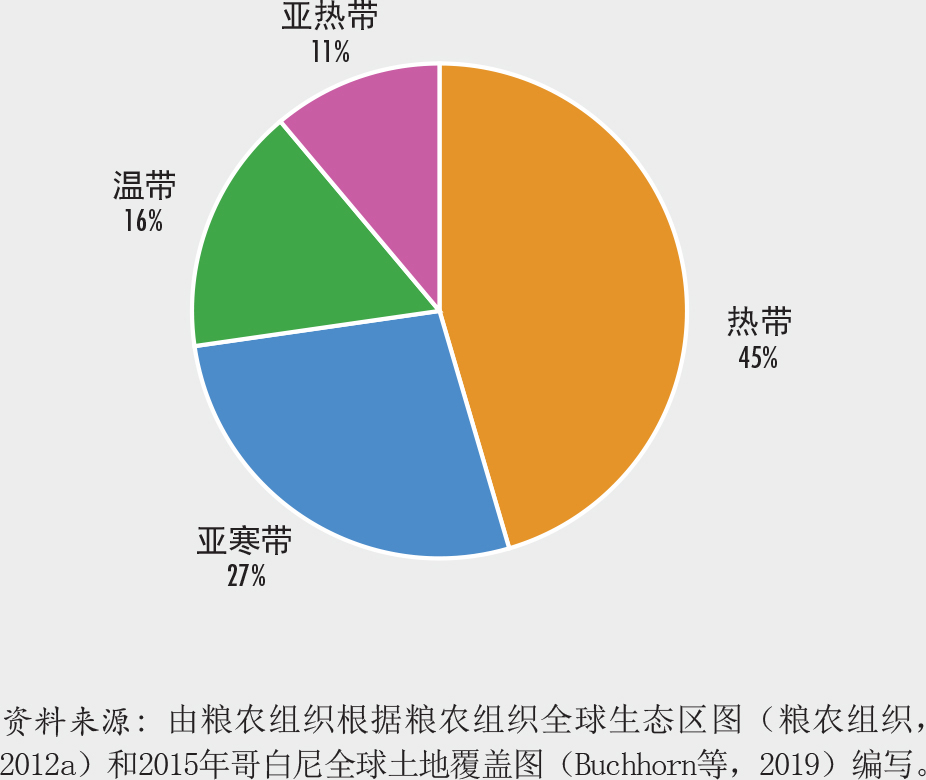
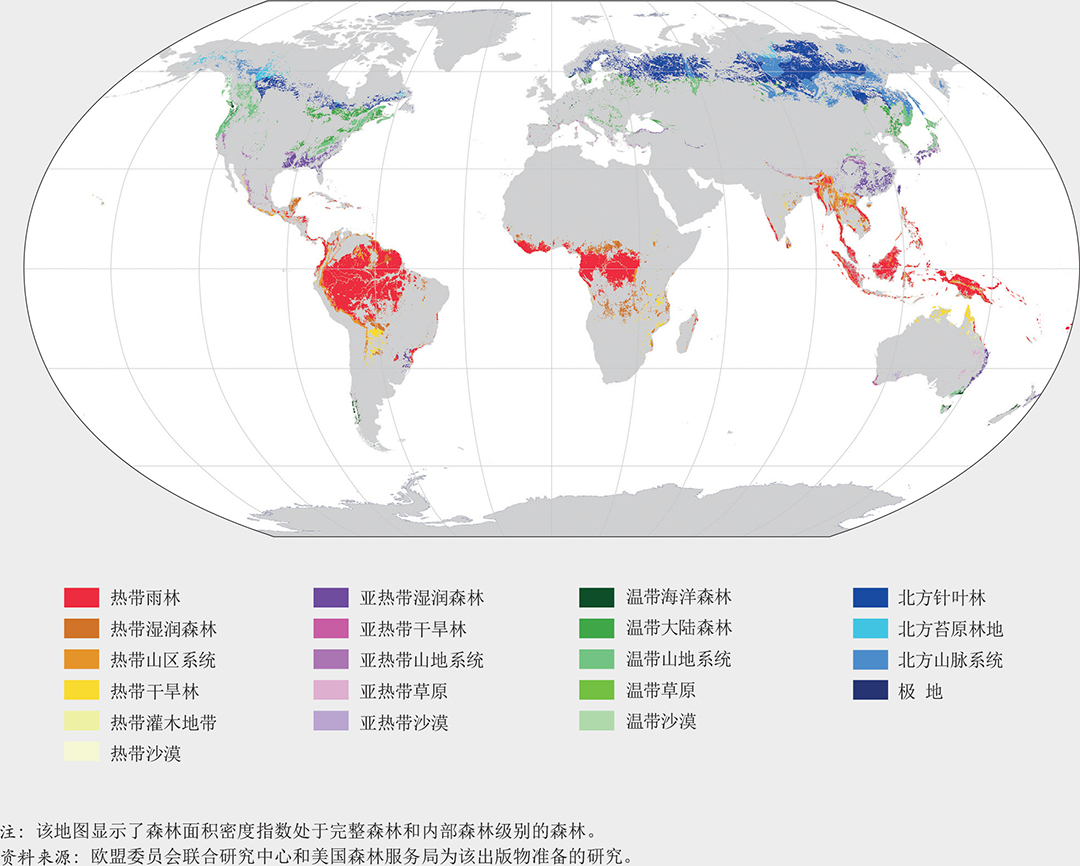
在世界范围内,有五个主要气候带:亚寒带、极地、温带、亚热带和热带。森林的最大部分(45%)位于热带地区,其次是亚寒带、温带和亚热带区(图7)。这些气候带又被进一步划分为陆地全球生态区,其中20个包含一定的森林覆盖(图8)。世界保护监测中心为《2020年世界森林状况》进行的树木覆盖变化分析发现(见第vii页),十个全球生态区在1992-2015年间的树木覆盖面积出现了净损失,十个生态区获得净增长。树木覆盖损失最大的生态区是热带雨林,涵盖了中部非洲、亚马逊河流域、印度尼西亚和巴布亚新几内亚的大部分地区,而在加拿大和俄罗斯的北方苔原林地上树木覆盖净增长最多。
从干旱地区(插文9)到湿地(插文10)和潮汐地区(插文11)都可以找到森林的踪迹。
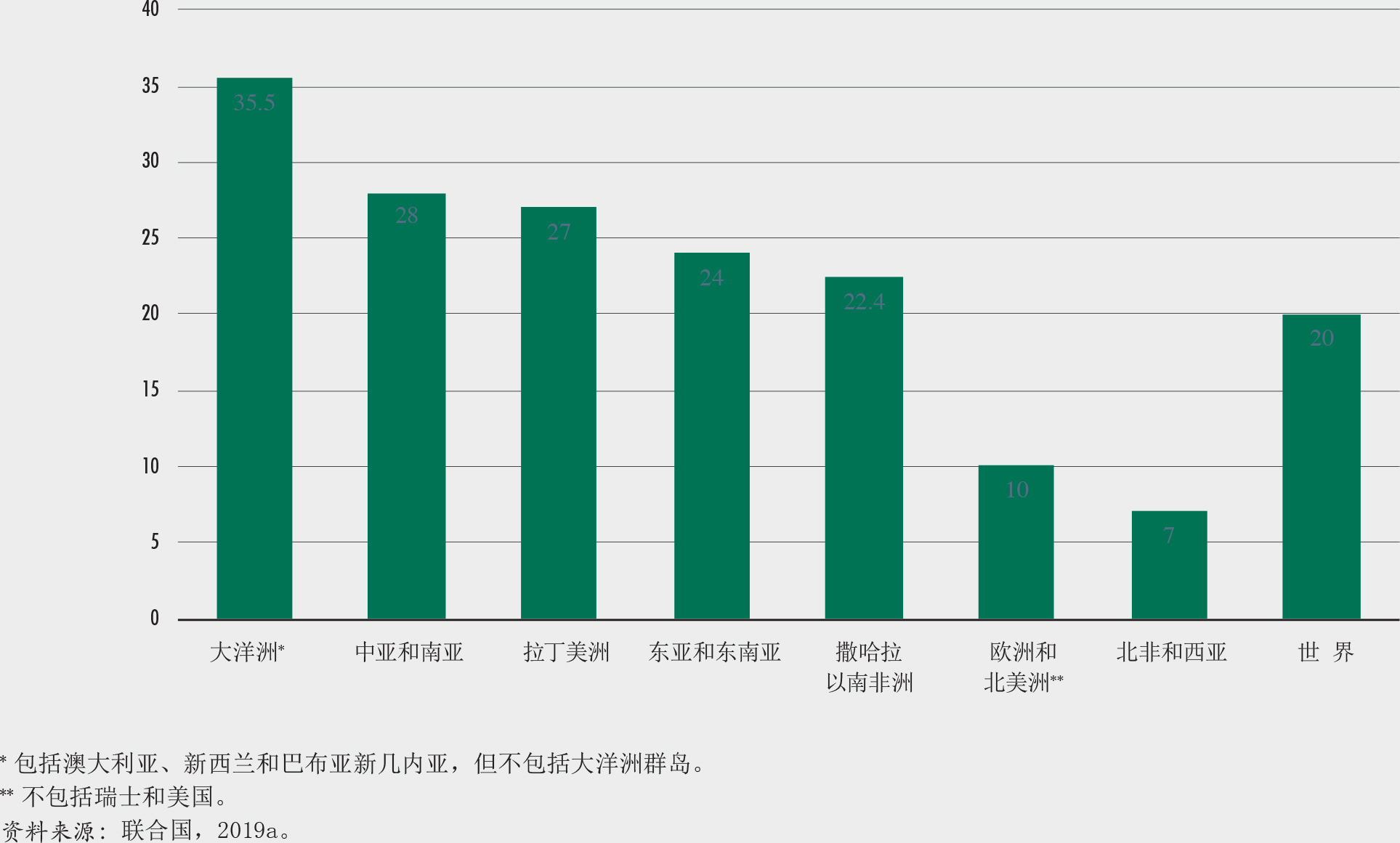
尽管湿润的热带森林生物多样性最为丰富,而干旱地区亦是具有可观经济、社会和环境价值的生物多样性丰富和多产的生态系统。在目前36个生物多样性热点地区中,干旱地区占其中7个热点地区土地面积的三分之二以上(Myers等,2000;CEPF, 2020),并在134个被列入优先保护目标陆地生态区的24个区中存在(Olson等,2015)。有超过20亿人居住在干旱地区,其中90%生活在发展中国家(MEA,2005)。这些人中有许多依靠森林和林地系统来满足其基本生存需求。尽管干旱地区具有重要的生态和社会意义,但到目前为止,关于这些地区的森林和树木覆盖的信息却相当有限。
第一次全球干旱地区评估(粮农组织,2019c)基于对被世界保护监测中心(2007)定义为干旱地区的世界上超过20万个样地的卫星图进行的影像分析。超过200名区域专家参与了该项分析。
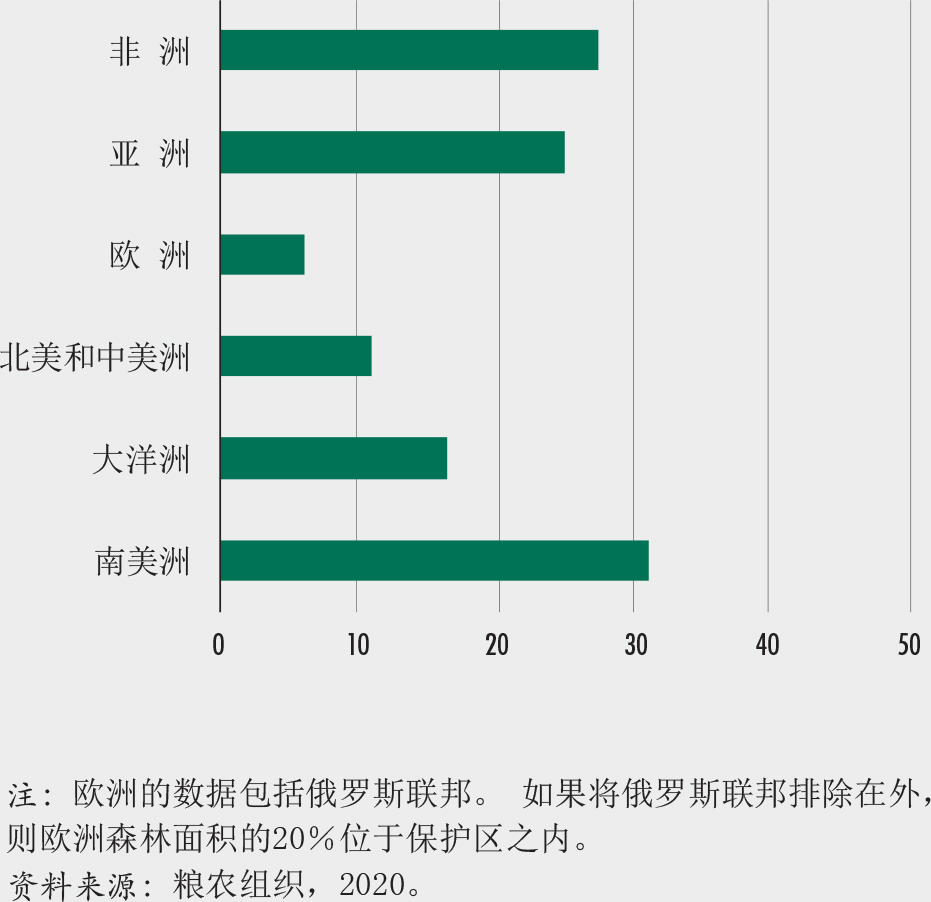
结果表明,世界干旱地区拥有11亿公顷森林,相当于世界森林面积的27%和干旱地区面积的18%。其中约51%为茂林,林冠覆盖率为70%至100%。不同区域之间的旱地森林面积差异很大(图A和B)。
许多旱地上的树木生长在森林之外。在干旱和半干旱地区,将近30%的农田和60%的建成地至少有一定程度的树木覆盖,具有大面积的牧场。处于农田中的森林以外树木在西部、中部非洲以及南亚的比例最高,其次是东非和南部非洲(图C);在这些区域,树木通常是传统混农林业或农林牧区景观和粮食系统的组成部分,起着支持农业生产、增强生态系统和当地社区应对变化的适应力的作用。
这些评估结果可以为确定对旱地森林及其种群产生威胁的新动因提供基础,确定行动优先级,关注以恢复和可持续管理这些脆弱的生态系统为目标的投资项目,这正是处于气候变化中的景观和社区提高其适应力的关键所在。用于评估的数据收集于2015年,因此可以用作监测森林、树木和土地使用随时间变化的基准值,有助于对实现可持发目标15的进展状况进行报告。
地处刚果盆地的Cuvette Centrale泥炭地被认为是世界上最大的连续热带泥炭地群,占地约1450万公顷,主要为硬木沼泽森林和以棕榈为优势树种的沼泽森林(Dargie等,2017)。该地区拥有大面积生物多样性丰富的完整雨林,并且拥有世界上密度最高的西部低地大猩猩(Gorilla gorilla gorilla)、倭黑猩猩(Pan paniscus)、黑猩猩(Pan troglodytes)和非洲森林象(Loxodonta cyclotis)。非洲侏儒鳄(Osteolaemus tetraspis)在泥炭地中产卵。这个庞大的淡水生态系统在调节水流以及为处于下游的刚果民主共和国和刚果共和国的大量居民提供食物上发挥着至关重要的作用。除了丰富的生物多样性之外,该泥炭地还蕴含至少300亿吨的碳 — 相当于两年的全球碳排放量(Dargie等,2017),这些大型碳储库增强了其生物多样性和生态系统服务的价值。
非洲侏儒鳄。
©Francesco Veronesi红树林是生长于热带和亚热带海岸线的耐盐灌木和树木,发挥着重要的环境和社会经济功能。这些功能包括提供各类木材和非木材产品,保护海岸线和珊瑚礁,以及为陆生和水生物种提供栖息地。
根据《2020年全球森林资源评估》报告,113个国家拥有红树林,估计总计1479万公顷。亚洲地区报告的红树林面积最大(555万公顷),其次是非洲(324万公顷),北美洲和中美洲(257万公顷)和南美洲(213万公顷)。大洋洲报告的红树林面积最小(130万公顷)。
40%的红树林集中在四个国家中:印度尼西亚(占总量的19%),巴西(9%),尼日利亚(7%)和墨西哥(6%)。自1990年以来,红树林面积减少了104万公顷,但在1990-2020年间减速降低了一半以上,从1990-2000年间的每年减少47000公顷降到最近十年间的每年减少21000公顷。
资料来源:粮农组织,2020。
尽管对森林退化没有公认的定义,但从更广义上来说,森林退化引起生物或经济生产力以及森林生态系统复杂性的损害或丧失,导致长期森林总效益供给的减少,效益包括木材、生物多样性和其他产品或服务。
为便于将来就与森林退化有关的相关目标进行报告(插文12),粮农组织要求各成员国报告其是否正在监测森林退化,以及使用的方法。共有58个国家作出了回应(占全球森林面积的38%),表明它们正在尝试监测森林退化的程度。但是,它们中的许多国家仅评估了一个或某几个特定要素。
▸ 可持续发展目标15.3:到2030年,防治荒漠化,恢复退化的土地和土壤,包括受荒漠化、干旱和洪水影响的土地,并努力实现无土地退化的世界。
— SDG 15.3.1:已退化土地占总土地面积的比例。
▸ 爱知生物多样性目标5:到2020年,包括森林在内的所有自然栖息地的丧失速度至少减半,并且在可行的情况下接近零,而且退化和破碎化的程度大大降低。
▸ 联合国森林战略计划目标1:通过可持续森林管理,包括保护、恢复、造林和再造林,扭转全球范围内森林覆盖下降的趋势,并加大努力防止森林退化,应对气候变化。
在本报告中,森林生态系统健康和森林破碎化作为估计森林退化状况的指标,我们对其现状和趋势进行了研究。
森林受到多种可能损害其健康和生命力的不利自然干扰的影响(如山火、病虫害、灾难性天气事件等),从而造成树木死亡或令其提供各类物品和服务的能力受损。在一些国家、地方层面和/或对特定森林物种来说,这些影响可能是毁灭性的。
森林大火。在某些生态系统中,自然山火对于维持生态系统动态平衡、生物多样性和生产力至关重要。火也是实现土地管理目标的一个重要且广泛使用的工具。大多数火灾是人为造成的,有时甚至达到难以控制的程度。每年,故意纵火和山火烧毁无数的森林和其他类型的植被。一项对2003-2012年间全球受火影响的森林面积的分析表明,每年约有6700万公顷的土地被烧毁(van Lierop等,2015)。在2015年,约9800万公顷的森林受到大火影响(粮农组织,2020)。这些大火主要发生在热带地区,约4%的森林面积受到影响。三分之二以上被烧毁的森林在南美洲和非洲。
90%的火灾得到及时控制,占总燃烧面积的10%或更少。其余10%的火灾占燃烧面积的90%。这些严重和备受瞩目的山火事件,例如2018年和2019年在澳大利亚、巴西、希腊、俄罗斯和美国(加利福尼亚州)的山火,导致人员和动物丧生、财产和基础设施的巨大破坏,造成环境和经济上不可计量的损失(包括被破坏的资源价值和灭火成本)。除非天气或可燃物状况有所改变,否则消防员对此类火灾无能为力。
未来,预计气候变化将在全球大部分地区带来更长的防火期,并引发更严重的火灾,包括以前从未发生过火灾的一些地区。尽管森林火灾无法避免,但通过采用综合林火管理和森林防火管理的措施,并充分考虑引起火灾的社会文化和生态因素,可以大大减少森林火灾的发生和减缓其产生的影响(粮农组织,2006)。
其他干扰。在2003-2012年间,除火灾以外的其他灾害影响了1.42亿公顷森林,其中包括主要在北美温带地区的虫害干扰;主要在亚洲的恶劣天气;主要在亚洲和欧洲的森林病害(van Lierop等,2015)。2015年,大约有4000万公顷的森林受到此类干扰的影响,主要在温带和亚寒带地区(粮农组织,2020)。
刚果民主共和国
洛马科森林为赤道上的一个社区保护区,它是Cuvette Centrale泥炭地的一部分。.
外来入侵物种(非本地害虫、病原体、脊椎动物和植物)以及本地病虫害的爆发对全球天然林和人工林的健康、可持续性和生产力构成越来越大的威胁(插文13)。仅森林虫害的爆发每年就对约3500万公顷的森林构成破坏(粮农组织,2010b)。现在,入侵的动植物物种被认为是造成生物多样性丧失的最重要原因之一,尤其是在许多岛屿国家(《生物多样性公约》,2009)。但是,除了某些发达国家以外,几乎没有关于入侵物种总影响的量化数据。
日益增加的国际贸易和人类流动性加剧了将动植物物种引入新环境从而变成入侵物种的状况,这一情形受气候变化的影响也变得更为频繁。例如,黄杨斑蛾(Cydalima perspectalis)导致伊朗和高加索地区特有的黄杨木(Buxus colchica)森林枯死,源于东亚的真菌白蜡鞘孢菌(Hymenoscyphus fraxineus)引起了英国白蜡树枯梢病。气候变化和各年气候波动通常与不良的森林管理方式(如改变森林构成和多样性)共同作用,对本地和引入的害虫和病原体产生强烈的影响,尤其是对其生物习性(如发育更快)和行为(如对宿主的喜好)产生的影响。来自较高的温度、恶劣和极端的天气事件以及干旱的压力会导致树木活力降低,使其在本地和外来病虫害爆发时更容易受到损害。例如,本地树皮甲虫的爆发导致中美洲、欧洲和北美洲数百万公顷松树林的死亡,这与气候变化、极端天气事件的影响以及在某些情况下森林管理方式的不当有关(Billings等,2004;Bentz等,2010;Hlásny等,2019)。
增强森林和森林生态系统对病虫害和入侵物种侵袭的抵御能力,需要协调国家、区域和全球在预防、早期发现、及早行动、采取植物检疫措施和提高公众意识上的活动。它还需要可持续的森林管理措施,既要减少森林对气候变化影响的脆弱性,又要考虑到生物多样性的保护和可持续利用。
在过去的一个世纪,森林破碎化(连续生境被分为更小、更孤立的碎片)已深刻改变了森林的特征和连续性,并造成了严重的生物多样性丧失(Haddad等,2015)。了解森林破碎化的程度、原因和后果对于保护森林生物多样性和生态系统功能至关重要(见插文14)。
森林的破碎化导致栖息地形态的改变、森林面积和连通性的丧失、森林斑块间隔的增加以及森林边缘与人类使用土地接触的加大(见图A)。森林孔洞化(即完整的森林斑块中引入孔洞)是碎片化的主要组成部分之一。森林孔洞化经常伴随着修建道路而来,导致不受干扰的核心森林栖息地区域大大减少。森林碎片化引发了其余森林碎片在结构和功能上的长期变化,对生境和森林生态系统服务产生了影响(Lindenmayer和Fischer,2006;Hermosilla等,2019)。
森林破碎化有可能是因自然环境变化和干扰(气候、地质变化进程、自然灾害、山火和病虫害)引起,这些变化可能导致森林被分裂成更小的斑块;也有可能是因人为因素引起,例如掠夺性森林采伐(不当的森林采伐或薪柴采集),由于农业扩张而引起的土地使用变化、转为人工种植林地、转为牧场、新增移民、城市化和基础设施建设。森林碎片化通常发生于从森林转化为其他用地的第一阶段。
森林破碎化过程改变了景观系统的组成、形态和功能,通常意味着栖息地遭到破坏或孤立。许多研究表明,尽管具体的反应会因物种和森林类型而异,但栖息地的长期破碎化,尤其是森林栖息地的破碎化,严重地影响了生物多样性和生态系统的进程(Skole和Tucker,1993;Pereira等,2010)。碎片化影响从基因到生态系统层面几乎所有的生态过程,并影响动植物种群的组成和动态。它也可能增加牲畜和野生动物之间的接触,从而加大疾病传染的风险。虽然属数、多生境、边缘或入侵物种的数量可能会增加(Laurance等,2006)(见第3章插文18中关于森林授粉者的内容),但森林破碎化大多会降低物种丰富度(Turner,1996;Zhu等,2004)。它减少养分的留存,影响营养动态平衡,并且在更孤立的碎片森林中改变动物的活动。研究表明,减少森林斑块的面积和增加斑块隔离度,将鸟类、哺乳动物、昆虫和植物的丰富度降低了20%至75%,影响了如种子传播等生态功能,进而影响了森林结构,导致如固碳、控制水土流失、授粉和养分循环等生态系统服务的减少(Haddad等,2015)。
欧盟委员会联合研究中心为本报告近期进行的一项空间分析,利用卫星遥感技术来识别出最完整和连通性最好的森林以及那些破碎化最严重的森林。分析是在全球范围内进行的,同时也对15个全球生态区中拥有森林超过世界森林面积1%的生态区进行了分析。
研究者将两个森林破碎度指标作为属性变量绘于2015年全球哥白尼土地覆盖地图之上(Buchhorn等,2019),然后与粮农组织全球生态区域地图进行叠加(见图7)。在此过程中,研究者尝试将油棕榈和木本农作物的面积排除出去以减少误差。第一个指标称为核算,用于评估森林斑块的大小和分布,即与其他森林面积分隔至少100米的单独森林区域(Vogt,2019a)(图9和10)。第二个指标是森林面积密度,它衡量了在指定邻域内森林像素的比例(Vogt,2019b)(图11至图13)。森林面积密度高表明森林连通性高、森林面积紧凑和森林破碎度低,而森林面积密度低则表明森林斑块被隔离,形成森林孔洞,且通常高度破碎。
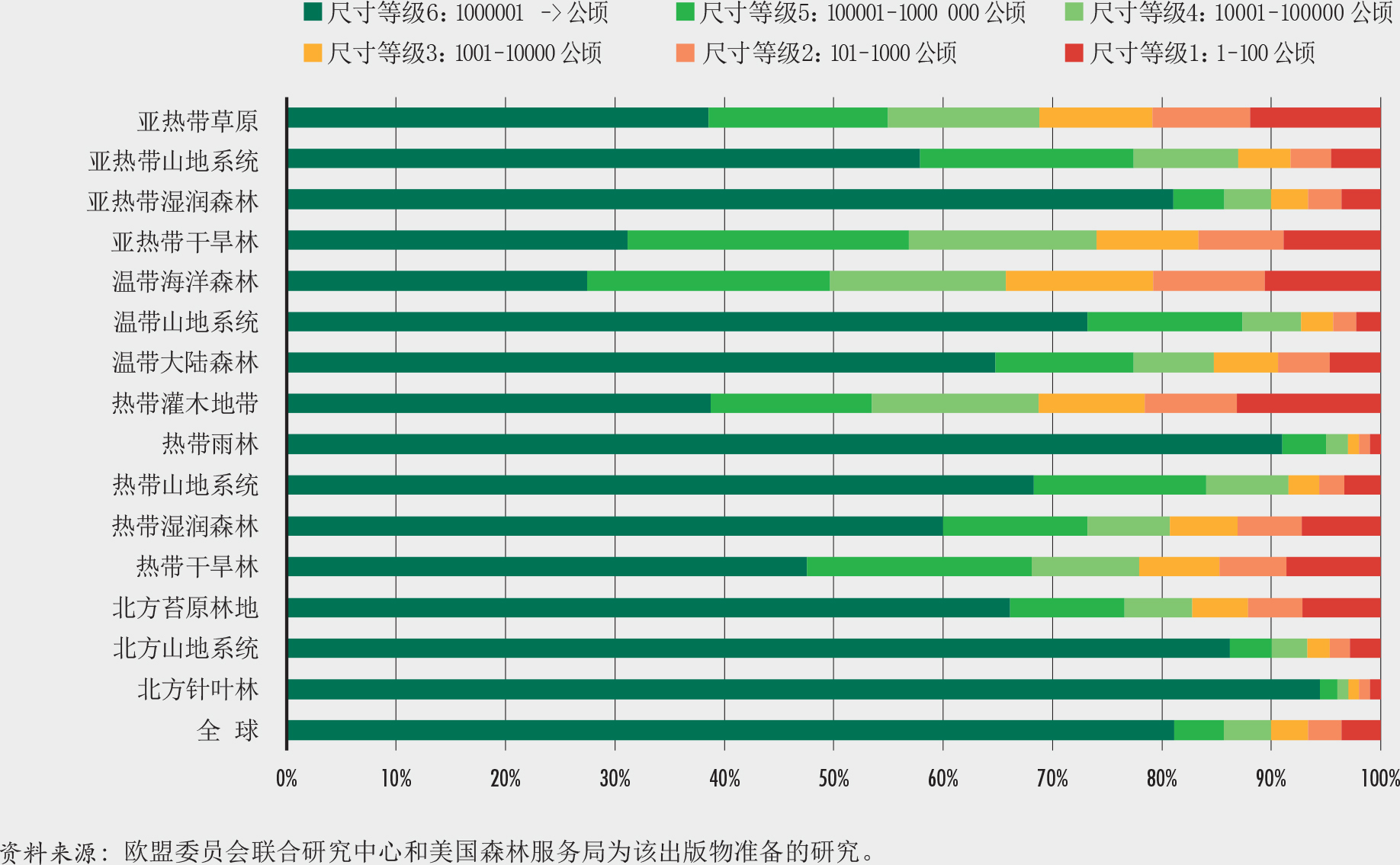
该研究发现全球3480万个森林斑块,面积从1公顷(在地图上为1个像素)到6.8亿公顷不等。全世界约80%的森林处于单个面积超过100万公顷的森林中;对于所有类型的森林来说,这种规模的森林均占其森林面积的25%以上(图9)。然而,这样的森林斑块只有149个,这意味着大多数的世界森林面积集中于极少数的地方。世界上其余的森林分散、且每个面积相对较小。
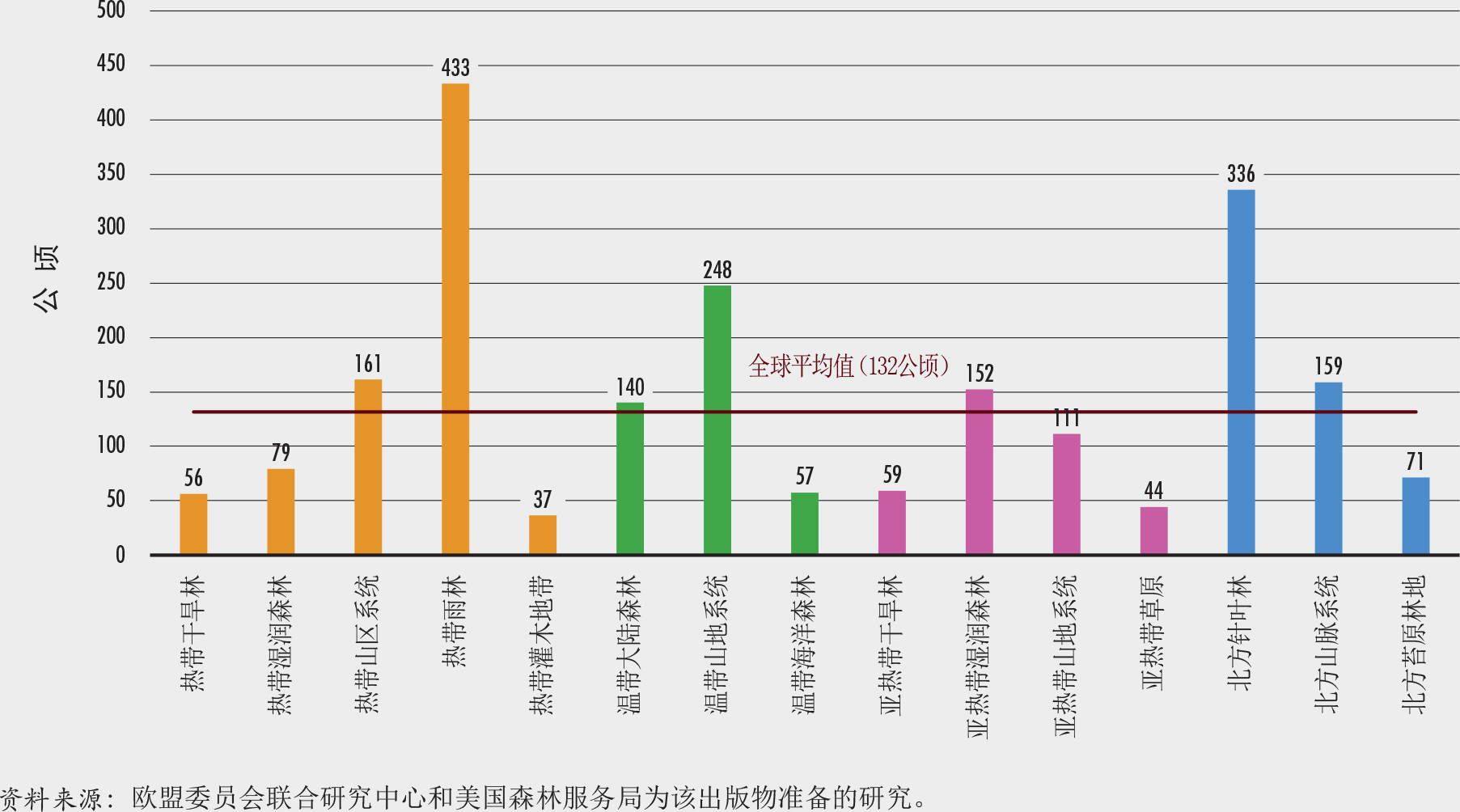
约有3470万个小于1000公顷的森林(占森林总数的99.8%),合计占全球森林面积的7%。所有森林斑块的平均面积仅为132公顷,但森林斑块平均尺寸在生态区之间差异很大(图10)。北部针叶林和热带雨林地区的森林平均斑块面积最大。
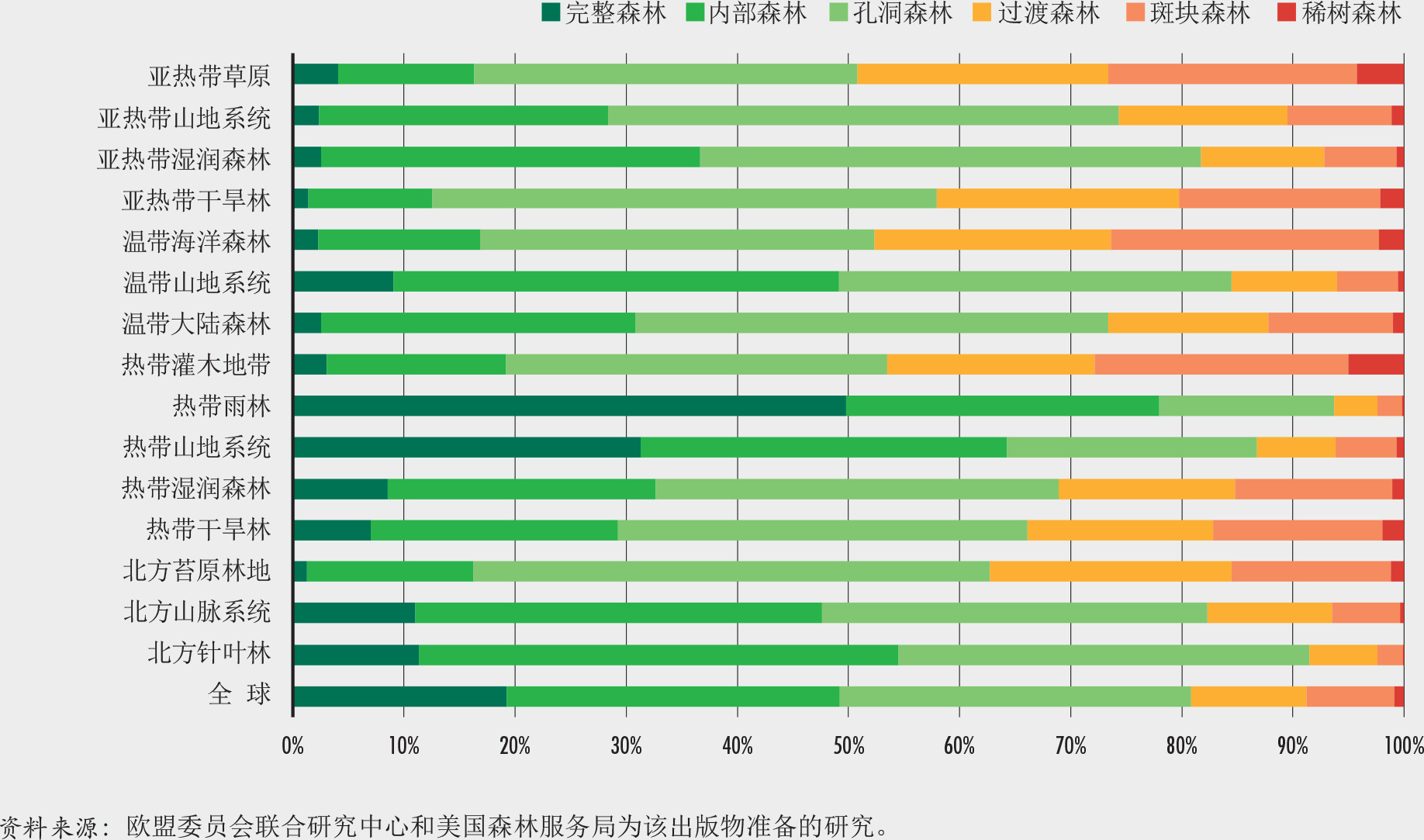
全球几乎一半的森林面积(49%)属于两个最高的森林面积密度类别(完整森林和内部森林),因此具有较高的完整度(图12和14)。在森林面积密度谱的另一极端,世界上9%的森林属于稀树状况和斑块森林类,几乎没有连通性,可以认为是严重破碎的(图12和15)。
哪里的森林最完整?热带雨林和北方针叶林(森林最多的生态区)是破碎化最少、最完整的森林生态系统。这些区域中超过90%的森林面积在面积超过100万公顷的斑块中,其森林斑块面积远大于全球平均水平(图9和10)。在这些地区中,只有不到2%的森林属于稀树状况和斑块森林类别,而超过50%的属于内部和完整森林类别(图12)。这些生态系统的特点是难以进入和人口密度低。
剩余的热带雨林中有一半属于完整森林,并且94%毗连度良好。亚马逊河流域和刚果河流域的森林斑块破碎化最少,且毗连度最高(图14)。但是,这些地区土地使用上的变化正在引起迅速的变化。由于这些森林具有独特生物多样性,因此需要特别注意森林保护和可持续管理。
在北方针叶林生物群落中,11%的森林是完整的,主要分布在加拿大和俄罗斯。北方森林破碎化主要与自然干扰(山火和虫害爆发)有关。日益严重的、与全球变暖有关的山火状况可能会增加北方森林的破碎化程度(Walker等,2019)。
处于亚寒带、温带和热带气候中的山区系统也是可达性有限且人口密度低的生物群落,而且这些生物群落明显没有其他生态区森林破碎片程度严重。它们的平均森林斑块面积大于全球平均水平(图10),仅6%属稀树和斑块森林类,40%以上属完整森林和内部森林类(图12)。这些生物群落的森林完整性也可能与这些区域中存在大量为保护水源和避免土地被侵蚀而设立的保护区有关。破碎化程度低的山区森林包括温带的北美山地森林(阿巴拉契亚山脉、喀斯喀特山脉),亚寒带俄罗斯森林(乌拉尔山脉、斯塔诺夫山脉和锡霍特山脉,有西伯利亚虎等濒危物种栖息)和中部非洲湖区的热带山脉(物种极其丰富,是大部分山地大猩猩种群的栖息地)。不幸的是,由于人口增加的压力,这些森林中的一些目前正面临边缘受到侵蚀和破碎化的高风险。
最破碎的森林在哪里?森林面积有限(不到土地总面积的三分之一)的生态区,例如热带灌木地带、亚热带草原、亚热带干旱林和温带阔叶森林,具有最高的破碎度和最低的平均森林面积密度(图10和13)。这些区域的平均斑块面积小于60公顷,而相当一部分(约占森林面积20%)小于1000公顷(图9和10);有20%的森林属稀树和斑块森林类,有不到20%的森林属完整森林和内部森林(图12)。尽管森林破碎状况对这些生态区中的一些区来说是自然形成的(如亚热带草原),但对其他生态区来说却是过去土地使用变化和森林经营方式的结果。
亚寒带苔原林地、热带干旱林和热带湿润森林生态区的森林覆盖更为显著(占总土地面积的40%以上),但平均斑块面积远小于全球平均水平(图9和10),并且超过30%属稀有、斑块和过渡林类别(图12)。这些生物群落少于30%的森林属完整和被其他森林包围类,而这一比例在亚寒带苔原林地中只有16%。
亚寒带苔原林地的森林破碎主要是自然条件和干扰(气候、山火和害虫)的结果。相反,热带干燥和湿润的森林,例如巴西的Cerrado森林、南美的查科(Gran Chaco)森林、南部非洲的Miombo林地以及印度和湄公河地区的热带干旱林,已经受到土地使用变动的影响。这些森林在生物多样性和为人类提供生计方面都非常重要,但是在这些生态区中只剩下很少的连续森林。
一旦森林被破碎化,就很难扭转这种状况,特别是就生物多样性的丧失而言。需要通过包括营建廊道、缓冲带或落脚石等恢复手段来重新连接森林碎片(参阅第5章:扭转毁林和森林退化)。
正如第2.1节森林面积的现状和趋势所示,人类在扭转全球森林覆盖流失方面取得了一些进展,森林面积的净损失从上世纪90年代的年均784万公顷降低到了2010-2020年间每年474万公顷(表1)。然而,我们还没有步入正轨可以顺利实现《联合国森林战略计划》(联合国,2017)设立的到2030年全球森林面积增加3%(相对于2015年)这一目标。
在过去30年中,天然再生林面积减少了7%(3.01亿公顷)(粮农组织,2020)。尽管天然再生林的流失率一直在下降(图16),但仍不足以实现爱知目标5和《纽约森林宣言》目标1,到2020年至少将全球天然林的流失率减半(相对至2010年)(插文5)。
尽管欧盟委员会联合研究中心的森林破碎化研究并未涉及随时间变化的趋势,但从毁林的模式来看,许多国家森林破碎化状况正在加剧。稍微值得欣慰的是,122个国家承诺将设立土地退化零增长的目标,而80多个国家已经设定了该目标(《防治荒漠化公约》,2019a)。
构成森林的不仅是树木,而且还包括栖息于土壤、林下和森林冠层中的各类动植物。据估计,地球上物种总数为300万至1亿(May,2010)。2011年的一项研究估计该数字约为870万(±130万),包括650万种陆生和220万种海生(Mora等,2011);而IPBES(2019a)则估计该数字为约800万,其中590万种为陆生。虽然广为报道森林为80%陆生动植物提供生境,但鉴于对生物多样性了解的不断变化,这样精确度的估计不太可能是精确的。
热带湿润森林是全球生物多样性极为重要的储库。比如,源自单个树种的1200种甲虫(Erwin,1982),1公顷森林里的365个树种(Valencia、Balslev和Paz y Miño,1994),0.1公顷森林里的365个植物物种(Gentry和Dodson, 1987),以及据估计世界上一半的物种存在于其陆地面积的6-7%之上(Dirzo和Raven, 2003)。热带和亚热带森林(干旱和潮湿)蕴含十个生物多样性热点地带,特有高地陆生脊椎动物总数最多,濒危物种数量最多(Mittermeier,2004;Mittermeier等,2011,引自IPBES,2019b)。
因此,虽然树木是森林的重要组成部分,树木的多样性可以表明总体的多样性,但还有许多其他方法可以确定森林生物多样性的显著性。本章在探讨物种和遗传水平上实现与保护森林生物多样性有关的重要目标的进展时,考察其中的一些方面(插文15)。
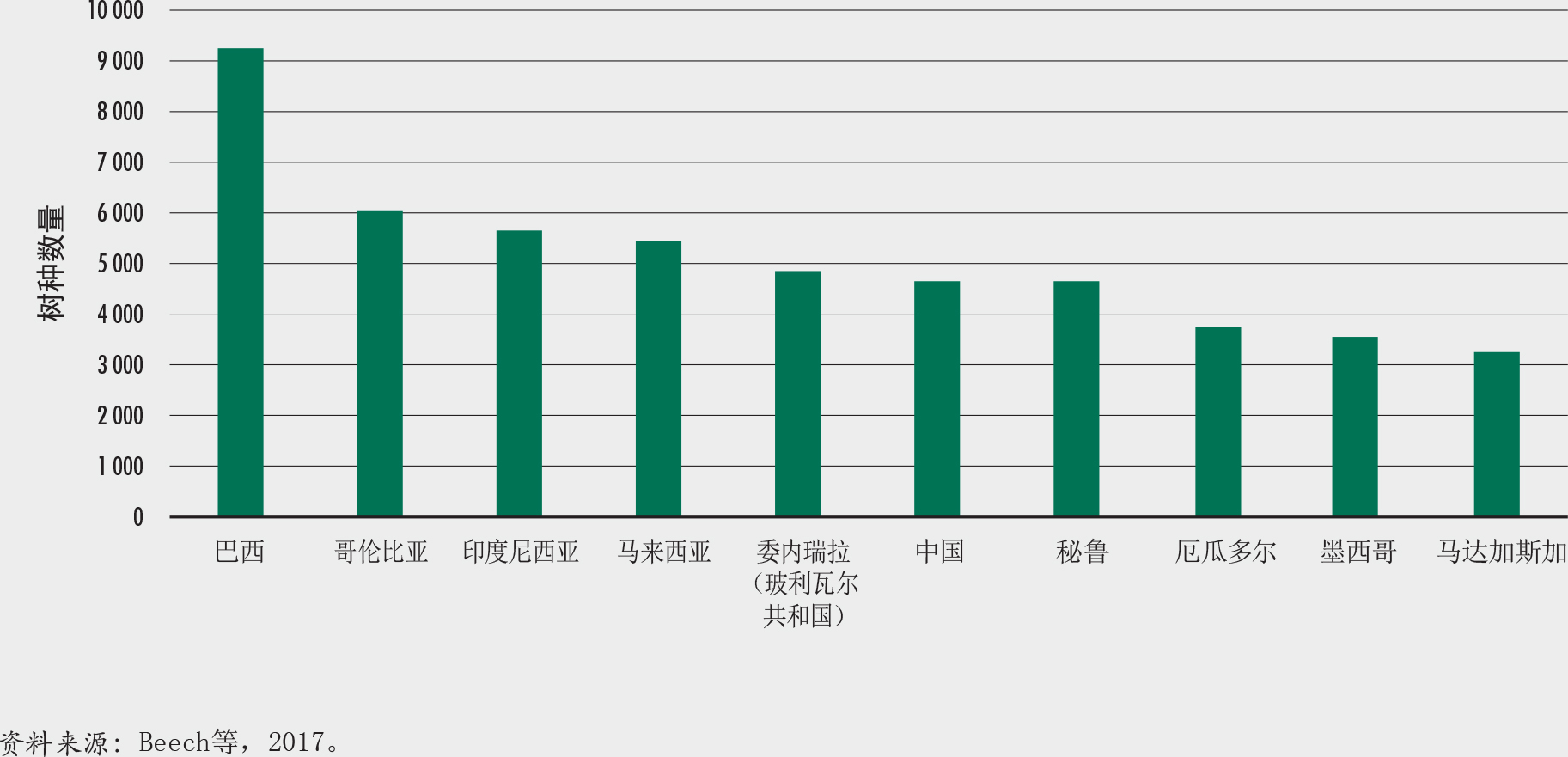
保护联盟GlobalTreeSearch数据库(BGCI, 2020)汇集了60082个树种。该数字包括棕榈和森林中不常见的许多木本农作物(例如果树、咖啡和油棕)。
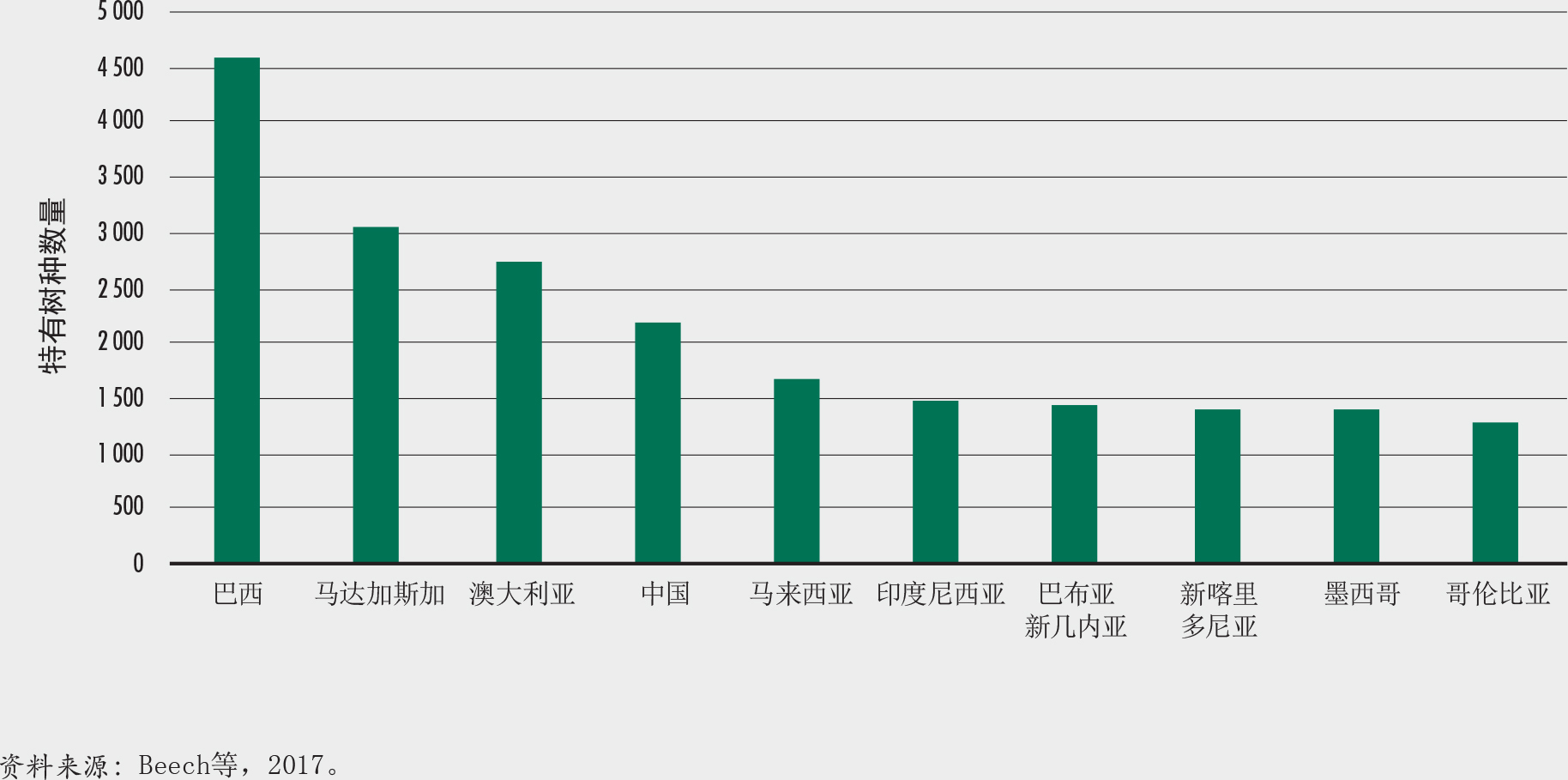
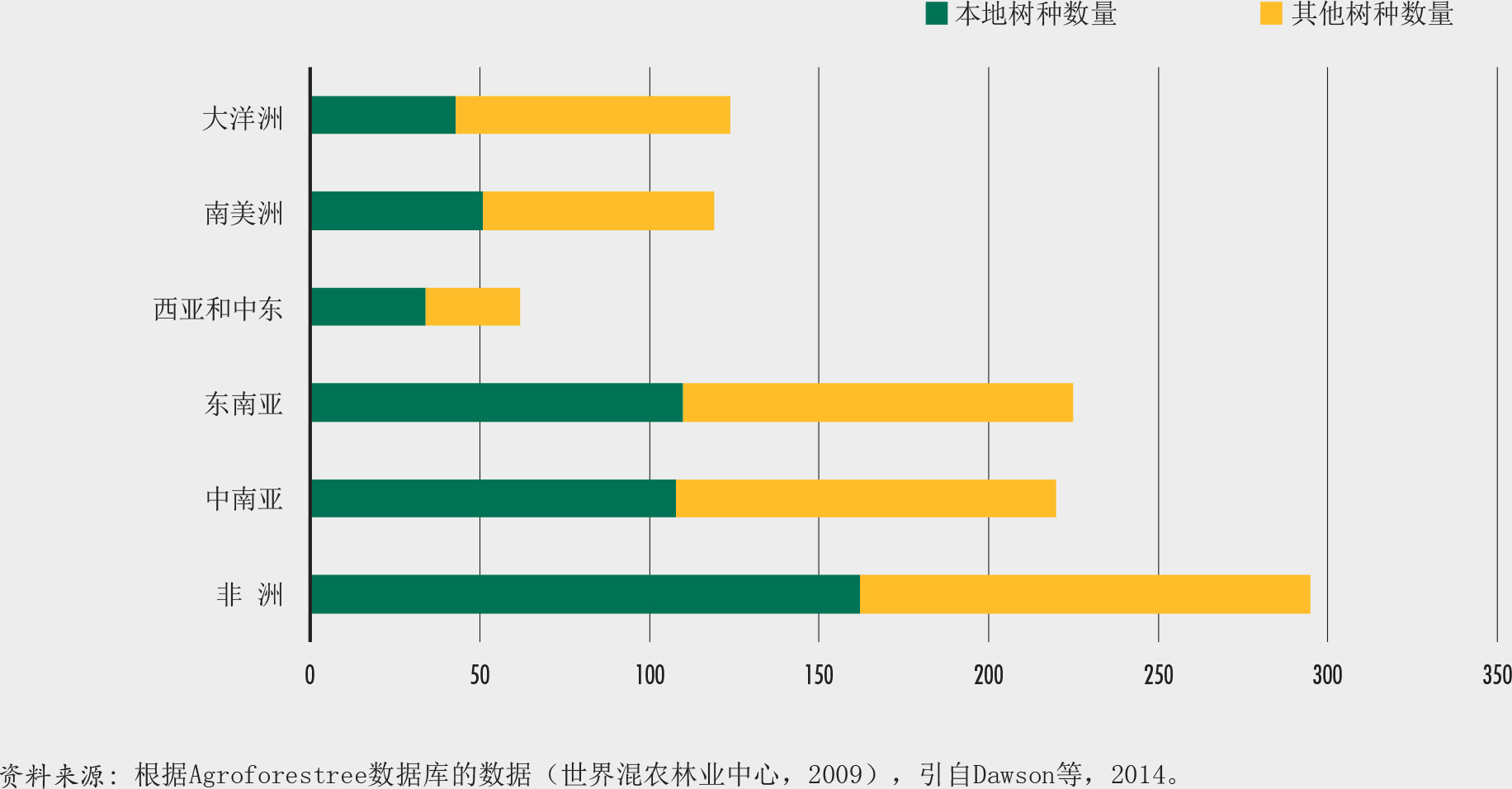
将近一半(45%)的树木种类来自10个科。树木种类最多的三个科是豆科、茜草科和桃金娘科。巴西、哥伦比亚和印度尼西亚是树木种类最多的国家(图17)。国家特有树种最多的国家体现了植物多样性更广的趋势(澳大利亚、巴西和中国),或反映出岛屿孤立导致物种增加的状况(印度尼西亚、马达加斯加和巴布亚新几内亚)(图18)。所有树种中近58%是单一国家特有物种(Beech等,2017)。
▸ 爱知生物多样性目标12:到2020年,防止了已知濒危物种免遭灭绝,且其保护状况(尤其是其中减少最严重的物种的保护状况)得到改善和维持。
▸ 爱知生物多样性目标13:到2020年,保持了栽培植物和养殖和驯养动物及野生亲缘物种,包括其他社会经济以及文化上宝贵的物种的遗传多样性,同时制定并执行了减少基因损失和保护其遗传多样性的战略。
▸ 爱知生物多样性目标16:到2015年,《关于获取遗传资源以及公正和公平地分享其利用所产生惠益的名古屋议定书》已经根据国家立法生效和实施。
截至2019年12月,世界自然保护联盟濒危物种红色名录(IUCN,2019a)总共收录了20334个树种,其中8056种被评估为全球受胁物种(极度濒危、濒危或易危)。共有32996个树种得到了一定层面上的保护评估(国家、全球、区域),其中12145个树种被认定为受胁。其中的1400多个树种被评估为极度濒危,急需采取保护行动(Global Trees Campaign,2020)(见插文16)。近年来,由于担心许多商业上有价值的树种可能受到过度采伐的威胁,《濒危野生动植物种国际贸易公约》收录的涉危树种数量激增。《濒危物种公约》附录中现已包含如檀木、黑檀木和桃花心木等900多个树种,通过该《公约》对其贸易进行了管制(《濒危物种公约》,2019)。
《欧洲濒危树种红色名录》(Rivers等,2019)对欧洲454个本地树种的保护状况进行了评估,结果表明,该区域58%的特有树种(地球上其他地方都没有的树种)为受胁,而42%的本地树种濒临区域灭绝。在特有物种中,有15%(66种)被评估为极度濒危,距离灭绝仅一步之遥。入侵害虫、疾病和植物是对欧洲树种的最大威胁。
花楸属树种所受影响尤为严重;欧洲170个花楸属树种中有四分之三被评估为受胁。
欧洲七叶树(Aesculus hippocastanum)受到源自巴尔干山区并扩散至欧洲其他地区的入侵物种潜叶蛾(Cameraria ohridella)的侵害,在经历了数量的减少之后,已被列为易危树种。
资料来源:IUCN,2019b。
在一些国家,人们致力于识别和保护森林外的树木,这些树木因其形态、树龄、历史意义或其他特质而引人注目(插文17)。
近几十年中,一些国家、州、地区或城市已经致力于识别和保护文化遗产树(有时被称为树王、历史树、地标树或重要树)。这些树由于其特殊的树龄、稀有性、树形大或美观、或具文化、历史或生态价值等原因而被认为具有独特价值。一个树种最古老的个体代表着重要的基因库,还是一个鲜活的气候变化博物馆,记录了数百或数千年间的变化(美国/ICOMOS,2019)。
在世界各地,不同的名树录一般关注有价值的、有时甚至是濒临灭绝的标志性树木。一些名树录是由国家非政府组织管理的,例如美国的国家冠军树名录、英国和爱尔兰的名树录以及澳大利亚的名树簿。这些名录通常不与任何监管机构和机制相关。然而,一些特殊的文化遗产树得到了国家、州、地区或市镇法律的保护(美国/ICOMOS,2019)。例如,在新加坡,根据2001年通过的“文化遗产树计划”,文化遗产树被依法保护。该计划是其国家保护措施的一部分,它不仅保护位于保护区内的树,而且还保护位于城市和乡村任何地方的树。在美国,许多城市都有禁止砍伐一些特定树木的文化遗产树木条例。
皮乔尼先生树(Albero del Piccioni)是一棵位于意大利马尔凯大区阿斯科利· 皮切诺(Ascoli Piceno)附近高24米、胸径8.7米的古法国梧桐(Platanus orientalis),其曾在一张1718年版的地图上被提及。
©Infinitispazi/维基共享资源在意大利,2014年国家法律颁布了一个名树录条例,其中包括在农林牧区或城市环境中的单棵树和树群,这些树因其树龄、大小、形态、稀有性、为动物提供栖所、以及历史、文化和宗教价值而被称为“绿色地标”。信息的收集由意大利农业部(MIPAAF)协调指导,并由法律规定的大区、自治省和直辖市实施。2017年发布的第一份名录包括2407棵树,并于2018和2019年分别更新增加了332和509棵新列名树。研究中心、学术机构、林业专业人士、环境协会和公民协助确定了这些名树(MIPAAF,2017;MIPAAF, 2019)。
目前科学上已知约有391000种维管植物(包括上述60082个树种和1600多个竹种(Vorontsova等,2016)),约94%为开花植物。其中有21%濒临灭绝(Willis,2017)。这些物种的60%存在于热带森林中(Burley,2002)。迄今为止,约144000种真菌得以命名和分类。然而,据估计,绝大多数(超过93%)的真菌物种至今尚不为科学所知,暗示地球上的真菌物种总数可能在220到380万之间(Willis,2018)。
已知并被描述的脊椎动物有近7万种(IUCN,2019a)。其中,森林为近5000种两栖动物(占所有已知两栖物种的80%)、近7500种鸟类(占所有鸟类的75%)和3700多种不同的哺乳动物(占所有哺乳物种的68%)提供了栖息地(Vié、Hilton-Taylor和Stuart,2009)。依赖森林生存的标志性物种包括美洲豹、北美熊、中非大猩猩、马达加斯加狐猴、中国熊猫、菲律宾鹰和澳大利亚考拉。
已知并被描述的无脊椎动物有约130万种。但是,还有更多种未被了解的无脊椎动物,其物种数估计在500万到1000万之间(例如见Ødegaard,2000)。大多数是昆虫,绝大多数生活在森林中(见插文18中的例子)。
野生和家养的传粉动物在森林景观中都起着重要作用,为农作物、野生植物和树木提供授粉服务。因此,它们对于维持生物多样性和相关的生态系统功能至关重要,也对生产木材和非木质林产品(NWFPs)的树木的繁殖再生十分重要,因此对森林的适应性和确保粮食安全和可持续生计举足轻重。全球约有87.5%的野生开花植物由动物授粉(94%的热带物种和78%的温带物种)(Ollerton、Winfree和Tarrant,2011),而115类主要粮食作物的75%在产出水果、蔬菜或种子时需要授粉动物一定的帮助(Klein等,2007)。然而,许多授粉动物(尤其是野生蜜蜂和蝴蝶)正受到威胁(IPBES,2016)。粮农组织和国际生物多样性组织正在进行的一项新研究(Krishnan等,即将出版)的证据表明,家养和野生授粉者种群的减少都可能对森林的自然再生和维持森林的遗传多样性产生严重影响,因此对其应对气候变化的适应力以及对病虫害的抵抗力产生严重后果。
尽管对社会型蜂种的研究最多,但是各类生境不同、食物迥异的动物也可提供授粉服务。例如,猴面包树(猴面包树属)和热带雨林树(Syzygium cormiflorum)由蝙蝠授粉。蜜蜂是花最常见的授粉者,其次是飞虫、蝴蝶和飞蛾(Winfree等,2007)。
授粉者受益于多种多样的自然栖息地,赖以觅食和筑巢。影响授粉者数量和多样性的因素包括土地使用变化、景观构成、森林经营措施和气候变化(IPBES,2016;Krishnan等,即将出版)。气候条件的变化可以改变植物展叶、开花和果实成熟的时间、质量和持续时间。对植物与动物之间相互作用关系的破坏可能对两个群落都产生负面影响。
授粉者生境的破碎和退化以及授粉者生境之间的连通性破坏会降低繁殖成功率,从而降低授粉者的种群规模。已经发现,规模较小的授粉昆虫种群会导致某些桉树树种后代自交增加和遗传变异降低,引起总体适应性下降,进而对其应对不断变化的环境的适应能力产生不利影响(Breed等,2015)。取决于破碎程度和所涉物种,增强跨森林区块的远距离授粉(例如鸟类授粉者)或许可以部分地弥补这一不足(Aguilar等,2008)。
另一方面,适度的干扰可以改善授粉者栖息地的质量和可用性,从而对授粉者多样性产生积极影响(IPBES,2016)。例如,与封闭森林相比,大多数蜜蜂似乎更喜欢开放森林,而森林碎片化仅在极度时才对蜜蜂产生负面影响(Winfree等,2009)。飞虫具备比蜜蜂和其他传粉者更强的适应生境变化或生境丧失的能力;随着土地使用的变化,某些物种的数量增加,而另一些物种的数量则减少(Stavert等,2007)。因此,森林经营可以在维持和持续提供授粉者方面发挥重要作用(Krishnan等,即将出版),但是选择最佳措施并不是一件容易的事,需要将诸多因素考虑在内。一些森林经营措施,如择伐和幼林萌生、保留死木、计划焚烧和不频繁除草等,可能会产生多样性更为丰富的生境,因而不仅对授粉者有益,而且也有利于森林其他生物多样性。保持足够的花卉多样性和林下植被也有助于为授粉者提供有利的生存条件。
昆虫在林下授粉者种群中占主导地位,而鸟类和哺乳动物更喜欢森林冠层。因此,景观管理需要考虑整个授粉者群体。森林景观中由鸟类和哺乳动物授粉的树种的多样性应通过积极的管理做法来保持,如树木的保留和种植。例如,在巴西,树木为授粉鸟在一马平川的农田中提供了落脚的地方。在高度碎片化的景观中,此类落脚石可以促进森林再生(Barros等,2019)。
全球范围内,已被描述记载的土壤细菌和真菌分别超过15000和97000种,与之相比的是20000-25000种线虫,21000种原生生物(原生动物、原生植物、原生菌类),以及40000种螨类(Orgiazzi等,2016)。然而,土壤生物群内许多物种仍然是未知的。土壤微生物、依赖森林的传粉动物(昆虫、蝙蝠、鸟类和某些哺乳动物)(插文18)、和腐木甲虫(插文19)在维持森林生物多样性和生态系统功能方面起着非常重要的作用。
腐木甲虫是一群在其生命周期的某些时期内依赖死木或腐木真菌的昆虫。它们在分解过程中起着重要作用,因而对森林养分循环具有重要意义。它们还是一些高营养级类群(如鸟类)的食物来源。许多腐木甲虫物种参与授粉。在地中海地区,它们的原生中心位于欧洲西南和东南部、土耳其、近东和北非地形变化多样的地区(如阿特拉斯山脉)。以橡树为主导树种的森林是腐木甲虫最重要的栖息森林类型。针叶人工林仅支持有限数量的腐木甲虫,通常与在半天然橡树林中发现的种群不同。对地中海森林中腐木甲虫构成主要威胁的是因砍伐树木、过度放牧和森林火灾而造成的栖息地丧失。
资料来源:粮农组织和Plan Bleu,2018。
同样,哺乳动物、鸟类和其他生物在森林生态系统结构中能起到主要作用,包括通过在散播和采食种子中的直接作用来决定树木的分布,以及在这样的生态架构中进行捕食的间接作用(Beck,2008)。
在热带海岸,红树林为无数鱼类和贝类提供了繁殖和抚育场所,并吸纳了可能对海草床和珊瑚礁(无数海洋物种的栖息地)产生不利影响的沉积物。
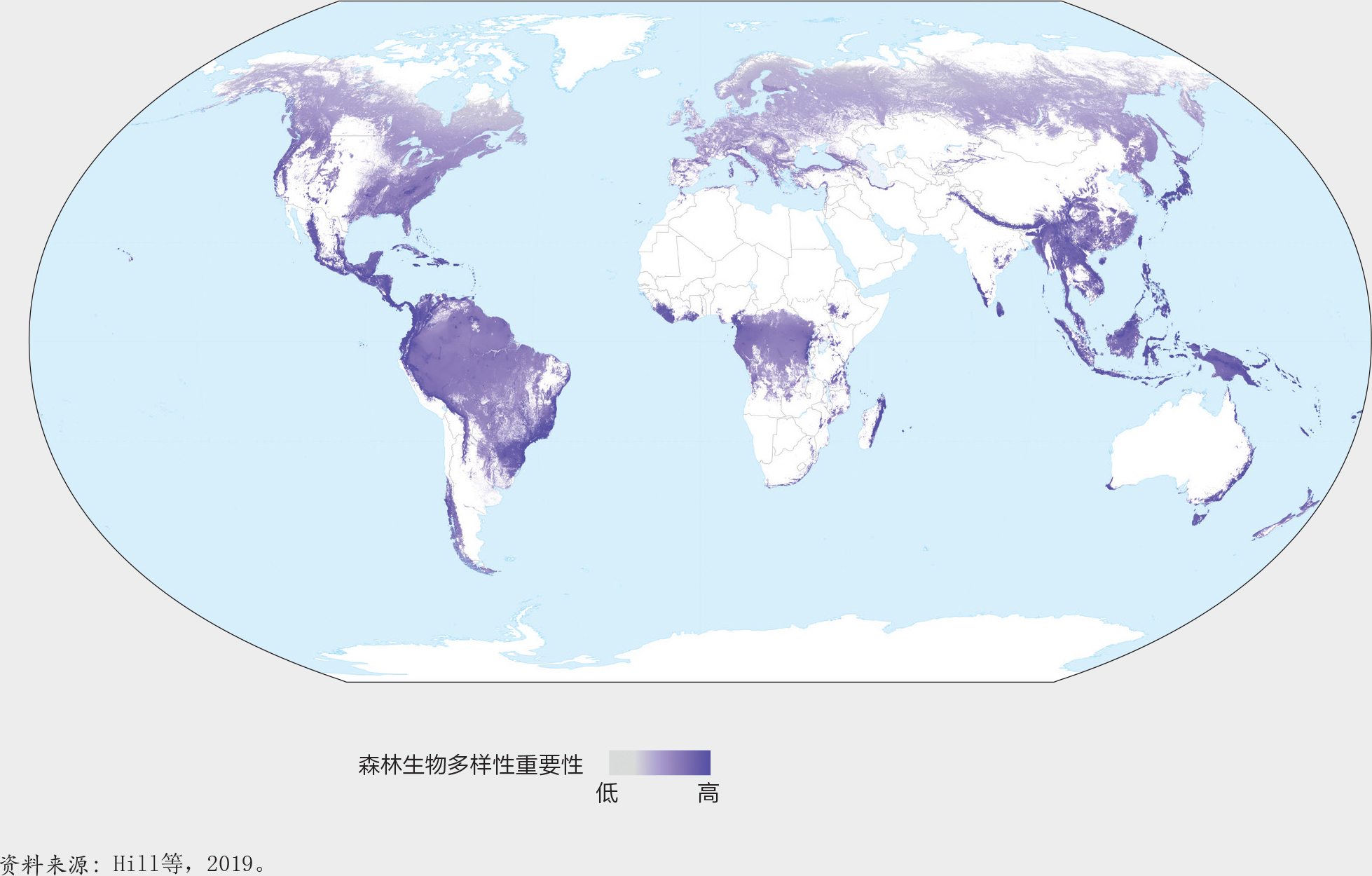
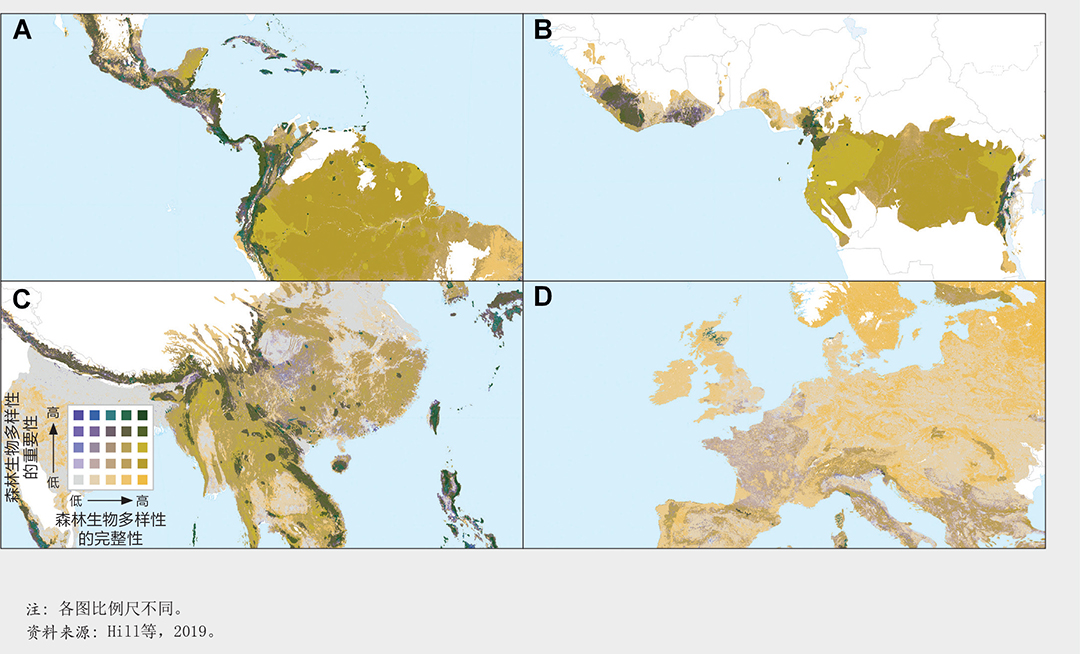
森林生物多样性的意义。森林生物多样性因森林类型、地理、气候和土壤等因素而存在很大差异。由世界养护监测中心主导的一项研究(Hill等,2019)显示了这些因素对哺乳动物、鸟类、两栖动物和针叶树种在全世界分布上的影响。该研究基于自然保护联盟红色名录中的数据(包括每个物种的分布地图),估算了这些物种以稀有程度为权重的种群丰富度(之所以选择这些物种,是因为它们是当时仅有的被全面评估的生物群体)。生物多样性重要性地图(图19)显示了与原生鸟类分布图和生物多样性热点地带分布图高度的相似性,但是与二者相比基于更多的物种(Myers,1990;Stattersfield等,1998;Mittermeier等,1998;Mittermeier等,2004)。
温带区域的大多数森林生物多样性的重要性较低,因为与热带区域相比,它们所支持的物种较少,而且与世界其他区域相比,它们所支持的物种的地理分布往往更广(图19)。亚马逊河流域和刚果盆地的低地热带森林生物多样性的重要性处于中等;尽管这些森林物种丰富,但目前其分布较广,因此任何单个位点对这些物种总体分布的贡献都很低。最具生物多样性重要性的区域是那些物种种类繁多、地理分布较集中的区域,例如南美、非洲和东南亚的山地森林以及东南亚岛屿、巴西沿海岸、澳大利亚、中美洲和加勒比海岛屿的低地森林。
图20基于一项关于2000-2018年间树木覆盖的丧失对森林生物多样性意义的分析,展示了森林生境的丧失可以对世界上依赖森林生存的物种产生不成比例的影响。影响最大的地区包括马达加斯加、巴西东部部分地区、中美洲、东南亚、西非、澳大利亚和新西兰北部。
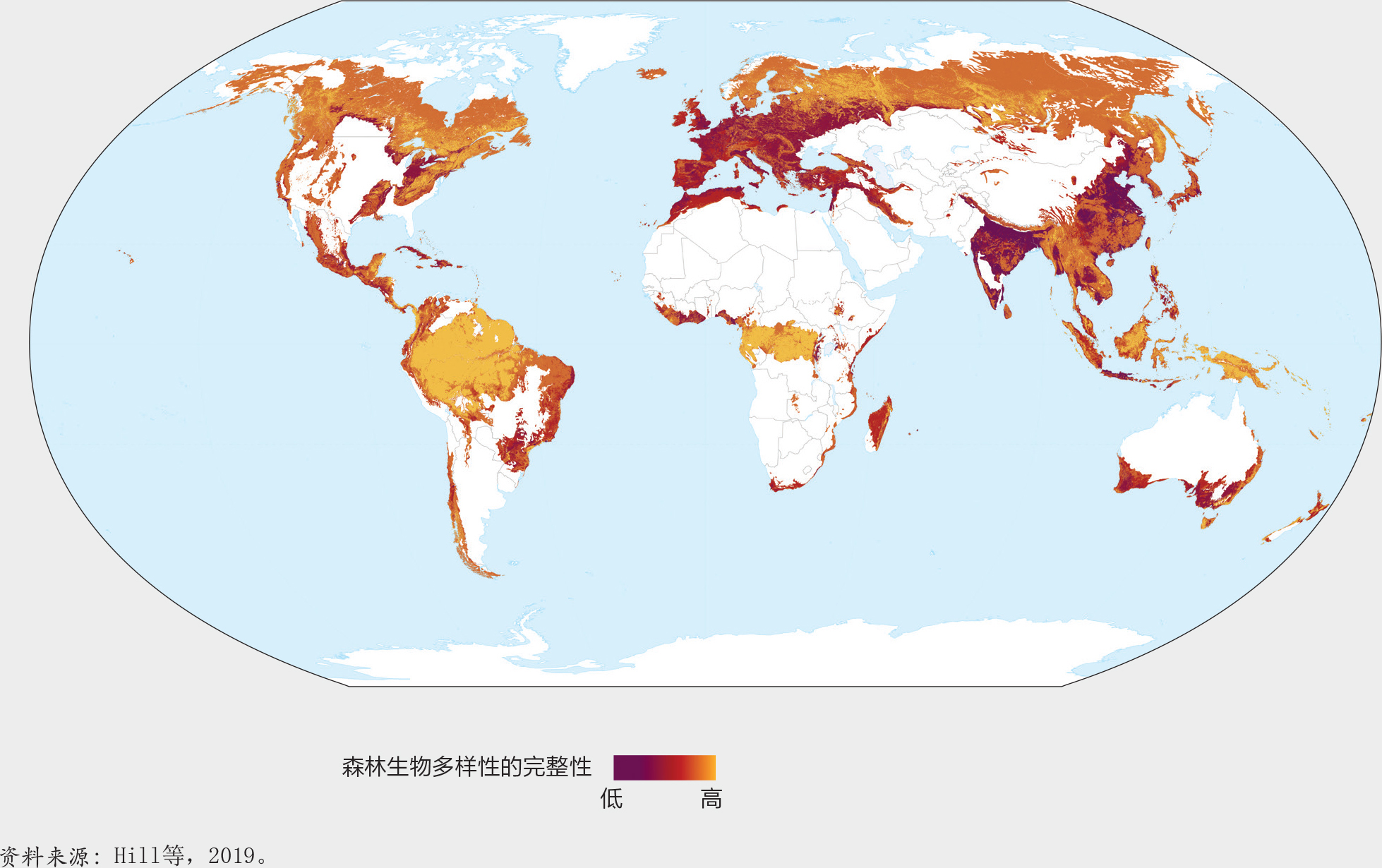
森林生物多样性的完整性。图21显示了森林生物多样性的完整性,说明了森林变化和人口密度对物种类群集合结构的影响;它是基于对来自人类活动的压力与物种群落组成变化之间关系进行的模拟。正如预期的那样,森林生物多样性在人口稠密和农业土地集约使用的地区完整性较差(如欧洲和孟加拉国局部地区、中国、印度和北美)。南澳大利亚、巴西沿海、马达加斯加、南非和北非也被认定为生物多样性完整性遭受重大损失的地区。
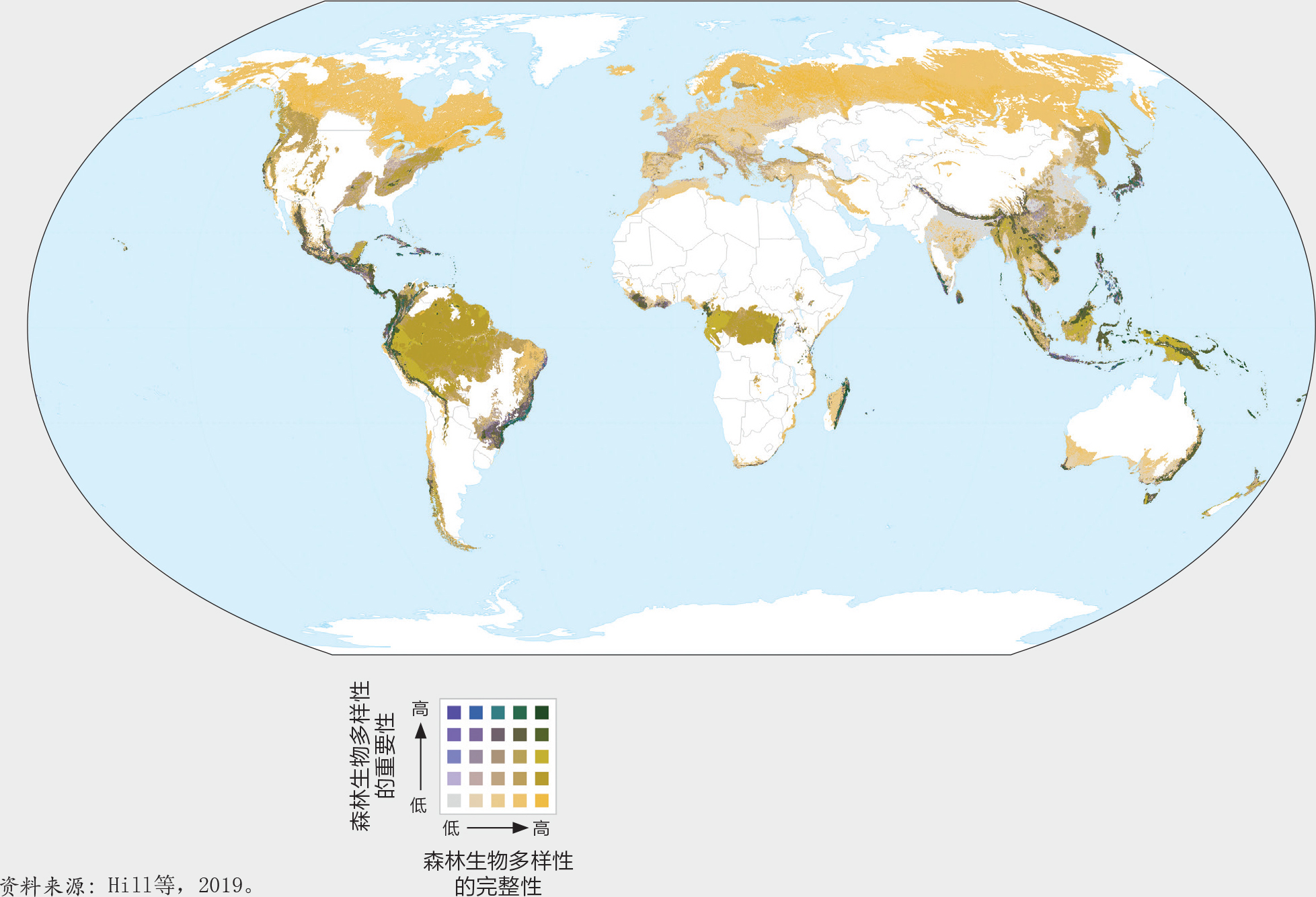
将完整性和重要性两项指标叠加进行保护规划。生物多样性的重要性和完整性指标对保护政策和措施具有互补性。保护具有重要性的地区非常重要,因为它们的损失会加剧物种灭绝的风险。维护完整性高的地区对于维持生态系统功能、保持应对如气候变化等压力的抵御力以及缓解气候变化带来的影响非常重要(Steffen等,2015)。
将生物多样性重要性和完整性指标两个地图层叠加后(图22)突显了两个指标均具有较高价值的区域,例如安第斯山脉中部北部和中美洲、巴西东南部、刚果盆地部分地区、日本南部、喜马拉雅山地区以及东南亚和新几内亚的多个地区(图23)。其他值得关注的是那些一个指标较高、而另一指标不高的地区。以欧洲为例,其东北地区有大片的生物多样性完整地区,而其南部以生物多样性重要性的地区为主(图23D)。
这样的叠加为保护规划提供了有关的信息。例如,重要性高但完整度较低的景观可能是开展恢复工作的恰当目标。完整性和重要性均高的景观具有较高密度的本地物种,因而通过广泛的政策措施和实地保护对其进行保护就非常重要,譬如指定保护区。相应的生态区内保护区森林覆盖率已经相对较高(参阅第6章:森林和森林生物多样性的保护和可持续利用), 但其中尚未被列为保护区的地方应当被视作保护区扩展的重点对象,譬如第斯山脉北部的山地森林。
这里强调的结果也与国际和国家政策有关,包括《生物多样性公约》下的国家生物多样性战略和行动计划。另外,绘制森林生物多样性重要性或完整性流失分布图随时间的变化有助于追踪一些目标实现的进展状况,如爱知目标5(生境的丧失和退化)、爱知目标11(对生物多样性有重要性的区域)和爱知目标12(防止已知濒危物种的灭绝和减少)。与生物多样性有关的森林损失数据还可以为减少毁林和森林退化的国家计划以及投资政策提供信息。
不久的将来,将会有工具将遥感数据与计算整合,可以几近实时地显示出森林流失的地区以及森林流失对生物多样性产生的后果,为及时对实地采取措施、进行干预提供可能。为了这个目标,生物多样性的重要性完整性和生物多样性图层均已被纳入全球森林观察平台(www.globalforestwatch.org)。
设定目标和监测进展状况的全球进程通常使用基于森林面积的指标来衡量森林生物多样性;例如,爱知目标5是到2020年将森林和其他自然栖息地的丧失速度减少一半。然而,最近的一项研究(Green等,2019a、b)质疑了用森林面积变化作为衡量森林脊椎动物种群趋势的可靠性。
这项研究使用了“生命星球数据库”1668种栖居于森林里的脊椎动物的种群数量的时间序列数据(伦敦动物学会和世界自然基金会, 2014),来评估树木覆盖变化对森林脊椎动物种群可能的影响。利用卫星图像来评估1982-2016年间树木覆盖的变化。又对175个“仅存于森林的物种”的种群进行了同样的分析,这些物种仅生存在森林中,而从没出现在其他生态系统里。
从全球数据整体来看,分析结果并未显示出树木覆盖变化与栖居于森林里或仅存于森林里的脊椎动物的种群变化之间存在统计上的显著关系。因此,在全球范围内,森林脊椎动物种群对其附近的树木覆盖变化似乎并未作出一致的反应。树木覆盖增加的地区并不一定会看到森林生物多样性的恢复,也许是由于其所受压力与生境丧失无关。然而,在局部范围内,这种关系在特定情况下在统计上是显著的。175种仅在森林里栖居的脊椎动物中有40种的年种群数量与树木覆盖变化呈正相关,而其他物种与树木覆盖变化呈负相关或不相关。由于森林脊椎动物可能需要数年时间才能对栖息地的变化作出反应,因此该研究考虑到了树木覆盖变化与种群变化之间存在一定的滞后性。关于这些只栖居于森林里的脊椎动物种群数据和文献还暗示,在局部地区存在着其他影响种群数量的因素(见插文20中的例子),依靠森林覆盖变化这一单一指标来衡量脊椎动物种群变化是欠妥的。
哥斯达黎加的圣罗莎国家公园始建于1971年,源于开垦的牧场。自1971年被指定为公园以来,该公园一直受到保护,免受狩猎、人为活动和伐木等干扰,因而以前的牧场已渐渐重归森林。
对鬃毛吼猴(Alouatta palliata)和白面卷尾猴(Cebus capucinus)的长期监测表明,这些种群的恢复与森林的重建有关(图A),但同时也揭示了还存在除森林面积和状况外影响其种群规模的其他因素(Fedigan和Jack,2012;Green等,2019a)。卷尾猴可以居住在幼林中。圣罗莎的最新调查表明,自上世纪80年代以来,该种群一直在持续增长。然而,鬃毛吼猴更喜欢较成熟的森林(林龄至少达60年),而且自上世纪90年代以来种群处于稳定状态,这表明该种群在国家公园中已达到其目前的最大承载能力。
在圣罗莎也发现了黑掌蜘蛛猴(Ateles geoffroyi),但它们仅出现在大片的老熟林中(林龄至少达100至200年)。该物种的种群可能需要数十年才能对森林覆盖的提高和成熟树的增加做出反应。
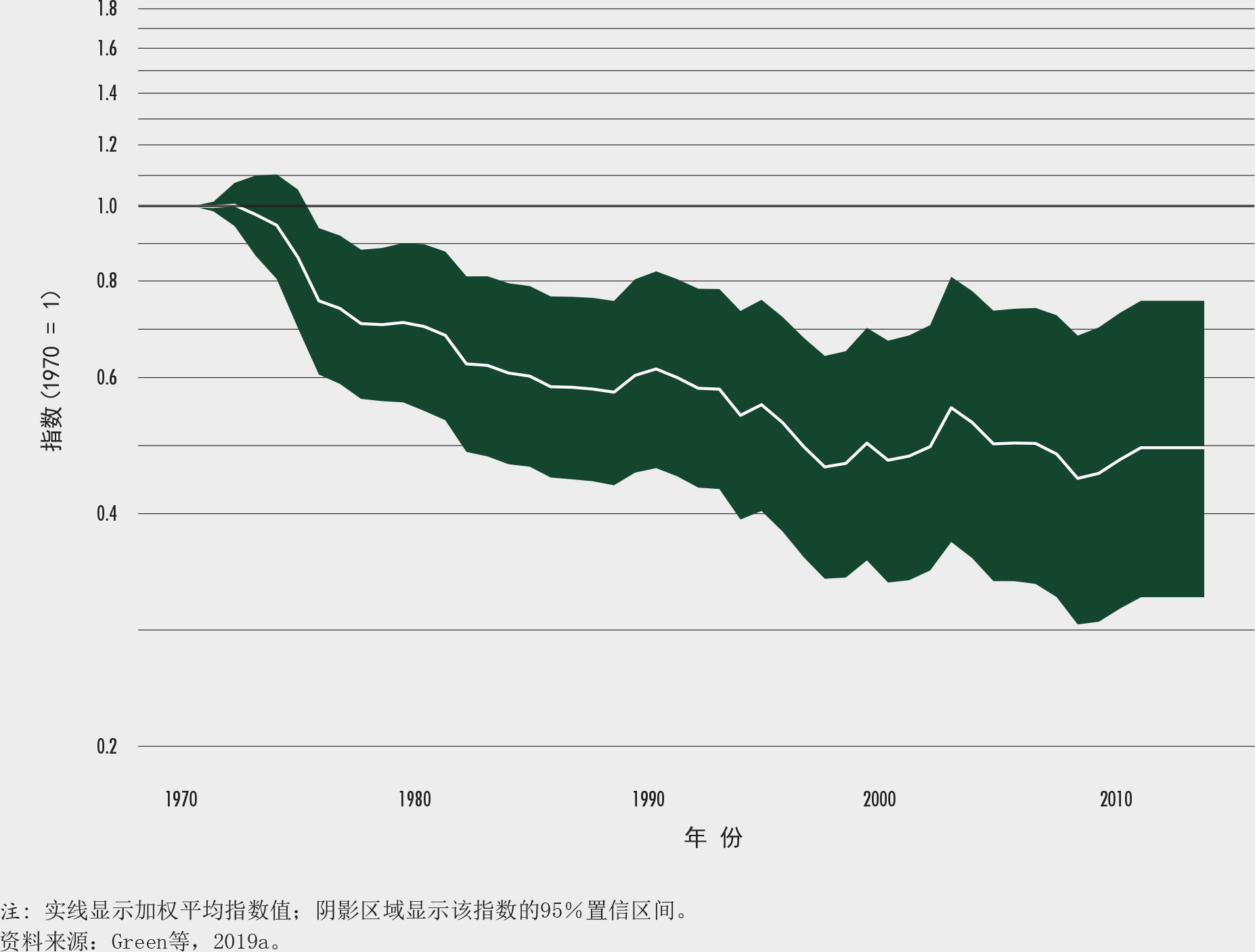
制定一个基于仅存于森林的物种指数。作为上述森林脊椎动物生物多样性研究的一部分,Green等(2019a)定义了一个仅在森林里生存的物种的指数,作为一项衡量全球森林冠层下生物多样性趋势的指标。该指数的制定汲取了“地球生命力指数”中仅存于森林的物种相关的信息(伦敦动物学会和世界自然基金会, 2014)。“地球生命力指数”追踪记录了全球数以万计的脊椎动物种群规模的平均变化。约75%仅存于森林的物种来自热带森林,全球生物多样性最丰富的森林。
1970-2014年间,仅存于森林的物种指数从1.0下降到0.47,降幅53%(图24),被监测物种中有455个在此期间种群数量减少超过一半,年降幅为1.7%。这一发现在哺乳动物、两栖动物和爬行动物中是一致的,但在鸟类中则不然,尤其是温带森林中的鸟类。该指数在1970-1976年间跌幅最大,此后虽继续下降但速度放缓。在研究期间的最后两年,物种种群增加的数量超过了物种种群减少的数量。然而,鉴于先前的种群改善全都伴随着种群的下降,因此尚不确定这种好转是否标志着仅存于森林的物种在种群数量上的长期改善。在热带和温带森林中,单一物种显示出改善、稳定和恶化的趋势,不一而同;热带森林中恶化的状况较为普遍,温带森林中改善的情形较多。
仅存于森林的物种指数可以作为现有指标的一个有益的补充,用以监测在实现可持续发展目标15、《生物多样性公约》 2020年后全球生物多样性框架和《巴黎协定》目标上的进展状况。《生物多样性指标伙伴关系》(2018年)已提出建议将该指数作为衡量实现爱知目标5、7和12进展状况的一项指标。
野生动物狩猎对森林生物多样性的影响。不可持续的野生动物狩猎是生物多样性丧失的主要因素之一,仅次于农业扩张(Maxwell等,2016)(另参见第5章:扭转毁林和森林退化)。一项对世界自然保护联盟红色濒危物种名录(IUCN,2019a)上8688种动物濒危信息进行的全球荟萃分析估计,热带地区哺乳动物和鸟类在狩猎地区的相对丰富度比在非狩猎区分别低83%和58%(Benítez-López等,2017)。红色名录中近20%濒危物种(极度濒危、濒危和易危)和近乎濒危的物种受到狩猎的直接威胁(Maxwell等,2016),其中包括300多种哺乳动物(Ripple等,2016)。繁殖率低、世代时间长、体型大的物种特别容易受到狩猎的不利影响(Ripple等,2016);因此,在狩猎森林中的脊椎动物中,体型小的物种占较大比例,例如大鼠、鸟类和松鼠。在狩猎压力极大的情况下,森林最终可以达到有树但却没有大型哺乳动物踪影的地步,这种现象被称为“空林综合征”(Redford,1992)。热带森林中最常被捕食的哺乳动物是食果动物,这些物种、大型鸟类和冲积林中一些鱼的减少或灭绝会对种子的传播和成活以及森林的再生产生重大影响(Galetti等,2008;Peres等,2016;Gardner等,2017)。因此,在非洲、亚洲和美州热带等一些地区,种子大、需要动物散种的树种比例较高,因此森林脊椎动物的丧失或减少可导致树种多样性的减少(Poulsen、Clark和Palmer, 2013;Bello等,2015;Osuri等,2016)。另一方面,在许多森林覆盖率高的国家,可持续狩猎不仅可以带来收入,也是一项重要的娱乐活动,从而成为保持森林的动力(如Reimoser,2000;Bengston、Butler和Asah,2008)(见第6章第128页可持续狩猎和野生动植物管理)。
森林遗传资源是具有实际或潜在的经济、环境、科学或社会价值的林木和其他木本植物物种(灌木、棕榈和竹类)的可遗传物质(粮农组织,2014b)。《世界森林遗传资源状况》(粮农组织,2014a)有史以来首次汇集了来自86个汇报国家的信息,占全球森林面积的85%。这些国家报告了近8000种树木、灌木、棕榈和竹,其中约有2400种被积极管理用于提供林产品或林业服务。
共计1000个物种予以原地保护,1800个被实施迁地保护(见插文21关于保护类型利弊的讨论)。大多数森林遗传资源的就地保护是在保护区之外各类公有、私有和传统拥有土地之上,特别是在多功能经营之下的森林中。被报道的迁地保护的物种也许比原地保护的物种更多,这主要是因为迁地保护通常比原地保护被更好地记录和登记。各国对原地保护的解释也不同。有时在一个保护区内发现某物种的踪迹就会被称为原地保护,即便该保护区的设立主要是为了保护栖息地或野生动植物,而不是为了保护森林遗传资源。
面对不断变化的社会需求和气候变化,动态的原地保护方法对森林遗传资源的长期保护至关重要。迁地保护大多是静态的,主要依靠保护和管理收集来的遗传多样性样本(如组织、种子或活植物收集)。
森林遗传资源的原地保护通常是在经营的天然林或保护区内,通过指定专门的保护林分或保护单位来实现(粮农组织、DFSC和IPGRI,2001)。这些单位可能包含一个或多个树种的被保护种群。如有必要,可以采取营林措施,以维持或增强树木种群内的遗传过程,并确保其再生。理想情况下,这些保护单位构成的网络应覆盖该树种的整个分布范围。除了物种的分布范围外,还需要有关其生殖生物学和遗传特征以及现有保护措施的信息,来评估当前遗传资源保护策略的有效性,指出可以改进的地方(如Lompo等,2017)。
森林遗传资源的迁地保护(如在种子库、种质圃、种植试验园和植物园中)通常作为原地保护的补充,特别是在野生种群数量极少或无法保证原地保护的情形下。利用种子库进行迁地保护对那些种子在干燥和低温保存后仍能保持活力的物种相对容易些。然而,这种方法不能用于无休眠并且对干燥和低温敏感的树木。在潮湿的热带地区,超过70%的树木是这种情况。迁地保护这些物种必须依靠野外收集、设立迁地保护林分和育种群(Sacande等,2004)来实现。更先进的技术手段,例如种子的冷冻保存、组织的体外保存、花粉储存和DNA储存,也可以用于此类物种(粮农组织、FLD和IPGRI,2004)。
天然再生依赖可在当地或附近容易获得的遗传物质,而植树通常意味着利用外部来源的种质。由于林分的轮伐期可能长达数十年,甚至超过100年,因此,关键要确保引入的种质起源适合当地的环境条件,并且其材料具有足够的遗传多样性使新森林能够应对不断变化的环境条件和可能的病虫害。
一旦建立了天然林或人工林,随后的营林干预措施可能对其遗传组成产生深远影响。这些影响的程度取决于特定的营林措施和林分结构,以及该物种的生物特性和生态(Ratnam等,2014)。
全球范围内的树木改良计划包括700多个树种,主要关注与商业利益相关的特征,例如生长、木材特性以及对病虫害的抗性或耐受性。然而,最近,林木育种计划越来越多地考虑了与气候变化相关的特征,例如可塑性和耐旱性(粮农组织,2014b)。
在全球范围内,旨在增加立木蓄积量的林木种质的供应仍主要依靠从林分中收集来的、未经改良的种子,但是各个区域和国家在林木种质的来源和生产上存在很大差异。一个极端是,大多数种植的林苗都是由改良种子培育而成的。另一极端则是,几乎所有种子都来自现有的森林或来源不明的人工林,甚至来自农田中的单个树木(粮农组织,2014b)。北寒带、温带和热带及亚热带速生树种的种子供应基本满足了造林的需求,但是许多昂贵热带硬木和混农林业系统中使用的树木的种子供给不足,难以满足其需求(Koskela等,2014)。最近,日益增加的森林恢复项目对本地树种的种子提出了很高的要求,许多项目已经面临供求上的挑战,难以充分地获取具有良好生理和遗传质量的种子(Jalonen等,2017)。
2019年,粮农组织开始编写第二版《世界森林遗传资源状况》,该报告将于2023年发布。预计第二版全球评估将增进人们对知识上存在差距的认识,并强调获得更好信息和森林遗传资源数据的重要性,以加强国家、区域和全球各个层面对这些资源的管理(见插文22中的例子)。
混农林业绿地是撒哈拉以南非洲许多地区的传统土地使用系统。农民在这些绿地中保留的树木为农村居民提供了野生水果、坚果和蔬菜,特别是在农作物收获季之间和长期干旱期间。不幸的是,许多食用树种面临着来自过度开发、火灾和气候变化的威胁。
为了加强布基纳法索这些树种遗传资源的保护,国际生物多样性组织的科学家及其合作者开发了一种威胁因素空间模型,以预测当前和未来可能对树木种群产生负面影响的威胁的空间位置(Gaisberger等,2017)。1该研究有针对性地选择了16个食用树种,主要基于它们对当地居民食物的重要性及其空间数据的可得性(对构建空间模型来说至关重要):猴面包树(Adansonia digitata)、非洲番荔枝(Annona senegalensis)、沙漠椰枣(Balanites aegyptiaca)、木棉树(Bombax costatum)、汉莎(Boscia senegalensis)、小果荚髓苏木(Detarium microcarpum)、非洲葡萄(Lannea microcarpa)、非洲刺槐豆(Parkia biglobosa)、Senegalia macrostachya、阿拉伯胶树(Senegalia senegal)、非洲漆树(Sclerocarya birrea)、大象橙(Strychnos spinosa)、酸角(Tamarindus indica)、乳油木(Vitellaria paradoxa)、海檀木(Ximenia americana)和印度枣(Ziziphus mauritiana)。它们中的一些分布广泛(例如Parkia biglobosa),另一些则提供多种可食用的食品(例如猴面包树的叶子、种子和果肉)。
该模型通过结合可得数据、物种分布模型、气候模型和专家调查结果中的信息,界定了这些树种在当前和未来条件下适宜的栖息地。在确定的六种主要威胁因素中,过度开发和土地用途转为棉花生产在短期内被认为是最重要的,气候变化对16个树种中的14种构成长期威胁。该研究还表明,所有16个物种在布基纳法索的大部分地区都面临严重威胁,暗示急需采取紧急行动来保护该国的物种及其遗传资源。
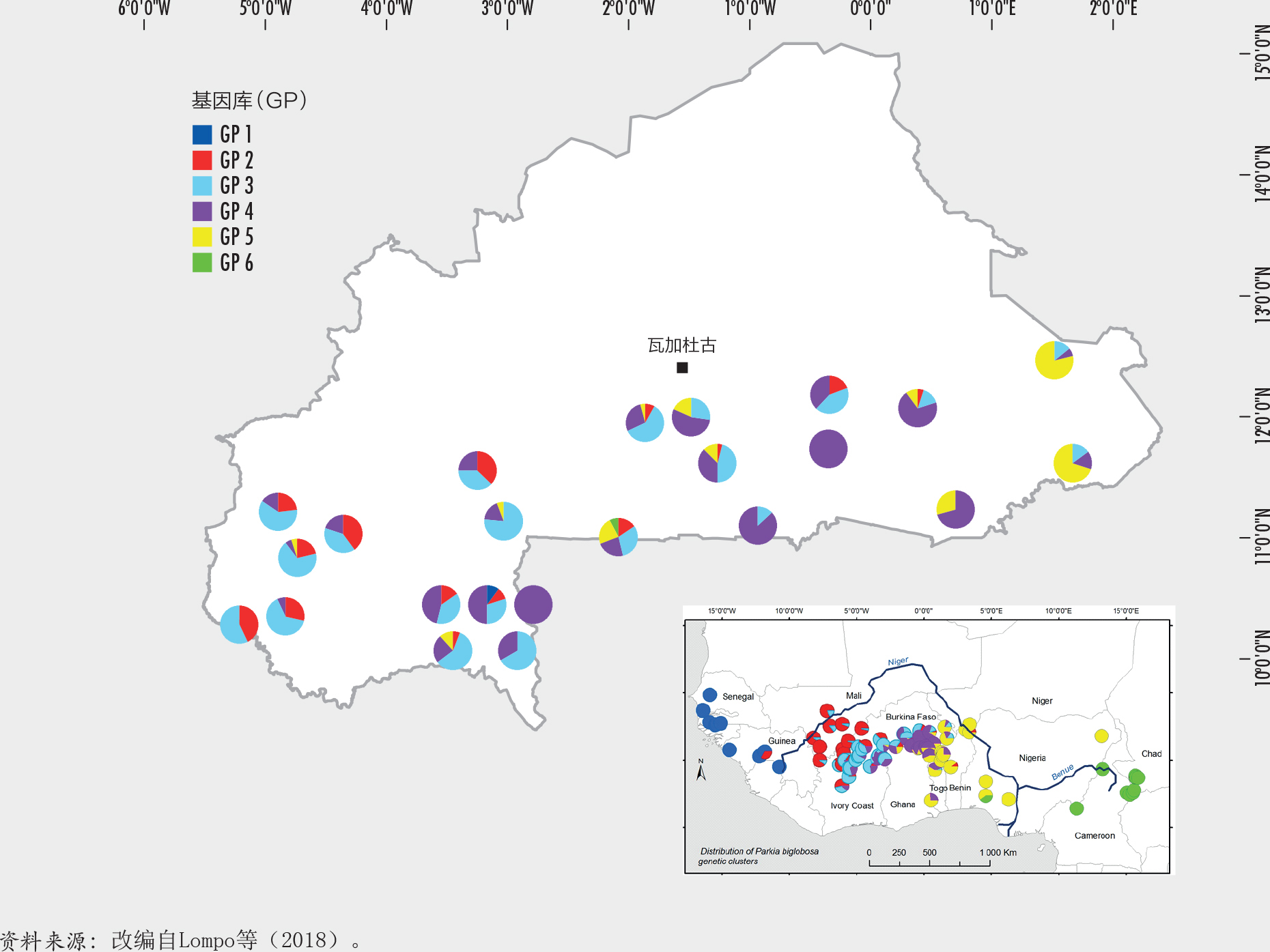
观察哪里物种遗传多样性丰富但受胁程度也较高,有助于设计出效率更高的保护措施,也便于利用有限的资源来维持物种分布范围内树木种群的遗传多样性。例如,非洲刺槐豆(Parkia biglobosa)在布基纳法索中部遭到严重的过度攫取(图Aa),当该物种种群数量在未来气候条件合适的地区增长的时候,应该在这些地区致力于保护和辅助再生。非洲刺槐豆位于其分布范围北缘的种群受到气候变化的严重威胁(图Ab),除非实施迁地保护或者收集这里的种子并将其种植在更适宜的气候中,否则该地区有价值的种质来源可能会永久丧失。一项范围广泛的基因分型研究为深度了解刺槐豆种群在西非的空间遗传结构分布提供了重要信息(Lompo等,2018)。通过比较Gaisberger等(2017)提供的威胁因素分布地图和来自Lompo等(2018)的布基纳法索遗传多样性分布地图(图B),可以判定出那些高危的具备特殊遗传特质的树木种群,急需对它们予以优先保护。这些信息还可用于指导植树工作。
实现爱知目标12(防止已知濒危物种灭绝和改善其保护状况)的进展一直以来比较缓慢。
表3汇总了截至2019年12月的自然保护联盟红色名录(2019a)中所评估的森林动植物和真菌的涉危状况。
地球生命力指数的计算基于全球监测的代表4005个物种的16704个种群的数据,该指数表明脊椎动物的种群规模在1970-2014年间总体下降了60%(世界自然基金,2018)。基于这一模型的仅存于森林的动物指数在1970至2014年间下降了53%(图24,第48页),突显了268种森林脊椎动日益面临灭绝的风险。
实现爱知目标13(维持栽培植物、养殖和驯养动物以及野生近缘物种的遗传多样性)和16(落实《名古屋议定书》)的进展相比而言较为乐观。截至2020年1月:
▸《名古屋议定书》已经得到包括欧盟在内122个缔约方的批准(比2016年增长74%)(《生物多样性公约》,2020a);
▸95个国家和欧盟已就《名古屋议定书》的执行情况向获取和利益分享信息交换中心提交了临时国家报告(《生物多样性公约》,2020b);
▸2018年,提交进度报告的44个国家平均已经实现了《森林遗传资源养护、可持续利用和发展全球行动计划》中三分之二的行动要点(插文23);
▸一项泛欧洲战略加强了在欧洲保护森林遗传资源的区域合作(插文24);
▸146个缔约方批准了《粮食和农业植物遗传资源国际条约》(粮农组织,2019d)。
2013年粮农组织大会通过自愿、无约束力的《森林遗传资源养护、可持续利用和开发全球行动计划》(粮农组织,2014b),确定了国家、区域(见以下插文24)和全球层面的四个优先行动领域,以加强森林遗传资源管理:
▸ 改善森林遗传资源信息的提供和获取;
▸ 保护森林遗传资源(原地保护和迁地保护);
▸ 森林遗传资源的可持续利用、开发和管理;
▸ 政策、机构和能力建设。
2017年,粮食和农业遗传资源委员会通过了森林遗传资源的目标、指标和验证指标,可用于监测《全球行动计划》的执行情况。这些目标和指标还可用于监测在实现爱知生物多样性目标13(以及一项可能在2020年后取而代之的新目标)以及可持续发展目标相关具体目标方面取得的进展。
2018年,有44个国家提交了进展报告,粮农组织基于这些报告提供的信息编写了关于《全球行动计划》执行情况的第一份报告(遗传委,2019)。尽管报告国家数量有限,无法对各国在执行《全球行动计划》方面取得的进展作出全面的结论,但仍可得出一些结论:
▸ 报告国平均已实现该计划中67%的行动要点,并已开始努力更进一步,再实现10%;
▸ 在44个报告国中,只有四个实现了所有15个行动要点;
▸ 许多国家缺乏人力和财力来开展和报告对所有重要和有用的森林物种的保护行动,特别是对濒危、受胁和稀有物种的保护。
许多树种的分布范围跨越具有巨大环境差异的广大区域。这些范围通常包含许多具有不同森林管理措施、所有权模式和行政管理的国家。由于这些原因,森林遗传资源的管理和保护在物种分布范围内经常有很大差异。
在原地保护欧洲树种的遗传多样性以及制定森林遗传资源保护区域战略方面的努力,曾经在相当长的一段时期内受到阻碍,主要是由于各国对于如何管理树木种群、指定保护林分以及记录上的不健全存在差异。
为解决这个问题,作为欧洲森林进程(欧洲森林,日期不详)下的一个合作机制,欧洲森林遗传资源计划(EUFORGEN,www.euforgen.org)为森林保护点进行遗传资源保护制定了通用的最基本要求,规范了如何记录和管理这些保护点的信息(Koskela等,2013)。欧洲森林遗传资源信息系统(EUFGIS,http://portal.eufgis.org)收集了这些森林遗产资源保护点的地理位置信息,使得在国家和区域层面发现保护工作可改进的地方成为可能(Lefèvre等,2013),同时也方便分析气候变化对欧洲森林树木遗传保护的预期影响(Schueler等,2014)。
基于这些信息,欧洲森林遗传资源计划制定了一项保护森林遗传资源的泛欧洲战略(de Vries等,2015)。在此进程中,将树种的分布范围按国家和欧洲的八个主要环境区划分为较小的地理区域,进而确定了每个树种的区域最低保护目标。对于每个树种,该战略以在该树种发现国所在环境区建立至少一个该树种的保护点为目的;这样就可以对整个物种分布范围内的所有国家和环境区进行系统覆盖(不考虑保护工作实施方面的偏差)。欧洲森林遗传资源计划还为保护森林遗传资源工作中考虑气候变化的影响提供了建议(Kelleher等,2015)。
截至2019年12月,欧洲森林遗传资源信息系统囊括35个国家的3593个遗传保护点和108个树种的数据(见图A中的实例)。该数据库还在不断更新,欧洲森林遗传资源计划定期监测该区域保护战略的执行情况。
这种区域合作促使许多国家采取行动来改善其森林遗传资源的管理,同时改善了专家、森林所有者、管理者和更广泛的生物多样性界人士之间的合作伙伴关系,探索用新的方法来增进生产林和保护区对森林遗传资源保护的贡献。
当今大部分人类社会与森林及其蕴含的生物多样性存在某种联系,所有人都从森林的碳、水和养分循环及其与粮食生产的关联中享受到森林生物多样性带来的益处。
人们与森林生物多样性的关系因地区而异,因国家而异,也因背景和实际情况而异:从人类活动有限的保护区,到居住在森林深处的族群,到农田和牧场,到城镇和较大的城市中心,到世界最大的城市。本章从生计、粮食安全和人类健康方面探讨人们从森林中获得的益处。
在发展中国家和发达国家以及所有气候带中,生活在森林中的人群都最直接地依靠森林生物多样性得以生存和维持生计,他们从森林获得食物、饲料、居所、能源、药品并获取收入。其他农村人口大多生活在有草原、农田和树木的地方,常常参与森林生物多样性的价值链,例如从附近森林中收集木材和非木质产品供自家使用或出售,或在林产品产业中就业,参与增加林产品附加值的活动(Zhang和Pearse, 2011)。尽管以下所举例子表明了依赖森林获取生计(或部分生计)的人数,然而目前就依赖森林的人数而言仍然没有一个精确的估计(插文25)。
对那些关注与森林、生物多样性和人息息相关的政策、措施、规划和投资的人们来说,他们面临的一个难题是确定最依赖森林资源的人群数量及其人口、社会和经济特征,这些人群经常被称为“依赖森林的人”。人们与森林互动的异质性使得很难用一个标准且有意义的方式来定义对森林的依赖(Newton等,2016)。例如,世界上许多粮食的生产都依赖于像淡水、授粉者和气候调节这样的森林生态系统服务。此外,缺乏可靠的数据以及衡量和追踪记录对森林依赖状况的方法;一般而言,与人口、社会、经济、健康和贫困指标有关的国家和国家以下各级统计数字并未对生活在森林中和森林附近的人口进行分类。参与非木质林产品的采集和贸易的通常绝大多数是妇女,对其活动的记录统计状况尤为不佳(Gurung, 2002;Watson,2005)。
然而,许多人口统计数据被用来估计人类对森林的依赖程度,并由此推断人类对森林生物多样性的依赖。随着世界各地农村人口的变化,经常引用的全球16亿人口依赖森林(世界银行,2002)这一数字在一定程度上已经过时。根据国际农业发展基金(农发基金)和其他来源的数据,粮农组织(2018b)估计发展中国家约有8.2亿人生活在热带森林和热带稀树草原中。基于世界银行、热带雨林基金会和世界热带雨林运动的数据,Chao(2012)估计约有12亿人依靠混农林业生产系统。此外,还有3亿至3.5亿人居住在茂密的森林内或其附近,以求取生存和获得收入。农发基金和联合国环境署(2013)给出了更广泛的估计,据估计有25亿从事小农农业的人得益于其生产系统中森林和树木所提供的供给型和调节型服务。此外,在发展中国家和发达国家的城市和农村地区,有24亿人使用木质能源进行烹煮、取暖和饮用水消毒(粮农组织,2017a)。
总体而言,就2019年12月世界人口约为78亿而言,这里提出的估计数字表明,大约三分之一的人类对森林和林产品高度依赖。但是,很难估计这个数字是如何随着全球趋势(例如从农村到城市的迁移)而变化的,以及如何随全球人口预计在2050年增加到100亿左右而变化。
由于关于依赖森林的人数的信息稀少,因此很难去设计有针对性的干预措施和政策,所以这个群体有在实现可持续发展目标的进程中被落下的风险。需要采取一些行动来确保实施适当的政策、措施和计划,最终防止这种情况的发生:
▸ 需要更加明确地界定对森林的依赖,以识别生活在森林内和森林附近的人以及在某种程度上依靠森林资源谋生的人。
▸ 国家和国际层面的人口普查和其他住户调查都需要对生活在森林地区内及其周围的人口进行充分的抽样调查,即便抽样成本因这些地区地处偏远而有所增加。
▸ 在已有的问卷调查中,有关依赖森林的人口和社会经济数据应被单独分类列出。
▸ 需要制定标准指标来衡量依赖森林的人群的贫困状况,可以基于其收入与国际贫困线的相对关系(如SDG具体目标1.1 1)和各国制定和调整的贫困指数(如SDG目标1.2 2)。理想状况下,后者应基于整合了森林特有因素的多维度标准,例如森林资源对生计的直接贡献以及传统森林社会有时较高的社会资本和非正规社会保障机制。
森林合作伙伴关系(CPF)已经制定了一套包含21项与森林有关的指标的全球核心指标体系,来支持《2030年议程》(特别是SDG 15 — 陆地生命)和联合国《2017-2030年森林战略计划》(联合国,2017),当前正在研究实施方法。目前的工作重点是那些在数据收集上存在一定挑战的指标,特别是社会经济指标,包括“依赖森林的极端贫困人口数”。
在发展中国家,木质燃料(薪柴和木炭)对家用和出售都尤为重要。全球估计有8.8亿人至少花一部分时间用于收集薪柴或生产木炭(粮农组织,2017a)。超过4000万人(占全球劳动力的1.2%)从事与薪柴和木炭生产相关的商业活动,来供应城市中心的需求。木质燃料的生产在2011年创造了330亿美元的全球收入。因此,木质燃料生产的可持续性至关重要。
木材和非木质林产品(NWFP)为发展中国家拥有中等及良好获取森林资源条件的农村家庭提供了大约20%的收入(Angelsen等,2014)。如果将直接、间接和衍生就业都计算在内,正规森林部门据估计每年在全球范围内提供4500万个就业机会,劳动收入超过5800亿美元(粮农组织,2018b)。中小林业企业(SMFE)约占2000万个工作岗位,每年创造1300亿美元的价值。全球范围内,2015年采集的非木质林产品总值约为80亿美元(粮农组织,2020)。这些估计值可能远低于实际数字,因为全球大多数林业部门活动为非正规经济活动,并没有在国家统计中得到很好的计量和体现。
非正规部门被定义为非商业性、自给性的或者不受监管和未经报告的小型生产单位。据估计,2011年非正规林业部门创造了1240亿美元的收入,为约4100万人提供了就业机会(粮农组织,2014c)。非木质林产品在此尤为重要,它为全世界数亿人口,尤其是妇女、儿童、无地农民、土著居民和其他处境脆弱的人们提供了粮食、收入和营养多样性(见插文25和粮农组织,2018b)。食物、药用植物、工艺材料、其他非木质林产品和木质燃料的采集是妇女对家庭生计做出贡献的重要组成部分。在一些偏远地区,出售非木质林产品所获收入是妇女可获得的唯一现金来源(Shackleton等,2011)。
森林生物多样性的非消耗性用途(如休闲和旅游),也是农村现金经济中越来越重要的组成部分(Hegetschweiler等,2017)。每年,据估计人们约有80亿次对保护区(许多被森林覆盖)进行游览,相应的国内消费支出约为6000亿美元(Balmford等,2015)。
此外,森林生物多样性为数亿人口在生计艰难时期提供了粮食、能源和收入安全网(Sunderlin等,2005),尽管有些学者(如Paumgarten、Locatelli和Witkowski,2018)指出该功能可能受到季节性波动的限制,在极端事件期间可用性会降低。
城市居民长久以来受益于各类木材产品和非木质林产品,从纸和家具到蘑菇、森林水果和野生动物。很大比例的城市贫困人口依靠薪柴和木炭来烹煮食物,尤其是在非洲(参见如Mulenga、Tembo和Richardson, 2019)。在较为繁荣的经济体中,城市居民对来自森林的食品、化妆品和其他产品表现出越来越大的兴趣,譬如像巴西棕榈果(Euterpe oleracea)和猴面包树(Adansonia digitata)等林产品出现在了全世界超市的货架上或最新潮厨师的食谱中(如McDonell,2019)。此外,在发达国家和发展中国家,越来越多经济富裕人口选择至少一部分时间在林区生活,生物多样性是吸引他们做出这样选择的主要原因之一,被称为舒适迁移(Gosnell和Abrams,2011)。
土著居民的生计在很大程度上取决于森林生物多样性,然而随着他们与国家和全球货币经济的联系日益紧密,这种关系也处在不断变化的过程中。由土著居民管理的地区(目前约占全球陆地面积的28%)包括一些生态最完整的森林和许多生物多样性热点地带(Garnett等,2018)。土著社区经常与其祖先的林地有着深厚的文化和精神联系,并拥有关于生物多样性流传已久的知识(Verschuuren和Brown,2018年),其中许多知识正濒临失传(Camara-Leret、Fortuna和Bascompte,2019)。在许多经济评估中,森林及其生物多样性对人们幸福感的认知和感受这样的无形贡献都被低估了。
世界最贫困人群在不同程度上依赖森林(Sunderlin等,2005;Camara-Leret、Fortuna和Bascompte,2019),但是通常比富裕些的人群更依赖生物多样性和生态系统服务(Reid和Hu,2005;《生物多样性公约》,2010b)。在森林覆盖率高和森林生物多样性丰富的中低收入国家,人口较少的这些地区的贫困率往往较高(Fisher和Christopher,2007)。粮农组织(2018b)估计,有2.52亿人生活在森林和大草原中,每天收入低于1.25美元。总的来说,这些农村贫困人口中约有63%生活在非洲,34%生活在亚洲,3%生活在拉丁美洲。拉丁美洲800万依赖森林的贫困人口约占该区域农村极端贫困人口的82%。
了解贫困与森林之间的关系对全球消除贫困和保护生物多样性具有至关重要的意义。人类与森林之间的关系受到复杂、变动的、有时是彼此相左的因素的影响(如Busch和Ferretti-Gallon,2017)。了解社会和经济因素与环境结果之间的关系是一项艰巨的挑战(Ferraro、Sanchirico和Smith,2019)。
一方面,减贫和收入增长可以增加对土地密集型商品和生产的需求,加大人类将森林转变为牧场、农田和生活空间的需求。另一方面,收入的增加可能会改变就业方式,趋向于远离土地密集型生产,增加对娱乐和环境质量的需求,并增强人们保护自然的能力和意愿。这些因素的影响是通过制度和政策条件过滤并形成的(Deacon,1995)。
Alix-Garcia等(2013)对墨西哥和Heß等(2019)对冈比亚关于收入增长与毁林之间因果关系的研究表明,基于一定条件的现金支付计划和社区驱动的发展计划引起的收入增长均导致了森林的丧失。与之不同,墨西哥和乌干达的研究表明,对森林保护行动提供补偿的付费计划成功地降低了毁林速度(Alix-Garcia等,2015;Jayachandran等,2017)。
一系列社会和经济因素与森林覆盖和贫困相互作用,影响着它们之间的关系。这些因素包括农业扩张、人口增长、交通基础设施、技术变化、获取信贷和国际贸易。交通基础设施就是此类互动的一个很好的例子。公有和私有林一般地处偏远,经常与服务对象和市场毗连性差;这一状况又因许多森林人口是少数族裔或土著居民等社会边缘化群体而进一步恶化。新建和条件更好的道路可以降低开发森林资源的成本,扩大当地林产品的市场,但同时也可能为林区居民提供更多的经济机会和社会服务,减少他们对森林的依赖。
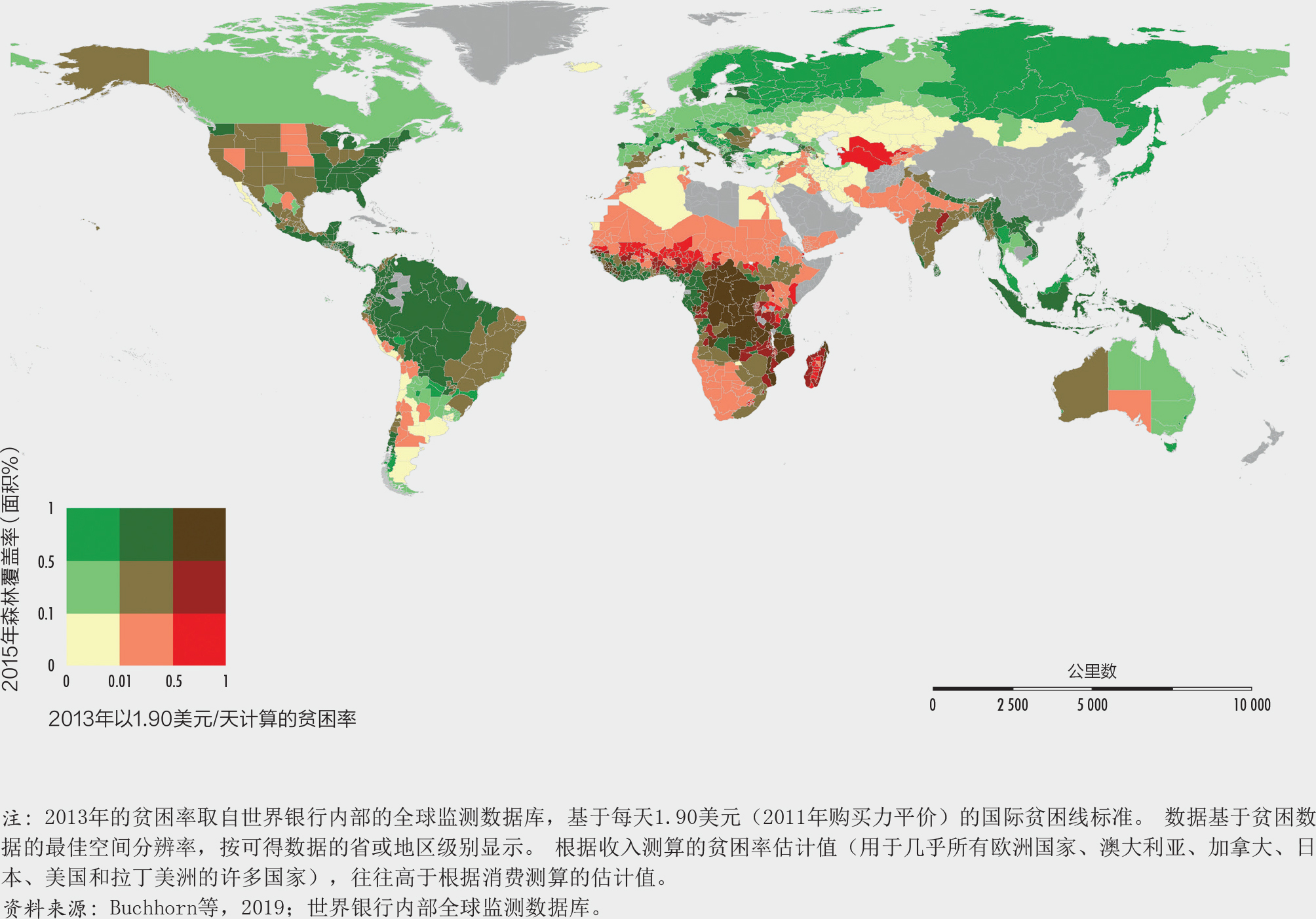
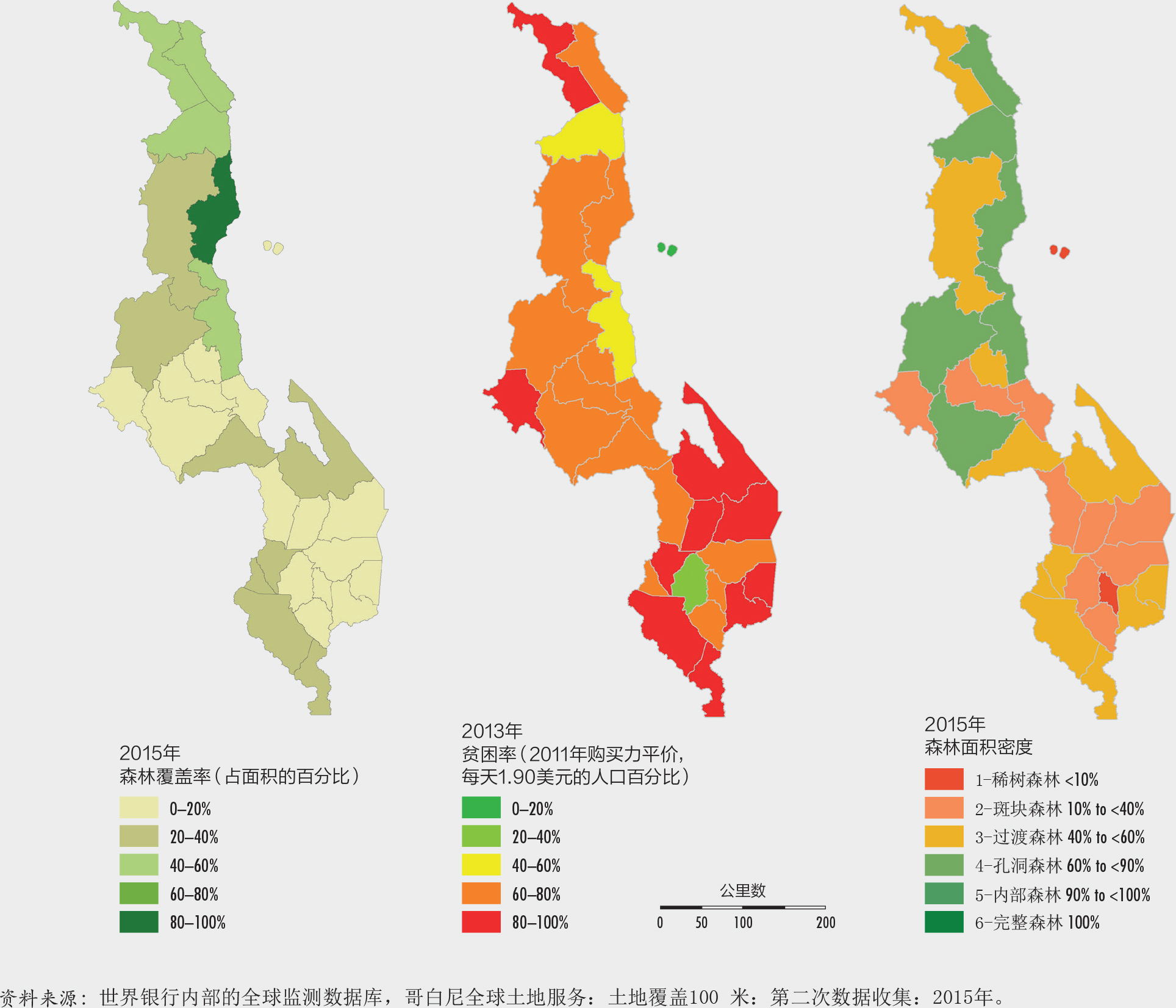
世界银行为本版报告进行的一项研究发现,贫困与森林覆盖之间的关系存在极大的异质性(图25)。中部非洲的贫困率高同时森林覆盖率也高,而欧洲和北美的许多地区贫困率低但是森林覆盖率高。由于地区级贫困数据的存在,马拉维成为一个独特的案例(图26)。这里的地理信息分析表明贫困与森林完好程度之间存在负相关,该国南部地区的森林密度较低(用作衡量森林完整性的变量),但贫困率较高。
虽然这样的结果无法推断因果关系,但仍然有助于为旨在结合发展和保护的国家计划和战略指明优先干预的地域。将来,获得分类细化的空间数据将会变得更为容易,理论上讲利用多维度的指标也可以更好地反映森林的状况,这些改善均有利于更深地了解贫困和森林之间的因果关系。
粮农组织(2009)将粮食安全定义为所有人在任何时候都能通过物质、社会和经济手段获得充足、安全和营养丰富的食物,满足其膳食需要和饮食偏好,过上积极健康的生活。根据这一定义,粮食安全应被理解为具有四个维度:可供量、获取、利用和稳定性。
森林和森林以外的树木(包括混农林业系统中的树木,农田中的其他树木以及农村和城市中非森林景观中的树木)通过提供营养食品、收入、就业、能源和生态系统服务来对粮食安全的四个维度做出贡献(粮农组织,2013a;粮农组织,2017b;高专组,2017)。因此,森林的枯竭或退化可能对粮食安全和营养产生负面影响。森林大规模地转化为其他土地用途,特别是用于农业,可能会在短期和中期改善这些农民和社区的粮食安全,但也可能对人们的环境、生计和粮食安全产生长期的负面影响;这些影响将主要影响森林社区当地居民,但也会影响全国和全球居民。此外,由于森林损失而造成的生物多样性和生态系统服务的丧失的长期影响很可能导致农业生产力下降。因此,森林对粮食安全和营养的贡献,需要我们对多数国家的森林政策给予更直接的关注。
食物的可供量(食物的实际或潜在供应)。在全球范围内,大约有10亿人在某种程度上依赖野生食物,例如野生动物、食用昆虫、食用植物产品、蘑菇和鱼类(Burlingame,2000)。一些研究表明,在发展中国家,这些家庭的收入倾向于处于最低水平(Angelsen等,2014)。尽管来自森林的食物据估计仅占全球食物消费的不到0.6%(粮农组织,2014c),但在许多地方对于确保人们获得营养丰富的食物以及重要的维生素和微量元素来说至关重要。
森林和森林以外的树木还通过提供牲畜食料(作为嫩叶或动物饲料)来支持食物供应。饲料对粮食供应的贡献是双重的:牲畜是肉和奶的来源,并且通过提供畜力和肥料来支持农业生产,而肥料可以提高农地的生产力。
农林牧系统中的森林和树木通过调节水循环和微气候、提供遮阳和防风、保持土壤、养分循环、生物虫害控制和授粉等生态系统服务来支持农林牧渔生产活动(Reed等,2017)(参见插文26中的实例以及森林生物多样性与可持续农业”)。它们在应对和减缓气候变化风险中的作用对于确保许多地区的粮食供应至关重要(见第5章案例研究1,关于旨在增强非洲小型农户和牧民的复原力而进行的大规模旱地恢复)。
亚马逊河下游洪泛平原的森林,在这些生物多样性极为丰富的生态系统中,为大量捕捞湖泊和河流中的鱼类做出了贡献,这些区域的鱼类丰富度据发现与森林面积直接相关(Lobón-Cerviá等,2015;Castello等,2018)。在尼日利亚,森林覆盖密度与村庄的新鲜鱼类消费量成正相关(Lo、Narulita和Ickowitz, 2019)。内陆渔业对全球粮食安全的贡献远远超过了以往认知,是动物蛋白和必需微量营养素的主要来源,特别是在发展中国家。例如,小鱼是维生素A、铁和锌的重要来源,通常比大鱼、其他肉类或蔬菜更易负担得起,也更容易获得(Kawarazuka和Béné,2011;Fluet-Chouinard、Funge-Smith和McIntyre, 2018)。
食物的获取。如4.1节人们受益于森林和生物多样性所述,正规和非正规森林部门(包括木材、木质燃料和非木质林产品的采获、加工和销售)是就业和收入的重要来源,因而为获取粮食提供了经济保障。尽管在全球范围内,林产品对家庭收入的现金贡献可能不大,但对于在正规和非正规森林部门中超过8000万的人们而言,对他们的生计、粮食安全和营养至关重要。得到保障的林权和资源权对于充分实现采集和销售林产品带来的经济利益十分关键,从而对依赖森林的人们的粮食来说非常重要。
尽管按性别分类的数据有限,但研究表明,农村妇女在可持续采集非木质林产品和收集薪柴中发挥着核心作用,并全年依赖这些销售收益(粮农组织,2014d;高专组,2017)。虽然在改善非木质林产品的数据上已经有许多努力,但还是需要更多的信息来更准确地估计这些产品在哪些方面以及对谁在粮食安全和营养中起着关键作用(粮农组织,2017c)。
由于与森林社区的紧密联系以及对与森林有关的生计的关注,中小型林业企业对许多农村社区具有特殊的增强粮食安全和营养安全的潜力。实现这种潜力通常取决于克服挑战,例如有限的地方能力、官僚主义的规章制度、不平等的地方权力结构、权属不安全以及地方精英攫取利益。
食物的利用(消耗足够的营养和能量)。烹煮是确保食物营养得到吸收的主要方法,全世界约三分之一的人口(24亿人)使用木质燃料烹煮食物,而全球约十分之一的人使用木质燃料烧煮和消毒饮用水,以确保饮用水和食品加工的安全(粮农组织,2014c)。作为在食品利用中使用林产品的另一个例子,鼓槌树的粉状种子(Moringa oleifera)还因其抗菌特性而用于净化家庭用水(Delelegn、Sahile和Husen,2018)。木质燃料还用于食品保存的加工过程中,例如熏制和烘干,这些过程可延长非生产时期的食物资源供应,并使其可以在更广阔的地区分布使用。
然而,使用木质燃料也会带来负面影响,包括森林退化和烟尘对人类健康的危害(插文27)。由于在中期未来内,木质燃料可能仍将是世界上相当一部分人最负担得起的能源,因此,确保木材的可持续利用和有效利用十分重要。
在许多发展中国家,特别是在非洲和南亚,木质燃料仍然是贫困农村家庭烹煮食物时使用的主要燃料。若不进行烹煮则可能是生食,因此木质燃料的贡献绝对是这些人群粮食安全的核心要素。随着时间的推移,大多数区域的木质燃料消费量已经下降或保持稳定,但在撒哈拉以南非洲,木质燃料的消费量仍在增加。由于习惯、口味、习俗和经验,即使在有替代能源的情况下,薪柴也被视为首选(粮农组织,2017a)。
尽管木质燃料占全球伐木消耗量的近50%,占非洲伐木总量的90%以上(粮农组织,2019e),但由于对森林使用没有加以节制,仍有三分之一的木质燃料以不可持续的方式被采伐(粮农组织,2017a)获取木质燃料是造成森林退化的常见原因之一。
如果使用不当,木质燃料还会引起烟尘污染(通常由于烹煮过程中燃烧效率低下而致),对健康产生负面影响。据估计,每年有近400万人死于因使用固体燃料烹煮和低效的烹煮方式而导致室内空气污染的疾病(清洁厨灶联盟,2015;世卫组织,2018a)。改进的灶具系统可以减缓这种风险,还可以减少所需的燃料量。
森林及其蕴含的生物多样性还提供了具有广泛常量和微量元素的食物,为维持地方居民的营养状况做出贡献。野生食品通常含有大量的关键微量营养素。例如,森林水果是矿物质和维生素的丰富来源,而森林中收获的种子和坚果则增加了膳食中的热量、油脂和蛋白质。野生食用块根和块茎是碳水化合物的来源,而蘑菇则是硒、钾和维生素等重要营养素的来源。树木和灌木的叶子(新鲜或干燥)是最为广泛消费的林产品之一。它们是蛋白质和微量营养素(包括维生素A、钙和铁)的丰富来源,而营养脆弱的人群的膳食通常缺乏这些营养。此外,全球维生素C和A、钙和叶酸的大部分供应来自由动物授粉的农作物(Eilers等,2011)。研究表明,森林覆盖与膳食质量之间存在密切联系(插文28)。
对森林和基于树木的系统的可得性与水果和蔬菜的消费以及膳食多样性有关,而森林的丧失与当地膳食的营养质量下降相关(Ickowitz等,2014)。个人或家庭的膳食多样性(在一定时期内食用的不同食物或食物组的数量)可以作为一项衡量营养状况的指标,包括微量营养素摄取是否充分、能量和儿童的成长(Jamnadass等,2015)。在坦桑尼亚的一项研究中,食用更多森林食物与更高的膳食多样性、食用更多肉类和更高营养膳食呈正相关的关系(Powell、Hall和Johns,2011)。Ickowitz等(2014)将21个非洲国家的树木覆盖卫星图像与其膳食信息匹配叠加,发现在树木覆盖率最高达到45%的情况下,树木覆盖较高的地区儿童膳食的多样性较高;对水果和蔬菜的消费量随树木覆盖增加而增长。同样,在非洲的27个国家中,森林与至少25%儿童膳食多样性的增加相关(Rasolofoson等,2018)。
森林覆盖的下降也会对营养产生负面影响。在一项对撒哈拉以南非洲15个国家的地理分析中,Galway、Acharya和Jones(2018)观察到毁林与幼儿膳食多样性减少之间的联系,特别是豆类、坚果、水果和蔬菜消费量的减少。他们发现西非这种关系最为明显。
粮食安全的稳定性(任何时候无风险地获取和利用粮食)。在季节性粮食短缺以及饥荒、农作物歉收和经济、社会及政治变动期间,从森林获取的收入和野生食物充当了一个安全网(粮农组织,2017b)。从森林中获取粮食是应对粮食不安全时期的一项重要战略,尤其是对于生活在森林中和森林附近的脆弱家庭而言。当传统农产品不可用以及库存不足或资金短缺时,林产品通常可以供应相对较长的时期,包括在“饥饿”或“缺粮”的季节(见插文29中西非案例)。
在西非,人们通过发酵非洲刺槐豆(Parkia biglobosa)来获得富含蛋白质(占干物质的40%)和脂肪(占35%)的营养食品,无需冷藏即可保存超过一年(粮农组织,2016a)。豆类在干旱季节成熟,因此在新作物收获之前的传统“饥饿季节”中提供了宝贵的食物。由于豆子不进入常规商品贸易,因此难以获得年度产量数据,但据估计仅在尼日利亚北部每年就收获20万吨非洲刺槐豆(Nwaokoro和Kwon-Ndung,2010)。
在加纳西部地区,非木质林产品对于缺粮季(6月至8月)的家庭粮食安全、营养和健康尤为重要。据报道,低收入家庭每周有五至六次消费从森林里获取的食物,例如野生动物(包括大蔗鼠Thyoyonomys swinderianus)、蜗牛、蘑菇、蜂蜜和水果(Ahenkan和Boon,2011)。
在塞内加尔,某些树木的果实,例如常年结果的Boscia spp和在旱季末期结果的马鲁拉果(Sclerocarya birrea)常被用来丰富膳食,有助于解决季节性维生素不足问题(粮农组织,1989)。
除了提供应对短期粮食供应不稳定(可能导致严重的粮食不安全)的措施外,森林及其生物多样性还提供了生态系统服务,这些服务对于确保粮食供应的中长期稳定至关重要(利于防止长期粮食不安全),包括通过对可持续农牧渔生产的支持(见食物的可供量中的描述;另参见森林生物多样性和可持续农业一节)。森林作为粮食作物和药用作物的基因库保护了生物多样性,对于确保提高膳食长期质量所需的多样性至关重要。
森林食物占农村粮食不安全人群日常膳食的很小(以卡路里计)但很重要的部分,它也增加了主食的多样性。在一些消耗大量森林食物的社区中,仅野生森林食物就足以满足对水果、蔬菜和动物蛋白的最低膳食要求(Rowland等,2015)。
森林食物作为营养资源的价值不仅限于发展中国家。欧盟有超过6500万公民偶尔收获野生食物,至少有1亿消费可食用林产品(Schulp、Thuiller和Verburg,2014)。野生食物,特别是野生动物和其他林产品,在北美也常被食用(Mahoney和Geist,2019)。一些森林食物被广泛交易。例如,据估计每年全球食用蘑菇市场价值为420亿美元(许多采自森林)(Willis,2018)。
森林食物对土著社区在营养上(和文化上)具有特殊的重要性。一项针对22个亚洲和非洲国家(包括工业化国家和发展中国家)的研究发现,土著社区平均每个社区使用120种野生食物(Bharucha和Pretty,2010)。
在全球范围内,大量树种提供了重要的食物和营养来源(图27)。许多树种的多个部分可以食用。例如,猴面包树(Adansonia digitata)是多用途热带树种,不仅果实能食用,叶子也可以食用,在非洲干旱地区是人们经常食用的食物。每100克猴面包果的脱水果肉中,维生素C高达300毫克,含量近于柑橘的6倍(Odetokun,1996,引自Manfredini、Vertuani和Buzzoni,2002),同时还含维生素A、B1、B2和B6。儿童每日食用0-20克果肉就能满足其维生素摄入需求。猴面包树的树叶富含钙、蛋白质和铁(Mbora、Jamnadass和Lillesø,2008)。
同样,鼓槌树(Moringa oleifera)的叶子富含维生素B、维生素C、β-胡萝卜素、镁、铁和蛋白质。同时还含有酚类和类黄酮化合物,具有抗氧化、抗癌、免疫调节、抗糖尿病和护肝作用。仅5克叶粉就能满足三岁以下儿童维生素A每日总需求的60%(医学研究所,2001;Witt,2013)。
坚果。坚果是最富营养的人类食物之一,富含蛋白质、油脂、能量、矿物质和维生素。尽管坚果是高能量食品,但研究和临床试验显示,坚果会引起较强饱腹感,食用不会增加体重(或减轻体重),并且可以降低肥胖风险(见Liu等,2019)。EAT-《柳叶刀》健康饮食委员会(Willett等,2019)指出,实现到2050年向健康膳食的转变将需要大量膳食上的转变,包括将坚果、水果、蔬菜和豆类等健康食品的消费量增加一倍以上。尽管在某些西非人群中坚果的消耗量传统上就很高,但就总体人口而言,坚果是实际膳食摄入量与EAT-《柳叶刀》健康饮食委员会所建议的“健康”参考摄入量之间差距最大的食物。
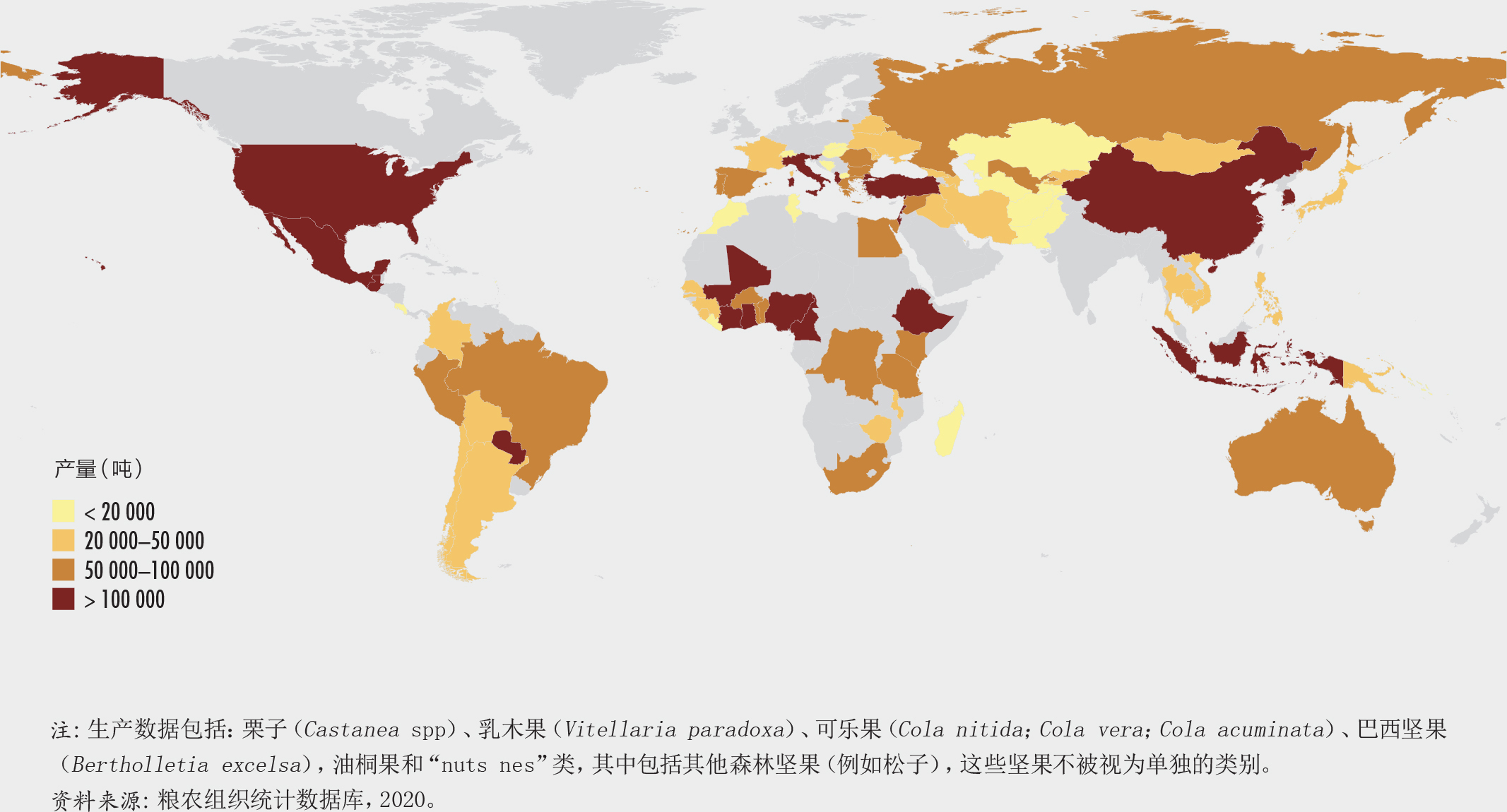
在许多国家,主要或完全来自森林的坚果年产量很大(图28)。一些坚果为农村社区和森林居民的自给生活做出了贡献,而另一些坚果(巴西坚果)则具有重要的商业意义(插文30)。清理土地后,人们经常将结食用坚果的树木和灌木丛留在农田和家园中。
巴西坚果(热带雨林树Bertholletia excelsa的种子)是目前唯一在全球进行贸易的、由专门的采收者从野外采集的树坚果。在过去的几十年中,采收巴西坚果为成千上万的农村家庭通过“以用促保”的方式保护千百万公顷的亚马逊森林做出了贡献。玻利维亚多民族国、巴西和秘鲁的坚果年出口价值达数千万美元,极大改善了当地居民的民生,促进了国民经济以及大范围区域的森林发展。树木也对当前实行的坚果采收类型和水平做出了强烈的回应。资源用户也已经发展了管理系统以维持生产力。
野味肉类。Redmond等(2006)列出了全世界将近1800种用作野味的昆虫、哺乳动物、鸟类、两栖动物和爬行动物,其中许多在热带和亚热带森林中。鉴于其中只有45%(约800种)是昆虫(其他来源表明已有1900种食用昆虫,参见下文),并且不包括鱼类和贝类,因此捕自森林的动物物种总数可能会更高。在一些农村森林社区和小城镇中,虽然人们获得支付得起的家养禽畜肉的机会有限,但是他们可以获得野生动物。对他们来说,野生动物通常是蛋白质和脂肪等常量营养素的主要来源(Sirén和Machoa,2008),而且也是例如铁和锌等重要微量营养素的来源(Golden等,2011)。最近对非洲、亚洲和拉丁美洲24个国家的近8000个农村家庭进行的一项调查发现,有39%的家庭捕获过野生动物,并且几乎都食用了野生动物(Nielsen等,2018)。在全世界至少62个国家的农村人口膳食中,野生动物肉至少占动物蛋白的20%(Nasi等,2008)。在亚马逊和刚果盆地,野生动物肉的消费量占当地人日常蛋白质需求的60%至80%(Coad等,2019)。研究表明,在森林食物消费量很高的地方,膳食中来自森林的肉、鱼、水果和蔬菜的比例可能高于来自家畜、水产养殖和农业的食物(Rowland等,2017)。相比之下,在家养禽畜肉类较便宜的城市中心,野生动物通常不会在粮食安全中发挥显著作用(Wilkie等,2016)。然而,在一些森林较匮乏的国家,城市中心可能对丛林肉有大量需求,特别是在家养牲畜蛋白质来源有限的地方(Van Vliet等,2019)。
当其他食物变得不易获得时(例如在经济困难、内乱或干旱时期),野味肉可能是蛋白质、脂肪和微量营养素尤为重要的来源(Coad等,2019)。
在城市中心出售野生动物也可能成为狩猎社区人们获取多样化收入的来源,尤其是在家畜蛋白质稀少或价格昂贵的地区(Nasi、Taber和Van Vliet,2011)。同样,其他野生动植物产品的贸易也可以为森林社区的人们提供现金收入,例如作为捕获动物副产品的兽皮。以秘鲁为例,全国每年平均出口41000张获得《濒危野生动植物种国际贸易公约》许可的供时装业使用的野猪皮(Sinovas等,2017)。
然而,随着城市化速度的加快,城市对野生肉类和野生动植物产品的需求正在推动狩猎的增加。供应商包括乡村猎人和其他地方的专业商业猎人。即使是较低的人均城市消费量也可能导致生境内野生动物摄取不可持续,尤其是当狩猎技术得到改进,野生生物生产力低下以及栖息地丧失和破碎化时(Fa、Currie和Meeuwig,2003;Coad等,2019)。
在一些野生动物对当地人生计至关重要但狩猎活动已变得不可持续的农村地区,除非能够在野生肉类商品链上采取可持续管理措施,否则野生物种数量的下降可能会对人类产生重大影响(Golden等,2011)(见第6章:保护和可持续利用森林和森林生物多样性)。重要的是,管理策略必须灵活,且与不同的利益、需求和优先目的有机整合,协调一致(Coad等,2019)。
昆虫。据估计,昆虫是至少20亿人传统膳食的一部分。据报道,已有超过1900种物种被用作食物,其中甲虫类(鞘翅目)占食用物种的31%,毛虫类(鳞翅目)占18%,蜜蜂、黄蜂和蚂蚁(膜翅目)占14%(粮农组织,2013b)。
尽管将食用昆虫作为一种商品资源进行管理具有巨大潜力,但过度捕食可能会带来保护和粮食安全问题,例如将Mopanie毛虫(Imbrasia belina)商业化(粮农组织,2013b)。其他挑战还包括缺乏立法和食品安全标准,然而情况正在逐步改善:例如,欧盟根据《新食品法规》认可了昆虫食品的合法性,该法规促进了以昆虫为基础的食品的市场化(Belluco、Halloran和Ricci,2017)。
培养食用昆虫已经成为减轻对野生物种的压力、在更大食物范围内加强粮食安全的一种方式。例如,在泰国,小规模昆虫饲养已经是行之有效的做法(粮农组织,2013c)。最近,肯尼亚和乌干达等国家已经成功建立了食用蟋蟀和蚂蚱的养殖模式。
养殖食用昆虫的价值和意义超出了其营养和经济价值,因为养殖食用昆虫对本已有限的资源(如土地、土壤、水和能源)造成的压力要比其他形式的畜牧生产少得多。例如,用黄粉虫(Tenebrio molitor)生产蛋白质比用牛肉生产蛋白质更为环保(粮农组织,2013b)。近年来,在如比利时、芬兰和荷兰等一些传统膳食不包括昆虫的欧洲国家中,养殖食用昆虫也已在环境、社会和经济层面上得到人们的接受(如Luke,2018)。
森林和农业生产系统经常有不同程度的交叉重叠。有时它们会像混农林业一样完全重叠。全球约40%的农业用地的树木覆盖率超过10%(Zomer等,2009)。
森林具有比农田更丰富的动植物多样性。这便于森林提供生态系统服务,从而对森林附近农业生产系统的生产力和复原力产生积极影响(Duffy、Godwin和Cardinale,2017;高专组,2017)。据估计,世界上可利用的淡水中有75%来自森林流域。这些水用于农业、家庭、工业和生态目的(MEA,2005)。
森林在缓解和适应气候变化方面也起着至关重要的作用,从而有助于预防与气候有关的粮食不安全。可持续经营的森林生态系统还有助于最大程度地减少因土壤侵蚀、山体滑坡和洪水而造成农业损失的可能性。
森林还为农民提供了源自本地的农业投入(例如饲料、纤维和有机物),从而降低了从更远的地方生产和运输这些投入的成本和不良的外部效应。
一些森林植物的生产已经转移到了农场(例如咖啡、可可和花生),但是森林生态系统仍然经常为适应和改良现有作物提供重要的遗传资源。森林是许多驯养牲畜和栽培农作物物种的野生近缘种(祖传或相关物种)基因库,这些驯养和栽培的物种因为高产和其他特征而被繁殖。驯养品种在遗传上可能是高度同质的,因此在受到生物和气候变化影响时比较脆弱。相反,野生物种在自然、多样化甚至有时是极端的条件下不断进化和多样化;与野生近缘种的杂交可能为驯养物种提供适应驯化的品种的来源。
森林为许多授粉者提供了栖息地,对于可持续粮食生产至关重要(见插文31中的例子)(另见第3章插文18:森林授粉者)。
坦桑尼亚与撒哈拉以南非洲的大多数国家一样,在很大程度上依靠农业来维持生计、获取收入和粮食安全。全国大多数农民都是小型农户,他们的自给自足和农业生产力依靠自然生态系统服务。一项国家评估显示,森林作为野生授粉者的自然栖息地对农业具有重要作用(Tibesigwa等,2019)。结果表明,依赖授粉者的农作物(占该国农作物绝大多数)受益于与森林毗邻,从而获得了可观的生产力(和收入)。随着农田和森林之间距离增加,这种收益呈指数下降,与森林距离超过2至3公里后这些额外收益就消失了。此外,森林覆盖的下降也导致了农作物收入的减少。该研究表明了保护森林在农业系统中的重要性。
115种全球主要粮食作物中有87种(约占75%)在某种程度上受益于授粉者进行水果、蔬菜或种子的生产,这些作物占全球粮食产量的35%(Klein等,2007)。这些授粉者中有许多可以在森林中发现。
然而,也有必要强调不可持续的农业给森林生物多样性带来的威胁。二十世纪末的农业转型提高了农作物和牲畜的产量,改善了粮食安全状况。但是这一转型依赖于加大农业投入,实现大规模集约化生产,因此对环境造成了严重危害,例如农药对水源的污染。目前,农业部门是导致全球73%毁林的主要原因(Hosonuma等,2012),引发生物多样性的严重下降(见第6章)。没有充分认识到森林及其提供的服务对农业的惠益(包括生物多样性)有时导致人们选择对生物多样性产生不利影响的管理措施,从而造成更大的损失。有利于生物多样性的土地使用方式有助于保持森林生态系统服务带来的惠益,提高农业生产力。在这方面,土著居民和本地居民的知识可以成为宝贵的资产(IPBES,2019a)(见插文32中的例子)。
位于干旱日益严重和反复发生的中国西南地区,哈尼水稻梯田是适应恶劣地形条件和抵御缺水状况的一个有极具启发性的例子。1300多年来,这些梯田已成为农民智慧的见证。2009年,一场严重干旱使该地区其他地方的农业生产产量锐减,而哈尼水稻梯田则为农民维持了正常水平的单产和水量。位于丘陵高处和高山地区的森林、以及梯田结构和巧妙的水资源管理方法,在增强当地抗旱能力方面发挥了核心作用(Min,2017)。没有来自森林的充足水源,就不可能种植不少于123个当地品种的哈尼水稻。实际上,森林在整个地区生态系统的稳定性和可持续性上起着至关重要的作用。
哈尼水稻梯田的适应能力取决于四个主要支柱:
“四合一”(森林-村庄-梯田-河流)景观管理。村庄和梯田高处山顶上茂盛的森林促进了水蒸气上升形成水滴,有助于水库和小溪中水的蓄积。森林拦截雨水并增强了土壤的蓄水能力。它们还有助于保护土壤、减少侵蚀、防止村庄受到滑坡的影响。
已适应水资源保护的森林物种。那里的森林主要由豆蔻(Alnus nepalensis)组成,豆蔻是一种在水含量高的土壤上生长良好的树种,其发达的侧根系统为易流蚀的土壤提供了一定的稳定性。
基于森林环境服务的高效灌溉系统。山顶森林所积聚的水和当地的地形为稻田提供了一种独特有效的灌溉方式(见图A)。林木的深层根系有助于降雨渗入地下水。此外,地表径流沿着森林、村庄和梯田顺势流下斜坡。森林地块不仅为稻田提供水,而且还提供了肥料,因为流水将来自森林的营养物带入梯田的各层。
中国云南省元阳县哈尼水稻梯田。
©粮农组织/Min Qingwen.森林是农民日常生活和文化的一部分。哈尼族人民崇拜自然并尊重树木,它们被视作保护和祝福人类的神灵。他们的信仰与森林在他们生活中的重要作用密切相关。森林提供了包括木材、薪材和药品在内的各类物品,同时也提供了生物多样性丰富的栖息地。每个村庄至少维护一个神圣的森林或“神灵”林地。这种与自然的文化联系是保护和保存森林的动力。
混农林业,无论是以农田中树木的方式还是以森林中农业的方式,都可以优化农业与森林和树木生物多样性之间的联系。对混农林业景观尺度方法的日益关注增强了其在生物多样性保护中的作用。混农林业在保护生物多样性中具有五个主要作用(Udawatta、Rankoth和Jose,2019):
▸ 为可以忍受一定程度干扰的物种提供了栖息地。
▸ 有助于保护敏感物种的种质。
▸ 通过提供一个比传统农业系统(有时意味着彻底清除自然栖息地)生产力更高和可持续性更强的替代方式,降低了自然栖息地的转化率。
▸ 提供了残余栖息地之间的连通性。
▸ 提供了如侵蚀控制和地下水补给一类的生态系统服务,从而防止了周围生境的退化和丧失。
森林、树木及其生物多样性提供了多种有助于人类健康的产品和服务,包括药品、食品、洁净水和空气、树荫、抑或仅仅是一个绿色的空间,人们可以在其中运动和放松(Nilsson等,2010)。森林或树木系统的生物多样性越丰富,可以提供的产品和服务范围就越广泛。
除上述森林和树木对营养和粮食安全(对人类健康至关重要)所做贡献之外,森林生物多样性还蕴含着许多具有已知或潜在药用价值的植物、动物和微生物原料。这些物质不仅在当地具有重要意义,而且还在全国和国际市场上得到商业化或用来合成新药物(大多数最初源自森林植物的活性化合物现在在实验室合成)。目前有超过28000种植物被记载有药物作用,其中许多是在森林生态系统中发现的(Willis,2017)。
在印度阿育吠陀医药、传统中医和其他土著医药体系中,采自森林的药材尤为突出。西药所依赖的许多药材均来自森林植物,并成为依赖森林的人群传统医药系统的一部分(Fabricant和Fransworth,2001)。例如,几个世纪以来,金鸡纳树皮(奎宁)是从金鸡纳属的几种安第斯树种中获得的,是世界上使用最广泛的抗疟药。它最初是经野生采集的,但后来从种植园种植的树木中获得。最终,奎宁被青蒿(Artemisia annua)提取物所取代,该提取物在中国药典中已有数千年的历史了。其他植物来源的药材有的通过药理筛选而成。紫杉醇就是一个例子,紫杉醇是一种生物活性化合物,最初来源于太平洋紫杉(Taxus brevifolia)的树皮,被视作取自天然原料的最好的抗癌药之一。
因此,世界各地依靠森林的人们的传统医学系统是一个重要的知识来源。世界卫生组织(世卫组织,2019)将传统医学定义为“在维护健康以及预防、诊断、改善或治疗身心疾病方面使用的,种种以不同文化所特有的、无论可解释与否的、理论信仰和经验为基础的知识技能和实践的总和。”这些系统对世界各地依赖森林的人们健康做出贡献,它们通常是最具可得性、可负担得起、有时在文化上较易被接受的卫生保健来源。世卫组织(2002年)指出,非洲多达80%的人仍然依靠传统医学来满足其基本医疗保健需求。据估计,至少有10亿人(不包括欧洲和北美)使用草药来治疗儿童的腹泻(粮农组织, 2014c)。2010年,基于传统医学和知识的世界草药市场价值为600亿美元(Nirmal等,2013)。
随着工业化的加速以及社会经济和文化潮流对当代土著居民社会的影响,以及世界生物、语言和文化多样性的下降,人们正在逐渐丧失有关药用植物及其益处的传统知识(Reyes-Garcia等,2013)。由于毁林、生态系统退化和这些知识的丧失,农村人口正在失去获得粮食和药品的机会,从而加剧了粮食不安全、营养不良和疾病。
显然,如《名古屋议定书》(《生物多样性公约》,2011)所认可的,保存和维护与森林生物多样性有关的传统知识,并保护农村居民分享和使用这些知识和资源的权利,对当地居民以及全球人类健康和身心极为重要。
越来越多的证据表明,与自然环境接触对社会各经济阶层和性别的人们的身心健康都有积极影响,特别是在城市地区(Triguero-Mas等,2015),尤其是对在社会经济方面处于不利地位的城市人口而言(Maas等,2006;Mitchell和Popham,2008)。在工业化国家和城市中,绿色环境可以激发人们进行体育锻炼(荷兰卫生委员会,2004),减少因久坐的生活方式而导致的健康问题,例如体重过重、长期压力和注意力疲劳。绿色空间还可以减缓精神压力,提高幸福感(Hartig、Mang和Evans,1991;Groenewegen等,2006;White等,2013)。据推测,接触大自然可以通过激发不耗费体力的无意识认知过程来减轻精神疲劳(Kaplan和Kaplan,1989)。然而,一些城市居民将自然绿色空间与不安全性联系在一起,暗示了对城市绿色空间进行仔细规划的必要性(Jorgensen、Hitchmough和Dunnet,2006)。
接触森林环境似乎也具有积极的生理作用,例如降低血压和脉搏(Tamosiunas等,2014),增强认知控制(Berman、Jonides和Kaplan,2008),甚至增强人类的免疫力(Li等,2008)。一些研究表明,居住在离自然和生物多样性丰富的环境较近的人们体内的微生物群更加丰富,特应性过敏的情形也较少(导致易感性过敏的因素)(Ege等,2011;Hanski等,2012;Rook,2013;Ruokolainen等,2015)。日本人认识到了“森林浴”(shinrin-yoku)(即身处大自然、吸纳森林气息的做法)的疗愈价值(Park等,2010;Hansen、Jones和Tocchini,2017)。
“森林学校教育”在斯堪的纳维亚国家长期以来很流行,现在在其他地方也被采用。它利用森林和树木作为儿童和年轻人发展身体、社会、认知和生活技能,建立独立和自尊的一种手段(O’Brien,2009)。在森林学校就读的儿童较少超重或肥胖,也较少出现多动症或感染传染病(Isted,2013;Blackwell,2015)。
世界上超过90%的人口生活在空气污染超过世卫组织准则上限的地方(世卫组织,2016),并且世卫组织(2018b)估计每年有700万人死于空气污染中的细小颗粒。森林仅通过改善空气质量就使整个人类受益(Nowak、Crane和Stevens,2006)。森林和树木有助于减缓许多在城市地区居住的问题,例如减少城市的热岛效应(Bowler等,2010;Shisegar,2014)(热浪可能致命)和缓冲隔离噪音(Irvine等,2009;González-Oreja等,2010)。鉴于森林和树木的这些以及其他好处,在澳大利亚、英国和美国等国家,一些领先的卫生政策已开始认识到利用自然来改善城市人口的健康(Shanahan等,2015)。例如,澳大利亚正在率先倡导“健康公园健康人类”的活动,这是全球运动的一部分,旨在释放自然和公园的预防和疗愈功能,同时保护生物多样性。
森林还可间接通过过滤水源并提供用于烹煮食物和烧开水的木质燃料,减少通过食物和水传播的疾病。这是至关重要的,以水传播的腹泻病为例,该疾病每年造成200万人死亡,其中大多数是五 岁以下的儿童(世卫组织/儿基会,2000)。此外,基于从森林和森林中收集的各种动植物食品为基础的传统饮食,可以减少二型糖尿病和肥胖症等疾病,因为这些食物大多是低脂肪、富含蛋白质和复合碳水化合物(Sarkar、Walker-Swaney和Shetty,2019)。
幸福不仅是单个人的状况,而且是更大范围一个社区的境况。许多人和社区,特别是土著居民,与特定的森林地区有着长期、世代的联系;他们不仅从森林中直接获益,而且还获益于与森林和本地物种之间无形的深厚精神联系,以信仰、习俗、传统和文化体现表达出来(Fritz-Vietta,2016)。
未能考虑文化价值的生物多样性保护举措可能对森林居民的个人和社会健康产生不利影响。例如,即使可以通过其他途径满足营养需要,限制获取某些传统上重要的食品也可能引起心理上的不安定,并影响其福利;例如,在刚果盆地的几个种族群体中,人们在没有丛林肉时会产生心理压力(Dounias和Ichikawa,2017)。
森林(特别是热带地区的森林)丰富的生物多样性蕴含着范围惊人的病原体、寄生虫及其媒介。人类大多数新的传染病是人畜共患的,这意味着它们起源于动物(Olival等,2017)。其出现可能与森林面积的变化和人口向森林的扩张有关,这两者都增加了人类与野生动植物的接触(Wilcox和Ellis,2006),而且在一些情形下也增加了人类消费野生动物的机会。与森林有关的疾病包括疟疾、查加斯病(又称美洲锥虫病)、非洲锥虫病(嗜睡病)、利什曼病和莱姆病(表4)。艾滋病毒和埃博拉病毒均为人畜共患病,又都是全球关注的焦点,它们有非常清晰的森林起源。与树木和森林有关的其他鲜为人知的病原体包括亨尼帕(Henipah)病毒,与此同时新的病原体还在不断涌现,例如导致当前COVID-19新型冠状病毒大流行的SARS-CoV2萨斯病毒。虽然尚不能确切确定人类最初是如何感染的,但COVID-19病毒也被认为是源于动物传染给人类所致(世卫组织,2020)。
大多数森林中的病原体并不构成对人类的直接威胁。许多潜在的病原体已与野生生物共同进化,不会对宿主造成健康问题,但如果传播到其他宿主物种(例如人类),则可能会成为问题。森林的改变可能会导致病原体宿主和媒介在丰度或分布上产生变化,而水文功能的改变则可能会有利于水生病原体(Wilcox和Ellis,2006)。因此,随着采掘业、毁林、栖息地退化以及人们对林地侵占的日益增加,新型病原体影响人们的风险也愈益增加。然而,有证据表明,生物多样性丰富的地区可能通过所谓的稀释效应使人们免受某些传染病的侵害(Rohr等,2019)。
十七种大型食肉动物曾被记录伤及人命。然而,其中只有五到六种似乎是经常性的,捕食动物对人类的袭击并不常见(Linnell和Alleau, 2016;Hart, 2018)。相比之下,每年有毒动物攻击多达250万人,造成2万至10万人死亡(世卫组织,2017)。蛇咬伤是任何森林活动中的职业危害。其他森林动物也可能造成人员伤亡。在亚洲和非洲,与大象的冲突每年导致数百人死亡(仅印度就因冲突事件每年造成400人死亡和100头大象死亡)(Shaffer等,2019)。世界范围内,通过以社区为基础的有创新性的自然资源管理计划、补偿体系以及激励和保险计划,为减少此类事件的发生做出了巨大努力(IUCN,2013)(另见第6章插文52)。
其他潜在的致命健康风险包括与伐木或在林间的其他操作有关的事故;立木倒下或树枝落下,尤其是在暴风雨期间更为频繁和危险;山火,特别是在近郊森林中发生时对人们及其房屋和企业造成的破坏最为严重,譬如2019年12月澳大利亚发生的森林大火。森林还含有会对人体有毒的过敏原(Cariñanos等,2019)、真菌和其他生物体。
这些问题均表明了负责任经营森林在保障人类福利方面的作用(McFarlane等,2019)。
鉴于人类、动物和环境健康之间有着千丝万缕的联系,“同一个健康”方法旨在通过在人类、动物及他们所处环境之间进行预防和减少风险来增进健康和福利。例如,在非洲,粮农组织、世界卫生组织和世界动物卫生组织(OIE)正在共同实施“同一个健康”计划,该计划将林业、自然资源、农业、畜牧和公共卫生领域的专业人士和政策决策者整合在一起,以确保所有相关部门和学科之间的平衡。
不仅在农村地区,而且在城市周边地区和城市地区,不论发达国家(例如插文33)还是发展中国家,都应当将实现人类最佳健康纳入森林经营和规划的目标之中。城市或农业扩张的土地使用规划还应考虑缓冲带的重要性,因为缓冲带将减轻因野生动物、牲畜和人之间较多接触而产生的潜在影响。
▸ 加强森林所有者、管理者、景观规划专家与其他部门的专业人士之间的合作,特别是公共健康、教育、体育、娱乐和旅游业。
▸ 通过与当地社区的沟通来鼓励公众参与性和包容性。
▸ 在评估提供森林生态系统服务的机制和资金时,应将人类福利作为生态系统服务概念的一个中心组成部分。
▸ 监控森林游客、他们对户外休闲的需求以及与健康相关的益处。
▸ 对研究进行投资,例如剂量-反应关系,从疾病中复原和恢复过程中的长期健康影响,以及森林健康效益的经济价值评估。
▸ 投资于包含森林多种功能的教育和培训,让从业者为新的绿色工作做好准备,这些工作将社会和健康纳入可持续森林管理。
▸ 增强森林的可获得性,以方便城市居民定期走近和接触森林。
▸ 积极沟通,以增进公众对与森林有关的决定的理解,减少森林地区使用和管理中的冲突。
迄今为止,对森林生物多样性威胁最大的是因毁林和森林退化而导致的生境和物种的丧失。
本章着眼于实现预防、制止和逆转第2章和第3章所述森林损失的措施。了解导致毁林或森林退化的因素有助于理解如何制止森林和生物多样性进一步丧失。在已经造成破坏的情况下,森林景观恢复可以开启扭转损失的局面。
长期以来,人们一直认为人口增长、人口构成趋势和经济发展是环境变化的主要驱动力。在过去的50年中,人口翻了一番,全球经济增长了近四倍。经济发展使许多国家的数十亿人摆脱了贫困。然而,全球大部分地区的自然状况在此过程中发生了巨大变化,伴随最多的是对生物多样性的负面影响,也常常对包括土著居民在内的社会最脆弱群体产生了最不利的影响。众所周知的主要压力包括:栖息地的变化、丧失和退化;不可持续的农业经营;入侵物种;包括非法采伐和野生动植物贸易在内的资源利用效率低下和过度攫取。气候变化和波动加剧了这些压力的影响。
全球市场压力、饮食偏好和农业价值链上的损失和浪费均驱动着人们对农产品和林产品的需求,而这些又引发了毁林和森林退化(IPCC, 2019)。总的来说,不断增长的人口对粮食和能源的需求是森林及其生物多样性丧失的主要诱因。在非洲,人口压力和贫困是森林保护的主要威胁,驱使贫困农民将森林转变为农田(Uusivuori、Lehto和Palo,2002;Lung和Schaab,2010),并以不可持续的数量获取木质燃料。在其他地方,毁林是由富裕人群的消费方式变化所驱使的。然而,毁林和森林退化实际上由许多政治和社会经济力量在全球和地方层面相互作用所驱动(Lambin等,2001;Carr、Suter和Barbier,2005)。
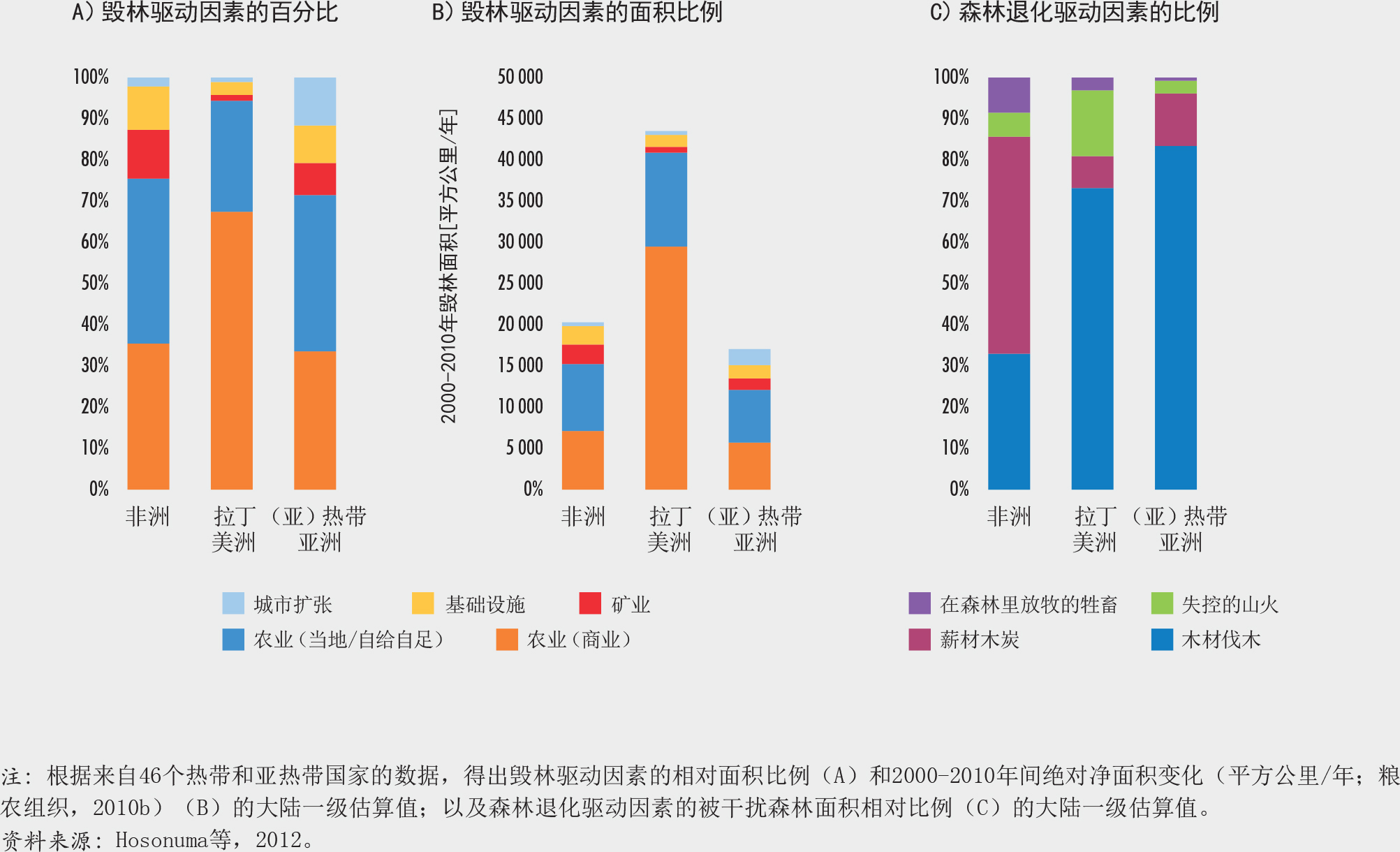
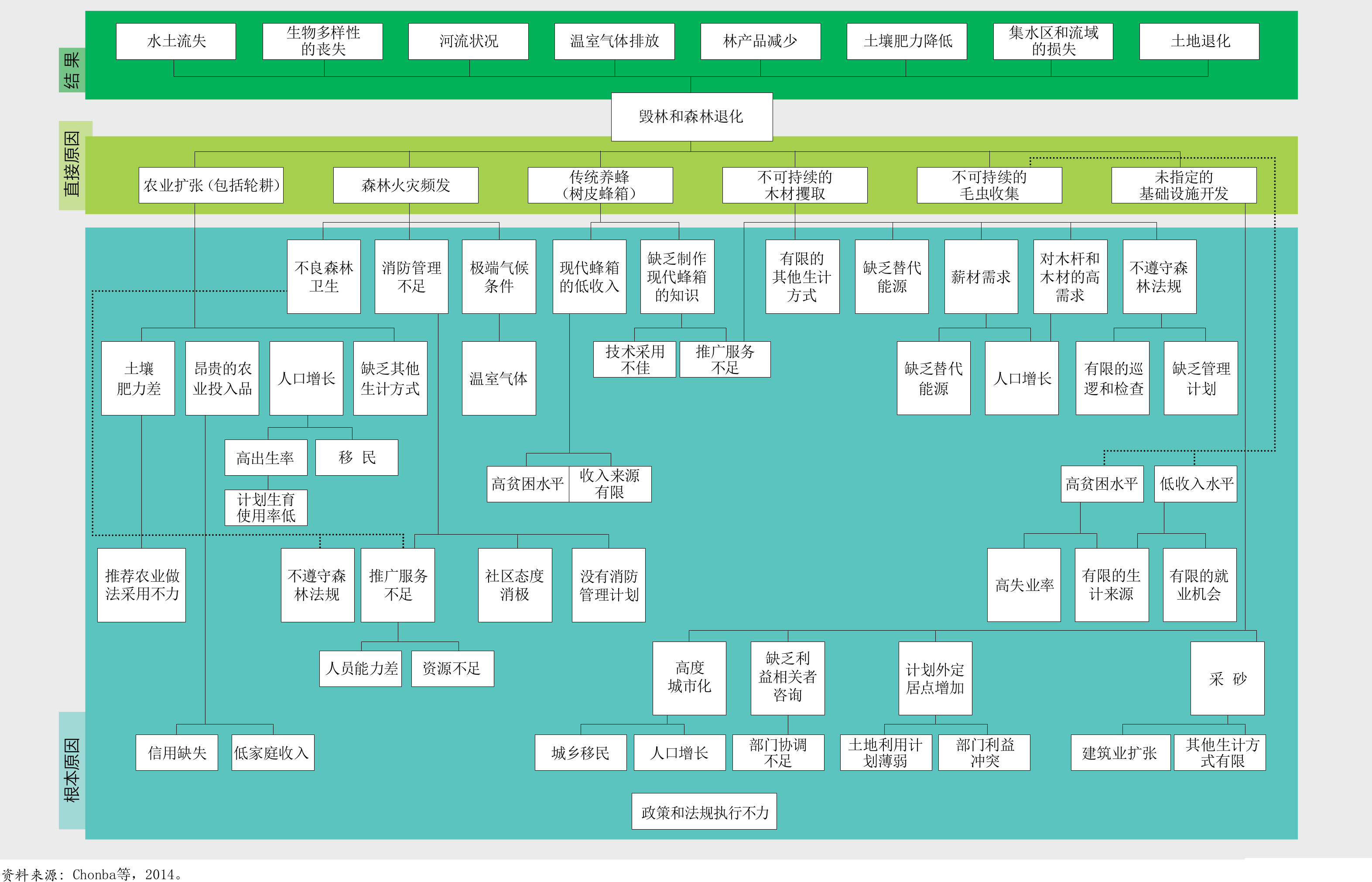
对46个热带和亚热带国家(约占这些气候地区森林面积的78%)进行的一项数据分析表明,大规模商业化农业生产(主要是养殖牛、种植大豆和油棕)是毁林最普遍的原因,约占40%(Hosonuma等,2012)。据估计,当地自给农业占毁林的33%,城市扩张占10%,基础设施建设占10%,采矿业占7%。在某些情况下,森林退化发生在土地使用变化之前,例如由不可持续或非法采伐木材引起的森林退化。该分析还显示,驱动因素在区域之间(图29)甚至在国家内部也存在显著差异。
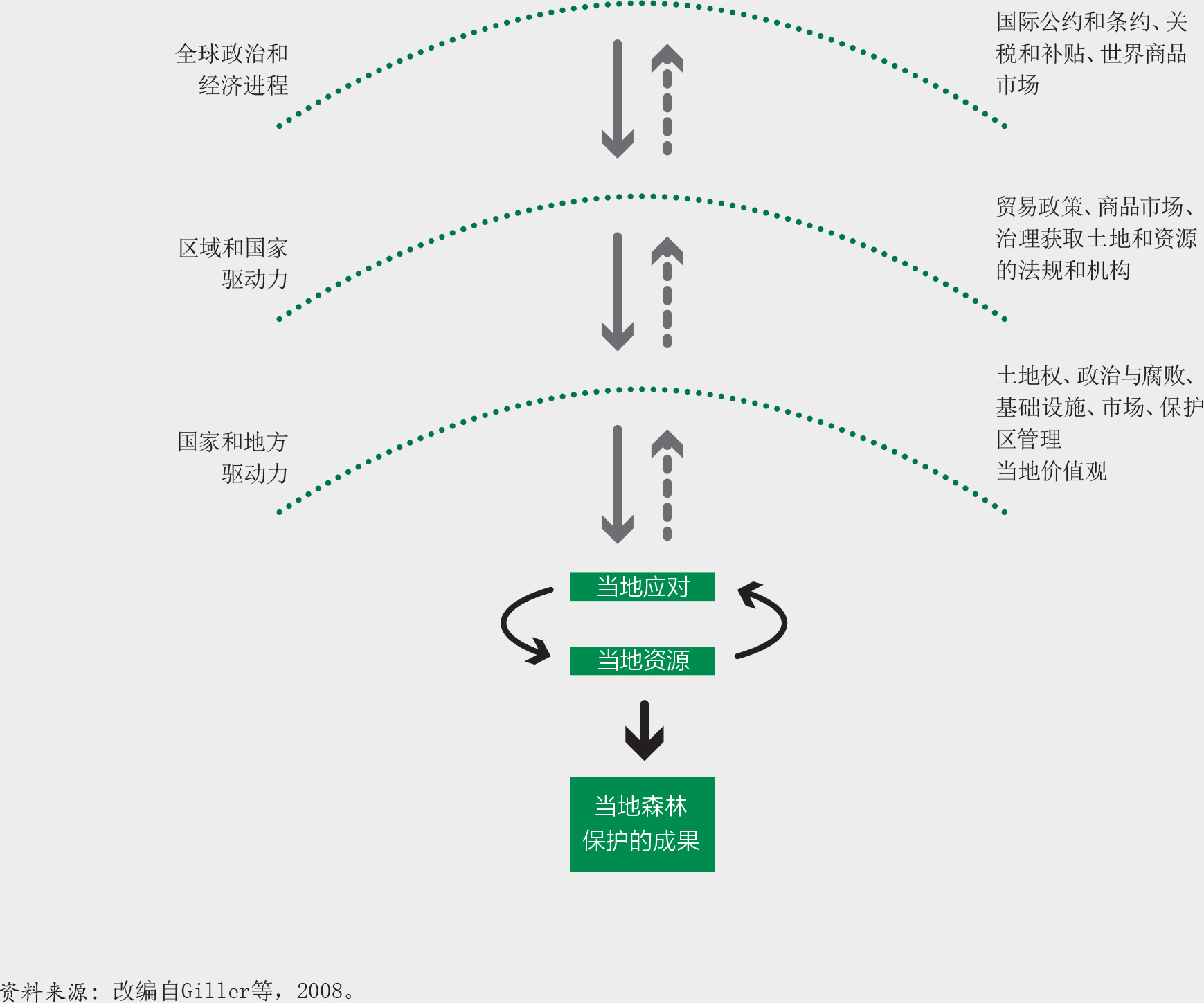
人们对资源的使用在很大程度上取决于获得的收益,并与获取时发生的成本或制度障碍导致的成本进行比较权衡(Schweik,2000),同时也受到地方和历史因素不同层面的影响,例如对传统森林所有权和习惯管理的认可以及实际操作措施、保护区协议在本地的实施、当地道路状况、商品价格和文化偏好。了解在不同层面驱动因素相互作用下的当地环境(包括全球和全国的政治和经济状况、资源管理的体制框架、利益相关者的价值诉求、以及资源的生态特征[图30])能有助于管理者做出知情决策(Ostrom和Nagendra,2006)。
如插文34中的案例所示,简单的森林变化驱动力模型并不能反映出当地复杂的社会和生态现实状况。它们导致简化的政策措施,基于这些措施的干预也因此常常无法实现其目标(另参见Nel和Hill,2013及Molinario等,2020)。重要的是要考虑到森林变化的潜在环境和动因,并认识到它们在影响当地人民决策中的重要性。各地影响人们进行森林可持续经营的激励措施各不相同,因此不能在全球范围内统一设计。
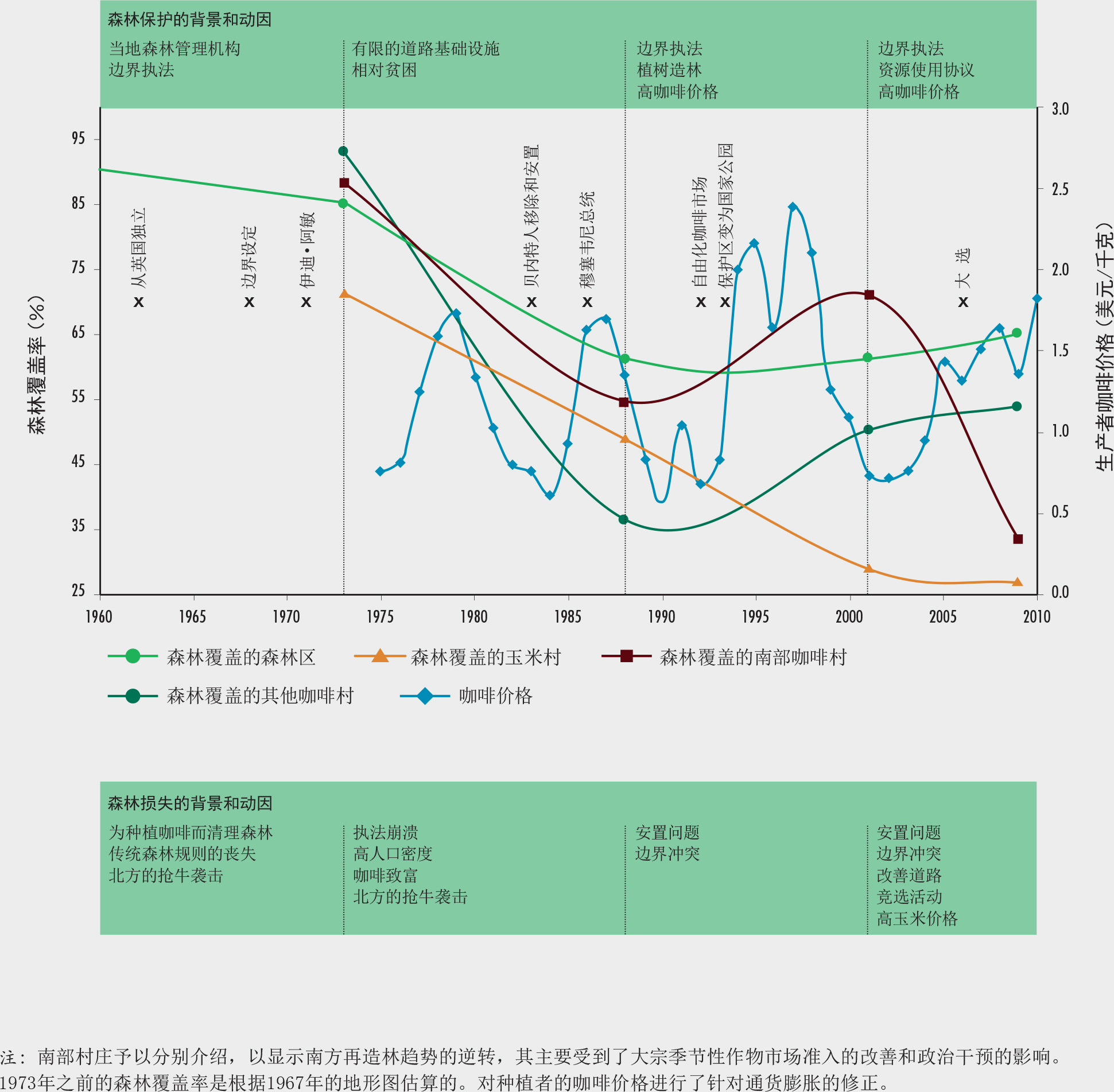
乌干达的埃尔贡山(Mount Elgon)体现了人口稠密地区在生物多样性保护上的挑战。它的森林为当地社区提供了木材、薪材、非木质资源和森林服务,由于该山是该地区的主要水源,因此它在水文上的贡献尤其突出。森林也是农业用土地的来源。埃尔贡山历史上在各种或多或少的排他性管理制度下都曾得到保护。高人口密度(每平方公里多达1000人)对森林资源的压力日益增大。资源获取和使用方面的冲突司空见惯(Norgrove和Hulme,2006;MERECP,2007)。
在1973-2009年间,该地区丧失了超过25%的森林覆盖,但是在某些地方森林也有所恢复(Sassen等,2013)。Sassen(2014)结合遥感技术和实地考察,研究了各种因素(包括土地使用目标、财富水平、市场准入状况、以及与公园管理的关系)在整个公园和整个时期内的变化如何对森林产生不同的结果。
该研究发现,在过去36年中,埃尔贡山的人口密度、贫困状况与农业扩张和毁林之间不存在简单直接的关系。人口数量仅在极少数情况下导致毁林,例如,二十世纪70年代和80年代间保护区管理机构崩溃,那里的人们开始通过种植咖啡致富。当保护区边界被重新建起后,森林在一些人口最密集的地区附近得到了恢复;这些包括居民可以进行农业集约化投资的地区、尽管市场准入有困难但产品是易运输的经济作物(咖啡)的地区、以及与公园管理几乎没有冲突的地区(参见图A中1988年后“其他依赖咖啡的村庄”附近森林覆盖的趋势)。总体而言(尽管这也取决于具体情况),与贫困相比,追逐财富(用资产衡量的)更可能导致毁林。上世纪90年代在森林外重新安置了牧民,并且鼓励他们从事农业劳动进行谋生(玉米),尽管人口密度较低,这些政策仍然导致了冲突和森林大面积被侵占(见图A中“以玉米为基础的村庄”附近森林覆盖的趋势)。价格高昂的经济作物与毁林相关主要发生在有良好的市场可以进行大宗季节性农作物交易(例如玉米、卷心菜和土豆)的地区,或对于公园边界的界定存在冲突的地方(即图A中2001年以后的“南部咖啡村”)。
森林退化也因当地对土地使用的需求(例如对固定香蕉和豆类植株材料的需求,或对供牛群吃草的土地的需求)和市场准入状况(例如出售木炭的机会)而变化。该研究还发现,在社区管理协议下允许收集薪柴等森林资源有可能成为一把双刃剑。一方面,为破坏性活动创造了机会。另一方面,可以帮助改善当地人与公园工作人员之间的关系,从而促进更优的管理措施,达成更好的森林效果。
这些发现表明,基于单一毁林驱动因素(例如人口或贫困)的简单模型无法解释在森林保护结果上的局部差异。相反地,正是在当地状况(例如执法、协作管理、政治干预)的作用之下,诸如人口、财富、市场准入和商品价格等驱动因素才随着时间的推移影响着森林覆盖、退化或再生的结果,而不是驱动因素本身独自构成影响。这一理念对设计更适合当地情况、且在生态和社会上可持续的管理措施具有重要意义。
了解对森林构成干扰的人类活动有助于在REDD+的框架下制定政策和行动,明确毁林和森林退化的驱动力通常是制定REDD+战略和行动计划的第一步。图31中赞比亚的例子则展示了诸多驱动力之间的互动关系。
过去十年来,打击毁林的行动加快了步伐,这主要是因为人们认识到森林的消失和用火来开地正在对全球碳循环产生着不利的影响。REDD+(减少毁林和森林退化造成的排放以及发展中国家的森林保护、持续管理和森林碳储存的作用)现已作为一项行动建议被列入《巴黎协定》。一项对31个REDD+战略的分析(粮农组织,即将出版)展示了导致毁林和森林退化的直接和潜在驱动力在国家内部以及国家之间的多样性(图32)。迄今为止,已有九个国家向《联合国气候变化框架公约》报告了毁林的减少,这意味着减少了约90亿吨二氧化碳排放(插文35)。各国现在正在从绿色气候基金和其他类似机制获得REDD+基于结果的支付奖励 — 减排奖励。许多国际举措为这些努力提供了支持,包括由粮农组织、联合国开发计划署(开发署)和环境署共同运作的联合国减少毁林和森林退化所致排放的计划(UN-REDD)(插文36)、森林碳伙伴基金以及世界银行的森林投资计划。
REDD+使发展中国家能够通过减少毁林和森林退化实现减排,可持续管理森林,养护和增加碳储,经核实结果之后可以获得资金。《联合国气候变化框架公约》关于REDD+的框架包括查明毁林动因,制定REDD+战略和行动计划。同时还包括一系列环境和社会保障措施,例如确保所采取的行动与天然林和生物多样性的保护相一致,并尊重土著居民和地方社区的知识和权利。
迄今为止,已有50个国家提交了基准排放,以监测其减少毁林和森林退化所致排放的进展。这些国家占全球森林面积的30%以上,占全球森林损失的70%以上。30多个国家制定了国家REDD+战略,而54个国家已将REDD+纳入其国家自主贡献。截至2020年1月,九个国家汇报因减少毁林和森林退化而致减排量达到88.2亿吨。
联合国减少毁林和森林退化所致排放计划是粮农组织、开发署和环境署的一项合作计划,支持由国家主导的REDD+进程。该计划促进了包括土著人民和其他依赖森林的社区在内的所有利益相关者的知情有意义的参与,使其参与国家和国际REDD+的实施。自2008年创建以来,该计划为非洲、亚洲及太平洋和拉丁美洲的65个伙伴国家的REDD+准备工作提供了支持。该计划已发展成为联合国关于森林与气候的知识和咨询平台,重点为推进《巴黎协定》第5条和第6条(森林作为碳储的保护、REDD+和其他可持续管理森林的方法[第5条]和碳市场[第6条]),支持政策和战略的实施,促进以制止毁林和保护恢复森林为目的的活动获得气候资金支持和建立合作伙伴关系,从而同时实现气候、生物多样性和生计的多重目标。
《纽约森林宣言》是一项于2014年启动的自愿且不具约束力的国际宣言,旨在采取行动以制止全球毁林。该宣言目前已得到200多个支持方的拥护。包括国家和地方政府、跨国公司、代表土著社区的组织和非政府组织。重要的是,它明确包括了私营部门在2020年前消除主要农产品供应链中的毁林现象的承诺,并对实现这一承诺给予支持(见插文37中的示例和图43)。
大约70%全球可可的供应来自西非小型农户,可可在这些生产地区是生产者的主要现金收入来源(Gayi和Tsowou,2016)。然而,历史上可可一直是毁林的重要驱动力和直接原因(Ruf和Zadi,1998)。因为刚清除天然植被的土壤通常更肥沃,所以已经建立的人工种植可可产量低通常是促使森林扩张的原因。
各国政府和私营部门做出了一系列承诺以终止可可供应链中的毁林现象,保护生物多样性和生态系统服务,同时避免收入受损和影响当地人们生计(Carodenuto,2019)。最近的公私合作举措,例如加纳和科特迪瓦的可可森林举措(世界可可基金会,2017)和喀麦隆的绿色可可计划(IDH,2019),旨在支持可可生产的可持续集约化和气候适应力,防止进一步的毁林,实现退化森林的恢复。它们通常与各国REDD+政策和计划一致。
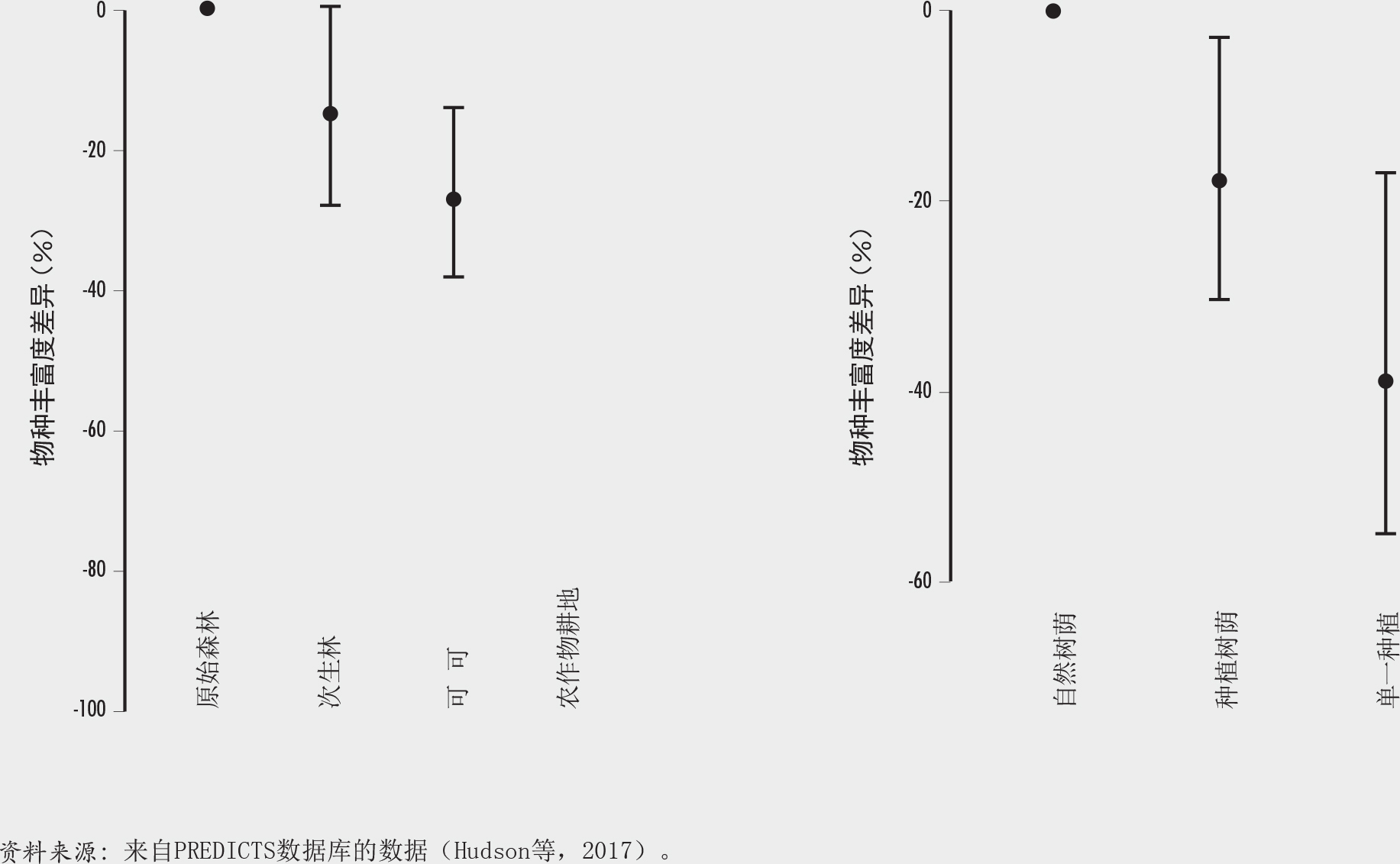
为了支持可可生产发展和可持续集约化的政策和规划,CocoaSoils研究和推广计划(Sassen、Arnel和van Soesbergen,即将出版)的一项研究认定了对生物多样性至关重要(基于世界自然保护联盟红色名录)同时也适和可可生长的森林区域(基于Schroth等[2016]的建模),因而这些地区具有较高的毁林风险(图A中的暗褐色区域)。
该研究还利用在变化陆地系统中预测生态多样性反应数据库(PREDICTS)(Hudson等,2017)中非洲、亚洲、美洲和大洋洲的数据,分析了生物多样性如何受不同可可系统土地使用状况的影响并发生相应变化。结果表明,就物种丰度和群落构成而言,建立可可生产系统的影响不如受农作物耕地影响严重,自然遮荫的混农林业系统具有的物种丰度比单一种植可可更高(图B)。随着时间的流逝,尽管在可可种植园的一个周期(约25年)内该系统未能完全恢复原有的森林群落,但可可混农林业系统变得与森林更加相似。因此,尽管可可混农林无法代替天然林,但它们仍然是保存和保护生物多样性同时又兼顾保持农业系统高水平生产力的宝贵工具(另见Schroth等,2004)。
复杂多样的结果突显了西非可可产区内不同地区面临的不同风险和机遇。在土地非常适合可可生产、与其余森林重叠且具有较高生物多样性价值(如利比里亚和喀麦隆)的地区,需要通过仔细周全的规划来保护现有保护区并限制可可生产向未保护森林的进一步发展。在这些地方,支持小型农户在多样化生产系统中进行可持续、零毁林的可可生产至关重要。
在如科特迪瓦和加纳这样已将许多原有森林转为农业生产的地方,可可混农林业系统有可能为增加农业系统中树木覆盖和恢复已退化土地(例如在REDD+框架下)发挥重要作用。这些系统有助于至少维持一些生物多样性,并为本地和全球生态系统服务以及生计多样化提供支持。
由于小型农户不太可能承受改变其一贯做法的相关费用,因此还需要一定的融资和财务机制来激励可可的可持续生产(例如信贷、环境服务支付或碳融资)。
在自给自足农业或采集薪柴是毁林主驱动力的地方,通过将可持续林产品生产和林业服务多样化来发展依赖森林的生计、发展中小企业、以及利用对碳固存或其他环境服务进行补偿这些手段,可以增加森林对当地社区的价值,促进森林保持完整。
森林合作伙伴关系在2018年2月召集了一次全球会议,邀请主要利益相关者团体讨论如何遏止毁林(插文38)。2019年7月,欧洲委员会启动了关于加强欧盟在保护和恢复世界森林方面行动的通告(欧盟委员会,2019a),列出了五个优先事项:
▸ 减少欧盟在陆地上的消费足迹,并鼓励消费欧盟零毁林供应链产出的产品。
▸ 与生产国合作,减轻对森林的压力,并进行“零毁林”的欧盟发展合作。
▸ 加强国际合作,遏止毁林和森林退化,鼓励森林恢复。
▸ 将资金导向更可持续的土地使用方式。
▸ 支持有关森林和林产品供应链信息的可用性、质量和获取,并支持研究和创新。
2018年2月,森林合作伙伴关系(成立于近20年前,一个由15个国际组织和重要涉林秘书处组成的自愿性伙伴关系,粮农组织担任主席)召集了一个全球会议,题为“跨部门努力制止毁林和增加森林面积:从愿望到行动”。300名来自政府、国际组织、学术界、私营部门、民间社会和农民组织的人士参加了会议。会议列出了以下制止和扭转毁林所需采取的措施:
▸ 作为森林监管者同时也通常是大规模森林所有者,各级政府必须带头制定适宜的条件,以确保所有森林得到可持续管理,并为此目的吸引长期融资和投资。这包括建立具有参与性、包容性和透明性的规程,以便社区和公司利益相关者参与土地使用规划和决策。
▸ 农业企业应兑现其到2020年农产品生产和加工中实现零毁林的承诺。尚未做出零毁林承诺的公司也应这样做。商品投资者应采用对环境和社会负责的商业模式,并通过如推广活动和共同计划可持续使用公司土地来鼓励当地生产者、分销商和其他价值链参与者参与并受益。
▸ 林产工业企业应确保林产品的合法性和可持续价值链,包括通过森林管理和产销监管链认证,并在此过程中与当地社区合作。
▸ 民间社会组织通过要求政府和企业承担责任来充当变革的监督者和推动者。非政府组织应通过多方利益相关者的倡议和平台来加强其话语权和影响力,这些倡议和平台促进人们对价值链各环节和各行业间所有利害相关方(包括男性和女性)的角色、贡献和兴趣的理解和认可。
▸ 公共和私人行动方应充分利用民间社会,特别是妇女和青年的潜力。青年可以促进集体行动、参与、创新、能力建设、建立联系和伙伴关系,并提供长远的视角。
资料来源:粮农组织 和CPF,2018。
尽管已经取得了一些进展(另见第2章),但仍有许多工作要做。
偷猎、非法采伐以及木材和其他森林资源非法贸易是一个全球现象,对保护生物多样性(参见第3章关于对物种生物多样性的影响)、生态系统服务和国民经济产生严重影响。同时还对城市和农村社区造成直接和间接的不利影响,主要是由于攫取森林资源动摇了这些社区人们赖以生存和获得福利的基础。
非法森林活动包括违反国家或国家以下各级法律的林产品获取、运输、加工、购买或销售。驱动森林资源非法攫取和贸易的因素很复杂,因时间的不同以及所涉商品类型和所处地点以及非法活动的不同而有很大的差异。非法活动的直接原因包括生产国森林管理薄弱以及由此导致的执法不到位;法律框架不明确;制定和实施土地使用计划上能力有限。然而,消费国由于进口林产品(包括木材、野生动植物和衍生产品)而未确保其来源的合法性,从而加剧了这些问题。例如,在撒哈拉以南非洲,野生动物非法贸易的主要驱动力包括消费国(例如东南亚)需求的增加、生产国的贫困和缺乏其他获得生计的途径、以及文化和殖民地遗留问题(Price,2017)。
除了物种和生态系统的丧失和破坏对环境产生影响之外,非法攫取森林还具有经济和社会影响。据非洲开发银行(ADB)估计,自然资源非法贸易对非洲的不利经济影响约为每年1200亿美元,相当于非洲大陆国内生产总值(GDP)的5%。在这一总额中,约有10%属于森林部门(非洲开发银行,2016)。非法贸易导致税收严重流失,对国家和地方各级都有影响。由于损失的收入无法再投资于该部门,收入损失损害了林业部门为国民生产和社会可持续发展做出贡献的能力。因为非法产品通常比合法产品更便宜,非法活动还扭曲了全球市场,破坏了促成可持续森林管理的激励机制。在社会影响方面,非法采伐和贸易往往与腐败有关,缺乏对森林社区或土著居民土地所有权和使用权的认可,这可能对当地居民的生计产生不利影响并导致冲突。
非法采伐林木。违反国家法律的木材采伐、运输、购买或销售(通常统称为“非法采伐”)是一个长期以来一直存在的全球性问题,尽管为此做了许多努力,但仍影响着温带和热带地区的许多森林国家。量化非法木材采伐具有挑战性并可能引起争议,但国际刑警组织(INTERPOL)估计包括企业犯罪和非法采伐在内的森林犯罪价值每年在510亿美元至1520亿美元之间(Nellemann等,2016)。Hoare(2015)估计,2013年全球贸易中约有50%的非法木材来自印度尼西亚,另有25%来自巴西(两国位居全球森林面积最大的十国之列)。自那时起,这两个国家均做出了巨大的努力(例如参见粮农组织,2020和第94页解决非法性问题)。其他热带木材生产国非法采伐量可能较少,但或许占各国木材总产量的比例更大。人们对木材的需求如此之大,以至于除非在全球范围内为控制非法采伐做出持续的努力,否则它仍将是森林资源未来的关注焦点(Hoare,2015)。
非法采伐可能是对木材资源需求的直接结果,包括针对最昂贵木材种类特定需求,也可能是对油棕和大豆等商品种植园进行土地清理的副产品。如上所述,毁林的最重要驱动力(合法和非法)源于对农业生产用地的需求;这种压力也最可能导致大规模非法采伐。
在大多数发展中国家,森林部门以非正规经营者为主,主要为国内市场生产的中小型企业。除了这种非正规性外,林业部门的特点是能力低下、资源有限以及资源可得性的不断变化,所有这些都使该部门容易受到非法活动的损害。
显然,由于缺乏森林管理计划,非法采伐会导致森林的丧失或退化,由此造成的栖息地和生物多样性丧失威胁到了一些物种的生存,特别是灵长类和某些大型哺乳动物。非法采伐活动经常针对和危害某些需求旺盛并可立即获得收入的珍贵木材品种。玫瑰木(Dalbergia spp.)就是一个典型例子。据估计,尽管玫瑰木被列入《濒危野生动植物种国际贸易公约》附录II,但在2009-2014年间,通往中国的红木出口增长了14倍(Bolognesi等,2015;Ong和Carver,2019)。在马达加斯加,非法采伐和走私红木导致严重的森林退化和生物多样性丧失(Ong和Carver,2019)。
木炭的非法生产比高价值木材的采伐和贸易更具挑战性,因为该部门非常零散和非正式。然而,它也造成了森林的丧失和退化。例如,Bolognesi等(2015)估计,2011-2013年间,索马里的木炭非法贸易达到24000吨,导致树木覆盖降低了2.7%。
非法攫取野生动植物。国际刑警组织估计,野生动植物非法贸易的年价值在70亿美元至230亿美元之间(Nellemann等,2016)。尽管某些类型的非法野生动植物贸易与特定地区密切相关,但世界上所有地区都在违禁野生动植物的来源、过境点或目的地这个过程中有所参与;例如,鸟类与中美洲和南美洲有关,哺乳动物与非洲和亚洲有关,爬行动物与欧洲和北美有关(联合国毒品和犯罪问题办公室,2016)。
可以说,非洲象是最著名的关键物种(即对特定生态系统构成的巨大影响与其生物量不成比例的物种)被过度攫取的案例,在上个世纪内大约损失了总数90%的(TRAFFIC,2019)。森林象对森林和其他自然生态系统特别重要,因为它们散播大种子,保持森林冠层开放,并在森林中散布稀有养分,非洲热带地区的许多物种均从中受益(Maisels等,2013)。
解决非法性问题。在过去十年中,消费国的贸易法规带动了应对非法采伐的行动,这些法规要求进口商证明木材是合法采伐的。需求方面的重要立法包括美国的《雷斯法案修正案》(2008年),《欧盟木材法规》(2013年),《日本清洁木材法案》(2016)以及《韩国木材可持续使用法案》的修正案(2017)。许多热带木材生产国正在做出相应努力,以加强遵守合法性法规和木材合法性验证。印度尼西亚尤为突出,已经实施了国家木材合法性保证体系(Sistem Verificasi Legalitas Kayu,SVLK),并于2016年首次颁发了符合欧盟木材进口要求的森林执法、治理和贸易(FLEGT)木材出口许可证(EU FLEGT Facility, 日期不详)。随着执法的加强,印度尼西亚的官方数据显示,被授予许可的企业量从2015年的25个增加到2017年的88个(MEF,2018)。其他14个热带木材生产国正在建立国家体系,实现在FLEGT机制(EU FLEGT Facility,日期不详)之下确保合法性。作为该机制的一部分,各国被要求采取措施以防止非法狩猎(见插文39)。
雨林覆盖了喀麦隆40%以上的面积,构成了刚果盆地森林生态系统的重要组成部分(粮农组织, 2020)。在农业和木材采伐的驱动之下,这一生物多样性极度丰富的生态系统受到了毁林和森林退化的威胁(MINEPDED,2013)。据估计,该国815种开花植物受到威胁(Onana、Cheek和Pollard, 2011),26种哺乳动物目前被列为濒危或极度濒危(IUCN,2019a)。
作为其为解决非法采伐、非法偷猎及野生动植物贸易问题而做出努力的一部分,喀麦隆于2010年与欧盟就森林执法、治理和木材及衍生产品贸易问题签署了《自愿伙伴关系协议》(欧盟, 2011)。该协议的一个基本要素是合法性验证系统,该系统基于一系列用于验证木材合法来源的标准和指标。该系统的标准5规定,所有允许采伐木材的区域(例如森林特许权区域、社区森林、委员会森林)必须遵守与生物多样性保护相关的国家法规(MINEF,1998;MINEF,2001),并采取措施防止非法狩猎野生动物。
为促进标准5的实施,喀麦隆野生动物保护协会(WCS)在粮农组织-欧盟FLEGT计划的资金支持下,开发了一套综合工具,可协助森林管理和经营者遵守有关监测和评估野生动植物管理的规定。其中包括SEGeF(Suivi de la gestion de la faune dans les forêts de production)监测和评估指标体系,该体系已被整合到网络和移动终端应用程序中(SEGeF,2018)。2019年,喀麦隆政府签署了一项立法,规定在喀麦隆的生产林中必须使用该体系(MINFOF,2019)。喀麦隆野生动物保护协会与森林经营者和森林社区密切合作开发和实施了该工具,并提供了使用方面的培训。
2015年7月,联合国大会通过了有史以来的第一个《关于解决野生动植物非法贩运问题的决议》(69/314)(联合国大会,2015b),该决议还涉及木材走私问题。该决议第四版于2019年9月通过(联合国, 2019b),呼吁加强国家立法,支持可持续生计,改善政策执行和反腐败措施,协助部署信息技术,并促进针对减少需求的工作。
可持续野生动物管理合作伙伴关系(CPW)(粮农组织,2019f)提供了一个平台,用于解决需要国家和跨越国家做出反应的野生动物管理问题,包括非法野生动物贸易。成立于2013年的可持续野生动物管理合作伙伴关系是14个在野生动物方面有重要工作的国际组织的自愿合作伙伴关系,旨在促进野生动物资源的可持续利用和保护。
《2019可持续发展目标报告》(UN,2019a)表明,在2000-2015年间地球表面的20%处于退化状态(图33)。2019年3月1日,联合国大会宣布2021-2030年为联合国生态系统恢复十年,旨在预防、制止和逆转生态系统退化,提高对生态系统恢复重要性的认识,并加快实现全球(插文40)和区域生态系统恢复目标的进程。
▸ 可持续发展目标15.3:到2030年,防治荒漠化,恢复退化的土地和土壤,包括受荒漠化、干旱和洪涝影响的土地,努力建立一个无土地退化的世界。
— SDG 15.3.1:已退化土地占总土地面积的比例。
▸ 爱知生物多样性目标15:到2020年,通过养护和恢复行动,生态系统的复原力以及生物多样性对碳储存的贡献得到加强,包括恢复了至少15%退化的生态系统,从而对气候变化的减缓与适应以及防治荒漠化做出了贡献。
▸ 《波恩挑战/纽约森林宣言》目标5:到2020年恢复1.5亿公顷退化景观和林地,此后显著提高全球恢复速度,到2030年至少恢复另外2亿公顷。
▸ 联合国森林战略计划目标1:通过森林可持续管理,包括保护和恢复森林、造林和再造林,扭转全球森林覆盖下降的趋势,并加大努力防止森林退化,应对气候变化。
—具体目标1.3:到2020年,推动所有类型森林的可持续管理,停止毁林,恢复退化森林,在全球大力开展造林和再造林。
恢复是《生物多样性公约》的生物多样性战略计划和爱知目标(《生物多样性公约》,2010a)的关键部分。森林景观恢复已被认为是实现爱知指标5、7、11、13和15的一种措施(Dave等,2019)。
迄今为止,《联合国防治荒漠化公约》的土地零退化目标设定计划已经收到122个国家土地零退化的承诺(UNCCD,2019a)。区域土地恢复目标包括拉丁美洲20x20倡议(20x20倡议,日期不详),计划到2020年恢复2000万公顷的退化土地;AFR100(非洲森林景观恢复计划),其目标是到2030年使1亿公顷的退化土地得到恢复(AFR100,日期不详);《地中海阿加迪尔承诺》,计划到2030年恢复至少800万公顷退化的森林生态系统(粮农组织,2017d);ECCA30计划,一项欧洲、高加索和中亚国家的项目,计划到2030年恢复3000万公顷的退化土地;撒哈拉沙漠和萨赫勒地区的绿色长城倡议,计划到2030年恢复1亿公顷土地(绿色长城,2019a)。
森林恢复可以具有与扭转土地退化或提供商品和服务(如粮食、生物多样性和水)的生态系统生产力丧失有关的多种目标,它们包括:
▸ 恢复:将所需物种、结构或过程恢复到现有生态系统;
▸ 重建:在用于其他目的的土地上恢复本地植物;
▸ 开垦:恢复没有植被的严重退化土地;
▸ 替代:最激进的一种恢复形式。由于气候变化迅速,将不能适应某处环境且无法迁移的物种或种源用新植被替代(Stanturf、Palik和Dumroese,2014)。
在实施恰当的情况下,森林恢复有助于恢复栖息地和生态系统、创造就业机会和收入,是一项以基于自然的应对气候变化的有效解决方案(CBD,2016a;粮农组织与UNCCD全球机制,2015;IPBES,2019a)。见案例研究1。
“防治荒漠化行动”(AAD)是一项由粮农组织及其合作伙伴执行,由欧洲委员会和非洲、加勒比及太平洋国家集团秘书处资助的行动,为撒哈拉和萨赫勒倡议的绿色长城计划提供实地支持。其目标是通过大规模恢复退化土地来增强受气候多变性和气候变化影响严重的旱地社区和农林牧生态系统的应变能力,从而减少贫困,实现粮食、饲料和营养安全,增强复原力。该行动通过提供多种环境和社会经济效益,为实现《2030年可持续发展议程》做出贡献。
防治荒漠化行动的旱地大规模恢复蓝图强调基于植物的解决方案,其中包括:
▸ 通过机械耕作和补植投资大规模整地;
▸ 通过生物物理和生物干预措施来防止沙土侵蚀,实现固土的目的;
▸ 在土壤种子库和残余植物允许的任何地方促进自然再生;
▸ 从丰富的旱地植物生物多样性中调取高质量的种子和种质;
▸ 在农村地区发展非木质林产品价值链,使妇女、男子和青年受益;
▸ 支付得起的、有参与性的信息传播系统;
▸ 具创新性的用以评估进展情况的生物物理和社会经济监测系统。
五年来,防治荒漠化行动已恢复了53000公顷退化的农林牧用地,种植了2500万棵农村社区广为使用的本地树种。在九个国家收集并种植了包含110种木本和草类饲料的100吨种子,带来了巨大的经济和环境效益。例如,布基纳法索和尼日尔种植的草场仅在种植后一年就达到每公顷1200公斤生物质的平均产出,每公顷收入40美元,相当于该国人均每月最低工资的一半;因此,布基纳法索1万多公顷恢复的土地每年可为当地农民带来40万美元的收益。在塞内加尔,在干旱季节(11月至5月)从约4000公顷恢复的退化土地上收获草料的村民可以从每个驴车获得2美元或每车(约100公斤饲料)获得4美元。按每亩产1吨来估计,这样的操作在2017-2019年间平均每年向当地经济贡献80000美元。此外,根据种植后3年至20年的结果推算,萨赫勒地区用本地树种恢复的土地每年每公顷可以固存7.15吨二氧化碳当量。
防治荒漠化行动为了增加适应能力而进行的土地恢复方法,将社区和植物知识置于干预措施的核心位置。促成防治荒漠化行动行动成功的因素包括:
▸ 动员社会和赢得当地社区对社区土地干预的支持;
▸ 利用植物知识和专门知识来优先考虑对社区有用的适应性良好的植物物种,确保获得支持;
▸ 将经过良好测试的方法和传统知识有机结合起来,克服技术和研究上面临的挑战,例如在正确的位置和正确的时间识别和种植正确的物种,实现从雨水中获得最大的收益,在不利环境下最大程度地保证植物的存活和生长。
这种方法非常适应各种生态和社会经济条件,因此,在持续投资允许的情况下,非常适合在非洲及其他地区进行复制和大规模推广使用。防治荒漠化行动最近开始将其干预措施扩展到南部非洲。在那里,南部非洲发展共同体(SADC) 各国在该共同体的协调以及非洲联盟委员会的支持下启动了绿色长城行动。
资料来源: 粮农组织,2019g。
森林与景观恢复全球伙伴关系(GPFLR,日期不详)制定了六项全球公认的森林与景观恢复准则:
▸ 专注于景观系统。
▸ 鼓励利益相关者介入,支持参与式治理。
▸ 还原多种森林功能以获得多种益处。
▸ 维护和增强景观内的自然生态系统。
▸ 根据当地情况量身定制恢复方法。
▸ 适应性管理以获得长期复原力。
现在有许多森林恢复指南,包括从业者森林景观恢复指南(Stanturf、Mansourian和Kleine,2017),专门针对退化旱地森林的(粮农组织,2015b)、针对红树林的(Field,1996)、针对自然再生在森林和景观恢复中的作用的(Chazdon等,2017),以及将对生物多样性的考量整合到生态系统恢复之中的(《生物多样性公约》,2016a)各种具体指南。国际热带木材组织正在更新其关于恢复、管理和养护热带退化林和次生林的准则(国际热带木材组织,2002)(另见插文41)。
森林的自然再生是一个可以加以辅助和管理的生物过程,以达到增加森林覆盖并实现复原本地生态系统或其中某些功能的目的。辅助自然再生(ANR)是指旨在增强和加速天然林自然再生的任何干预措施,例如提供保护使其免受外部干扰(火灾、觅食的家畜,和人类),减少来自阻碍自然再生树木生长的草、灌木和藤类植物的竞争。
辅助自然再生是一种通过消除或减少自然障碍来恢复森林的简单、成本低且有效的技术。除了增强抵御能力并提供多种林产品和生态服务之外,辅助自然再生在恢复生物多样性、物种间互动和在不同生境内迁移等方面非常有效。在辅助自然再生的过程中,通过以下方式丰富了本地生物多样性:
▸ 从种子、根萌苗、树桩或萌芽自然生成的树木和灌木;
▸ 适应当地土壤和气候条件的当地遗传资源的再生;
▸ 树木的相关授粉媒介、草食动物和种子传播媒介。
使用直接播种和植树方法也可以实现许多这些好处,但成本要高得多。在热带地区,自发和辅助的自然再生比植树能更有效地实现生物多样性和森林结构的恢复,并且比起一般的植树只种植有限的树种来说,其生成的多层植被也更为多样化。
资料来源:粮农组织,2019g。
恢复森林生态系统远远超越植树或辅助树木的自然再生。见案例研究1和插文42中重新野化的例子。
野化旨在恢复生态的自然演替,达到自我维持的生态系统和生态系统过程,并强调基于过程的保护方法(《生物多样性公约》,2014)。
在某些情况下,这是以被动方式实施的,即顺其自然。在另外一些情况下,它包括重新引入高阶捕食者和关键物种。一个著名的例子是将狼重新引入美国的黄石公园。
狼(Canis lupus)曾经在北美地区从北极到墨西哥自由漫游,但是截至1926年,作为当时消灭所有捕食者的政策的一部分,最后一个狼群在美国最古老的国家公园黄石被消灭。
在短短几年之内,作为鹿种群最大的物种之一,红鹿(Cervus elaphus)的数量急剧增加,过度地啃食了柳树(Salix spp.)和美州山杨(Populus tremuloides)。没有这些树木,鸣鸟开始衰落,海狸(Castor canadensis)无法再建坝,河岸就开始出现侵蚀现象。海狸和河畔木本植被的丧失导致河流和河岸生态系统功能的严重退化。河道变得更宽,更浅,温度也有所升高。总而言之,这些河流的变化严重破坏了鱼类的栖息地。
1995年,与加拿大机构合作,14头在贾斯珀国家公园捕获的狼被引入黄石公园,于1996年又增加了17头狼。红鹿和鹿种群立即有反应。在大约十年之内,柳树在许多地区逐渐恢复,尽管美州山杨仍旧受到鹿和野牛吃食的影响,但最近在一些林分上它们也正在恢复。鸣鸟、海狸、鹰、狐狸和獾也返回了公园。
尽管在黄石公园重新引入狼群耗费约3000万美元,但是狼生态旅游每年可带来3500万美元的收入,使周边社区受益。
资料来源:Monbiot,2013;Boyce,2018;Kay,2018;《卫报》,2020。
森林恢复的主要挑战来自于如何使从业者和政策制定者共同努力,确保恢复活动计划得当、实施成本效益高、并在众多发展目标中得到足够的重视和优先考虑(Sabogal、Besacier和McGuire,2015;粮农组织和《荒漠化公约》全球机制,2015;Strassburg等,2019)。许多有公私部门共同参与的双边和多边项目计划都在努力处理好这一挑战。第二个挑战是让生产者组织、农民和中小型企业参与恢复,并确定和启用可使人们通过可持续的土地使用过上体面生活的商业模式。为了给发展商业模式提供基础,一项新计划旨在提供有关生态系统恢复的成本效益的信息。见下面插文43。
始于2019年的“生态系统恢复经济学”倡议旨在为现有和未来全球主要生物群落中和不同背景下的生态系统恢复项目的成本和收益估算提供参考,主要通过一个标准化的框架,从具可比性的项目收集信息。
该倡议由粮农组织牵头,并与多个组织合作,包括《生物多样性公约》和《荒漠化公约》秘书处,国际生物多样性委员会,国际林业研究中心,自然保护联盟,Tropenbos国际,WeForest和世界资源研究所(WRI),共同建立了一个信息平台,开发了决策工具,供捐助者、投资者、项目实施者、政府和其他利益相关者参考使用,获取可靠的成本和收益数据,进行生态系统恢复决策。
这项倡议将于2020年发布的第一项产出是一个框架,用于收集有关生态系统恢复成本和收益的一致而可靠的数据,有助于进一步的分析和决策。正在萨赫勒地区进行一项试点研究,数据收集将很快扩展到所有主要生物群落。
最近研究表明,大约有17亿至18亿公顷的潜在林地(定义为可以支持树木覆盖率达到10%以上的土地)存在于以前退化和以稀疏植被、草地和退化的裸露土壤为主的地区(Bastin等,2019);这些地不包括现有的森林以及农业和城市用地,相当于9亿公顷连续森林覆盖,占全球目前森林面积的25%以上。然而,需要谨记的是,该研究仅关注土地支持森林生成的生物物理潜力,而未将当前生态系统和现有土地所有权这些重要因素考虑在内。因此,需要结合了当地状况的更为详尽的评估,来确定国家或地方层面最合适进行森林恢复的区域。
粮农组织已开发出用于土地监测的地球观测数据获取、处理和分析系统(SEPAL)中的一个模块,该模块结合了计算树木恢复潜力的公式,有助于各国确定适合恢复的区域。粮农组织及相关各国政府机构将于2020-2021年在柬埔寨、肯尼亚、缅甸和乌干达试用该模块。
作为对世界自然保护联盟(IUCN)开发的恢复机会评估方法的补充,一些现有的特定指南可以将生物多样性纳入景观恢复机会评估中(Beatty、Cox和Kuzee,2018)。
对亚洲、非洲和拉丁美洲的62个国家进行的一项研究发现,每个区域都有超过半数的国家在其国家生物多样性战略和行动计划或向《生物多样性公约》提交的第五次国家报告中有既定或初步的恢复目标(《生物多样性公约》,2016b)。虽然建立目标是很好的第一步,但履行承诺仍然具有挑战性(图34)。此外,恢复工作难以衡量,目前还没有全球数据来衡量森林景观恢复的进展(NYDF,2019)。粮农组织正在与几个伙伴组织机构合作,为联合国生态系统恢复十年建立一个全球监测系统,同时粮农组织和世界资源研究所(2019)已制定了指南,帮助各国和从事恢复工作的人员确定监测森林和景观恢复的优先事项和指标。
许多目标缺乏定量要素,而开展恢复活动是一个复杂的过程。但是,还是有一些实现恢复的成功范例(图35)。例如,得益于政府主导的森林政策或举措,森林覆盖在中国、哥斯达黎加、大韩民国和越南都得到显著的增加。在尼日尔南部,农民管理的混农林业下的天然再生法在过去的三十年中使500万公顷土地的生产力得到提高(Reij、Tappan和Smale, 2009)。另一个例子是非洲联盟于2007年发起的“撒哈拉-萨赫勒绿色长城”倡议,该倡议旨在2030年前恢复目前退化的1亿公顷土地,固存2.5亿吨碳,创造1000万个绿色就业机会,在非洲旱地上修建8000公里的绿墙(参阅案例研究1)。自2007年以来的进展(绿色长城, 2019b;《荒漠化公约》,2019b)包括:
▸ 300万公顷布基纳法索土地通过采用当地做法得以修复;
▸ 1500万公顷埃塞俄比亚退化土地得以恢复,林权状况得到改善;
▸ 500万公顷尼日利亚退化土地得以恢复,在11个州建立了639公里防护林带,建立了309公顷社区果园和293公顷社区林地;
▸ 500万公顷尼日尔土地得以恢复;
▸ 在不到十年的时间里,塞内加尔种植了1200万棵抗旱树木。
截至2019年10月,61个国家在“波恩挑战”下作出在2020或2030年前恢复总计1.706亿公顷土地的承诺(图36)(Dave等,2019)。然而,自2000年以来,就森林或树木的覆盖而言,仅实现了2020年目标的18%(到2020年恢复1.5亿公顷的退化景观和林地)(NYDF,2019)。《波恩挑战晴雨表》(IUCN,2018;Dave等,2019)致力于获取关于实质性实施进展更精确的信息,包括恢复土地公顷数、提供的相关生态系统惠益(包括碳固存和生物多样性保护)和创造的就业机会(Dave等,2019)。
许多国家在2019年9月于美国纽约举行的气候行动峰会上宣布了恢复森林和树木的新承诺(插文44)。2020年初,世界经济论坛启动了一项栽种、恢复和养护1万亿棵树的倡议(世界经济论坛,2020)。
▸ 巴巴多斯:到2020年将种植100万棵树
▸ 哥伦比亚:到2022年将恢复30万公顷(1.8亿棵树),并在混农林业和可持续森林管理下恢复90万公顷
▸ 刚果民主共和国:森林覆盖率稳定在60%
▸ 欧洲、高加索和中亚:到2030年,将3000万公顷退化和毁林土地恢复原状
▸ 埃塞俄比亚:一年将种植40亿棵新树
▸ 斐济:将种植100万棵新树,并努力争取再种植3100万棵树
▸ 危地马拉:到2022年将恢复150万公顷土地
▸ 匈牙利:到2030年森林覆盖将增加30%
▸ 肯尼亚:到2022年将植树20亿棵
▸ 马里:到2030年将恢复1000万公顷*
▸ 新西兰:到2028年将植树10亿棵
▸ 尼日利亚:青年人将植树2500万棵
▸ 巴基斯坦:未来五年将植树100亿棵
▸ 塞内加尔:到2030年将恢复200万公顷的土地*
▸ 塞拉利昂:到2023年将植树200万棵
资料来源:除带星号(*)外,均来自Nature4Climate,2019。带星号信息来自提交给AFR100的国家报告。
本章探讨如何以保护和可持续利用森林生物多样性的方式来管理世界森林生态系统。
从历史上讲,建立保护区一直是实现生物多样性目标最常采用的森林治理措施(Watson等,2014)。许多森林保护区的管理都设法协调当地人们的生计与生物多样性的保护。尽管现有证据还无法对该措施在保护最稀有物种方面的作用做出定论,但保护区方法在遏止毁林和保护物种方面已取得了积极成果。
然而,从生物物理学的角度来看,有证据表明仅有自然保护区是不足以保护生物多样性的。保护区通常太小,而且形成了物种迁移的障碍,也很容易受到如气候变化等外来因素的影响(Bennett, 2004;Fung等,2017)。此外,保护区仅占现有森林生物多样性的一小部分。因此,需要着眼于保护区之外的方法,将保护生物多样性纳入主流森林经营措施。
一些兼顾资源保护和社会经济发展目标、支持可持续资源利用并下放森林管理权于民的方法逐渐涌现出来。这些异于严格保护的措施,可以成为与之互补的方法(Agrawal、Chhatre和Hardin,2008;Lele等,2010;Mace, 2014)。许多基于利益相关者的治理方法,就自然资源的多种用途有时甚至是相互矛盾的用途进行协商,以保护当地人使用和珍惜的资源,以及那些在更广泛范围内支持和满足社会需求的资源(Kaimowitz & Sheil,2007;McShane等,2011)。例子包括土著社区、民间组织和私营部门管理和保护的区域(Stolton等,2014;Drescher和Brenner, 2018),日益强调综合景观和以权利为基础的方法。在许多情况下,协调森林利用和森林保护意味着协调当地和全球的需求。
认识到保护区之外地区的保护(包括生产林)的重要性体现在将其他有效的基于地区的保护措施(即那些在保护区之外的被保护的区域)包括在内,并在全球保护目标中明确提出这些区域的可持续利用(插文45)。
▸ 可持续发展目标15.1:到2020年,根据国际协定的义务,保护、恢复和可持续利用陆地和内陆的淡水生态系统及其服务,特别是森林、湿地、山麓和旱地。
— 可持续发展目标指标15.1.2:保护区内陆地和淡水生物多样性的重要场地所占比例,按生态系统类型分列。
▸ 爱知生物多样性目标11:到2020年,至少有17%的陆地和内陆水域以及10% 的沿海和海洋区域,尤其是对于生物多样性和生态系统服务具有特殊重要性的区域,通过有效而公平管理的、生态上有代表性和相连性好的保护区系统和其他基于保护区的有效保护措施的得到保护,并被纳入更广泛的土地景观和海洋景观。
▸ 联合国森林战略计划目标3:大幅增加世界各地森林保护区及其他可持续管理林区的面积,提高可持续管理林区林产品所占比例。
— 具体目标3.1:大幅增加全球森林保护地面积或其他通过有效地域保护措施保护的森林面积。
在过去几十年中,全球保护区规模迅速扩大,达到约240000个指定的保护区,其中大部分是在陆地上。总的来说,这些地区保护了超过20亿公顷土地,相当于地球陆地表面的15%(UNEP-WCMC、IUCN和NGS,2020)。成千上万个保护区是专门为保护森林而设计的;其中一些是世界上最古老的保护区之一。例如,斯里兰卡的马拉凯尔森林自然保护区自1875年起就一直用于保护森林。
保护区可以根据其管理目标进行分类(插文46)。
第Ia类涵盖了为保护生物多样性以及可能的地质/地貌特征而保留的严格保护区,在这些区域中,严格控制和限制人类的访问、使用和影响,以确保保护价值。这些保护区可以作为科学研究和监测必不可少的参考区。
第Ib类保护区通常是未经改变或稍加改变的大块区域,保留其自然特征和影响力,没有永久或重要的人类居住地,对其进行保护和管理以保持其自然状况。
第II类保护区是为保护大规模生态过程而留出的大块自然区或近自然区,以及该地区特有的生态系统特征和物种,这也为在环境和文化上兼容的精神、科学、教育、娱乐和参访机会提供了基础。
第III类是为了保护特定的自然古迹而设立的保护区,这些自然古迹可以是地貌、海底山、海底洞穴、象洞穴一样具备特殊地质特征的区域、甚至可以是如古树林这样的生态园。它们一般很小,通常具有很高的旅游价值。
第IV类保护区旨在保护特定的物种或栖息地,因而管理也体现了这一优先事项。许多第IV类保护区将需要进行定期、积极的干预措施,以解决特定物种的需求或维持栖息地,当然这也不是该类别必须的。
第V类保护区是指随着时间的推移,通过人与自然的相互作用产生了具有显著生态、生物、文化和风景价值的独特区域,并且维护这种相互作用的完整性对于保护和维持该区域及其相关的自然保护和其他价值至关重要。
第VI类保护区保护生态系统和栖息地,以及相关的文化价值和传统的自然资源管理系统。它们通常很大,大部分地区处于自然状态,其中一部分处于可持续自然资源管理之下,兼顾自然保护与较低程度的对自然资源的非工业使用,被视为该区域的主要目标之一。
资料来源:UNEP-WCMC、IUCN和NGS(2020)。
从全球来看,世界森林面积的18%(超过700万公顷)处于如国家公园、保护区和野生动物保护区等(第I到第IV类保护区)依法设立的保护区范围内。南美洲保护区森林占的比重最大(31%),而欧洲则最低(5%)(图37)(粮农组织,2020)。
根据《2020年全球森林资源评估》,自1990年以来第I到第IV类保护区范围内森林面积增加了至少1.91亿公顷,但年均增长速度在过去十年已逐渐放缓(图38)。就2020年森林资源评估而言,只有129个国家报告了整个时间序列数据,共计森林总面积的84%(粮农组织,2020),因此保护区森林面积的实际增长可能会略高。
世界养护监测中心为本报告专门进行了一些新研究,考察按森林类型和全球生态区划分保护区的趋势以及关键生物多样性地区内(KBAs,即对全球生物多样性有重要贡献的点)森林面积的趋势。这些研究基于四个地理信息数据库:
▸ 保护区:2019年6月发布的世界保护区数据库(WDPA)(UNEP-WCMC和IUCN, 2019)。
▸ 关键生物多样性地区:2019年3月发布的世界关键生物多样性地区数据库(BirdLife International,2019)。
▸ 土地覆盖:来自欧洲航天局气候变化计划(ESA CCI)土地覆盖数据产品(Bontemps等,2013)(2.0.7版)的1992-2015年间年度土地覆盖数据(分辨率约300米)。
▸ 生态区:全球生态区数据库,第二版(粮农组织,2012a)。
虽然数据上不可能排除木本农作物土地覆盖,但由于这类作物很少处于保护区内,所以现有数据不太可能显著地扭转以下所列结果。
需注意的是,尽管粮农组织要求各国报告2020年第I至第IV类保护区内森林的面积,但本研究还包括第V和VI类。因此,以下所列保护区中的森林总面积比《2020年森林资源评估》报告的要显著地大。
按森林类型划分的保护区现状和趋势。在1992-2015年间,全球保护区的树木覆盖面积骄人地增加了3.96亿公顷,平均每年增加1700万公顷(图39),截至2015年总计达到8.33亿公顷(表5)。不能确定这些增加是由于保护区的普遍广泛扩展与森林碰巧重合,还是表明了对森林生态系统有针对性的保护。
受保护面积增加最大的是常绿阔叶(热带)森林(图39),它们自1992年起增加了2.26亿公顷,到2015年达到3.97亿公顷,是所有森林类型中面积最大的,也是保护区森林占比第二大的森林类型(表5)。自1992年以来,每年被保护的常绿阔叶林的增长占全球被保护森林增长的一半以上。在这23年间,所有其他森林类型都经历了明显较小的增幅(图39)。
按全球生态区划分的被保护森林的现状和趋势。在全球范围内,20个陆地生态区包含一定树木覆盖。所有生态区在2015年的被保护树木所占比例都比其在1992年时更大(图40)。在其中的三个全球生态区中(热带雨林、亚热带干旱林和温带海洋林),30%以上的树木覆盖现在已在受法律保护的地区。在另外三个全球生态区(亚热带湿润森林、温带草原和北方针叶林)中,不到10%的树木覆盖在保护区中(表6)。保护区森林比例如此低的生态区大多位于较高的纬度(图41)。鉴于保护有代表性的陆地生态系统是爱知目标11的关键组成部分,因此应将这些生态区视为进一步保护的重点。
有趣的是,尽管热带雨林森林覆盖损失率最高,但这一生态区保护区内树木覆盖增幅最大。这也许主要归因于巴西的保护区,其目前拥有世界上最大的保护区系统(UNEP-WCMC和IUCN,2019)。
截至2015年,在欧洲、智利和大洋洲部分地区的温带海洋森林在保护区所占比例最大。这部分归因于欧洲广泛存在的保护区,其几乎占有世界一半的保护区(UNEP-WCMC、IUCN和NGS,2020)。
关键生物多样性地区的森林趋势。关键生物多样性地区是明确满足11个生物多样性标准至少一项的区域,例如占某一全球濒危或极危生态系统类型的5%以上(IUCN,2016)。目前,世界上有超过15000个关键生物多样性地区,总面积超过19亿公顷(Birdlife International,2019)。其中约有95%是陆生的,而超过75%蕴含一定森林覆盖。
世界保护监测中心的研究表明,1992-2015年间这些关键生物多样性地区的森林覆盖略有下降,这一结果与其他关于某些关键生物多样性地区的研究发现是一致的(Tracewski等,2016)。尽管与保护区完全或部分重叠的关键生物多样性地区或位于较偏远地区的关键生物多样性地区与其他关键生物多样性地区相比,土地覆盖变化的可能性较低,但关键生物多样性地区本身并不提供任何正式的森林保护。虽然关键生物多样性地区森林覆盖略有减少,但是这些地区保护区的面积随着时间的推移一直稳步增长,尽管在不同国家保护水平存在显著的不同(Ritchie等,2018)。
越来越多旨在保护生物多样性的保护区采用了被称之为生物廊道或生态网络的方法(见Bennett和Mulongoy, 2006),该方法协调了生物物理和人类的视角,为更广义意义上农业生态景观的完整性做出了贡献。案例研究2展示了来自哥伦比亚的例子,该国是世界生物多样性最丰富的国家之一。过去30多年实施生态廊道中吸取的经验教训证明了廊道对保护森林覆盖有益处,然而不一定对保护所有种类的树种均有利(Bennett和Mulongoy,2006)。
自2016年以来,为期五年的加勒比生态连通性倡议(Conexión BioCaribe)一直致力于减少哥伦比亚北部加勒比海地区宝贵生态系统的退化和破碎。尽管该地区始自殖民时期前的资源开发曾经促进了经济增长,但不可持续的做法越来越多地威胁到该地区丰富的生物多样性以及农村社区和粮食安全的复原能力(粮农组织,2019i)。
该计划的核心是设计150万公顷的连通廊道,以连接彼此孤立的保护区(图A和B)。这些廊道由环保型的生产系统组成,包括林牧系统、农林系统、混合果园、水源和海岸恢复、红树林恢复、湿地恢复与水产农业,混合了既支持生物多样性保护又支持粮食生产的物种。该过程包括土地规划、具有跨文化视野的社会参与、对现有保护区的有效管理、创建新的保护区和建立连接保护区的缓冲带,以及对潜在的保护性奖励措施和认证计划进行可行性分析。
该计划取得的成果(粮农组织,2019i)包括如下对生态系统连通性以及对恢复养护鸟类和哺乳动物所做贡献:
▸ 约1.35万公顷的新保护区,另外11.6万公顷保护区的正在准备中;
▸ 在可持续生产的不同模式下耕种了约5000公顷的土地,有1500多个家庭参加了农民田间学校培训;
▸ 建成了1300公顷在可持续生产计划之下的保护区缓冲区;
▸ 建立了6.8万公顷保护和可持续利用自然资源斑块。
廊道是通过一个有当地社区和机构参与的过程进行设计的。这样可以设计出适合该民族社区的价值观和社会文化传统的活动。最后,两个土著社区和三个非裔社区将连通性方法纳入了他们的集体土地使用计划。
该倡议还促进了创建一个旨在信息传播和提高对社区活动认识的综合交流网络,该网络让儿童和青年人充分参与如何应对各个社区面临的挑战。2020年,哥伦比亚国家公园系统预计将接手负责管理这个网络,维护这些群体之间交流对话的文化主权。
近40%受保护的生态完整的生态系统(如亚寒带和热带原始森林、草原和沼泽)目前由土著居民管理(Garnett等,2018)。人们越来越认识到,与生物多样性保护地相关的当地社区的需求、知识和价值观可以为保护生物多样性做出贡献(Pretty和Smith,2004;Sayer等,2017)。这一认识为实现在保护自然遗产的同时改善生计这一双赢战略铺平了道路。保护区内人与生态系统之间的互动是否可持续以及保护水平是否足够是问题的关键,因为通常很难监测保护的有效性(Andam等,2008;Leverington等,2010)。在许多情况下,允许在保护区内开展支持当地人生计的活动,例如可持续采伐木材、采集非木质林产品(案例研究3和插文47)和可持续旅游业(案例研究4),为当地人保护资源提供了有效的激励机制。
玛雅生物圈保护区创建于1990年,旨在保护中美洲最大的热带森林地区。占地约210万公顷,其中767000公顷受到严格保护,848400公顷在多用途管理之下(包括特许经营),497500公顷在私人持有的缓冲区内。在具有明确保护目标的多用途地区,已授予约533000公顷的特许权(见图A)。
1994-2002年间,保护区授予了14项特许权,包括规模从2公顷到约130000公顷的工业用木材特许权。遵照1996年《和平协议》,十二个特许权被授予了社区,该协议明确规定政府到1999年将向中小型农民授予100000公顷的特许权。其余两个特许权被授予给私有木材公司。此后,由于耕种压力大、经济效益低以及毒品走私的存在,取消了两项社区特许权,并暂停了一项特许权。特许权目前覆盖485122公顷(Gretzinger,2016)。
维持任何特许权都必须获得森林管理委员会(FSC)的认证。它起着问责机制的作用,并作为公共机构有限的监督能力的补充。
社区特许权森林的综合管理用于实现多种用途,包括收集非木质林产品和旅游业。然而,大部分收入来自木材,特别是高价值树种,如大叶桃花心木(Swietenia macrophylla)(Rodas和Stoian,2015)。这些利润的约三分之一通过消防巡逻和森林保护的形式被重新投资到森林中。
总体而言,据报告社区特许经营的采伐强度较低。2012-2016年间,每公顷桃花心木的采伐量为0.7米3(每公顷0.29棵树),所有树木总计1.6米3(Rodas和Stoian,2015)。采伐的木材种类从4到19种不等,工业特许权采伐的木材种类通常多于社区特许权。
在特许权区保护生物多样性的成果包括木材可持续采伐(Grogan等,2016)、在厄尔尼诺和拉尼娜年份成功控制森林火灾以及降低森林火灾的发生率(CONAP和WCS 2018)、维持美洲豹种群(Polisar等,2016)以及近乎于零毁林,这为2016-2017年森林覆盖增加0.1%做出了贡献(CONAP和WCS,2018)。相比之下,核心保护区(未包括在特许权中)的毁林率差异性大一些,平均约为1%(Hodgdon等,2015)。
与发展有关的成果包括:由于特许经营者的信誉提高,木材收入增加,向外移民减少,就业机会增加,社会投资增多,能力建设改善,获得银行信贷的机会增加:
▸ 2012-2016年间,社区特许权的木材销售收入约为2500万美元。由于产品多样化(木材和非木质林产品)和添加附加值能力增强,参与家庭的森林收入是贫困线的1.6到2.8倍(Stoian和Rodas,2018)。
▸ 森林收入(约占家庭收入的38%)加上特许权提供的社会服务(如奖学金和医疗保健)有助于减少向外移民。平均而言,特许经营区的汇款仅占家庭收入的2%(Stoian等,2018)。
▸ 非木质林产品的生产和商业化中的就业机会对妇女尤为重要,林产品如枣椰树叶(Chamaedorea spp.)、面包树(ramsimum alicastrum)的玛雅拉蒙坚果、蜂蜜和甜椒。
▸ 特许经营权将获得利润投资于社区项目,例如基础设施(道路建设和维护)、卫生服务和教育(奖学金、教师薪酬)。调查显示,与现金相比,社区成员更喜欢实物分配和将森林收入进行再投资(Bocci等,2018;Stoian等,2018)。
▸ 特许经营管理和认证要求为增强社区企业的技术和管理能力提供了机会和动力。
▸ 社区可以从接受年度运营计划作为抵押的银行获得融资。许多社区通过预付款(包含利息)来为采伐活动提供资金。
山地合作伙伴产品(MPP)倡议是一项认证和贴标计划,旨在通过改善山地产品(如有机食品、纺织品和旅游服务)的价值链,向山地小农生产者提供技术和财务支持,帮助其创建企业,增强营销技能,改善生计。该倡议促进较短的国内价值链,同时也确保生产者和消费者之间的透明度和信任,对初级生产者提供公平补偿,以及保护农业生物多样性和古老技术。每件产品都有一个描述性标签,说明产品的起源和栽培、加工和/或保存方法、营养价值(对于食品而言)以及在当地文化中的作用,使消费者能够在知情的情况下进行购买。迄今为止,该倡议已支持了约1万名农民,其中6000名是妇女。
山地合作伙伴产品计划支持的产品之一是无刺蜜蜂(Tetragonisca angustula)的蜂蜜,这是一种土著林产品,由多民族玻利维亚国查科省依尼奥国家公园(Serranía del Iñao)的160名瓜拉尼社区妇女组成的合作社精心采集。瓜拉尼社区人们自古以来就养殖蜜蜂。然而,毁林和高产欧洲蜜蜂的引进导致350种已知无刺蜂(麦蜂族)分布的减少,蜂蜜已成为一种稀有商品。无刺蜂是完全适应当地环境的重要授粉者。它们的丧失可能导致玻利维亚(多民族国)森林生物多样性的大量流失。因此,该计划不仅为养蜂人提供了生计,保护了蜜蜂,而且通过保护授粉维持了现有植物的生物多样性。
约旦是一个干燥且干旱多发的国家。它有限的88000公顷森林覆盖集中在以地中海气候为特征的高原地区。这些森林在保护约旦的动植物种群中发挥着至关重要的作用,但森林和牧场的退化导致了土壤侵蚀、流域面积受损、生物多样性丧失和宝贵的生态系统服务流失(MoP和MoE,2008)。为了保护其有限的森林资源和与森林有关的生物多样性,该国已将其中一些森林宣布为国家级自然保护区,并将管理这些森林的权力下放给一个非政府组织 — 皇家自然保护协会(RSCN)。
32000公顷的达纳生物圈保护区(DBR)建立于1989年(图A),是约旦最大的自然保护区。它蕴含了四个不同的生物地理区和六种植被类型,包括一个重要的相对完整的刺柏林(Juniperus phoenicea),也是柏树(Cupressus se mpervirens)森林群落最南端的家园。此保护区内共记发现了891种植物物种(其中3种是科学上新发现的物种)(RSCN,2018)。该保护区还是449种动物的栖息地,其中许多是稀有动物,有些还受到灭绝的威胁;它们包括沙猫(Felis margarita)、叙利亚狼(Canis lupus arabs)、努比亚山羊(Capra nubiana)、黄爪隼(Falco naumanni)和埃及刺尾蜥蜴(Uromastyx aegyptia)(RSCN,2018)。到目前为止,在该保护区中发现了25种被列为濒危或易危动物,使其成为具有全球重要性的地区(RSCN,2018)。达纳生物圈保护区是由国际鸟盟指定为更大的达纳重要鸟类区域的一部分。这一区域最重要的树种是地中海柏(Cupressus sempervirens)。
皇家自然保护协会灵活的保护方法将环境、社会和经济目标以及当地人民的生计和经济整合于一体。达纳生物圈保护区是四个民族社区的家园,他们分布在保护区内外的约16个村庄或定居点,总人口达31000人,他们均以一种或另一种方式融入保护区的管理。保护区管理计划也与当地经济和农村发展计划很好地融合在一起。该保护区为当地社区提供了85个长期就业机会和数百个兼职工作。当地社区还从出售手工艺品、药用和芳香植物产品、打猎获得的猎物、以及接待游客参观他们的家舍和提供当地传统食物这些活动中获取收入。
在管理计划之下进行牲畜放牧的规定取得了积极效果。该计划包括一项条款,允许社区成员在干旱季节,当保护区以外的草料不足时,将其牲畜带入保护区的某些地方进行放牧。社区还接受轮牧的培训。大多数当地社区的人们具有游牧和农牧背景,该计划中采用的有控制的放牧对他们的生计来说意义很大;这样做同时还增强了当地社区人们的主人翁意识以及对该保护区进行保护的意愿。每年该保护区提供给当地社区17500头牲畜的饲料,价值约为221.9万美元(RSCN,2018)。
生物圈保护区因其生物学和考古意义吸引了许多本地和国际游客。生态旅游基础设施的发展、保护区收费、木材和非木质林产品的销售收入以及旅游活动收入,使皇家自然保护协会获得了可观的收益,用来支持保护区的保护和可持续管理。皇家自然保护协会修建了旅馆、生态小屋、可容纳多达120人30个帐的露营地和一系列步行登山道(RSCN和Wild Jordan,2017)。旅游业在保护区的成功使皇家自然保护协会获得政府和当地人民的信任,有机会从国家和外部进行融资用于保护活动和支持当地社区的生计。皇家自然保护协会还为当地社区提供了能力建设的机会,帮助它们获得经营小型企业的技能以及组织具有法律地位的合作社的方法,从国家金融机构获得贷款以资助社区项目的开展和实施。
标志性的努比亚山羊在国际自然保护联盟濒危物种红色名录中被列为脆弱物种。
©自然保护协会,约旦安曼来自Finan的当地妇女受到皇家自然保护协会开展的手工艺品生产培训,以作为替代创收活动。
©自然保护协会,约旦安曼保护区使森林状况得到改善,特别是在顾及到了当地人和依赖森林的人的需求的情况下。来自巴西的证据表明,不同治理制度下的保护区(可持续利用、土著居民的土地、严格保护和其他方式)的效果与地处、毁林压力、执行力度密切相关(Soares-Filho等,2010)。研究表明,巴西的采伐保护区使每年的毁林面积从2004年的278万公顷急剧降至2012年的46万公顷,减少了74%(Instituto Socioambiental,2015,摘自RRI,2015)。
不丹超过50%的土地都位于保护区内,在距其1997年制定第一个《生物多样性行动计划》20年之际(不丹政府,1997),该国进行了一次评估。该评估显示了保护区在保护物种和提高人们生物多样性意识上的积极成果。然而,他们也发现了挑战:例如,众多利益相关者之间缺乏协调;与保护区管理和技术手段的财务可持续性相关的不确定性;各政策之间的矛盾;监测现状和进展;以及支持当地利益相关者方面存在困难。人类与野生动物之间的冲突也已成为一个重要问题。当地人掌控野生动物对农作物和牲畜影响的权限被限制,这在一些情况下引发了人们对保护政策的反对(Mongbo等,2011;Lham等,2019)(另见可持续狩猎和野生动物管理中插文51)。
有证据表明保护区实施基于权利的森林保护方法可以产生裨益,但不一定每个物种都能从中受益(Campese等,2009)。比如,旅游和运动狩猎或许对某些物种产生积极影响,但对其他物种则没有(Sayer等,2017)。一个成功的基于权利的保护区方法取决于是否具备能力进行监测、能否为社区的传统做法提供支持和对规章制度的执行能力。
根据各国提供的《2020年全球森林资源评估》数据,主要用于保护生物多样性的森林面积有4.22亿公顷,比1990年增加了1.11亿公顷。被指定的森林面积相当于世界森林面积的10%。在全球范围内,最大的部分是在2000-2010年间指定的;在过去十年中,年增幅有所下降(粮农组织,2020)(图42)。其中一些地区位于受法律保护的地区,而其他一些地区则不在其中。这一数字远低于上述保护区森林面积的原因在于,许多保护区被指定用作多种用途(例如,将生物多样性与娱乐活动或生态旅游结合起来)或主要用于其他目的。以巴西为例,其报告的几乎所有的保护区都指定为主要用于社会服务(对依赖森林的人们的文化和生活方式进行保护),只有限制使用的区域才被划为主要用于生物多样性保护。
2010年,在《生物多样性公约》缔约方大会通过的《2011-2020年生物多样性战略计划》(《生物多样性公约》,2010a)中,爱知生物多样性目标11引入了“其他有效的基于区域的保护措施”(OECM)一词,为实现保护保护区以外的生物多样性提供了一种方式,在这些区域主要的管理目标不一定是保护生物多样性。
2018年《生物多样性公约》缔约方大会通过的第14/8号决定将OECM定义为“除保护区以外的地理区域,其治理和管理是为了实现生物多样性就地养护的积极、持续的长期成果,并取得相关的生态系统功能和服务,以及在适用情况下实现文化、精神、社会经济价值和其他与当地相关的价值”(《生物多样性公约》,2018a)。该决定还定义了认定其他有效的基于区域的保护措施的四个标准:该区域当前未被指定为保护区;该地区受到治理和管理;该地区为原地保护生物多样性作出了持续和有效的贡献;以及提供了生态系统功能和服务以及文化、精神、社会经济及其他与当地相关的价值。
IUCN WCPA(2018)和Jonas等(2018)认定了一些森林生境中潜在的OECM的例子,包括:
▸ 由土著居民和当地社区保护的但没有被正式定为保护区的区域(见插文48);
▸ 与国家公园或保护区毗邻的野生生物保护区;
▸ 在国家报告中尚未定为保护区,但以保护为主要目标并颇具成效的私有管理区域;
▸ 正在积极进行生境恢复的区域,其主要目的为恢复具有较高生物多样性和生态系统价值的退化生态系统,例如恢复好的沿海湿地和红树林;
▸ 保存自然生境和动植物的狩猎保护区,以及被狩猎和未被狩猎的本地动物种群;
▸ 森林里一些被永久保存起来免受威胁的区域,如老熟林、原生林或其他生物多样性丰富的森林(见案例研究5);以及
由土著居民和当地社区保护的区域被公认为可为实现爱知目标11做出贡献的重要组成部分(ICCA,“土著和社区保护区域”),无论其是正式的还是非正式的保护区,抑或是其他有效的基于区域的保护措施。土著和社区保护区域各不相同,但通常具有以下三个特征(Borrini-Feyerabend等,2013):
▸ 一个土著居民群体或当地社区与该区域具有紧密而深厚的关系(领域、区域或栖息地);
▸ 这些居民或社区是制定与该区域相关决策的主要参与者,并且具有事实上和/或法律上制定和执行规定的能力;
▸ 不管初始或主要动机为何,这些居民或社区的决定和努力成就了该区域生物多样性、生态功能和文化价值的保护。
土著和社区保护区域包括集体管理的领土、文化场所、圣地、特定物种的庇护所、以及诸如社区森林和牧场、季节性迁徙放牧场、本地管理的海域等可持续使用公共领域。世界养护监测中心拥有一个土著和社区保护区域名录(UNEP-WCMC,2020)。尽管尚未对土著和社区保护区域的数量和范围进行正式的评估,但据估计其覆盖面积可能等于或大于政府指定的保护区。
治理和管理土著和社区保护区域的规章和制度不一而同,从数代人口口相传的不成文习惯法,到正式的成文法。土著和社区保护区域不须成为官方指定的保护区的一部分,而事实上一些土著居民或当地社区并不愿意他们的领地被指定为正式保护区。
许多内陆鱼类依赖由森林维护和支持的淡水生境。高地森林可提供土壤稳定性,减少暴雨期间的破坏性径流,并减少山体滑坡流入下游河流的风险。健康的洪泛区森林为蜿蜒的河道、海狸池塘和水流缓慢的侧渠提供支持。河流沿岸森林为水生食物网提供了遮荫、防止土壤侵蚀、过滤化学品和营养丰富的陆地食物。在美国和加拿大的西北部地区的太平洋沿岸,人们管理和恢复森林来支持淡水生物多样性。
历史上在该地区的森林生境中发现的许多淡水鱼类在1973年出台的《濒危物种法》(美国政府,1973)之下被列为受胁或濒危物种。为了保护水生生物多样性和内陆鱼类社会文化效益,政府成功地实施了一些规模大、协调一致的项目,其中与森林有一定关系的包括西北森林计划、鲑鱼之灵计划(Wy-Kan-Ush-Mi Wa-Kish-Wit),和俄勒冈州白鲑恢复计划。
西北森林计划(美国农业部,日期不详)是有史以来规模最大的协调一致的土地管理计划之一,它前所未有地将管理目标从可持续木材产量转为保护。始于1994年,该计划通过指定庞大的成熟林和河岸林保护区系统,并结合在其他土地上实行有控制的采伐,为1000万公顷联邦土地制定了100年的管理方针。证据表明,在该计划实施之初20年,它保护了茂密的原始森林,并成功地维护了受胁和濒危的鸟类以及一系列水生生物的栖息地(Spies等,2018)。气候变化和与之相关的山火活动的增加导致了该计划之下土地上原始森林意外流失。然而,有益于内陆鱼类生物多样性的水生生境的三个基本要素 — 水温、水生无脊椎动物和河岸地区的自然条件 — 都得到了改善。这些改善可能源于道路的减少以及河岸森林中大树数量的增加(Spies等,2018)。公有土地上的低坡度溪流条件的改善归因于上世纪90年代森林管理标准和准则的变化(Roper、Saunders和Ojala,2019)。
Wy-Kan-Ush-Mi Wa-Kish-Wit意为“鲑鱼之灵”,是一个由内兹佩尔塞(Nez Perce),尤马蒂拉(Umatilla)、暖泉(Warm Springs)和雅克玛(Yakama)四个部落创建并由哥伦比亚河部落间渔委会协调实施的计划,旨在恢复在文化和营养上具有重要意义的太平洋大鲑鱼(Oncorhynchus spp.)(CRITFC,2020)。到上世纪70年代后期,洄游至哥伦比亚河的成年鲑鱼已从欧洲人接触之前的每年1500万条下降至不到50万。在各州、联邦政府以及25个部落的合作之下,该计划通过种植河岸树木、协调流域内森林管理以及在森林健康的地区重新引入鲑鱼等行动,成功地改善了1000多公里溪流。哥伦比亚河下游的邦纳维尔大坝的鱼类统计表明,成年的奇努克鲑鱼(Oncorhynchus tshawytscha)从2001年开始大量增加,到2015年达到130万条鱼的峰值。不幸的是,近年来奇努克鲑鱼的数量急剧下降,这可能是因2015年不良的海洋条件和较高的水温所致,提醒人们任务尚未完成。在鲑鱼洄游增加的地方和时间段,部落成员可以捕获种类和数量都更多的鲑鱼,捕鱼期也更长,更多的部落成员(包括较年轻的一代人)从从捕鱼中获得了就业和收入。太平洋鲑鱼还通过将营养物质(如氮)从海洋运送到它们产卵的有森林覆盖的溪流,从而促进了陆地生物多样性。鲑鱼还直接地(通过其腐烂的尸体)和间接地(通过棕熊(Ursus arctos)(Hilderbrand等,1999)或其他以它们为食的动物)将营养物质转移到河岸土壤中。这些土壤养分通过增加针叶面积从而加速光合作用促进了锡特卡云杉(Picea sitchensis)的生长,提高了它们的活力(Reimchen和Arbellay,2019)。
俄勒冈州白鲑恢复计划于1998年发布实施,其目标是扭转俄勒冈州白鲑鱼(Oregonichthys crameri)的减少。俄勒冈州白鲑鱼是俄勒冈州西部威拉米特河谷特有一种小型淡水鱼(美国鱼类和野生生物管理局,1998)。该计划的活动包括保护现有野生种群、在其历史上活动区域内的洪泛区重新引入该物种、以及提高公众对这一保护问题的认识。在各政府机构、行业组织、科学家和公众人士的不懈努力之下,俄勒冈州白鲑鱼终于于2015年2月从濒危物种名单中被删除,使其成为美国有史以来第一个因恢复管理措施得当而被除名的鱼类。西北森林计划管理之下的威拉米特国家森林对于这些鱼类赖以生存的栖息地的恢复和维持至关重要。
这三个案例的成功都取决于系统之下的多学科的规划和管理,需要森林生态学家、水文学家、淡水生物学家、鱼类生物学家和其他人员的积极参与,以此作为当地实地行动的基础。为了促进水生生物多样性,人们在管理和恢复森林上做出了协调一致的努力,这些努力很大程度上建立在对上游和下游地区之间、森林与河流之间以及人为主导的地区与荒地之间各种关系的理解。来自不同机构、有时甚至是存在互相竞争关系的机构的人们,以及来自不同文化背景的人们,大家共同协作也是成功的一个关键因素。
总之,其他有效的基于区域的保护措施为记录用于生物多样性保护的连续区域提供了机会,从国有保护区到在其他管理形式之下可以对生物多样性保护做出重要贡献的公有、私有和传统拥有的土地,即便保护也许并不是这些土地的管理目标。具体而言,其他有效的基于区域的保护措施可以通过填补空白、连接栖息地和保护在指定保护区之外出现的物种作为保护区有益的补充。然而,正如Dudley等(2018)所指,只有解决了生物多样性丧失的关键驱动因素,并且关键的扶持条件到位(例如尊重人权、保障所有权和给予社会保障),其他有效的基于区域的保护措施才能真正实现为保护生物多样性这一目标做出贡献。
生物多样性已经是可持续森林管理概念公认的要素。《2017-2030年联合国森林战略计划》(联合国,2017a)也明确承认了森林在维持生物多样性方面的作用。
在墨西哥坎昆举行的2016年联合国生物多样性会议呼吁将生物多样性纳入所有农业部门和旅游部门的主流工作。全球环境基金科学和咨询委员会将生物多样性主流化描述为:“将对生物多样性的考虑纳入影响或依赖生物多样性的关键公共和私有部门参与者的政策、战略和实践行动之中,以便养护生物多样性,使其在本地和全球都实现公平和可持续使用。”(Huntley和Redford,2014)。
将生物多样性纳入林业主流意味着优先考虑对生态系统、物种和遗传水平上的生物多样性以及对生态系统服务有积极影响的林业政策、计划、方案、项目和投资(见插文49中的例子)。这涉及加强森林和生态系统中生物多样性的可持续利用,并尽量减少森林部门对所有其他生态系统的影响。
蒙古是一个高度依赖自然资源的贫困国家。大部分人口分布在小城市中心和广袤的草原上,主要活动是放牧牛、羊、山羊、马、牦牛和骆驼。这些(连同社区林业)起到了提供就业、缓解贫困、促使边缘化社区参与国民经济活动的作用。蒙古的森林可持续管理为该国许多穷人提供了其他收入来源,最近在该国试点还引入了参与性森林管理。
粮农组织-全球环境基金-蒙古政府的项目“将生物多样性保护、可持续森林管理和增强碳储纳入蒙古森林景观主流”旨在改善超过460000公顷森林的管理,其中包括濒危物种的重要栖息地,如麝香鹿(Moschus moschiferus)和猎隼(Falco cherrug)。在粮农组织的协助以及全球环境基金的资金支持之下,该项目由蒙古环境与旅游部与各省、地区级政府实施,直接与101个森林用户小组合作。在该项目支持之下制定的所有森林管理计划均包括生物多样性保护目标和野生动物监测活动。
除了旨在增强森林健康、生产力和碳储的活动(如病虫害防治、防火、林分改良)之外,该项目还促进基于薪材、小手工艺品和非木质林产品的创收活动;这些为森林用户小组提供了进行多用途经营森林的机会。迄今为止获得的项目监测数据表明,项目区域内一些野生物种(包括麝香鹿和野猪)的数量有所增加。
认证计划(见插文50中的示例)和REDD+都具有旨在保护生物多样性的强制性环境和社会经济保障措施。现有一些指南可用于将生物多样性纳入森林管理的主流,分别针对生产林(ITTO和IUCN,2009),人工林(Carnus等,2006)和森林恢复(Beatty、Cox和Kuzee,2018)。
大西洋雨林曾覆盖巴西超过1亿公顷的土地,但截至2000年为止,仅剩7%。即便如此,这些分散的小森林碎片仍为某些世界上最丰富的生物多样性提供着庇护之所。一公顷土地上可发现约450种树种,其中一半以上在地球上其他任何地方都找不到踪迹。在这些森林中迄今发现记录的2万种植物约占全球总数的8%,其中有8000种是该地区独有的(Ribeiro等,2009)。
制浆造纸企业及其桉树种植园的到来也许终将成为压倒骆驼的最后一根稻草。根据《巴西森林法典》(1992年第12.651号法律),该地区的土地所有者必须在其个人土地(“法律保护地”)的20%之上以及用于维护生态系统完整性的“永久保护区”中保留自然植被,例如水道周围的植被或防止陡坡土壤侵蚀的缓冲区。一些公司甚至试图规避这一最低标准(Azevedo等,2017)。然而,使用森林认证计划(例如森林管理委员会的认证)的林业公司提供了合法性的保证,其中一些公司甚至做得更进一步,它们指定并管理了一些具有较高保护价值的区域,并率先在森林恢复上做出了工作(新一代种植林,2018)。
参与世界野生动物基金会(World Wildlife Fund)新一代种植林(New Generation Plantations)平台的四家纸浆和造纸公司遵循第三方认证计划制定的可持续发展原则,管理超过200万公顷位于大西洋森林生物群落的土地(Silva、Freer-Smith和Madsen,2019)。该地区约有一半土地种植着桉树,主要在以前用于牧场的退化严重的土地之上。其余的则留作保护用地。这些公司已经恢复了数万公顷的土地,为旨在重新连接森林碎片、确保大西洋雨林的未来的景观规模计划做出了贡献。
同时,由于遗传改良和森林管理研发方面取得的成果,巴西桉树自1970年以来平均生产力(每公顷木材产量)翻了一番还多。这意味着就木材而言,这些人工林是世界上生产力最高的生产林之一,其所有者在营林的同时还增强了景观中的生物多样性。
越来越多的研究证明,由土著居民和当地社区管理的森林在维持森林覆盖方面至少与严格保护的森林一样有效(Porter-Bolland等,2012,Stevens等,2014;Blackman等,2017;Blackman和Veit,2018,Tauli-Corpuz、Alcorn和Molnar,2018)。保护区以外社区管理的森林不仅可以提高森林覆盖,而且还可以提供如野生动物种群的保持和增加这类其他的保护效益,澳大利亚、巴西和加拿大(Schuster等,2019),尼泊尔(Anup,2017),和坦桑尼亚(案例研究6)都有这方面成功的例子。
坦桑尼亚联合共和国拥有约4810万公顷森林,约占总土地面积的55%。林地为本国农村和城市提供了95%的能源,以及75%的建筑材料。森林还提供各种非木质产品,并在储水方面起了很大的作用。然而,森林正在承受来自人类居住、非法采伐、木炭生产、火灾、采矿和基础设施建设等诸多方面的压力,导致每年约372816公顷森林被清除(MNRT,2015)。
坦桑尼亚联合共和国在其应对气候变化的国家自主贡献中,已经认识到森林对于应对气候变化和实现该国减排目标的重要性。其国家自主贡献是少数几个强调扩大参与性森林管理(PFM),协调一致地执行REDD+行动,并加强对天然林的保存和保护的国家之一。
坦桑尼亚联合共和国具备非洲最先进的习惯法法律体系之一,认可土地所有权和参与式森林管理。习惯性土地权利在村庄范围内得到承认,参与式森林管理也已被纳入政府工作计划的主流。社区总共拥有近2200万公顷林地。参与式森林管理在米奥姆波(Miombo)林地最为普遍,据估计占该国林地的90%以上(Lupala等,2015)。
在参与式森林管理之下的地区,不受约束的采伐和其他森林干扰有所减少;森林状况明显恢复;土壤侵蚀和过度放牧状况减少,水质和水量得到相应的改善;蜂箱重新变得旺盛;野生动物数量总体上有所增加(Patenaude和Lewis,2014)。相比之下,开放式森林地区则遭遇不可持续的行为,例如农业扩张、山火、过度放牧、非法攫取木材和非木质林产品(Blomley等,2008;Burgess等,2010)。
遵循《土地权属负责任治理自愿准则》(粮农组织,2012b),承认习惯性土地所有权和将土地和资源权利下放到地方的举措,赋予了当地人民管理自己资源的自主权。允许社区组成自己的理事机构并制定自己的规则,是赋权当地人民进行可持续管理森林和其他自然资源的第一步。例如,允许对巴加莫约(Bagamoyo)沿海村庄森林保护区进行集体管理避免了一系列不利威胁(包括不可持续的狩猎,为获得木材、木杆、木炭和手工艺品而进行的砍伐),因此保护区内的毁林得到了控制(见图A)。
坦桑尼亚联合共和国Shimbwe Juu村庄的查加部落社区成员。
©粮农组织/Felipe Rodriguez然而,坦桑尼亚联合共和国的参与式森林管理计划在促进生计方面尚未充分发挥其潜力。面临的挑战包括执行上的延误、对土著居民的认可不足、权利下放有限(特别是在联合森林管理中)以及难以让牧民参与其中。虽然在承认集体权属方面取得了进展,但仍需要注意一些更大的森林治理问题,包括激励制度、加强社区机构以及增加投资和人力资源。
许多研究旨在评估保护和开发项目对当地社区的影响(Plumptre等,2004;West、Igoe和Brockington,2006;Sayer等,2007)。然而,很少有研究同时考虑对保护区和地方社区的影响,而且在实践中也较少有双赢的解决方案(Southworth、Nagendra和Munroe,2006;Chan等,2007;McShane等,2011)。已经发现的一些弊端包括预定的保护目标和不可商议的保护区边界(Sharpe,1998);转移给地方机构的权力有限(Ribot,2002);森林管理权下放后,资源被精英阶级获取(Persha、Agrawal和Chhatre,2011);有限的排他权;此类项目在政府政策转变和支持不确定时表现出的脆弱性(RRI,2015)。
野生动物的获取和消费依然对千百万人民的粮食安全、健康、文化和生计至关重要。无监管的狩猎是某些物种流失的一个重要原因(见第3章)。然而,与许多人的看法相反,可持续利用其实是一种行之有效的野生动物保护机制。事实上,在一些地方,消费野生动物的人群仍然是野生动物管理和国家运作的保护工作的主要贡献者(案例研究7)。
当第一批欧洲定居者到达时,美国和加拿大的野生动物相对丰富,但是到19世纪后期,许多物种由于商业开发而濒危或濒临灭绝。例如,到1889年,美洲野牛(Bos bison)的数量已从2000万头减少到大约1000头。到1902年,曾经至少有30亿只的旅鸽(Ectopistes migratorius)已在野外灭绝。其他受胁的物种包括麋鹿(Cervus canadensis)、骡鹿(Odocoileus hemionus)、白尾鹿(Odocoileus virginianus)、野生火鸡(Meleagris gallopavo)、木鸭(Aix sponsa)和叉角羚(Antilocapra americana)。面临这些资源危机,人们的社会责任感触发了基于公民责任和自然极限的资源使用哲学的崛起,最终发展成为一套由惯例、政策和法律组成的系统,即北美野生动物保护模式(美国鱼类和野生生物管理局, 2018;Mahoney和Geist,2019)。这一模式基于七个主要元素:
▸ 野生动物是一个公共信托资源。
▸ 消除野生动物市场:禁止商业狩猎和野生动物买卖,以确保野生动物种群的可持续性。
▸ 野生动物是依据法律分配给公众的(而不是根据市场原则或土地所有权分配给公众)。
▸ 只可以出于合法目的(食物、毛皮、自卫和保护包括牲畜在内的财产)猎杀野生生物;在没有尽一切合理努力来获取和合理利用资源的情况下,残害鱼类或野生动物(即便是获得许可)被广泛认为是非法的和不道德的。
▸ 野生动物被认为是一种国际资源。
▸ 科学是执行野生动植物政策的恰当工具。
▸ 狩猎民主,即公开准入 — 因此,猎人是野生动物保护资金的重要贡献者。
自20世纪初以来,这种模式促进了已捕获和未捕获野生动物物种的显著恢复以及可持续消费。这种恢复的突出例子包括野生火鸡和白尾鹿,这两种动物在殖民之前对土著居民来说都是重要资源,种群数量据估曾经均在1000万以上。
到20世纪初,由于无节制狩猎和栖息地丧失,野生火鸡数量已减少到20万。狩猎组织推动了早期立法,促进了野生火鸡的保护和研究。最初在恢复上的尝试基于将圈养的火鸡放归自然,但大多没有取得成功。后来又发展成利用改进的捕获技术来捕捉野生火鸡,然后将其转移到合适的、无人居住的栖息地。从1986年开始,启动了一个复杂的在州与州之间转移野生火鸡的系统。如今,野生火鸡种群已经恢复到接近殖民前的数量,2013年估计为700万只。可进行种群自我维持的野生火鸡现在存在于美国50个州中的49个州、六个加拿大省以及墨西哥中部和东部(Hughes和Lee,2015)。
白尾鹿同样也在商业化狩猎和栖息地丧失面前非常脆弱,到十九世纪末总数减少到50万只。相应地,猎人做出了促进和协助执行狩猎法规、迁移鹿群以及资助养护和管理项目的行动来应对。许多猎鹿人甚至购买或租赁了可以保护或繁殖鹿群的土地。早期一些人将鹿引入美国八个州中无人居住的地方,企望有一天鹿群可以发展到进行狩猎的规模。今天,美国约有3000万只白尾鹿,加拿大约有40万只。该物种现在是北美最受欢迎的大型猎物,并且仍然是一个重要的食物来源,尤其是在农村地区。
为了配合《生物多样性公约》14/7号关于可持续野生动物管理的决定(《生物多样性公约》,2018b),国际林业研究中心(CIFOR)和《生物多样性公约》在与野生动物合作伙伴关系的合作之下,提出了以下一系列有关野生动物肉可持续利用的建议(Coad等,2019):
▸ 创建一个有效的有利环境。可包括:
— 与广大利益相关者协商,修订国家狩猎法,以确保这些法规虑及粮食安全和野生动物保护问题,并且可以公平、切实地得到执行;
— 在国家执法机构的支持下,将土地权属下放给土著居民和地方社区;
— 构建区域和国家野味肉类监测框架,促进有据可依的决策。
▸ 管理农村供应并减少城市对野味肉类的需求。商品链中彼此联系的干预措施可以包括社区管理或共同管理的保护区、野生动物牧场、社区保护组织、生态系统服务付费(PES)计划和认证机制。从事木材采伐、采矿或森林中集约农业活动的公司必须通过为员工提供替代食物(如牲畜肉类)、与当地社区合作帮助公平狩猎法规的实施、防止外部商业猎人使用特许经营的道路和车辆等途径,采取措施以确保其特许经营范围内野味肉类猎取和使用的可持续性。在新兴的城市地区,野生动物种群严重枯竭但其替代品还没有广泛可用,政府和发展机构应帮助开发如牲畜肉等替代食品。在大都市地区,野味肉通常被视为奢侈消费,干预措施可包括针对改变消费者行为的运动,同时执行对野生动物肉贸易进行适当管制的法律。一项可能实现保障粮食安全和营养、促进可持续的地方收入和环境健康的举措就是加强对速生野生动物物种的可持续管理。
▸ 促进有据可依的参与式管理。建立食肉为目的的野生动物管理项目必须在社区的充分参与和同意之下进行。此外,项目设计必须纳入变动理论和为应对变化而做的监测和评估,以便让项目的成败可以为将来的管理干预措施提供依据。
自2017年10月以来,包括粮农组织、国际林业研究中心、野生动物保护协会和法国国际发展农业研究中心(CIRAD)在内的合作伙伴联盟一直在实施一项为期7年的可持续野生动物管理计划。该计划的目的是在12个非洲、加勒比和太平洋国家制止不可持续的野生动物狩猎,保护生物多样性和自然遗产,并增强人民的生计和粮食安全。在每个国家中,该计划旨在改善体制和法律框架,使野生动物肉的可持续使用适应狩猎和捕鱼,以及对这些野生物种的管理;增加替代蛋白质的供给;将野生肉的消费减少到可持续水平。该计划强调了监控、评估、学习和知识对于最终升级的重要性。该计划由欧盟资助。
野生动物管理还需要处理人与野生动物之间的冲突,尤其是在为了方便野生动物迁徙而未在保护区周围设置围栏的情况之下。见插文51。
人类与野生动物之间产生冲突一般发生在当动物对人类的生计或安全构成直接和反复的威胁时,常常引发了对该物种的迫害。在许多区域,这些冲突随着人口增长和土地使用的变化变得更为严重。总的来说,人类与野生动物冲突的后果包括农作物遭到破坏、农业生产率降低、互相争夺牧场和饮水、牲畜被捕食、农民受伤亡、基础设施被破坏、以及疾病从野生生物传染到牲畜的风险增加。人类与野生动物冲突通常会引发对保护的消极情绪,尤其是在建立或扩大保护区时。
人类与野生动物冲突是非洲野生动物保护和人类福利的主要关注点。例如,仅纳米比亚在2017年就有超过8000 例人类与野生动物的冲突事件发生(世界银行,2019)。2011-2016年间,鬣狗在纳米比亚的赞比西地区杀死超过600头牛,同时期还发生了超过4000起农作物受损害的事件,大多数是由于大象在该地区活动而引起的(NACSO,2017a)。人类与野生动物冲突也已成为亚太地区许多国家的主要问题。例如,斯里兰卡每年有多达80人因大象而丧生,有230多头大象被农民杀害。斯里兰卡大象被列为濒危物种,只有2500-4000头在野外生存(IIED,2019)。
对森林来说,高密度的有蹄类动物群(例如鹿)会严重破坏森林,通过践踏或嚼食小树、在树上摩擦、蹭下树皮等方式威胁到森林的再生。这种行为可能具有重要的经济影响,并可能导致森林和野生动物管理者之间水火不相容的矛盾(CPW,2016)。
已经开发出许多对策预防和减轻人类与野生动物冲突,大致可分为致命和非致命的。从需要昂贵的基础设施(如电围栏)和政府介入(例如补偿和保险计划)的方法,到只需单人使用低成本工具即可实施的方法(如保护牲畜、烧辣椒砖),不一而同(Nyhus,2016)。建立和维护起来成本较低的蜂巢围栏,是解决人类与大象之间冲突的一个创新的方式,已经被肯尼亚农民自愿采用。这些栅栏是一种自然的威慑手段,它利用了大象避开非洲蜜蜂的本能,同时又提供了授粉服务和“保护大象的”蜂蜜(King等,2017;拯救大象,2019)。
为应对这一挑战,许多国家开始明确将解决人类与野生动物冲突纳入国家野生动物管理、发展和扶贫政策和战略之中。在国家一级,林业、野生动植物、农业、畜牧业和其他相关部门之间的跨部门合作是关键。粮农组织通过促进跨部门对话,在制定国家政策和法律框架方面提供技术援助,帮助分享有关好经验和工具的信息等做法,积极支持成员国为更好地管理人类与野生动物冲突所做的努力。例如,粮农组织在2010年开发了一个针对人类与野生动物冲突的工具包(Le Bel、Mapuivre和Czudek,2010),供南部非洲的农民和当地社区使用,现已将其改编并翻译成法文,供中部非洲使用(Nguinguiri等,2017)。
蜂巢围栏。
©Lucy King由《2020年全球森林资源评估》得到的数据以及世界保护监测中心专门为本版出版物所做的一项研究得出,在全球范围内,森林生态系统作为一个整体已经超越了爱知生物多样性目标11(到2020年保护至少17%的陆地面积)。虽然没有专门对森林保护区的总体成效做过评估,但是鉴于只依赖森林的人口指数在1970-2014年间下降了53%(见测量森林脊椎动物种群的趋势),毫无疑问还是有改进的空间。
就“生态学上具有代表性的、连通性良好的保护区系统”而言,按全球生态区划分的对保护区的分析(见关于保护区趋势最新研究)表明,目前不到10%的亚热带湿润森林、温带草原和北方针叶林处于被保护的状态。
其他应予以高度重视的领域包括对生物多样性的重要性和完整性有价值的地区,例如安第斯山脉北部和中美洲、巴西东南部、刚果盆地的部分地区、日本南部、喜马拉雅山、东南亚和新几内亚的部分地区(图22)。
鉴于其他有效的基于区域的保护措施是一个较新的概念,在将特定的森林区域归为此类区域方面的进展有限,但有关这一类别的指南正在制定中,对森林具有巨大的潜力。
如本章中的案例研究所示,对保护区内外的森林生物多样性进行保护采用的方法表明,在实现生物多样性与社会经济平衡上取得了一定程度的成功,也许可以进行复制或扩大规模。成功的基本要素包括参与性方法、对产权的重视、跨部门方法(也称为领域或景观方法)和能力建设。直接或间接地对当地收入或商业机会产生积极影响的经济方法,也可以在激励产生积极的生物多样性成果中发挥重要作用。
可持续森林管理,如联合国森林文书(联合国大会,2008;UNDESA,2016)所述,将森林生物多样性作为其七个主题要素之一。1如果能得到成功实施,它可以确保在保护和社会经济发展方面取得积极成果。由于没有任何单一的可量化和可衡量的特征能够充分描述可持续森林管理的诸多社会、环境和经济方面,所以可持续发展目标指标15.2.1(实现可持续森林管理的进展)(见插文52)不易衡量。在认识到这一点的同时,粮农组织与合作伙伴共同开发了一种监测汇报该指标的方法,并建立了一套由五个子指标组成的体系来衡量进展情况:
▸ 森林面积年净变化率;
▸ 森林中的地上生物质储量;
▸ 位于合法建立的保护区内的森林面积比例(表明为保护和维护生物多样性以及其他自然和文化资源而采取的行动);
▸ 长期森林管理计划中森林面积的比例(表明长期管理森林的意图);
▸ 经独立的森林经营认证计划验证的森林面积(进一步提供森林经营资格)。
▸ 可持续发展目标15.2:到2020年,推动对所有类型森林进行可持续管理,停止毁林,恢复退化的森林,大幅增加全球植树造林和再造林。
— 可持续发展目标15.2.1:实施可持续森林管理的进展。
▸ 爱知生物多样性目标7:到2020年,农业、水产养殖业及林业用地实现可持续管理,确保生物多样性得到保护。
▸ 联合国森林战略计划目标3:大幅增加世界各地森林保护区及其他可持续管理林区的面积,提高可持续管理林区林产品所占比例。
— 具体目标3.2:大幅增加具有长期森林经营方案的森林面积。
前三个子指标涉及森林的环境价值,而后两个考虑可持续森林管理的所有维度,包括社会和经济方面。前四个子指标的数据通过森林资源评估国家报告流程收集,而有关认证森林面积的数据则是从主要认证机构获得。对于每个子指标,在可持续发展目标元数据存储库中提供了有关定义和方法的详细说明(联合国,2020年)。所有数据呈现在一览表中,用来显示每个子指标的进度。虽然已经在后三个子指标上取得进展,但由于森林面积的净损失,前两个子指标在全球范围内显示出负增长的趋势。
关于《联合国2017-2030森林战略计划》的目标3.2(联合国,2017a)(见插文52),《2020年全球森林资源评估》的数据表明,在长期管理计划之下的森林面积在过去30年中显著上升到20.5亿公顷(相当于全球森林面积的54%)(粮农组织,2020)。
尽管前几章指出在保护森林和森林生物多样性方面正在取得进展,但生物多样性的广泛丧失继续对人类福利和安全构成严重威胁。IPBES(2019a)在评估可持续发展目标之间一系列的相互作用时发现,当前生物多样性和生态系统状况的不利趋势将对实现80%(44个中的35个)的SDG目标的进展造成负面影响。因此,现在的问题不仅是经济发展活动对生物多样性的影响,而且还有生物多样性(或更确切地说,生物多样性的丧失)对经济发展的影响。
本章探讨了生物多样性保护与其他可持续发展目标之间的取舍和协同作用,并提供了一些成功方法的实例。本章还进一步概述了有利于取得平衡解决方案的环境的一些关键要素,并展示了一些有创新意义的方法以实现对进展的监控。
《2018年世界森林状况》强调了森林对可持续发展目标的潜在贡献。国际森林研究组织联合会关于世界森林、社会与环境的特别项目的一个最新报告(Katila等,2019)分析了可持续发展目标对森林的影响。这两份报告都强调了森林在实现可持续发展目标方面的关键作用。虽然各个可持续发展目标之间相互关联、不可分割,增进可持续发展目标之间协同关系的不同行动会促进彼此,但短期内可能会需要权衡。
Katila等(2019)得到的三个关键信息尤其说明了这一点:
人的需求影响着人们对森林价值的认可。鉴于人及其利益千差万别,一个或多个可持续发展目标的实施在许多情况下对森林产生不同影响,因而有赢家也有输家。
预先设定的森林保护与社会发展之间是正相关的假设存在误导性。增加森林面积并非永远是满足复杂发展需求的最佳解决方案,而实现某些可持续发展目标可能会导致森林损失,可能会推动社会和经济发展,例如通过农业扩张或更多住房和基础设施空间。
至关重要的是,要理解可持续发展目标中与森林和其他土地用途有关的潜在的权衡取舍,并在社会和政策决策中予以充分考虑。这必须包括跨不同层面和代际的考量。它还必须包括让依赖森林的人们发声,他们有可能被旨在推进可持续发展目标议程的措施所忽视。
生物多样性的丧失往往给已经处于不利地位的人们,特别是最贫困人口、妇女、儿童和土著居民造成更大的损失。在损失危及人们生存的地区,这种退化往往加剧冲突或移徙,成为安全问题。生物多样性下降也日益威胁到粮食安全和营养(粮农组织, 2019a)。如第4章所述,粮食生产依靠森林的完整性为其提供重要的生态系统服务,以支持可持续农业和农业系统应对气候变化的适应力。然而与此同时,农业扩张是对森林生态系统完整性的最大威胁,而毁林是农业、林业和土地利用所造成的温室气体排放的主要来源之一,其占人为总排放量的23%(IPCC,2019)。因此,解决生物多样性丧失的办法不仅需要考虑森林和邻近人口的需求,而且还需要满足农民的需求,从广义上讲,他们也是依赖森林的人。对于生物多样性和人类而言,气候变化导致更广泛的生态系统和栖息地上的变化,增加了破坏和损失的风险。
处理各个可持续发展目标之间固有的权衡取舍关系是具有挑战性的,但是至少逐渐成型的评估框架可以使这些关系更明确可见,为政策制定者就如何处理不同类型的关系提供信息(如Nilsson、Griggs和Visbeck,2016)。
要取得确保生物多样性和人类的积极成果,就需要与所有利益相关方共同努力,在保护目标和为了生计而产生的资源需求之间找到现实的平衡(Kaimowitz和Sheil, 2007)。这可能意味着,至少在某些地方,可接受的标准要低于传统对未受干扰生境进行保护所规定的标准,但在满足当地需求(在资源、生计和赋权方面)的同时足以维持必要的生态系统服务和生物多样性,有助于树立对保护区和其他保护措施更积极态度。真正的赋权于当地人民的参与性措施,结合开发替代资源的激励机制,可以促进对人类和保护均有利的可持续性更好的森林经营模式。
尽管很少有案例能够完美地平衡生物多样性的保护和当地的生计需求(Hoffmann等,2012),但本版《世界森林状况》提供了一些积极的例子,表明是可能实现的。
如案例研究8所示,有机商品标准和公平贸易标准等市场工具可以激励可持续生态系统管理;使当地人能从林产品(在此例为药用植物)中获取经济利益,同时为脆弱的野生动物(在此例为大熊猫)维持栖息地。可以在世界其他有相似景观的地区探索类似途径,例如,猴面包树(Adansonia digitata)与非洲东部和南部濒危的非洲丛林象(Loxodonta africana);美国西洋参(Panax quinquefolius)和美国黄褐森鸫(Hylocichla mustelina);尼泊尔的印度大花甘松(Nardostachys grandiflora)和雪豹(Panthera uncia)(Jenkins、Timoshyna和Cornthwaite,2018)。
尽管人们从植物种植中获得了许多收益,但据估计市面上出售的60%至90%药用和芳香植物(MAP)物种仍然是从野外收集的。从森林中和森林附近收集的野生植物为卫生保健、化妆品和食物生产部门提供了重要的原材料,为千百万人的生计提供了支持。然而,过度攫取、土地转化和污染是对世界许多地区的野生物种及其采集者的主要威胁:五分之一的药用和芳香植物物种濒临灭绝(Jenkins、Timoshyna和Cornthwaite,2018)。
许多野生植物与其他受胁物种共享生境。因此,可持续的野生植物采集和贸易是系统地管理其他物种和广泛的生态系统的基础。
中国在药用和芳香植物国际贸易的重要参与者,2013年出口总量为130万吨,价值50亿美元(占全球药用和芳香植物出口的15.6%)。其中,野生采集材料价值约为18亿美元(ITC,2016)。这些贸易大部分与中药使用的成分有关,超过70%来自野生药用植物。仅中国甘草(Glycyrrhiza uralensis)、虫草(Cordyceps sinensis)、枸杞(Lycium barbarum)、茯苓和辽藁本根每年出口金额就达1.8亿美元。
在长江上游生态区的村庄里,药用植物的销售可达家庭收入的60%(Jenkins、Timoshyna和Cornthwaite,2018)。该地区有益于熊猫的南五味子藤(Schisandra sphenanthera)保护模式的十年经验提供了有力的证据,表明标准和规范可以在有效地促进可持续资源管理的同时,增强地方和农村社区人们的收入和健康,特别是那些贫困和边缘化人口(Brinckmann等,2018)。
五味子藤生长于为大熊猫(Ailuropoda melanoleuca)提供栖息地的高山落叶森林之中,其浆果用于四川少数民族的草药以及中药。中欧传统药用植物可持续管理生物多样性计划支持应用现有的可持续性标准,例如美国农业部(美国农业部,日期不详)和公平野生基金会(公平野生基金会,2019)的野生作物采集实践标准,同时也支持专门制定的大熊猫友好型产品认证标准(世界自然基金会-中国,2012)。收集者接受了有关五味子果可持续收获方法的培训;例如,他们学会了从五味子藤的下三分之二处采摘浆果,其余的则留给鸟类和野生动植物,使种子在森林中传播。这些标准的应用带来了新成立的本地贸易合作社与国际公司之间的长期公平贸易协议,因此价格上涨了30%。该模式已扩展到22个村庄,涉及的家庭数量从48个增加到300个,自2009年以来野生五味子的收获量增加了六倍,到2017年产量达到30吨(见图A)。
收入的增加为社区提供了激励机制,鼓励他们可持续采摘五味子果,并在大熊猫保护区之外维持次生森林栖息地(Brinckmann等,2018)。大熊猫种群数量目前已经稳定下来,甚至在部分区域内还有所增加(Brinckmann等,2018年引用四川林业局,2015),其在自然保护联盟红色名录中的地位已从濒危变为易危。
熊猫。
图片由 Zoe Nicolaou 于 Unsplash 上提供印度西高止山脉也采取了类似方法,一个采用公平野生(FairWild)标准的项目(公平野生基金会,2019)(目前除木材外最全面的野生真菌、地衣和植物认证体系)在那里鼓励当地社区(包括Mahadev Koli部落居民)采集和出售诃黎勒和毗黎勒果实,而不是为获得薪柴而采伐树木。该项目已经为约2000棵诃黎勒树和500棵毗黎勒树提供了保护,从而保护了该地区两种最具特色的鸟类 — 双角犀鸟(Buceros bicornis)和冠斑犀鸟(Anthracoceros coronatus)— 筑巢和栖息的地方(Jenkins、Timoshyna和Cornthwaite,2018;Yearsley,2019)。
如案例研究9所示,完美整合于一体的景观保护和管理方法有多重益处,不仅对生物多样性和社会经济发展(如收入多样化、就业和赋予妇女权能)有益,而且还有助于永续地提供其他生态系统服务(例如保护水资源、防止侵蚀和减轻灾害风险)。这些方法体现了可持续森林管理的理念。
在摩洛哥进行的一个具有参与性和适应性的流域管理项目,展示了减少社区所面临的灾害和气候变化风险如何可以在丰富生物多样性的同时减少贫困。
位于摩洛哥高阿特拉斯山脉与中阿特拉斯山脉之间的上穆洛亚盆地,由于其脆弱地形、干旱气候以及农村和邻近城市地区的农牧活动,非常容易遭受侵蚀、洪涝和土地退化的影响。1970-2010年间,树木覆盖下降了30%以上,土壤侵蚀增加了60%以上。从1995年到2011年,奥塔特(Outat)河水灾造成的损失价值约为540万美元。
在过去九年(2010-2019)中,分两个阶段实施的项目将景观和风险的理念应用于该流域的综合流域管理之中。在地点选择上,进行了危害和风险评估,识别和认定出风险最高的地点。在省和社区两级准备、讨论和商定了基于风险的计划,对两个流域进行共同管理,涉及面积约16万公顷。这些计划包括结构性措施(例如400公顷土地上的沟渠和泥沙控制设施),以及非结构性侵蚀控制措施(例如对裸露的斜坡进行再造林和重新植被)。
该项目通过围栏、修复和混农林业恢复了480公顷的森林和牧场。恢复工作包括修建保护圆叶栎(Quercus rotundoflia)和大西洋雪松(Cedrus atlantica)森林的围栏和种植梣树(Fraxinus dimorpha)。同时在腓尼基刺柏(Juniperus phoenicea)、大果刺柏(Juniperus oxycedrus)、Hertia maroccana、迷迭香(Salvia rosmarinus)和其他本地灌木的自然再生上取得了良好成果。
该项目通过一系列创收方案解决了社区中贫困和营养不良问题,包括:
▸ 种植本地药用植物;
▸ 生产经认证的苹果醋;
▸ 向九个合作社分配蜂箱,2018年生产了8700升蜂蜜,净收入为174000美元;
▸ 支持妇女合作社生产芳香和药用植物,例如迷迭香、薰衣草、鼠尾草和玫瑰,年产精油达到850升;
▸ 果树栽培、奶制品加工和牲畜计划。
除加强农业生物多样性外,方案还支持收入多样化、农村青年就业和增强妇女权能。
社区的认同和积极性对项目的成功至关重要。参与该项目的合作社、社区和个人愿意采用创新技术和方法,并在该项目初始投资的基础上取得主动,起到主导的作用。在大多数情况下,业务得到了扩大。例如,药用植物合作社成立了一个苗圃,出售产品并确保其精油生产原材料的稳定供应。
该项目展示了在流域综合管理的各个阶段考虑风险的必要步骤,包括选址、流域综合规划和项目实施。社区居民亲眼目睹这些措施是有效的,于是他们在自己的项目中主动复制使用了相同的干预措施。机械性水土流失控制等创新技术现在也正在其他地域推广实施。
由艾尔加泽瑞尔(Eljazera)芳香和药用植物生产和增值妇女合作社生产的草本精油。
©Yuka Makino/粮农组织尽管为建立和加强有关生物多样性的全球治理框架进行了数十年的努力,并且已取得了一些进展,但如本章所述,如果继续按照当前的步伐和节奏的话,显然是无法实现可持续发展目标、《生物多样性公约》和其他全球承诺和框架设定的保护目标的(IPBES,2019a;环境署,2019)。
有效的治理对于保护生物多样性至关重要,并且似乎是决定以生物多样性为导向的政策能否取得成功的最重要因素(Baynham-Herd等,2018)。尽管腐败和贸易已被公认为森林生物多样性的关键挑战,但与森林使用、权属和决策地相关的其他方面也对界定环境是否有利于生物多样性保护十分重要。
由于生物多样性是可持续发展的基础,而对森林生物多样性的大多数威胁都来自森林部门以外,因此,所有国家都必须制定和实施跨领域战略,以实现其生物多样性目标,并将其与实现《2030年议程》和可持续发展目标的努力结合起来。
为达到效果,这种跨领域战略必须包括专注于目标的部门和行政级别之间的协调。
在与相关利益方协商的前提之下,在国家和国家以下各级进行土地综合利用规划是另一个关键要求,应包括对发展状况的设定、确定其他保护区的优先级(牢记需要针对那些没有被充分代表的生态系统或森林类型、具有高度生物多样性意义和完整性的地区、关键物种或物种组)、以及恢复建立生物廊道和可持续管理现有森林的优先区域。第2、3、5和6章中描述的地理分析和评估可以相对容易地在国家和国家以下级别进行复制实施。
如果要改变土地利用方式,则需要采取一致的财政政策,其中首要的事项是研究农业补贴,因为农业是毁林的最大驱动力。
据估计,在经济温和增长的情况下,到2050年农业产量需要比2013年增长50%,才能满足因快速增长的人口和饮食习惯的改变而引发的需求(粮农组织,2017e)。如果不改变当前生产和消费食物的方式,那么产量的增加就可能对森林和生物多样性产生显著的不利影响。致力于零毁林的商品链、减少粮食损失和浪费、恢复农业用地的生产力、采取混农林业和可持续农业生产方式、以及减少土地转变需求的饮食,这些方面的努力都有助于减轻负面影响(例如参见粮农组织,2019a;粮农组织,2019j; IPCC, 2019 和Willett等,2019)。《2016年世界森林状况》提供了七个案例研究,展示了一些国家如何在提高粮食安全的同时能够增加森林覆盖。参见粮农组织(2016b)关于所获得经验教训一节。另见森林与土地利用联盟(2019)和插文53,了解向更可持续的农业和粮食系统迈进所需要的过渡。
在最近从粮食和农业视角评估生物多样性状况的权威报告中(粮农组织,2019a),众多国家均提到了对调节和支持生态系统服务产生负面影响的驱动因素,该因素就是土地和水资源利用的变化及其管理。森林和水生生态系统的丧失和退化,以及在许多生产系统中发生的向集约化生产少数物种和品种的过渡,仍然是丧失生物多样性和生态系统服务的主要驱动力。为粮食和农业提供大量必需服务的关键生态系统正在迅速减少。
同一项评估发现,有利于粮食和农业生物多样性的可持续利用和保护的管理措施和方法的使用日益增加。80%的报告国表示,在一种或多种类型的生产系统中使用了一种或多种以生物多样性为重点的措施(同上)。
粮食和农业生物多样性的可持续利用和保护要求在生产系统及其周围生境中采取综合管理生物多样性的方法。这就要求将原地或农场管理纳入生态系统或景观一级的战略,包括如可可或咖啡生产的农林系统、常绿农业(农林混植)、林牧或农林牧混作系统、或对生物多样性有利的红树林水产养殖。
一系列通过整合的方式使农业和粮食系统更具可持续性的方法涌现了出来,其中包括主流化生物多样性。粮农组织最近启动了全新的愿景和方法以促进可持续粮食和农业,明确要求考虑跨部门(例如农作物、畜牧、渔业、水产养殖和林业和多目标(例如经济、社会和环境)的政策和工具,并确定如何协调和权衡各个部门和目标之间的关系(粮农组织,2019j)。该方法的核心是会员国在2016年批准的五项基本原则:
▸ 提高粮食和农业资源的利用效率;
▸ 采取直接行动来保存、保护和增强自然资源;
▸ 保护和改善农村生计、公平和社会福利;
▸ 增强人们、社区和生态系统的适应性;以及
▸ 负责任和有效的治理机制。
资料来源:粮农组织,2019a;粮农组织,2019j。
可以通过以下两种方式协调粮食生产与生物多样性保护:节约土地法(一个区域的高产农业有助于节约其他区域的土地,使那些土地可以专门用于自然保护)或共享土地法(在同一块土地上整合生产和生物多样性保护,共同实现两个目标),例如生产性混农林业系统(Phalan等,2011)。后者可以同时为生物多样性和农民带来多种利益,包括遮荫和微气候调节、土壤肥力、疾病控制以及应对气候、疾病和市场风险的收入多样化(Schroth等,2004)。
大型农业公司的政策和实践也需要与生物多样性保护目标保持一致。2014年首次获得批准的《纽约森林宣言》在这方面是一个重要的里程碑,它将政府、公司、民间社会和土著居民组织在消除毁林方面所做的努力联系在一起。然而,正如其《五年评估报告》(NYDF,2019)所强调的那样,迄今为止所做的努力还不足以实现系统性的改变。同样,一项对企业零毁林供应链承诺的追踪举措表明(Forest Trends,2017;Ceres,2019),还有很多工作需要去做,尤其是针对驱动毁林和森林变化的四个最大的商品链而言(图43)。
正如参加全球“跨部门协作以制止毁林和增加森林面积:从愿望到行动”会议的与会者所建议(插文38),“农业企业应履行其到2020年在农产品生产和加工中零毁林的承诺。尚未做出零毁林承诺的公司也应该这样做。大宗商品投资者应采取对环境和社会负责的商业模式,并通过例如推广项目和在公司土地上推行可持续土地综合使用计划等措施,使地方/社区生产者、分销商和其他价值链上的行为方参与并获益。”
在这方面,世界粮食安全委员会于2014年批准的《负责任农业和粮食系统投资原则》(粮安委,2014)提供了一个重要参考。
一些农业银行正在发挥带头作用,设立资金,提供贷款、技术援助和其他降低风险的手段,并部署混合融资手段(使用发展融资或慈善资金来调动私人资本流向新兴市场和前沿市场)以支持对可持续农业的投资(另见下文利用私人融资杠杆)。
保障土地权属为保护生物多样性的举措奠定了基础。虽然世界上大多数森林是公有的,但据估计约有15亿当地人和土著居民通过基于社区的权属获得了森林资源的权利,这些地方团体管理着世界18%的森林面积(RRI,2015)。在这些权利得到有效执行的地方,非洲、亚洲和拉丁美洲的国家正在目睹毁林速度的逐渐降低。例如,最近在秘鲁进行的一项研究发现,有迹象表明给予土著社区土地所有权可以在授权后不久就看到毁林和干扰的减少,部分原因是对相关社区及其内部施加了正式和非正式的监管压力(Blackman等,2017)。另请参见第6章(第127页)将生物多样性纳入社区管理森林的主流。
在世界许多地区,为农业而清理森林以获得土地使用权仍然是一种普遍做法,通常是在界限划定不佳、管理不力的依习惯保有土地或公有土地之上。习惯保有制的领导人或国家可以通过向农民提供替代土地或在土地稀缺的情况下提供长期有条件的土地租约,使用户从事混农林业或与生物多样性保护相互兼容的其他土地和资源利用,来防止这种活动。例如,这种方法已在印度尼西亚苏门答腊的楠榜省成功实施;根据社区林业计划(又名Hutan Kamasyarakatan),贫困农民获得为期25年的国有林租赁合同进行混农林业。该方案增加了以木材为目的树木和其他多用途树木的种植,并增加了土地投资和土壤肥力管理。卫星图像显示,该区域森林损失减少,而混农林业面积有所增加(Kerr、Pender和Suyanto,2008)。
保障地方权属为以相对较低的成本进行有效保护提供了巨大的机会(Ding等,2016)— 这种解决方案不仅公正而且还可以减少冲突(Tauli-Corpuz、Alcorn和Molnar,2018)。如果实施得当,可以同时促进多个可持续发展目标的实现。2可以协商土地和森林的权利,以强调那些有助于生物多样性保护的权利。然而,与保障地方权属有关的干预措施需要对政治、经济和法律背景进行仔细审查,正如粮农组织制定的《在国家粮食安全背景下对土地、渔业和森林权属进行负责任治理的自愿准则》中所强调的那样(粮农组织,2012b)。
由于许多国家在1989年通过了《土著和部落居民公约》(国际劳工组织,2017),而且几乎全部国家都批准了2007年《联合国土著居民权利宣言》(联合国,2008a),所以越来越多的国家正通过法律和宪法改革对土著居民和地方社区的土地和森林权利给予法律上的承认。其中一些国家(例如,澳大利亚、巴西、哥伦比亚、厄瓜多尔、印度、秘鲁、菲律宾、南非和美国)明确规定了在保护区内对此类权利的认可(RRI,2015)。
自由、事先和知情同意(FPIC)是与土著居民有关的一项特定权利,已得到一系列国际法律承认,包括《土著和部落居民公约》、《联合国土著居民权利宣言》和《生物多样性公约》。获得自由、事先和知情同意不仅允许土著居民在任何阶段对某个项目给予或撤消认可,而且还包括确定哪种参与、协商和决策的过程类型是恰当的。
一些国家为自愿纳入保护区的社区(和私有)土地提供支持,并给予某些利益以补偿权利上受到的限制,例如保护免受第三方侵害和政府对特许权的分配,分享旅游收入,以及其他财政或技术利益帮助;澳大利亚的土著保护区计划就是一个例子(Davies等,2013)。
许多其他国家不承认保护区的地方社区权利,但已在公有和社区拥有的土地上采用了多种共同管理的系统,因此既针对保护又针对发展的需要。社区的权利可能包括获得、使用和管理的权利。共同管理安排可以为当地社区提供一种方法,维护在习惯法下对大片连续土地拥有的使用权和管理权。然而,它们往往是高度集中的,大多数计划都没有适当地考虑当地社区的需求或将传统知识纳入管理中(RRI,2015)。尽管如此,成功的案例还是展示了共同管理系统的潜力(见案例研究10中的实例)。另一个例子是第6章在保护区的保护成效中提到的巴西亚马逊地区的采伐保护区(第117页)。
马库依拉国家公园位于哥伦比亚东北部瓜希拉(La Guajira)半岛之上,占地25000公顷(图A),是韦乌族人的神圣文化景观,是由农业、放牧和选择性森林利用形成的(Premauer和Berkes,2012)。该公园包括一个小而孤立的山脉,其山峰和山坡上有永久性的湿润森林。在这里发现的山地矮林是特有物种的绿洲,并且在哥伦比亚是唯一一个这样生态系统(UAESPNN,2005)。在建立国家公园之前,韦乌族人由于其文化禁忌和对自然的尊重而保护了许多地区和景观特征(Premauer和Berkes,2012)。1977年宣布成立国家公园之时,没有考虑土著对领域的主张,所以导致一系列的冲突。然而,多年来逐渐形成一种协作式治理和解决问题的方法,使韦乌族人和生物多样性保护得到双赢的效果(Premauer和Berkes,2012)。
1984年,韦乌族人以集体土地保有权形式(名为resguardo,一种土著保护区)被授予对其祖先领土的土地所有权。在保护区内,土著人民拥有对经济、社会和文化发展进行管理的权利。保护区的土地覆盖了哥伦比亚三分之一的国家领土,80%以上的森林地具有很高的生物多样性价值。土地所有权不能出售或没收。韦乌族人对其祖传土地的所有权是成功保护马库伊拉的关键之一。
具有参与性保护性质的“有居民的公园”政策制定于1998-2000年间,并在在全国范围内土著领土与保护区有重叠的地方实施,例如马库伊拉国家公园(Premauer和Berkes, 2015)。该政策强调对土著权利的承认、地方管理、跨文化管理措施、以管理而不是保存来实现保护(Ingwall-King和Gangur,即将出版)。
为响应“有居民的公园”政策,马库伊拉公园的公园管理一直高度尊重习惯价值和治理。例如,公园管理部门花了三年时间与当地人和合法的习惯管理当局建立关系,并了解韦乌族人的社会和政治组织以及领域管理。因此,在2006年,大多数韦乌酋长们接受了与公园进行合作(Premauer和Berkes,2015)。
此外,还采取了共同决策的程序,通过建立由54名酋长组成的理事会来商讨决定共同治理协议的文化和保护目标。会议在瓦乌地区附近举行,省却了酋长们的长途旅行,而且主要使用韦乌语,方便韦乌人自由发言(Premauer和Berkes, 2015)。
将公园作为由土著居民和地方社区或ICCA(见插文48)保护的领域或区域进行管理,使韦乌人拥有了自治权,以他们认为合适的方式行驶其习惯价值和做法,例如,从事狩猎、采集林产品、饲养牲畜和园艺。这些人类与环境的互动已支持韦乌人的生活方式数百年(Premauer和Berkes, 2012、2015)。
共同管理的安排以多种方式帮助公园和韦乌人解决了分歧:
▸ 公园支持韦乌人保护其领土,并确保其对针对公园采取的任何行动有自由自愿、事先知情的认可权。
▸ 因为公园的工作人员太少,无法控制所有入侵者的出入,所以韦乌人帮助控制和监控公园中的活动。
▸ 韦乌人和公园管理层限制进入山地矮林,这尊重了韦乌人的文化禁忌和增加了公园的保护价值。
然而,目前仍然存在一些冲突,例如旅游业。然而,以共同利益为基础的合作治理关系已取得积极成果,尤其是保护领土免受外部威胁方面,例如阻止了公园内的采矿和勘探活动。这些共同利益有助于建立信任、尊重和互惠的关系(Premauer和Berkes,2015)。
公园管理部门和韦乌人之间的合作减少了该地区的非法活动,例如偷猎禽鸟和非法采伐木材(Premauer和Berkes, 2012)。尽管缺乏系统的数据使得很难准确地评估生物多样性趋势,但从景观层面上看,自上世纪70年代以来,马库依拉的五种植被(尤其是山地矮林)的完整度一直得以保持(Premauer和Berkes,2012)
资料来源:Premauer和Berkes,2015。
在保护区之外,一些其他有效的基于区域的保护措施还承认地方权利,以便在产生积极保护成果的同时实现可持续利用。例如,纳米比亚以社区为基础的野生动物管理方法,赋予形成保护组织的社区机构以合法权利,使其能使用从自己土地上的野生动物,从中获益。在过去的二十年中,这种方法不仅创造了可观的收入,而且野生动物的数量和多样性也急速增加(NACSO,2017b)。
需要筹集资金以解决毁林的驱动因素,并更好地保护、管理和恢复森林及其生物多样性。
每年估计约需2000亿美元来支持向零毁林的牛、大豆、棕榈油以及纸浆和造纸生产方式的转变(热带森林联盟,2020),而实施《生物多样性公约》的《2011-2020年生物多样性战略计划》的成本最初估计为每年1500亿美元至4400亿美元(包括但不限于森林生物多样性)(《生物多样性公约》,2012a)。这些数字貌似庞大,但与目前每年超过7000亿美元的农业财政激励措施(经合组织,2019a)或矿石燃料补贴(2017年估计约为5.2万亿美元,约占全球GDP的6.3%)相比则显得很小(Coady等,2019)。
尽管最近对森林在保护生物多样性和减轻气候变化中的作用的关注得到了重视,但目前的资金筹措仍远远没有达到这些目标。这必须改变也可以做得到。经合组织在2019年5月举行的七国集团环境部长会议上准备的报告(经合组织,2019b)明确提出了采取行动保护生物多样性在社会经济和商业上的重要性,许多已确定的规模化生物多样性行动的机会将对森林产生积极影响。图44显示了各种可能的资金来源。
长期筹资的解决方案越来越依赖于私营部门和可以实现自我维持的融资手段,例如环境基金。许多创新方法带来了希望。零土地退化基金的公私合作伙伴关系模型是由防治荒漠化公约全球机制(《荒漠化公约》,日期不详)推出的,它通过恢复土地的同时从已恢复土地的可持续生产中为投资者获取收入来支持向零土地退化的转变,而国际林业研究中心提议的景观基金计划则按照绿色债券的模式发行森林恢复债券(粮农组织和《荒漠化公约》全球机制,2015)。新的金融产品和行业投资通过企业社会责任和慈善事业成为传统资金有益的补充。尽管资金来源相对较少,但存在多种多样可以为保护森林和生物多样性筹集资金的手段(表7)。
利用私人融资杠杆。公有部门通过强有力的环境法规和积极的激励机制,在利用私人资金进行保护方面发挥着关键作用。然而即便这些措施到位,新的可持续土地利用模式也常常被视为风险投资,特别是如果要在发展中国家实施。因此,私有部门往往要求诸如政府或多边金融机构之类的合作伙伴通过提供次级债务、第一损失担保和其他信用增强结构来降低投资风险。这样做可以获得大量的私人投资。这方面的例子包括热带景观融资机制(一个由环境署、世界农林业中心、法国巴黎银行和亚洲债务管理有限公司达成的合作伙伴关系),筹集了近10亿美元的债券为可持续商品生产、加工和贸易提供融资,以及Agri3基金(由环境署、荷兰合作银行和IDH可持续贸易计划达成的合作伙伴关系),近10亿美元资金用于零毁林性商品的生产。
另一个例子是美国的栖息地保护银行,它将强有力的立法和有利的体制机制结合起来,鼓励私营部门参与保护濒危物种。保护银行是一种补偿机制,有利于促进人们遵守1973年美国《濒危物种法》(美国政府, 1973)。通过该工具,以提供永久性生境保护为目的进行土地管理的私有土地所有者,可以转让征得美国林务局批准的生态功能和服务积分。项目和开发商购买这些积分来抵消其对环境造成的不利影响。自2003年颁布国家保护银行准则以来至2016年止,保护银行的数量已达137个,该计划下的土地面积增加了288%(Poudel、Zhang和Simon,2019)。
虽然许多国家都提供了有关保护区内外森林管理成本的信息,但较少有人尝试评估进行恢复工作的成本和收益,而且往往由于缺乏基准数据和进行追踪、理解和分享结果和经验教训的统一的框架,那些做了的评估也没有得到很好的记录。例如,生态系统和生物多样性经济学倡议计划研究了2万多个恢复案例,发现只有96个包含有用的成本数据(经合组织,2019b)。缺乏信息阻碍了公共和私人部门对恢复活动的进一步投资,损害了实现恢复目标的机会,妨害了它们为实现可持续发展、减缓和适应气候变化的全球目标、保护和可持续利用生物多样性做出贡献。生态系统恢复经济学倡议计划(第5章插文43)旨在填补这一信息空白。一般而言,迹象表明收益通常会超过成本。例如,最近的一项分析估计,在全球范围内恢复3.5亿公顷的退化森林地区,每投资1美元可产生7-30美元的效益(Verdone和Seidl,2017)。
生态系统服务付费。基于结果的减少毁林和森林退化所致碳排放的付费是目前就森林提供的生态系统服务提供支付的最大全球计划,在减缓毁林速度及生物多样性的丧失方面已经产生了重大的积极影响。针对与水有关的森林生态系统服务进行的付费在许多国家很普遍,欧洲经委会和粮农组织(2018)列出了北美101个和欧盟国家70个正在实施的计划。
生态系统服务付费计划还用于奖励和规范某些更直接地支持私有土地上生物多样性保护的措施。此类计划已成功用于保护生物多样性丰富的地区,包括野生动植物种群的重要迁徙和扩散地区。然而,在土地权属不明确或不安全的地方,这些计划可能难以实施,因为很难将环境服务划归提供者(粮农组织,2016c)。这对于非洲农村的生态系统服务付费来说,是一个重大问题,那里90%的土地属于传统土地权制度,并没有任何正式的所有权(Blomley,2013)。在某些国家,非政府组织协助社区获得传统土地权证书,以帮助他们克服这一限制。例如,在坦桑尼亚联合共和国的西曼吉罗平原,基层组织乌贾马社区资源小组已通过获得传统土地占有权证书,帮助38个牧民和狩猎人社区在62万公顷土地上获得了有保障的使用权,使他们能够为超过100万公顷的土地制定土地使用计划(Nelson和Sinandei,2018)。在一些社区和旅游服务经营者之间建立的生态系统服务付费合同,帮助其获得社区的支持来通过传统的土地使用规则维持野生动物的扩散区,同时每年向社区支付的费用旨在防止将来转换为农业用地(Sachedina和Nelson,2012)。这种方法还有助于减少冲突,并为该地区一些最为边缘化的社区提供民生保障。
哥斯达黎加通过允许缺乏正式土地所有权的所有者选择提供一些证据来证明其拥有权的方式(FONAFIFO、CONAFOR和环境部,2012)或借用未来付款来支付办理土地使用权所需费用的方式(粮农组织,2016c),解决了生态系统服务付费中森林权属得不到保障的问题。表8列出了十个最大的国家生态系统服务付费计划。
保护地役权。保护地役权是“一项自愿的法律协议,它为了保护该土地的生态价值而对其使用进行永久限制”(NCED,2019)。与生态系统服务付费一样,保护地役权经常被用来激励对土地具有明确和有保障的所有权的私有土地所有者对他们的土地进行保护,包括管理国家公园附近的大片公共区域(粮农组织,2016c)。在这种情况下,土地所有者被要求放弃某些权利以获取某些特定利益,通常是经济激励机制(例如在欧洲和美国通常是减少税收)。在坦桑尼亚联合共和国北部,一些社区与私营部门之间订立的保护地役权协议为社区提供年度付款和就业机会,对其放弃进一步的农业扩张做出补偿(Sachedina和Nelson,2012)。
用自然抵偿债务。美国《热带森林保护法》(TFCA)于1998年颁布并于2019年重新授权(TNC,2019),为有资格的发展中国家提供了多种选择,以减轻其对美国政府的某些官方债务,同时以当地货币筹集资金来支持热带森林保护活动。美国国际开发署(2017)报告称,自1998年以来,已与14个国家缔结了20项美国《热带森林保护法》的用自然抵偿债务协议:孟加拉国、伯利兹、博茨瓦纳、巴西、哥伦比亚、哥斯达黎加(两项协议)、萨尔瓦多、危地马拉、印度尼西亚(三项协议)、牙买加、巴拿马(两项协议)、巴拉圭、秘鲁(两项协议)和菲律宾(两项协议)。这些协议涉及政府资金2.33亿美元,以及来自非政府组织(大自然保护协会、保护国际基金会和世界自然基金会)的2250万美元。另外又从利息收入、资本收益、受赠方成本分摊、以及其他捐助方对项目的共同供资等多种来源筹措了8300万美元,总金额超过3.3亿美元。
许多国家在非政府组织(如坦桑尼亚联合共和国,俄罗斯联邦和世界自然基金会[WWF,2018])的支持下,也正在与私有基金会就以债务换自然的协议进行谈判。此类计划为非洲减免债务和进行自然投资提供了非常积极有效的机会,非洲是近年来外债急剧增加的大陆。
在国家层面,需要制定更好的指标来监测自然资本的趋势以及森林对人类的效益,以帮助决策者确保在制定发展计划时将不同土地利用方案之间的权衡和协同作用考虑在内。
将国民核算体系扩展到包括衡量环境及其与经济之间关系的核算框架是一个需要满足的长期要求(如Repetto,1992)。这一要求于1992年在《21世纪议程》中被首次提出,采用环境经济核算体系(SEEA)核心框架则是向前迈出的重要一步。该框架作为一个国际统计标准,将环境资源及其对经济的贡献和它们的固碳储碳功能以实物和货币的形式加以核算(联合国等,2014a)。在环境经济核算体系中,森林作为一项特定的自然资本得到了特别关注(如世界银行,2017)。环境经济核算体系实验性生态系统核算旨在进一步扩展环境经济核算体系,提供关于自然资本的基于生态系统的指标(联合国等,2014b)。
环境经济核算体系通过提供一个一致的框架来组织有关自然资本的信息,并将其与国民核算体系联系起来,是将森林、森林生态系统服务和森林生物多样性的惠益纳入经济规划的重要工具(参见如Banerjee等,2016)。目前约有40个国家正在使用环境经济核算体系来辅助其与生物多样性相关的决策和管理(Ruijs和Vardon,2019)。许多国家对批准涉及转变公有森林的项目之前进行的环境影响评估也有详细要求。
虽然经常在国家范围内考虑政策和法律框架,但区域框架和合作在加强治理和在更大范围内采取行动方面会非常有效(见插文54)。例如,欧盟呼吁各国之间采取更加协调一致的行动,于1979年通过了《欧盟野生鸟类保护指令》,并于1992年通过了《栖息地保护指令》,以应对急剧发生的物种灭绝、栖息地破坏和生态系统退化,实现这些目标及其对《生物多样性公约》的承诺。栖息地保护指令的核心是创建“Natura 2000”,这是一个欧盟范围内的生态网络,包括在野生鸟类保护指令(特殊保护区)和栖息地保护指令(特殊保护区)保护之下的所有区域。该网络涉及28个欧盟国家,覆盖了18%的欧盟陆地面积和9.5%的海洋,包括一些受到严格保护的自然保护区,但大部分为私有土地(欧盟委员会,2019b)。森林生态系统约占该网络陆地面积的50%。2012年启动的《Natura 2000生物地理进程》促进了成员国之间的协调行动以及各种政府和非政府利益相关者之间的合作,实现有效实施、管理、监测、筹资和报告以及在整个网络中强制遵守法规。尽管面临挑战和实施缓慢,特别是在海洋栖息地,但Natura 2000已被证明在解决整个欧盟领土上关键栖息地的丧失、破碎和退化方面取得了成功(Medaglia、Phillips和Perron-Welch,2014)。
▸ 中非森林委员会(COMIFAC)(COMIFAC,2020)是一个政府间组织,负责协调与中非森林保护和可持续管理相关活动。活动由次区域融合计划进行指导。该计划第二版(2015-2025年)有六个优先事项,其中之一就是“保护和可持续利用生物多样性”。
▸ 于2007年启动的绿色长城(Great Green Wall,2019a)计划有着宏伟的目标,要在非洲萨赫勒地区的整个宽度上植树长8000公里的树木。近年来,该倡议已发展到更广义上的促进可持续地利用和恢复措施的综合应用。绿色长城被认为是非洲应对气候变化、生物多样性丧失、土地退化、荒漠化和干旱的旗舰型举措,旨在改善民生并提高粮食安全和抵御力。它强调了生物多样性对人类的重要性。
▸ 非洲可持续森林管理框架(2020-2030年)旨在指导非洲联盟和非洲区域经济共同体成员国在与森林有关的优先事项上实现非洲联盟(非盟)《2063年议程》的目标(非洲联盟,日期不详)和《联合国2030年议程》。优先事项包括提高森林、市场、加工和贸易、能力发展和知识管理的价值;促进可持续森林管理的支持性政治和体制框架,促进退化森林和景观的恢复;并加强伙伴关系和资源筹措。
▸ 欧洲森林(欧洲森林保护部长级会议)是泛欧自愿的高层政治进程,旨在就欧洲森林政策进行对话与合作。欧洲森林为其47个缔约体(46个欧洲国家和欧盟)制定了有关如何保护和可持续管理其森林的通用策略和指南。
▸ 2019年12月,欧盟理事会通过了行动框架,以加强欧盟保护和恢复世界森林的行动(欧盟委员会,2019a)。该框架有五个优先重点,以实现保护和可持续管理全球具有生物多样性的森林。它着重强调了这些行动将对实现与森林有关的多边环境协定以及扭转毁林趋势做出的贡献。
▸ 亚马逊合作条约组织致力于促进亚马逊流域各国森林资源的保护和可持续利用,与森林有关的生物多样性间接得益于该组织的某些活动。玻利维亚多民族国、巴西、哥伦比亚、厄瓜多尔、圭亚那、秘鲁和苏里南于2019年签署了《亚马逊莱蒂西亚条约》(Leticia Pact,2019),确认了为森林和生物多样性评估进行区域合作以及协调行动,防止毁林和森林退化,打击非法活动,预防火灾和其他灾难,以及恢复和再造林的倡议。
生物多样性的丧失或保护往往是人类行为的结果。因此,可持续的自然资源管理需要倾向保护的人类价值观、态度和行为,并将人类视为自然的一部分,而与自然又与人类息息相关(Saunders、Brook和Meyers,2006;St.John、Edwards-Jones和Jones,2010;Verissimo,2013)。
不幸的是,尽管公众越来越意识到环境问题的重要性,但大多数人并未积极参与到支持更可持续未来的行动中(Bickford等,2012)。有效的保护性干预措施需要激励人们改变行为,这需要具体了解对自然的态度是如何转化为行动的,以及人类行为如何产生积极的生物多样性成果(Verissimo,2013)。
增强环境素养。环境素养可以为实现生物多样性保护和可持续森林管理提供基础,并可以通过教育和知情沟通得到促进(McKeown,2002)。一种新的教育可持续性的方法必须强调批判性思维、综合原则以及利用获得的技能将知识转化为行动(Schelley等,2012)。环境素养通常通过与自然亲密接触的经验来建立,包括参与以生态为重点的户外活动并参与适应性管理(Saunders、Brook和Meyers,2006;Bickford等,2012)。森林学校让孩子们从小就开始欣赏和了解自然(O’Brien和Murray,2007)。
提高环境素养的一种方法是通过公民科学计划,让公众参与收集数据或生态研究,例如通过让居住在保护区附近或受到入侵物种威胁的社区居民参与计划(插文55)。科学家可以与基层民众组织、土著居民和当地社区合作,设计旨在传播有关当地生态系统的知识、增进对保护问题的了解、赋权于当地利益相关者以作出知情决定的一系列计划(Bickford等,2012)。
在全球范围内,许多森林不断遭受严重的入侵物种爆发,对环境和社会文化可能产生巨大影响。随着全球贸易和旅行的逐渐增多,森林受入侵物种的威胁日益增加,而气候变化的影响又加剧了这些威胁。管理入侵物种并避免引入已知具有潜在可能成为入侵物种的动植物,这需要许多各方在国家、区域和全球范围内共同努力。
《2025年新西兰生物安全计划》旨在推动变革,使该国每个公民、企业和组织都成为生物安全风险管理者。该计划强调了包容性和参与性,这使得国家生物安全系统更具适应性和面向未来,对保护本国免受病虫害的侵扰至关重要。
英国正在考虑采用相同的战略。该国环境审计委员会的一份报告援引新西兰在2025年前培训15万人的生物安全性计划(新西兰生物安全,2018),呼吁英国采用鼓励公众参与的方法来遏止外来物种入侵,后者已构成对英国自然环境的五大威胁之一。外来入侵物种不仅挑战了该国一些最稀有物种的生存,而且还破坏了自然生态系统,每年造成的经济损失估计为17亿英镑(超过22亿美元)。例如,橡树游蛾(Thaumetopoea processionea)可以破坏橡树树皮使其裸露,也对人体健康造成危害,而由真菌(Hymenoscyphus fraxineus)引起的白腊树枯梢病,造成的威胁可能使该国本地白蜡树在一个世纪之内损失一半,造成150亿英镑(约200亿美元)的经济损失。该委员会希望教会130万人如何发现入侵物种的爆发,并呼吁建立一支专门的边境部队,以改善国家边界处的生物安全。
资料来源:RNZ(2019)。
分享取得成效的成功保护案例,可以通过演示可以得到什么以及如何得到来增强人们的权能并促进行动(Nadkarni,2004;Saunders、Brook和Meyers,2006;Garnett和Lindenmayer,2011)(见插文56中的实例)。传统上,关于保护的故事是通过媒体传达给公众的,但是这种交流往往缺乏细节和准确性(Nadkarni,2004)。科学家、研究人员、宗教领袖和保护主义者除了可以参与公共领域的媒体活动以外,还可以通过许多其他方式与公众进行交流,例如,名人和有影响力的人可以通过担任知识大使来吸引更多的受众,尤其是年轻一代(Galetti和Costa-Pereira,2017)(见插文57中的实例)。根据听众的背景,通过故事和隐喻与他们进行交流,并使信息与他们的意识形态或精神和宗教信仰保持一致,这会有助于信息的传达和交流。与公众沟通是互惠互利的:公众增强了关于环境和可持续性问题的意识,从业者和科学界获得新见解,有助于其形成行动、研究问题、政策和决策支持工具。
世界树木城市是一项由粮农组织和美国植树节基金会推动的国际活动,以表彰鼓励致力于维护、可持续管理和欣赏其城市森林和树木的城镇。要获得认可,城镇必须符合五个核心标准:
▸ 树立权威:市领导以社区的书面声明将市政范围内树木的护理责任委托给工作人员、市政府或一组市民(树木委员会)。
▸ 制定规则:社区采用政策、最佳做法或行业标准来管理城市树木和森林,说明如何执行工作,在何处以及何时使用规则,以及对违规行为的处罚。
▸ 了解拥有的资源:社区会更新对本地树木资源的评估,从而可以为种植、护理和清除城市树木制定有效的长期计划。
▸ 分配资源:社区有专门的年度预算,用于例行实施树木管理计划。
▸ 庆祝成就:社区每年举行一次树木庆祝活动,以提高居民的意识并表彰执行城市树木计划的市民和工作人员。
由联合国环境规划署和Futerra公司于2016年创建的为生命呐喊(Wild for Life)运动(https://wildfor.life),旨在提高全球意识并动员千百万人,特别是年轻人,来支持对濒危物种的保护并倡导制止非法野生动物贸易。
“为生命呐喊”运动目的在于将保护工作拟人化:为这些濒临灭绝的物种起一个名字和一张面孔。为了实现这种拟人化联系,该活动设有一个在线个性测验,该测验根据特征和行为,为每个测验者分配一个物种。然后邀请用户拍摄#WildforLife自拍照,将用户的图像和该物种结合在一起,并在社交媒体上分享照片。
该运动吸引了来自联合国和政府机构、慈善机构和媒体的25个合作伙伴,其中包括世界银行,国际刑警组织,简·古道尔研究所和Rovio Entertainment公司(创作了《愤怒的小鸟》系列)。同时还为超过35位名人、有影响力的人物和亲善大使提供了测试和匹配,并得到了他们的支持,将他们的面孔和名字都与某种濒临灭绝的物种联系起来。
这场运动已经涉及超过10亿人,其中包括中国主要目标市场的近3.3亿人。更重要的是,该活动中的几个物种在《濒危野生动植物种国际贸易公约》的进程中获得了全球的更大支持,中国已宣布到2017年底全面禁止象牙商业贸易。
该运动的成功可以归因于:
▸ 以积极的榜样为先导,专注于人们对自然和濒危物种的热爱和与它们之间的联系;
▸ 以可行和可解决的方式陈述问题,表明这是一场在大家参与下可以赢得的战斗;以及
▸ 新鲜、引人入胜和英勇的身份形象,在视觉上脱颖而出。
在不断变化的环境中进行生物多样性规划和决策取决于准确的知识和信息。无论是动物还是植物,我们在种群、物种和遗传水平上对森林生物多样性的知识仍然有限。然而,为了弥补这一领域的差距,很多工作正在进行。
许多国际进程和可持续发展目标都需要准确、有效、成本效益高的森林信息监测和报告,它们也为促进改善森林管理和可持续发展提供了基础。有了新的工具(插文58),以前不具备所需数据收集能力的国家现在也可以用最少的资源和培训获得和分析内容广泛的信息(参见插文59中的实例)。
Open Foris(www.openforis.org)是粮农组织开发的一套创新且易于使用的森林监测平台和工具,使世界各地的用户能够自主收集和分析信息,并将这些信息报告给国际社会。这些工具易于使用和直观,不需要任何先前技能,并且是免费和开放源代码。
Open Foris通过降低成本、消除收集和分析数据的障碍、改善许多国家政府的森林监测,在打击毁林方面发挥了关键作用。
在Open Foris中,用于地球观测、数据访问、处理和土地监测分析(SEPAL)的系统是一个大数据处理平台,结合了超级计算能力、开源地理空间数据处理软件和现代空间数据基础设施(例如谷歌公司的Earth Engine),使世界各地的研究人员和技术人员都可以获取和分析卫星图像,并为本地决策提供相关依据。
巴布亚新几内亚是著名的生物特有病和物种多样化中心。尽管其生物多样性程度、规模和丰富度极高,但从科学的角度来看,人们对其森林知之甚少。为了增进对国家森林生物多样性的了解,政府扩大了国家森林资源清查的范围,除树木生物量、树木物种多样性和土壤的化学物理特性外,还包括了树木以外的植物、鸟类和昆虫(飞蛾、果蝇和蚂蚁)。
由于难以评估,所以国家森林资源清查极少包含生物多样性的详细信息。巴布亚新几内亚正在使用粮农组织开发的用于森林和土地利用监测的Open Foris来收集、记录和分析此信息(见插文58),其中包括“Collect Earth”,该工具将谷歌地球(Google Earth)的数据与必应地图(Bing Maps)和谷歌地球引擎(Google Earth Engine)相结合。人们仅需经过一两天的培训即可使用这些工具,这使本国研究人员能够进行生物多样性相关的研究,对该国制定恰当的森林管理计划和政策十分必要。九名学生已经完成了与国家森林资源清查有关的研究。
为巴布亚新几内亚国家森林清查收集信息。
©粮农组织/Hitofumi Abe遥感数据(见插文60)与地面数据相结合,对于跟踪监测地球自然资源的状态和趋势具有不可估量的价值。正如本卷中提出的许多研究所示,卫星图像和工具方面的最新技术发展显著提高了收集和分析大量数据的能力。
使用来自卫星传感器的数据来监测地球森林生物多样性已经进行了多年,复杂性和规模不一而同。一些生物多样性的指标是直接获得的(例如,仅用卫星获得的信息即可得出),而大多数指标是通过利用卫星图像中所观测到的现象推算出生物多样性状况和地面生物多样性变化相结合这样间接的方法进行的。尽管科学文献中有一些例子已使用遥感技术对图像中的动物进行识别和计数,但本节主要侧重于利用卫星图像对植被进行分类,因为这直接与森林生物多样性有关,也可以作为其他生境生物多样性状况的大致估计。
从最基本的角度讲,对地观测卫星对于监测土地覆盖的状态和趋势(例如土地表面的生物物理特性)非常有用。自1970年代初以来,专门为测量和监测土地覆盖而发射的卫星一直在提供数据,使我们可以对树木覆盖程度、分布和动态进行定性。这些数据可用于估计任何区域树木覆盖状况随时间的变化。因此,它们可以用来描述影响生物多样性的几个最重要的因素,包括是否存在树木覆盖、树木覆盖总面积(更多的区域通常意味着更多的生物多样性)和树木覆盖的变化(因为毁林往往导致生物多样性减少,而再造林可以增加多样性)。
根据卫星数据估算森林基于分类学的生物多样性则更为复杂,通常需要将卫星观测与现场观测联系起来。在大多数情况下,在地球表面测得的反射率会转换为一组光谱指数。每个光谱指数都以某种方式与植被状况相关,例如在水分含量、光合作用和冠层覆盖率方面。这些指标有助于对植物功能、健康、活力和其他关键参数做出判断。然后,这些参数可以与基于地面的对物种集合的观察相结合。一旦建立了这种关系,就可以在从国家到区域乃至全球的较大空间尺度上绘制植被状况的地图。
通过遥感绘制物种分布地图有两种形式:间接和直接。间接法绘制物种分布图可以通过融入其他遥感数据(例如来自天气和气候观测传感器的数据)以及例如海拔和地形(也可以从遥感数据中得出)等其他可用数据来进行改善。结合来自多方的数据,可以预测在何时何地可以满足特定植物的生长需求,并由此模拟大面积上植物物种分布范围。直接法绘制物种分布图可以通过从卫星图像观察和检测植物性状,直接对物种进行制图 — 例如,通过测量植被高度(例如,将矮小物种与高大物种区分开),跟踪有叶/无叶的状态(例如,由此区分常绿植物和落叶乔木)和观察大面积开花事件(例如监测热带或温带森林中的物种)。最近,高光谱遥感(例如,针对数百种特定光波长的遥感)使得仅根据每种物种的独特光谱特征就可以检测森林中的单个树种。
最后,卫星可以测量对大规模生态系统功能重要的参数,从而洞察对森林生物多样性具有重大影响的大面积变化。例如,卫星可以检测树木死亡、新增物种、降雨模式和其他对定性生物多样性至关重要的变量,并且该信息可以用于测量、监测和预测生态系统功能的变化,从而预测生物多样性。
下一代卫星有望在提供与森林生物多样性直接相关的指标方面更加有用,包括对树高、树冠特征和植物功能的直接和精细观测。这些先进技术,再加上越来越多更好的林间数据,以及无人驾驶飞行器(无人机)的使用,将会持续增强我们发现和监测生物多样性的能力。
进一步取得进展的一个重要领域是制定和应用监测生物多样性的指标。实例包括第2章中的破碎化状况研究(森林完整度和破碎度)和仅存于森林的物种指数(测量森林脊椎动物种群的趋势)以及生物多样性意义和完整性研究(评估森林生物多样性)。第3章中的其他示例在插文61和62中给出。
面对快速的城市化进程(联合国,2008b),需要将生物多样性保护扩展到城市,那里有可能具有丰富的生物多样性(《生物多样性公约》,2012b)。2008年,在《生物多样性公约》缔约方大会第九届会议上,新加坡表示愿意牵头制定生物多样性指数,以追踪城市中生物多样性保护举措的有效性。
由《生物多样性公约》秘书处(SI)、新加坡和全球地方生物多样性行动伙伴关系制定的《新加坡城市生物多样性指数》由三个部分组成。它衡量了在城市或评估区域中可以发现的本地生物多样性;其提供的生态系统服务;以及用于治理和管理生物多样性的措施(表A)。一本用户手册(Chan等,2014)提供了有关如何使用的详细信息。
截至2018年,六大洲的30多个城市已经使用了新加坡城市生物多样性指数(《生物多样性公约》,2018c)(图A)。
该指数具有多种用途,例如用于制定城市/区域/项目的总体规划;用于决策和决定资源分配的优先次序;用于补充其他环境可持续性或绩效指标;并为制定当地生物多样性战略的准则做出贡献。
在新加坡曼岱路沿线多层种植各种树木,模仿低地热带雨林结构,从而降低了周围环境温度;为包括灵长类动物、小型哺乳动物、鸟类和蝴蝶等动物提供了栖息地和生态联系;固存和储存碳;将人们与自然联系起来,从而改善身心健康。
©Lena Chan利用生物多样性指标进行栖息地评估是一种以科学为基础的经济有效的方式,可以衡量森林生态系统的健康状况,为制定维持生物多样性和通过可持续管理确保生态系统服务的决策提供信息。由于水量和水质(包括沉积物含量、化学物质含量和温度)对树木覆盖率和森林管理的变化敏感,因此淡水生物指标可以很好地反映河岸生态系统健康随时间的变化。
河岸栖息地评估工具通常着眼于与生物多样性有关的许多特征,包括动植物物种的存在、不存在和/或其数量,水质,植被类型,河岸植被结构以及河道和河岸变化。此类工具现已被用作公民科学监测计划的一部分(Gurnell等,2019)。这样的例子包括英国的河流栖息地调查、的由世界自然基金会瑞典分会和瑞典森林所有者协会开发的“”。
瑞典的小型林场主使用“蓝色目标工具”来确定保护内陆水域(特别是小河)所需的河岸缓冲区的宽度。该工具由一张双面纸构成,包括一系列基于科学参数的是非选择题的快速评估调查问卷和一个评分系统(Henriksen,2018)。工具中包含的保护价值包括特殊的栖息地/物种,水体和河岸带;对人类的影响,包括对水道的改造;土壤敏感性,包括地形和侵蚀风险;增值,例如娱乐,食品生产,文化价值和恢复。根据调查结果,将水体根据保护需求归为以下四类之一:
▸ 可以在离水很近的地方进行林业活动的;
▸ 需要较大河岸缓冲区的;
▸ 需要采取特殊保护行动的,例如消除迁徙障碍或恢复河岸缓冲区、栖息地或水文形态条件;以及
需要尽可能宽阔河岸缓冲区的,在这些地区营林操作需要认真考虑对水的影响。
由于其有效性和简便性,蓝色目标工具已针对其他国家(包括芬兰、拉脱维亚、立陶宛和波兰)进行了微调(Eriksson等,2018),目前正与圣保罗大学和巴西ABC联邦大学合作,为在巴西使用进行相应调整(Taniwaki等,2018)。
如本报告所示,森林是高度多样化的栖息地,为世界上绝大多数的陆地生物多样性提供庇护之所。森林生态系统、物种和遗传物质为地球上的生命奠定了基础。
人们与森林生物多样性的关系在区域、国家和生态区之间,从农村到城市都各不相同;然而,大多数人类社会至少与森林及其生物多样性具有某种相互作用。数十亿人依赖森林维持生计,获得粮食安全和福祉。据估计有24亿人使用木质能源来烹煮食物。森林和树木缓解气候变化、调节水量、提供遮荫、防风林和饲料,以及为许多传粉媒介提供栖息地,这些作用对可持续粮食生产至关重要。
在全景方法框架之下进行对森林和树木的保护和可持续使用,结合由完整森林至种植林再至混农林业系统、农田和退化土地上树木的全面连续性,是确保世界生物多样性、粮食安全和世界人民民生福祉的关键。因此,最重要的是将生物多样性保护纳入森林管理主流,并将本版报告中列举的许多产生积极成果的实例扩展到更大规模。
然而,仅仅这些还是不够的。根据编制本报告所收集的信息,显而易见的是,大多数与森林生物多样性的相关目标和指标尚未实现,而相关的可持续发展目标现在也未处于可以在2030年前顺利实现的轨道之上。同样明显的是,延续生物多样性和生态系统目前的不利趋势将会妨碍实现可持续发展目标的进展。
鉴于农业扩张是毁林的主要驱动力,我们生产和消费食物的方式需要开展颠覆性变革。我们必须摆脱目前对粮食需求导致不恰当的农业模式的情况,这些模式促使森林大规模地转变为农业生产同时丧失了与森林有关的生物多样性。采取混农林业和可持续生产方式,恢复退化的农业用地的生产力,采用来自可持续粮食体系的更健康的饮食,减少粮食损失和浪费,均迫切需要在更大规模上采取行动。农业企业必须履行其对零毁林商品链的承诺,而尚未做出零毁林承诺的企业也应这样做。商品投资者应采用对环境和社会负责的商业模式。在许多情况下,这些行动将需要调整当前的政策(特别是财政政策)和监管框架。
可喜的是,森林作为一种应对许多可持续发展挑战的基于自然的解决方案这一作用日益得到认可,这体现在政治意愿得到强化并作出一系列减少毁林速度和恢复退化森林生态系统的承诺之上。我们必须把握这一机会,采取果断有力的行动来预防、制止和扭转森林及其生物多样性的丧失,以造福今世后代。
ADB. 2016. Illicit trade in natural resources in Africa — A forthcoming report from the African Natural Resources Center. Abidjan. [also available at https://www.afdb.org/fileadmin/uploads/afdb/Documents/Events/IFF/Documents_IFF/ANRC_ILLICIT_TRADE_IN_NATURAL_RESOURCES.pdf].
AFR100. n.d. Home [online]. Midrand, South Africa. [Cited 18 December 2019]. https://afr100.org/
African Union. n.d. Agenda 2063: The Africa we want. In: African Union [online]. Addis Ababa. [Cited 13 January 2020]. https://au.int/en/agenda2063/overview
Agrawal, A., Chhatre, A., & Hardin, R. 2008. Changing governance of the world’s forests. Science, 320(5882): 1460–1462.
Aguilar, R., Quesada, M., Ashworth, L., Herrerias-Diego, Y. & Lobo, J. 2008. Genetic consequences of habitat fragmentation in plant populations: susceptible signals in plant traits and methodological approaches. Molecular Ecology, 17: 5177–5188.
Ahenkan, A. & Boon, E. 2011. Improving nutrition and health through non-timber forest products in Ghana. Journal of Health, Population and Nutrition, 29(2): 141–148.
Alix-Garcia, J., Sims, K.R. & Yañez-Pagans, P. 2015. Only one tree from each seed? Environmental effectiveness and poverty alleviation in Mexico’s payments for Ecosystem Services Program. American Economic Journal: Economic Policy, 7(4):1–40.
Alix-Garcia, J., McIntosh, C., Sims, K., & Welch, J. 2013. The ecological footprint of poverty alleviation: Evidence from Mexico’s Oportunidades Program. The Review of Economics and Statistics, 95(2): 417–435.
Alkire, S. & Santos, M.E. 2014. Measuring acute poverty in the developing world: robustness and scope of the multidimensional poverty index. World Development, 59: 251–274.
Andam, K.S., Ferraro, P.J., Pfaff, A., Sanchez-Azofeifa, G.A. & Robalino, J.A. 2008. Measuring the effectiveness of protected area networks in reducing deforestation. PNAS, 105(42): 16089–16094.
Angelsen, A., Jagger, P., Babigumira, R., Belcher, B., Hogarth, N.J., Bauch, S., Börner, J., Smith-Hall, C. & Wunder, S. 2014. Environmental income and rural livelihoods: a global-comparative analysis. World Development, 64: S12–S28. [online]. [Cited 3 January 2020]. https://doi.org/10.1016/j.worlddev.2014.03.006
Anup, K.C. 2017. Community forestry management and its role in biodiversity conservation in Nepal. In G.A. Lameed, ed. Global exposition of wildlife management [online]. [Cited 3 January 2020]. https://www.intechopen.com/books/global-exposition-of-wildlife-management/community-forestry-management-and-its-role-in-biodiversity-conservation-in-nepal
Azevedo, A.A., Rajão, R., Costa, M.A., Stabile, M.C.C., Macedo, M.N., Dos Reis, T.N.P., Alencar, A., Soares-Filho, B.S. & Pacheco, R. 2017. Limits of Brazil’s Forest Code as a means to end illegal deforestation. PNAS, 114(29): 7653–7658.
Balmford, A., Green, J.M., Anderson, M., Beresford, J., Huang, C., Naidoo, R., Walpole, M. & Manica, A. 2015. Walk on the wild side: estimating the global magnitude of visits to protected areas. PLOS Biology, 13(2): p.e1002074 [online]. [Cited 3 January 2020]. https://doi.org/10.1371/journal.pbio.1002074
Banerjee, O., Cicowiez, M., Horridge, M., & Vargas, R. 2016. A Conceptual Framework for Integrated Economic–Environmental Modeling. Journal of Environment and Development, 25(3): 276–305. [also available at doi: 10.1177/1070496516658753]
Barlow, J., Gardner, T.A., Araujo, I.S., Ávila-Pires, T.C., Bonaldo, A.B., Costa, J.E., Esposito, M.C. et al. 2007. Quantifying the biodiversity value of tropical primary, secondary, and plantation forests. PNAS, 104: 18555–18560.
Barros, F.M., Peres, C.A., Pizo, M.A. & Ribeiro, M.C. 2019. Divergent flows of avian-mediated ecosystem services across forest-matrix interfaces in human-modified landscapes. Landscape Ecology, 35(4): 879 [online]. [Cited 3 January 2020]. https://doi.org/10.1007/s10980-019-00812-z
Bastin, J.-F., Finegold, Y., Garcia, C., Mollicone, D., Rezende, M., Routh, D., Zohner, C.M. & Crowther, T.W. 2019. The global tree restoration potential. Science, 365(6448): 76–79.
Baynham-Herd, Z., Amano, T., Sutherland, W.J. & Donald, P.F. 2018. Governance explains variation in national responses to the biodiversity crisis. Environmental Conservation, 45(4): 407–418.
Beatty, C.R., Cox, N.A. & Kuzee, M.E. 2018. Biodiversity guidelines for forest landscape restoration opportunities assessments. 1st edition. Gland, Switzerland, IUCN.
Beck, H. 2008. Tropical ecology. In Jørgensen, S.E. & Fath, B.D. eds. General ecology: Encyclopedia of ecology, pp. 3616–3624. Elsevier, Oxford, UK.
Beech, E., Rivers, M., Oldfield, S. & Smith, P. 2017. GlobalTreeSearch: the first complete global database of tree species and country distributions. Journal of Sustainable Forestry, 36(5): 454–489.
Bello, C., Galetti, M., Pizo, M.A., Magnago, L.F.S., Rocha, M.F., Lima, R.A.F., Peres, C.A., Ovaskainen, O. & Jordano, P. 2015. Defaunation affects carbon storage in tropical forests. Science Advances, 1(11): e1501105 [online]. [Cited 3 January 2020]. https://doi.org/10.1126/sciadv.1501105
Belluco, S., Halloran, A. & Ricci, A. 2017. New protein sources and food legislation: the case of edible insects and EU law. Food Security, 9(4): 803–814.
Bengston, D.N., Butler, B.J. & Asah, S.T. 2008. Values and motivations of private forest owners in the United States: a framework based on open-ended responses in the national woodland owner survey. In D.B. Klenosky & C.L. Fisher, eds. Proceedings of the 2008 Northeastern Recreation Research Symposium, pp. 60–66. General Technical Report NRS-P-42. Newtown Square, Pennsylvania, USA, USDA Forest Service, Northern Research Station. [also available at https://www.nrs.fs.fed.us/pubs/gtr/gtr-p-42papers/09bengston-p-42.pdf].
Benítez-López, A., Alkemade, J.R.M., Schipper, A.M., Ingram, D.J., Verweij, P.A., Eikelboom, J. & Huijbregts, M. 2017. The impact of hunting on tropical mammal and bird populations. Science, 356(6334): 180–183.
Bennett, G. 2004. Integrating biodiversity conservation and sustainable use: lessons learned from ecological networks. Gland, Switzerland, IUCN.
Bennett, G. & Mulongoy, K.J. 2006. Review of Experience with Ecological Networks, Corridors and Buffer Zones. Technical Series No. 23. Secretariat of the Convention on Biological Diversity, Montreal, Canada.
Bentz, B.J., Régnière, J., Fettig, C.J., Hansen, E.M., Hayes, J.L., Hicke, J.A., Kelsey, R.G., Negrón, J.F. & Seybold, S.J. 2010. Climate change and bark beetles of the Western United States and Canada: Direct and indirect effects, BioScience, 60(8): 602–613.
Berman, M., Jonides, J. & Kaplan, S. 2008. The cognitive benefits of interacting with nature. Psychological Science, 19(12): 1207–1212.
Bernier, P.Y., Paré, D., Stinson, G., Bridge, S.R.J., Kishchuk, B.E., Lemprière, T.C., Thiffault, E., Titus, B.D. & Vasbinder, W. 2017. Moving beyond the concept of “primary forest” as a metric of forest environment quality. Ecological Applications, 27: 349–354.
BESNet. 2019. Thematic area: Biodiversity finance. In: Biodiversity and Ecosystem Services Network [online]. Nairobi. [Cited 3 January 2020]. https://www.besnet.world/biodiversity-finance-solutions
BGCI. 2019. GlobalTreeSearch. Botanic Gardens Conservation International. Richmond, UK. [Cited 31 December 2019]. www.bgci.org/globaltree_search.php
Bharucha, Z. & Pretty, J. 2010. The roles and values of wild foods in agricultural systems. Philosophical Transactions of the Royal Society B: Biological Sciences, 365(1554): 2913–2926.
Bickford, D., Posa, M.R.C., Qie, L, Campos-Arceiz, A. & Kudavidanage, E.P. 2012 Science communication for biodiversity conservation. Biological Conservation, 151(1): 74–76.
Billings, R.F., Clarke, S.R., Mendoza, V.E., Cabrera, P.C., Figueroa, B.M., Campos, J.R. & Baeza, G. 2004. Bark beetle outbreaks and fire: A devastating combination for Central America’s pine forests. Unasylva, 55: 10–15.
Biodiversity Indicators Partnership. 2018. Living Planet Index (forest specialists). In: Biodiversity Indicators Partnership [online]. Cambridge, UK. [Cited 3 January 2020]. https://www.bipindicators.net/indicators/living-planet-index/living-planet-index-forest-specialists
Biosecurity New Zealand. 2018. Biosecurity 2025 Implementation Plan. Strengthening the biosecurity system together. Ko Tātou This Is Us. Biosecurity New Zealand 2025. Ministry for Primary Industries, Government of New Zealand. [also available at: https://www.thisisus.nz/assets/Resources/163e2a594e/Biosecurity_2025_implementation_plan_full_version.pdf]
BirdLife International. 2019. World Database on Key Biodiversity Areas [online]. [Cited 3 January 2020]. http://www.keybiodiversityareas.org/home
Blackman, A. 2015. Strict versus mixed-use protected areas: Guatemala’s Maya Biosphere Reserve. Ecological Economics, 112: 14–24.
Blackman, A. & Veit, P. 2018. Titled Amazon indigenous communities cut forest carbon emissions. Ecological Economics, 153: 56–67.
Blackman, A., Corral, L., Lima, E.S. & Asner, G.P. 2017. Titling indigenous communities protects forests in the Peruvian Amazon. PNAS, 114(16): 4123–4128.
Blackwell, S. 2015. Resilience, wellbeing and confidence development at forest schools. In: Get children outdoors [online]. [Cited 3 January 2020]. http://getchildrenoutdoors.com/resilience-wellbeing-and-confidence-development-at-forest-schools
Blomley, T. 2013. Lessons learned from community forestry in Africa and their relevance for REDD+. Washington, DC, USAID-supported Forest Carbon, Markets and Communities Program. [also available at https://rmportal.net/library/content/fcmc/publications/CF_Africa.pdf].
Blomley, T., Pfliegner, K., Isango, J., Zahabu, E., Ahrends, A. & Burgess, N.D. 2008. Seeing the wood for the trees: an assessment of the impact of participatory forest management on forest condition in Tanzania. Oryx, 42(3): 380–391.
Bocci, C., Fortmann, L., Sohngen, B. & Milian, B. 2018. The impact of community forest concessions on income: an analysis of communities in the Maya Biosphere Reserve. World Development, 107: 10–21.
Bolognesi, M., Vrieling, A., Rembold, F., & Gadain, H. 2015. Rapid mapping and impact estimation of illegal charcoal production in southern Somalia based on WorldView-1 imagery. Energy for Sustainable Development, 25: 40–49.
Bontemps, S., Defourny, P., Radoux, J., Van Bogaert, E., Lamarche, C., Achard, F., Mayaux, P. et al. 2013. Consistent global land cover maps for climate modelling communities: current achievements of the ESA’s land cover CCI. In Proceedings of the ESA Living Planet Symposium, Edinburgh, UK, 9–13 September 2013, pp. 9–13. Paris, European Space Agency. [also available at https://ftp.space.dtu.dk/pub/Ioana/papers/s274_2bont.pdf].
Borrini-Feyerabend, G., Dudley, N., Jaeger, T., Lassen, B., Pathak Broome, N., Phillips, A. & Sandwith, T. 2013. Governance of protected areas: from understanding to action. Best Practice Protected Area Guidelines Series No. 20, Gland, Switzerland, IUCN.
Bowler, D.E., Buyung-Ali, L.M., Knight, T.M. & Pullin, A.S. 2010. A systematic review of evidence for the added benefits to health of exposure to natural environments. BMC Public Health, 10: Article number 456 [online]. [Cited 3 January 2020]. https://doi.org/10.1186/1471-2458-10-456
Boyce, M.S. 2018. Wolves for Yellowstone: dynamics in time and space, Journal of Mammalogy, 99(5): 1021–1031. https://doi.org/10.1093/jmammal/gyy115
Breed, M.F., Ottewell, K.M., Gardner, M.G., Marklund, M.H.K., Dormontt, E.E. & Lowe, A.J. 2015. Mating patterns and pollinator mobility are critical traits in forest fragmentation genetics. Heredity, 115(2): 108–114.
Brinckmann, J.A., Luo, W., Xu, Q., He, X., Wu, J., & Cunningham, A.B. 2018. Sustainable harvest, people and pandas: Assessing a decade of managed wild harvest and trade in Schisandra sphenanthera. Journal of Ethnopharmacology, 224: 522–534.
Buchhorn, M., Smets, B., Bertels, L., Lesiv, M., Tsendbazar, N.-E., Herold, M. & Fritz, S. 2019. Copernicus Global Land Service: Land Cover 100m: epoch 2015: Globe. In: Zenodo [online]. Geneva, Switzerland. [Cited 3 January 2020]. https://zenodo.org/record/3243509
Burgess, D., Bahane, B., Clairs, T., Danielsen, F., Dalsgaard, S., Funder, M., Hagelberg, N. et al. 2010. Getting ready for REDD+ in Tanzania: a case study of progress and challenges. Oryx, 44(3): 339–351.
Burley, J. 2002. Forest biological diversity: an overview. Unasylva, 209: 3–9.
Burlingame, B. 2000. Editorial: Wild nutrition. Journal of Food Composition and Analysis, 13: 99–100.
Busch, J. & Ferretti-Gallon, K. 2017. What drives deforestation and what stops it? A meta-analysis. Review of Environmental Economics and Policy, 11(1): 3–23.
Camara-Leret, R. & Denney, Z. 2019. Indigenous knowledge of New Guinea’s useful plants: A review. Economic Botany, 73(3): 405–415.
Camara-Leret, R., Fortuna, M.A. & Bascompte, J. 2019. Indigenous knowledge networks in the face of global change. PNAS, 116(20): 9913–9918.
Campese, J., Sunderland, T., Greiber, T. and Oviedo, G. (eds.) 2009. Rights-based approaches: Exploring issues and opportunities for conservation. CIFOR and IUCN. Bogor, Indonesia.
Canuto, M.A., Estrada-Belli, F., Garrison, T.G., Houston, S.D., Acuña, M.J., Kováč, M., Marken, D. et al. 2018. Ancient lowland Maya complexity as revealed by airborne laser scanning of northern Guatemala. Science, 361(6409): p.eaau0137 [online]. [Cited 3 January 2020]. DOI: 10.1126/science.aau0137
Cariñanos, P., Grilo, F., Pinho, P., Casares-Porcel, M., Branquinho, C., Acil, N., Andreucci, M.B. et al. 2019. Estimation of the allergenic potential of urban trees and urban parks: towards the healthy design of urban green spaces of the future. International Journal of Environmental Research and Public Health, 16(8): 1357 [online]. [Cited 3 January 2020]. https://doi.org/10.3390/ijerph16081357
Carnus, J.-M., Parrotta, J., Brockerhoff, E., Arbez, M., Jactel, H., Kremer, A., Lamb, D., O’Hara, K. & Walters, B. 2006. Planted forests and biodiversity. Journal of Forestry, 104(2): 65–77.
Carodenuto, S. 2019. Governance of zero deforestation cocoa in West Africa: New forms of public–private interaction. Environmental Policy and Governance, 29(1): 55–66.
Carr, D.L., Suter, L., & Barbier, A. 2005. Population dynamics and tropical deforestation: State of the debate and conceptual challenges. Population and Environment, 27(1): 89–113.
Castellanos, E., Regalado, O., Pérez, G., Montenegro, R., Ramos, V., & Incer, D. 2011. Mapa de cobertura forestal de Guatemala 2006 y dinámica de la cobertura forestal 2001–2006. Guatemala, Universidad del Valle de Guatemala, Instituto Nacional de Bosques, Consejo Nacional de Áreas Protegidas, Universidad Rafael Landívar.
Castello, L., Hess, L.L., Thapa, R., McGrath, D.G., Arantes, C.C., Renó, V.F. & Isaac, V.J. 2018. Fishery yields vary with land cover on the Amazon River floodplain. Fish and Fisheries, 19(3): 431–440.
CBD. n.d.a. COP decisions – COP2 Decision II/9: Forests and biological diversity. In: Convention on Biological Diversity [online]. Montreal, Canada. [Cited 19 December 2019]. https://www.cbd.int/decision/cop/?id=7082
CBD. n.d.b. What is forest biological diversity? In: Convention on Biological Diversity [online]. Montreal, Canada. [Cited 13 December 2019]. http://www.cbd.int/forest/what.shtml
CBD. 2006. Definitions. In: Convention on Biological Diversity [online]. Montreal, Canada. [Cited 13 January 2020]. https://www.cbd.int/forest/definitions.shtml
CBD. 2009. Invasive alien species. A threat to biodiversity. Montreal, Canada, Secretariat of the Convention on Biological Diversity. [also available at https://www.cbd.int/doc/bioday/2009/idb-2009-booklet-en.pdf].
CBD. 2010a. Decision adopted by the Conference of the Parties to the Convention on Biological Diversity at its tenth meeting. X/2. The Strategic Plan for Biodiversity 2011–2020 and the Aichi Biodiversity Targets. Tenth Meeting of the Conference of the Parties to the Convention on Biological Diversity, Nagoya, Japan, 18–29 October 2010. UNEP/CBD/COP/DEC/X/2. Montreal, Canada, Secretariat of the Convention on Biological Diversity. [also available at https://www.cbd.int/doc/decisions/cop-10/cop-10-dec-02-en.pdf].
CBD. 2010b. Linking Biodiversity Conservation and Poverty Alleviation: A State of Knowledge Review. CBD Technical Series No: 55. Montreal, Canada, Secretariat of the Convention on Biological Diversity. [also available at https://www.cbd.int/doc/publications/cbd-ts-55-en.pdf].
CBD. 2011. Nagoya Protocol on Access to Genetic Resources and the Fair and Equitable Sharing of Benefits Arising from their Utilization to the Convention on Biological Diversity - Text and Annex. Montreal, Canada, Secretariat of the Convention on Biological Diversity.
CBD. 2012a. Resourcing the biodiversity targets: A first assessment of the resources required for implementing the strategic plan for biodiversity 2011–2020. Montreal, Canada, Secretariat of the Convention on Biological Diversity. [also available at https://www.cbd.int/doc/meetings/fin/hlpgar-sp-01/official/hlpgar-sp-01-01-report-en.pdf].
CBD. 2012b. Cities and biodiversity outlook. Montreal, Canada, Secretariat of the Convention on Biological Diversity.
CBD. 2014. Global Biodiversity Outlook 4. Montreal, Canada, Secretariat of the Convention on Biological Diversity. [also available at https://www.cbd.int/gbo/gbo4/publication/gbo4-en-hr.pdf].
CBD. 2016a. Ecosystem restoration: short-term action plan. Decision XIII/5 of the Conference of the Parties to the Convention on Biological Diversity. Montreal, Canada, Secretariat of the Convention on Biological Diversity. [also available at https://www.cbd.int/doc/decisions/cop-13/cop-13-dec-05-en.pdf].
CBD. 2016b. Updated assessment of progress towards Aichi Biodiversity Targets 5 and 15. Thirteenth Meeting of the Conference of the Parties to the Convention on Biological Diversity, Cancun, Mexico, 4–17 December 2016. UNEP/CBD/COP/13/INF/12. Montreal, Canada, Secretariat of the Convention on Biological Diversity.
CBD. 2017. The ABS Clearing-House. In: Convention on Biological Diversity [online]. Montreal, Canada. [Cited 26 December 2019]. https://www.cbd.int/abs/theabsch.shtml
CBD. 2018a. Decision adopted by the Conference of the Parties to the Convention on Biological Diversity. 14/8. Protected areas and other effective area-based conservation measures. 14th meeting of the Conference of the Parties to the CBD, Sharm El-Sheikh, Egypt, 17–29 November 2018. CBD/COP/DEC/14/8. Montreal, Canada, Secretariat of the Convention on Biological Diversity. [also available at https://www.cbd.int/doc/decisions/cop-14/cop-14-dec-08-en.pdf].
CBD. 2018b. Decision adopted by the Conference of the Parties to the Convention on Biological Diversity. 14/7. Sustainable wildlife management. 14th meeting of the Conference of the Parties, Sharm el-Sheikh, Egypt, 17–29 November 2018. CBD/COP/DEC/14/7. Montreal, Canada, Secretariat of the Convention on Biological Diversity. [also available at https://www.cbd.int/doc/decisions/cop-14/cop-14-dec-07-en.pdf].
CBD. 2018c. Progress of the application of the Singapore Index on Cities’ Biodiversity. Note by the Executive Secretary. 14th meeting of the Conference of the Parties, Sharm el-Sheikh, Egypt, 17–29 November. CBD/COP/14/INF/34. Montreal, Canada, Secretariat of the Convention on Biological Diversity.
CBD. 2019. The Nagoya Protocol on Access and Benefit-sharing. In: Convention on Biological Diversity [online]. Montreal, Canada. [Cited 19 December 2019]. https://www.cbd.int/abs/
CBD. 2020a. Parties to the Nagoya Protocol. In: Convention on Biological Diversity [online]. Montreal, Canada. [Cited 13 January 2020]. https://www.cbd.int/abs/nagoya-protocol/signatories/
CBD. 2020b. The Access and Benefit-Sharing Clearing-House [online]. Montreal, Canada. [Cited 13 January 2020]. https://absch.cbd.int/
CEPF. 2020. Biodiversity hotspots defined. In: Critical Ecosystem Partnership Fund [online]. Arlington, VA, USA. [Cited 13 January 2020]. https://www.cepf.net/our-work/biodiversity-hotspots/hotspots-defined
Ceres. 2019. Out on a limb: The state of corporate no-deforestation commitments and reporting indicators that count. Boston, MA, USA. [also available at www.ceres.org/sites/default/files/reports/2019-06/OutOnaLimb.pdf].
CFS. 2014. Principles for responsible investment in agriculture and food systems. Rome. [also available at http://www.fao.org/3/a-au866e.pdf].
CGRFA. 2019. First report on the implementation of the Global Plan of Action for the Conservation, Sustainable Use and Development of Forest Genetic Resources. 17th regular session, Rome, 18–22 February 2019. CGRFA-17/19/10.2/Inf.1. Rome. [also available at http://www.fao.org/3/my877en/my877en.pdf].
Chan, K.M.A., Pringle, R.M., Ranganathan, J., Boggs, C.L., Chan, Y.L., Ehrlich, P.R., Haff, P.K., Heller, N.E, Al-Khafaji, K. & Macmynowski, D.P. 2007. When agendas collide: human welfare and biological conservation. Conservation Biology, 21(1): 59–68.
Chan, L., Hillel, O., Elmqvist, T., Werner, P., Holman, N., Mader, A. & Calcaterra, E. 2014. User’s manual on the Singapore Index on Cities’ Biodiversity (also known as the City Biodiversity Index). Singapore, National Parks Board, Singapore.
Chao, S. 2012. Forest peoples: numbers across the world. Moreton-in-Marsh, UK, Forest Peoples Programme.
Chazdon, R.L., Bodin, B., Guariguata, M., Lamb, D., Walder, B., Chokkalingam, U. & Shono, K. 2017. Partnering with nature: The case for natural regeneration in forest and landscape restoration. FERI Policy Brief. Montreal, Canada, FERI.
Chomba, B.M., Tembo, O., Mutandi, K., Mtongo, C.S. & Makano, A. 2014. Drivers of deforestation, identification of threatened forests and forest co-benefits other than carbon from REDD+ implementation in Zambia. A consultancy report prepared for the Forestry Department and the Food and Agriculture Organization of the United Nations under the national UN-REDD Programme. Lusaka, Ministry of Lands, Natural Resources and Environmental Protection. [also available at http://landforlions.org/data/documents/drivers-deforestation-Zambia-WEB_final.pdf].
CITES. 1983. Convention on International Trade in Endangered Species of Wild Flora and Fauna [online]. [Cited 19 December 2019]. https://www.cites.org/sites/default/files/eng/disc/CITES-Convention-EN.pdf
CITES. 2019. Projects and initiatives – Supporting sustainable management of endangered tree species. In: Convention on International Trade in Endangered Species of Wild Flora and Fauna [online]. Geneva, Switzerland. [Cited 4 January 2020]. https://www.cites.org/eng/prog/flora/trees/trees_project
Clean Cooking Alliance. 2015. Five years of impact 2010–2015. In: Clean Cooking Alliance [online]. New York, USA, United Nations Foundation. [Cited 4 January 2020]. https://www.cleancookingalliance.org/resources/reports/fiveyears.html
Coad, L., Fa, J., Abernathy, K., Van Vliet, N., Santamaria, C., Wilkie, D.S., El Biziri, H.R., Ingram, D.J., Cawthorn, D. & Nasi, R. 2019. Towards a sustainable, participatory and inclusive wildmeat sector. Bogor, Indonesia, CIFOR.
Coady, D., Parry, I., Le, N.-P. & Shang, B. 2019. Global fossil fuel subsidies remain large: an update based on country-level estimates. IMF Working Paper. Washington, DC, IMF.
COMIFAC. 2020. Commission des Forêts d’Afrique Centrale [online]. Yaoundé. [Cited 2 January 2020]. https://comifac.org/
CONAP & WCS. 2018. Monitoreo de la Gobernabilidad en la Reserva de la Biosfera Maya: Actualización al año 2017. Con el apoyo de USAID y el USDOI/ITAP. 56 pp. San Benito, Petén, Guatemala. [also available at: https://conap.gob.gt/wp-content/uploads/2019/10/MONITOREO-DE-LA-GOBERNABILIDAD-EN-LA-RBM.pdf]
Cook, B., Anchukaitis, K., Kaplan, J., Puma, M., Kelley, M. & Gueyffier, D. 2012. Pre-Columbian deforestation as an amplifier of drought in Mesoamerica. Geophysical Research Letters, 39(16): L16706 [online]. [Cited 4 January 2020]. https://doi.org/10.1029/2012GL052565
CPW. 2016. Sustainable wildlife management and human–wildlife conflict. CPW Fact Sheet 4. Rome. [also available at http://www.fao.org/3/a-i4893e.pdf].
CRITFC. 2020. The Plan: Wy-Kan-Ush-Mi Wa-Kish-Wit. In: Colombia River Inter-Tribal Fish Commission [online]. Portland, OR, USA. [Cited 1 January 2020]. https://www.critfc.org/fish-and-watersheds/fish-and-habitat-restoration/the-plan-wy-kan-ush-mi-wa-kish-wit/
Dargie, G.C., Lewis, S.L., Lawson, I.T., Mitchard, E.T.A., Page, S.E., Bocko, Y.E. & Ifo, S.A. 2017. Age, extent and carbon storage of the central Congo Basin peatland complex. Nature, 542(7639): 86–90.
Dave, R., Saint-Laurent, C., Murray, L., Antunes Daldegan, G., Brouwer, R., de Mattos Scaramuzza, C.A., Raes, L. et al. 2019. Second Bonn Challenge progress report – application of the barometer in 2018. Gland, Switzerland, IUCN.
Davies, J.D., Hill, R., Walsh, F., Sandford, M., Smyth, D. & Holmes, M.C. 2013. Innovation in management plans for community conserved areas: Experiences from Australian indigenous protected areas. Ecology and Society, 18(2): 14 [online]. [Cited 4 January 2020]. http://dx.doi.org/10.5751/ES-05404-180214
Dawson, I.K., Leakey, R., Clement, C.R., Weber, J.C., Cornelius, J.P., Roshetko, J.M., Vinceti, B. et al. 2014. The management of tree genetic resources and the livelihoods of rural communities in the tropics: Non-timber forest products, smallholder agroforestry practices and tree commodity crops. Global Forest Genetic Resources, 333: 9–21.
Deacon, R.T. 1995. Assessing the relationship between government policy and deforestation. Journal of Environmental Economics and Management, 28(1):1–18.
Delelegn, A., Sahile, S. & Husen, A. 2018. Water purification and antibacterial efficacy of Moringa oleifera Lam. Agriculture and Food Security, 7: Article 25 [online]. [Cited 4 January 2020]. https://doi.org/10.1186/s40066-018-0177-1
de Vries, S.M.G., Alan, M., Bozzano, M., Burianek, V., Collin, E., Cottrell, J., Ivankovic, M. et al. 2015. Pan-European strategy for genetic conservation of forest trees and establishment of a core network of dynamic conservation units. European Forest Genetic Resources Programme (EUFORGEN). Rome, Bioversity International.
Ding, H., Veit, P.G., Blackman, A., Gray, E., Reytar, K., Altamirano, J.C. & Hodgdon, B. 2016. Climate benefits, tenure costs: the economic case for securing indigenous land rights in the Amazon. Washington, DC, WRI.
Dirzo, R. & Raven, P.H. 2003. Global state of biodiversity and loss. Annual Review of Environment and Resources, 28: 137–167.
Dounias, E. & Ichikawa, M. 2017. Seasonal bushmeat hunger in the Congo Basin. EcoHealth, 14: 575–590.
Dourojeanni, M. 2017. [Opinión] ¿Las sociedades prehispánicas cuidaron mejor la Amazonía? In: SPDA Actualidad Ambiental [online]. Lima. [Cited 4 January 2020]. www.actualidadambiental.pe/opinion-las-sociedades-preshispanicas-cuidaron-mejor-la-amazonia/
Drescher, M. & Brenner, J.C. 2018. The practice and promise of private land conservation. Ecology and Society 23(2) [online]. [Cited 4 January 2020]. www.jstor.org/stable/26799076
Dudley, N., Jonas, H., Nelson, F., Parrish, J., Pyhälä, A., 2018. The essential role of other effective area-based conservation measures in achieving big bold conservation targets. Global Ecology and Conservation, 15: e00424 [online]. [Cited 4 January 2020]. https://doi.org/10.1016/j.gecco.2018.e00424
Duffy, J., Godwin, C. & Cardinale, B. 2017. Biodiversity effects in the wild are common and as strong as key drivers of productivity. Nature, 549: 261–264.
EC. 2019a. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions: Stepping up EU action to protect and restore the world’s forests. COM(2019) 352 final. Brussels. (also available at https://ec.europa.eu/info/sites/info/files/communication-eu-action-protect-restore-forests_en.pdf).
EC. 2019b. Nature and biodiversity – Natura 2000. In: European Commission, Environment [online]. Brussels. [Cited 4 January 2020]. https://ec.europa.eu/environment/nature/natura2000/index_en.htm
Ege, M.J., Mayer, M., Normand, A.C., Genuneit, J., Cookson, W.O., Braun-Fahrländer, C., Heederik, D., Piarroux, R. & von Mutius, E. 2011. Exposure to environmental microorganisms and childhood asthma. The New England Journal of Medicine, 364: 701–709.
Eilers, E.J., Kremen, C., Smith Greenleaf, S., Garber, A.K. & Klein, A.-M. 2011. Contribution of pollinator-mediated crops to nutrients in the human food supply. PLOS ONE, 6(6): e21363 [online]. [Cited 13 January 2020]. https://doi.org/10.1371/journal.pone.0021363
Eriksson, M., Samuelson, L., Jägrud, L., Mattsson, E., Celander, T., Malmer, A., Bengtsson, K. et al. 2018. Water, forests, people: The Swedish Experience in building resilient landscapes. Environmental Management, 62(1): 45–57.
Erwin, T.L. 1982. Tropical forests: their richness in Coleopteran and other arthropod species. The Coleopterists’ Bulletin, 36: 74–75., cited by Dirzo, R. & Raven, P. H. 2003. Global state of biodiversity and loss. Annual Review of Environment and Resources, 28: 137–167.
ESA CCI. 2017. Global Land Cover Maps for 2015. In: Land Cover CCI Climate Research Data Package [online]. ESA Climate Change Initiative – Land Cover led by UCLouvain. https://www.esa-landcover-cci.org/?q=node/164
EU. 2011. Voluntary Partnership Agreement between the European Union and the Republic of Cameroon on forest law enforcement, governance and trade in timber and derived products to the European Union (FLEGT). 6 April. Official Journal of the European Union, 92: 4–125.
EU FLEGT Facility. n.d. FLEGT licensed timber – Essential information [online]. Brussels. [Cited 4 January 2020]. www.flegtlicence.org/home
Evans, N.P., Bauska, T.K., Gázquez-Sánchez, F., Brenner, M., Curtis, J.H. & Hodell, D.A. 2018. Quantification of drought during the collapse of the classic Maya civilization. Science, 361(6401): 498–501.
Fa, J.E., Currie, D. & Meeuwig, J. 2003. Bushmeat and food security in the Congo Basin: linkages between wildlife and people’s future. Environmental Conservation, 30: 71–78.
Fabricant, D.S. & Fransworth, N.R. 2001. The value of plants used in traditional medicine for drug discovery. Environmental Health Perspectives, 109(1): 69–75.
FairWild Foundation. 2019. The FairWild standard [online]. Cambridge, UK. [Cited 18 December 2019]. https://www.fairwild.org/the-fairwild-standard
FAO. 1989. Forestry and food security. FAO Forestry Paper No. 90. Rome. [also available at http://www.fao.org/3/T0178E/T0178E00.htm].
FAO. 1997. The State of the World’s Plant Genetic Resources for Food and Agriculture. Rome. [also available at http://www.fao.org/tempref/docrep/fao/meeting/015/w7324e.pdf].
FAO. 2006. Fire management: voluntary guidelines. Principles and strategic actions. Fire Management Working Paper 17. Rome. [also available at http://www.fao.org/3/j9255e/j9255e00.htm].
FAO. 2007. The State of the World’s Animal Genetic Resources for Food and Agriculture. Rome. [also available at http://www.fao.org/3/a-a1250e.pdf].
FAO. 2009. Declaration of the World Food Summit on Food Security [online]. Rome. [Cited 4 January 2020]. http://www.fao.org/3/w3613e/w3613e00.htm
FAO. 2010a. The Second Report on the State of the World’s Plant Genetic Resources for Food and Agriculture. Rome. [also available at http://www.fao.org/3/i1500e/i1500e.pdf].
FAO. 2010b. Global Forest Resources Assessment 2010 – Main report. FAO Forestry Paper No. 163. Rome. [also available at http://www.fao.org/3/i1757e/i1757e00.htm].
FAO. 2011a. International Plant Protection Convention. Rome, Secretariat of the International Plant Protection Convention. [also available at https://www.ippc.int/static/media/files/publication/en/2019/02/1329129099_ippc_2011-12-01_reformatted.pdf].
FAO. 2011b. State of the World’s Forests 2011. Rome. [also available at http://www.fao.org/3/i2000e/i2000e00.htm].
FAO. 2012a. Global ecological zones for FAO forest reporting: 2010 Update. Forest Resources Assessment Working Paper 179. Rome. [also available at http://www.fao.org/3/ap861e/ap861e00.pdf].
FAO. 2012b. Voluntary guidelines on the responsible governance of tenure of land, fisheries and forests in the context of national food security. Rome. [also available at http://www.fao.org/3/a-i2801e.pdf].
FAO. 2013a. Forests and trees outside forests are essential for global food security and nutrition. Summary of the International Conference on Forests for Food Security and Nutrition, Rome, 13–15 May 2013. Rome. [also available at http://www.fao.org/3/aq110e/aq110e.pdf].
FAO. 2013b. Edible insects – future prospects for food and feed security. FAO Forestry Paper No. 171. Rome. [also available at http://www.fao.org/3/i3253e/i3253e.pdf].
FAO. 2013c. Six-legged livestock: edible insect farming, collection and marketing in Thailand. RAP Publication No. 2013/03. Bangkok, Thailand, Food and Agriculture Organization of the United Nations, Regional Office for Asia and the Pacific. [also available at http://www.fao.org/3/a-i3246e.pdf].
FAO. 2014a. The State of the World’s Forest Genetic Resources. Rome. [also available at http://www.fao.org/forestry/fgr/64582/en/].
FAO. 2014b. Global plan of action for forest genetic resources. Rome. (available at http://www.fao.org/3/a-i3849e.pdf).
FAO. 2014c. State of the World’s Forests 2014. Rome. [also available at http://www.fao.org/3/a-i3710e.pdf].
FAO. 2014d. Women in forestry: Challenges and opportunities. Rome. [also available at http://www.fao.org/3/a-i3924e.pdf].
FAO. 2015a. The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture. Rome. [also available at http://www.fao.org/3/a-i4787e.pdf].
FAO. 2015b. Global guidelines for the restoration of degraded forests and landscapes in drylands: building resilience and benefiting livelihoods. Forestry Paper No. 175. Rome. [also available at http://www.fao.org/3/a-i5036e.pdf].
FAO. 2015c. Global Forest Resources Assessment 2015. How are the world’s forest changing? Rome. 2nd edition. Rome. [also available at http://www.fao.org/3/a-i4793e.pdf].
FAO. 2016a. Follow-up to the second International Conference on Nutrition. 23rd session of the Committee on Forestry, Rome, 18–22 July 2016. COFO/2016/7.4. Rome. [also available at www.fao.org/3/a-mq485e.pdf].
FAO. 2016b. Payments for forest environmental services in sub-Saharan Africa: a practical guide. Accra, FAO. [also available at http://www.fao.org/3/a-i5578e.pdf].
FAO. 2017a. Sustainable woodfuel for food security. A smart choice: green, renewable and affordable. Working paper. Rome. [also available at http://www.fao.org/3/a-i7917e.pdf].
FAO. 2017b. Strengthening sector policies for better food security and nutrition results – Forestry. Policy Guidance Note 3. Rome. [also available at http://www.fao.org/3/a-i7215e.pdf].
FAO. 2017c. Non-wood forest products in international statistical systems. Rome. [also available at http://www.fao.org/3/a-i6731e.pdf].
FAO. 2017d. The Agadir commitment towards a Mediterranean regional initiative on forest and landscape restoration. AFWC/EFC/NEFC Committee on Mediterranean Forestry Questions – Silva Mediterranea, 22nd session, Agadir, Morocco, 22 March 2017. [also available at www.fao.org/forestry/45685-0ad87e3a1d4ccc359b37c38ffcbb5b1fc.pdf].
FAO. 2017e. The future of food and agriculture – Trends and challenges. Rome. [also available at http://www.fao.org/3/a-i6583e.pdf].
FAO. 2018a. Terms and definitions: FRA 2020. Forest Resources Assessment Working Paper 188. Rome. [also available at http://www.fao.org/3/I8661EN/i8661en.pdf].
FAO. 2018b. State of the World’s Forests 2018. Rome. [also available at http://www.fao.org/3/I9535EN/i9535en.pdf].
FAO. 2018c. REDD+ finance and investments. Rome. [also available at http://www.fao.org/3/CA0907EN/ca0907en.pdf].
FAO. 2019a. The State of the World’s Biodiversity for Food and Agriculture. Rome, FAO and Commission on Genetic Resources for Food and Agriculture. [also available at http://www.fao.org/3/CA3129EN/CA3129EN.pdf].
FAO. 2019b. The State of the World’s Aquatic Genetic Resources for Food and Agriculture. Rome. [also available at http://www.fao.org/3/CA5256EN/CA5256EN.pdf].
FAO. 2019c. Trees, forests and land use in drylands: the first global assessment – Full report. FAO Forestry Paper No. 184. Rome. [also available at http://www.fao.org/3/ca7148en/ca7148en.pdf].
FAO. 2019d. International Treaty on Plant Genetic Resources for Food and Agriculture [online]. [Cited 13 January 2020]. http://www.fao.org/fileadmin/user_upload/legal/docs/033s-e.pdf
FAO. 2019e. FAOSTAT. In: Food and Agriculture Organization of the United Nations [online]. Rome. [Cited 4 January 2020]. www.fao.org/faostat
FAO. 2019f. Collaborative Partnership on Sustainable Wildlife Management. In: Food and Agriculture Organization of the United Nations [online]. Rome. [Cited 18 December 2019]. http://www.fao.org/forestry/wildlife-partnership/en/
FAO. 2019g. Restoring forest landscapes through assisted natural regeneration (ANR) – A practical manual. Bangkok. [also available at http://www.fao.org/3/ca4191en/CA4191EN.pdf]
FAO. 2019h. Action Against Desertification. In: Food and Agriculture Organization of the United Nations [online]. Rome. [Cited 4 January 2020]. http://www.fao.org/in-action/action-against-desertification
FAO. 2019i. Championing sustainable agriculture in the Caribbean Region of Colombia: a case study. Rome. [also available at www.fao.org/3/ca6753en/CA6753EN.pdf].
FAO. 2019j. Sustainable Food and Agriculture – An Integrated Approach, by Campanhola, C. and Pandey, S. (eds). FAO and Elsevier.
FAO. 2020. Global Forest Resources Assessment 2020 – Main report. Rome.
FAO. forthcoming. Analysis of 32 REDD+ Strategies. Rome.
FAO & CPF. 2018. A joint initiative of the Collaborative Partnership on Forests (CPF). Co-chairs summary report. Presented to the international conference on Working across Sectors to Halt Deforestation and Increase Forest Area – From Aspiration to Action, FAO headquarters, Rome, 20–22 February 2018.
FAO & Global Mechanism of UNCCD. 2015. Sustainable financing for forest and landscape restoration: Opportunities, challenges and the way forward. Rome, FAO. [also available at http://www.fao.org/3/a-i5174e.pdf].
FAO & Plan Bleu. 2018. State of Mediterranean forests 2018. Rome, FAO, and Marseille, France, Plan Bleu. [also available at http://www.fao.org/3/CA2081EN/ca2081en.PDF].
FAO & WRI. 2019. The road to restoration: a guide to identifying priorities and indicators for monitoring forest and landscape restoration, by Kathleen Buckingham, Sabin Ray, Carolina Gallo Granizo, Lucas Toh, Fred Stolle, Faustine Zoveda, Katie Reytar, Rene Zamora, Peter Ndunda, Florence Landsberg, Marcelo Matsumoto & John Brandt. Washington, DC, USA.
FAO, DFSC & IPGRI. 2001. Forest genetic resources conservation and management. Vol. 2: In managed natural forests and protected areas (in situ). Rome, IPGRI.
FAO, FLD & IPGRI. 2004. Forest genetic resources conservation and management. Vol. 3: In plantations and genebanks (ex situ). Rome, International Plant Genetic Resources Institute.
Fedigan, L.M. & Jack, K.M. 2012. Tracking neotropical Monkeys in Santa Rosa: Lessons from a regenerating Costa Rican dry forest. In P.M. Kappeler & D.P. Watts, eds. Long-term field studies of primates, pp. 165–184. Berlin, Springer.
Ferraro, P., Sanchirico, J., & Smith, M. 2019. Causal inference in coupled human and natural systems, PNAS, 116(12): 5311–5318.
Field, C.D., ed. 1996. Restoration of mangrove ecosystems. Okinawa, Japan, International Society for Mangrove Ecosystems.
Fisher, B. & Christopher, T. 2007. Poverty and biodiversity: Measuring the overlap of human poverty and the biodiversity hotspots. Ecological Economics, 62: 93–101.
Fluet-Chouinard, E., Funge-Smith, S. & McIntyre, P.B. 2018. Global hidden harvest of freshwater fish revealed by household surveys. PNAS, 115(29): 7623–7628.
FONAFIFO, CONAFOR and Ministry of Environment. 2012. Lessons learned for REDD+ from PES and conservation incentive programs. Examples from Costa Rica, Mexico, and Ecuador. Washington, DC, The International Bank for Reconstruction and Development/The World Bank.
Food and Land Use Coalition. 2019. Ten Critical Transitions to Transform Food and Land Use. [also available at https://www.foodandlandusecoalition.org/wp-content/uploads/2019/09/FOLU-GrowingBetter-GlobalReport.pdf]
Forest Europe. n.d. Home page [online]. Zvolen, Slovakia. [Cited 26 December 2019]. https://foresteurope.org/
Forest Europe. 2019. Human health and sustainable forest management, edited by Ľ. Marušáková & M. Sallmannshoferet. Forest Europe Study. Zvolen, Slovak Republic. [also available at https://foresteurope.org/wp-content/uploads/2017/08/Forest_book_final_WEBpdf.pdf]
Forest Trends. 2017. Supply change: Tracking corporate commitments to deforestation-free supply chains, 2017. Washington, DC.
Forest Trends. 2020. Forest Trends Supply Change Initiative [online] [Cited 17 March, 2020]. http://supply-change.org/
Fritz-Vietta, N.V.M. 2016. What can forest values tell us about human well-being? Insights from two biosphere reserves in Madagascar. Landscape and Planning 147: 28–37.
Fung, E., Imbach, P., Corrales, L., Vilchez, S. Zamora, N., Argotty, F., Hannah, L. & Ramos, Z. 2017. Mapping conservation priorities and connectivity pathways under climate change for tropical ecosystems. Climatic Change 141: 77–92.
Gaisberger, H., Kindt, R., Loo, J., Schmidt, M., Bognounou, F., Da, S.S., Diallo, O.B. et al. 2017. Spatially explicit multi-threat assessment of food tree species in Burkina Faso: A fine-scale approach. PLOS ONE, 12(9): e0184457 [online]. [Cited 4 January 2020]. https://doi.org/10.1371/journal.pone.0184457
Galetti, M. & Costa-Pereira, R. 2017. Scientists need social media influencers. Science, 357(6354): 880–881.
Galway, L.P., Acharya, Y. & Jones, A.D. 2018. Deforestation and child diet diversity: A geospatial analysis of 15 sub-Saharan African countries. Health & Place, 51: 78–88.
Gardner, C.J., Bicknell, J.E., Struebig, M.J., & Davies, Z.G. 2017. Vertebrate populations, forest regeneration and carbon: a rapid evidence assessment. Canterbury, UK, University of Kent, Durrell Institute of Conservation and Ecology.
Garnett, S.T. & Lindenmayer, D.B. 2011. Conservation science must engender hope to succeed. Trends in Ecology and Evolution, 26(2): 59–60.
Garnett, S.T., Burgess, N.D., Fa, J.E., Fernández-Llamazares, Á., Molnár, Z., Robinson, C.J., Watson, J.E. et al. 2018. A spatial overview of the global importance of indigenous lands for conservation. Nature Sustainability, 1(7): 369–374.
Gayi, S. & Tsowou, K. 2016. Cocoa industry: Integrating small farmers into the global value chain. Geneva, Switzerland, UNCTAD. [also available at https://unctad.org/en/PublicationsLibrary/suc2015d4_en.pdf].
Gentry, A.H. & Dodson, C.H. 1987. Contribution of nontrees to species richness of a tropical rain forest. Biotropica, 19:149– 56, cited by Dirzo, R. & Raven, P. H. 2003. Global state of biodiversity and loss. Annual Review of Environment and Resources, 28: 137–167.
Giller, K.E., Leeuwis, C., Andersson, J.A., Andriesse, W., Brouwer, A., Frost, P., Hebinck, P. et al1. 2008. Competing claims on natural resources: what role for science? Ecology and Society, 13(2): 34 [online]. [Cited 4 January 2020]. http://www.ecologyandsociety.org/vol13/iss2/art34/
Global Trees Campaign. 2020. Red lists. In: Global Trees Campaign [online]. [Cited 4 January 2020]. https://globaltrees.org/threatened-trees/red-list/
Golden, C.D., Fernald, L.C.H., Brashares, J.S., Rasolofoniaina, B.J.R. & Kremen, C. 2011. Benefits of wildlife consumption to child nutrition in a biodiversity hotspot. PNAS, 108: 19653–19656.
González-Oreja, J.A., Bonache-Regidor, C. & de la Fuente-Díaz-Ordaz, A.A. 2010. Far from the noisy world? Modelling the relationships between park size, tree cover and noise levels in urban green spaces of the city of Puebla, Mexico. Interciencia, 35(7): 486–492.
Gosnell, H. & Abrams, J. 2011. Amenity migration: diverse conceptualizations of drivers, socioeconomic dimensions, and emerging challenges. GeoJournal, 76, 303–322.
Government of Bhutan. 1997. Biodiversity Action Plan for Bhutan. Thimpu, Bhutan. [also available at www.cbd.int/doc/world/bt/bt-nr-01-en.pdf].
Government of the United States of America. 1973. Endangered Species Act of 1973. Washington, DC. [also available at https://www.fws.gov/international/pdf/esa.pdf].
GPFLR. n.d. What is forest and landscape restoration (FLR)? In: Global Partnership on Forest and Landscape Restoration [online]. [Cited 4 January 2020]. www.forestlandscaperestoration.org/what-forest-and-landscape-restoration-flr
Great Green Wall. 2019a. The great green wall. In: Great Green Wall [online]. Bonn, Germany. [Cited 31 December 2019]. https://www.greatgreenwall.org/about-great-green-wall
Great Green Wall. 2019b. Results. In: Great Green Wall [online]. Bonn, Germany. [Cited 4 January 2020]. www.greatgreenwall.org/results
Green, E., McRae, L., Harfoot, M., Hill, S., Simonson, W. & Baldwin-Cantello, W. 2019a. Below the canopy: plotting global trends in forest wildlife populations. Woking, UK, WWF-UK.
Green, E., McRae, L., Harfoot, M., Hill, S., & Baldwin-Cantello, W., Simonson, W. 2019b. Below the canopy: global trends in forest vertebrate populations and their drivers. PeerJ Preprints, 7: e27882v1 [online]. [Cited 4 January 2020]. https://doi.org/10.7287/peerj.preprints.27882v1
Gretzinger, S. 2016. Latin American experiences in natural forest management concessions. Forestry Policy and Institutions Working Paper 35. Rome, FAO. [also available at http://www.fao.org/forestry/45023-0707f17f1cce86c7e4f4e870bf4edd2f0.pdf].
Groenewegen, P.P., Van den Berg, A.E., De Vries, S. & Verheij, R.A. 2006. Vitamin G: effects of green space on health, well-being, and social safety. BMC public health, 6(1), 149 [online]. [Cited 4 January 2020]. https://doi.org/10.1186/1471-2458-6-149
Grogan, J., Free, C., Pinelo, G., Johnson, A. & Alegria, R. 2016. Conservation status of five timber species populations in the forestry concessions of the Maya Biosphere Reserve, Guatemala. Turrialba, Costa Rica, CATIE.
Guariguata, M., Cronkleton, P., Duchelle, A. & Zuidema, P. 2017. Revisiting the ‘cornerstone of Amazonian conservation’: a socioecological assessment of Brazil nut exploitation. Biodiversity and Conservation, 26: 2007–2027.
Gurnell, A.M., England, J., Shuker, L. & Wharton, G. 2019. The contribution of citizen science volunteers to river monitoring and management: International and national perspectives and the example of the MoRPh survey. River Research and Applications, 35(8): 1359–1373.
Gurung, J.D. 2002. Getting at the heart of the issue: Challenging male bias in Nepal’s Department of Forests. Mountain Research and Development, 22(3): 212–216.
Haddad, N.M., Brudvig, L.A., Clobert, J., Davies, K.F., Gonzalez, A., Holt, R.D., Lovejoy, T.E. et al. 2015. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Science Advances, 1: e1500052 [online]. [Cited 4 January 2020]. DOI: 10.1126/sciadv.1500052
Hansen, M.M., Jones, R., & Tocchini, K. 2017. Shinrin-yoku (forest bathing) and nature therapy: A state-of-the-art review. International Journal of Environmental Research and Public Health, 14(8): 851.
Hansen, M.C., Potapov, P.V., Moore, R., Hancher, M., Turubanova, S.A., Tyukavina, A., Thau, D. et al. 2013. High-resolution global maps of 21st-century forest cover change. Science, 342(6160): 850–853.
Hanski, I., von Hertzen, L., Fyhrquist, N., Koskinen, K., Torppa, K., Laatikainen, T., Karisola, P. et al. 2012. Environmental biodiversity, human microbiota, and allergy are interrelated. PNAS, 109(21): 8334–8339.
Hart, D. 2018. Man the hunted: primates, predators, and human evolution. New York, USA, Routledge.
Hartig, T., Mang, M., & Evans, G.W. 1991. Restorative effects of natural environment experiences. Environment and Behavior, 23(1): 3–26.
Health Council of the Netherlands. 2004. Nature and Health. The influence of nature on social, psychological and physical well-being. The Hague, Health Council of the Netherlands and the Advisory Council for Research on Spatial Planning, Nature and the Environment in the Netherlands.
Hegetschweiler, K.T., Plum, C., Fischer, C., Brändli, U.B., Ginzler, C. & Hunziker, M. 2017. Towards a comprehensive social and natural scientific forest-recreation monitoring instrument – A prototypical approach. Landscape and Urban Planning, 167: 84–97.
Henders, S., Persson, U.M. & Kastner, T. 2015. Trading forests: land-use change and carbon emissions embodied in production and exports of forest-risk commodities. Environmental Research Letters 10, no. 12, doi:10.1088/1748-9326/10/12/125012.
Henriksen, L. 2018. Blue Targeting – manual. How to do Blue Targeting for best management practice (BMP) for forestry along small streams. Swedish Forest Agency, EU Interreg project Water Management in Baltic Forests. [also available at https://www.skogsstyrelsen.se/globalassets/projektwebbplatser/wambaf/blue-targeting/blue-targeting-manual.pdf].
Hermosilla, T., Wulder, M.A., White, J.C., Coops, N.C., Pickell, P.D. & Bolton, D.K. 2019. Impact of time on interpretations of forest fragmentation: three-decades of fragmentation dynamics over Canada. Remote Sensing of Environment, 222: 65–77.
Heß, S., Jaimovich, D., & Schündeln, M. 2019. Environmental effects of development programs: Experimental evidence from West African dryland forests [online]. [Cited 13 January 2020]. http://hesss.org/Gambia%20Forest.pdf
Hilderbrand, G.V., Schwartz, C.C., Robbins, C.T., Jacoby, M.E., Hanley, T.A., Arthur, S.M. & Servheen, C. 1999. The importance of meat, particularly salmon, to body size, population productivity, and conservation of North American brown bears. Canadian Journal of Zoology, 77: 132–138.
Hill, S.L.L., Arnell, A., Maney, C., Butchart, S.H.M., Hilton-Taylor, C., Ciciarelli, C., Davis, C., Dinerstein, E., Purvis, A. & Burgess, N.D. 2019. Measuring forest biodiversity status and changes globally. Frontiers in Forest and Global Change, 2: 70 [online]. [Cited 4 January 2020]. https://doi.org/10.3389/ffgc.2019.00070
Hlásny, T., Krokene, P., Liebhold, A., Montagné-Huck, C., Müller, J., Qin, H., Raffa, K. et al. 2019. Living with bark beetles: impacts, outlook and management options. From Science to Policy 8. Barcelona, Spain, European Forest Institute.
HLPE. 2017. Sustainable forestry for food security and nutrition. A report by the High Level Panel of Experts on Food Security and Nutrition of the Committee on World Food Security. Rome. [also available at www.fao.org/3/a-i7395e.pdf].
Hoare, A. 2015. Tackling illegal logging and the related trade: what progress and where next? Chatham House Report. London, Chatham House, The Royal Institute of International Affairs.
Hodgdon, B.D., Hughell, D., Ramos, V.H. & McNab, R.B. 2015. Deforestation trends in the Maya Biosphere Reserve, Guatemala 2000–2013. New York, USA, Rainforest Alliance.
Hoffmann, B., Roeger, S., Wise, P., Dermer, J., Yunupingu, B., Lacey, D., Yunupingu, D., Marika, B., Marika, M. & Panton, B. 2012. Achieving highly successful multiple agency collaborations in a cross-cultural environment: experiences and lessons from Dhimurru Aboriginal Corporation and partners. Ecological Management and Restoration, 13(1): 42–50.
Hosonuma, N., Herold, M., De Sy, V., De Fries, R.S., Brockhaus, M., Verchot, L., Angelsen, A. & Romijn, E. 2012. An assessment of deforestation and forest degradation drivers in developing countries. Environmental Research Letters, 7(4): 044009 [online]. [Cited 4 January 2020]. https://doi.org/10.1088/1748-9326/7/4/044009
Hudson, L.N., Newbold, T., Contu, S., Hill, S.L., Lysenko, I., De Palma, A., Phillips, H.R. et al. 2017. The database of the PREDICTS project. Ecology and Evolution, 7(1): 145–188.
Hughes, T.W. & Lee, K. 2015. The role of recreational hunting in the recovery and conservation of the wild turkey (Meleagris gallopavo spp.) in North America. International Journal of Environmental Studies, 72(5): 797–809.
Huntley, B.J. & Redford, K.H. 2014. Mainstreaming biodiversity in practice: a STAP advisory document. Washington, DC, USA, GEF.
Ickowitz, A., Powell, B., Salim, M.A. & Sunderland, T. 2014. Dietary quality and tree cover in Africa. Global Environmental Change, 24: 287–294.
IDH. 2019. Green Cocoa Cameroon. In: IDH, The Sustainable Trade Initiative [online]. Utrecht, The Netherlands. [Cited 4 January 2020]. https://www.idhsustainabletrade.com/contact-directions/
IFAD & UNEP. 2013. Smallholders, food security, and the environment. Rome, IFAD.
IIED. 2019. Darwin Initiative Main and Post Project Annual Report: Livelihoods Insurance from Elephants (LIFE) in Kenya and Sri Lanka [online]. London. [Cited 4 January 2020]. https://pubs.iied.org/pdfs/G04412.pdf
ILO. 2017. NORMLEX – C169 – Indigenous and Tribal Peoples Convention, 1989 (No. 169). In: International Labour Organization [online]. Geneva, Switzerland. [Cited 2 January 2020]. https://www.ilo.org/dyn/normlex/en/f?p=NORMLEXPUB:12100:0::NO::P12100_ILO_CODE:C169
INAB. 2019. Cobertura forestal. In: SIFGUA – Sistema de Información Forestal de Guatemala [online]. Guatemala. [Cited 4 January 2020]. http://www.sifgua.org.gt/Cobertura.aspx
Ingwall-King, L. & Gangur, A. forthcoming. Integrating traditional knowledge into conservation policy and practice: a good practice review. Cambridge, UK, UNEP-WCMC.
Initiative 20x20. n.d. Healthy lands for food, water and nature [online]. Washington, DC. [Cited 18 December 2019]. https://initiative20x20.org/
Institute of Medicine. 2001. Dietary reference intakes for vitamin A, vitamin K, arsenic, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. Washington, DC, National Academies Press.
Instituto Socioambiental. 2015. Advances and setbacks in territorial rights in Brazil. Brasilia. Cited in RRI. 2015. Protected areas and the land rights of indigenous peoples and local communities: current issues and future agenda. Washington, DC, RRI.
IPBES. 2016. The assessment report on pollinators, pollination and food production-policy platform on biodiversity and ecosystem services on pollinators, pollination and food production. Bonn, Germany.
IPBES. 2019a. Summary for policymakers of the global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Bonn, Germany.
IPBES. 2019b. Chapter 2.2 Status and Trends – Nature. Unedited draft chapter for IPBES Global Assessment on Biodiversity and Ecosystem Services [online]. Bonn, Germany. [Cited 13 January 2020]. https://ipbes.net/sites/default/files/ipbes_global_assessment_chapter_2_2_nature_unedited_31may.pdf].
IPCC. 2019. Climate Change and Land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems [P.R. Shukla, J. Skea, E. Calvo Buendia, V. Masson-Delmotte, H.-O. Pörtner, D. C. Roberts, P. Zhai, R. Slade, S. Connors, R. van Diemen, M. Ferrat, E. Haughey, S. Luz, S. Neogi, M. Pathak, J. Petzold, J. Portugal Pereira, P. Vyas, E. Huntley, K. Kissick, M. Belkacemi, J. Malley (eds.)]. [also available at: https://www.ipcc.ch/srccl/]
Irvine, K.N., Devine-Wright, P., Payne, S.R., Fuller, R.A., Painter, B. & Gaston, K.J. 2009. Green space, soundscape and urban sustainability: an interdisciplinary, empirical study. Local Environment, 14(2): 155–172.
Isted, A. 2013. An investigation into the benefits of forest school intervention for young people with ADHD in the education system (Examination paper). London, University of Greenwich.
ITC. 2016. Sustainable sourcing: Markets for certified Chinese medicinal and aromatic plants. Geneva, Switzerland.
ITTO. 2002. ITTO guidelines for the restoration, management and rehabilitation of degraded and secondary tropical forests. ITTO, CIFOR, FAO, IUCN and WWF International.
ITTO & IUCN. 2009. ITTO/IUCN Guidelines for the conservation and sustainable use of biodiversity in tropical timber production forests. ITTO Policy Development Series No. 17. Yokohama, Japan, ITTO.
IUCN. 2013. Mitigating human-wildlife conflict. In: IUCN [online]. Gland, Switzerland. [Cited 4 January 2020]. https://www.iucn.org/content/mitigating-human-wildlife-conflict
IUCN. 2016. A global standard for the identification of key biodiversity areas. Version 1.0. First edition. Gland, Switzerland.
IUCN. 2017. The IUCN red list of threatened species. Version 2017.3. http://www.iucnredlist.org.
IUCN. 2018. The Bonn Challenge barometer. In: InfoFLR [online]. Gland, Switzerland. [Cited 13 January 2020]. https://infoflr.org/bonn-challenge-barometer
IUCN. 2019a. The IUCN red list of threatened species. Version 2019-2. http://www.iucnredlist.org. Downloaded on 4 October 2019.
IUCN. 2019b. Over half of Europe’s endemic trees face extinction. In: IUCN [online]. Gland, Switzerland. [Cited 4 January 2020]. https://www.iucn.org/news/species/201909/over-half-europes-endemic-trees-face-extinction
IUCN WCPA. 2018. PARKS. The International Journal of Protected Areas and Conservation. Volume 24 Special Issue. Gland, Switzerland, IUCN.
Jalonen, R., Valette, M., Boshier, D., Duminil, J. & Thomas, E. 2017. Forest and landscape restoration severely constrained by a lack of attention to the quantity and quality of tree seed: Insights from a global survey. Conservation Letters, 11(4): e12424 [online]. [Cited 4 January 2020]. https://doi.org/10.1111/conl.12424
Jamnadass, R., McMullin, S., Iiyama, M., Dawson, I.K., Powell, B., Termote, C., Ickowitz, A. et al. 2015. Understanding the roles of forests and tree-based systems in food provision. In B. Vira, C. Wildburger & S. Mansourian, eds. Forests and food: Addressing hunger and nutrition across sustainable landscapes. Cambridge, UK, Open Book Publishers. http://dx.doi.org/10.11647/OBP.0085
Jayachandran S., de Laat, J., Lambin, E.F., Stanton, C.Y., Audy, R. & Thomas, N.E. 2017. Cash for carbon: A randomized trial of payments for ecosystem services to reduce deforestation. Science, 357(6348): 267–273.
Jenkins, M., Timoshyna, A. & Cornthwaite, M. 2018. Wild at home: exploring the global harvest, trade and use of wild plant ingredients. Cambridge, UK, TRAFFIC International.
Jonas, H.D., MacKinnon K., Dudley N., Hockings M., Jessen S., Laffoley D., MacKinnon D. et al. 2018. Editorial essay: Other effective area-based conservation measures: From Aichi Target 11 to the Post-2020 biodiversity framework. PARKS, The International Journal of Protected Areas and Conservation, 24 (Special issue on OECMs): 9–16.
Jorgensen, A., Hitchmough, J. & Dunnet, N. 2006. Woodland as a setting for housing-appreciation and fear and the contribution of residential satisfaction and place identity in Warrington New Town, UK. Landscape and Urban Planning, 79(3–4): 273–287.
Kaimowitz, D., & Sheil, D. 2007. Conserving what and for whom? Why conservation should help meet basic human needs in the tropics. Biotropica, 39(5): 567–574.
Kaplan, R. & Kaplan, S. 1989. The experience of nature – a psychological perspective. Cambridge, UK, Cambridge University Press.
Kapos, V., Lysenko, I. & Lesslie, R. 2002. Assessing forest integrity and naturalness in relation to biodiversity. FAO Forest Resources Assessment Programme Working Paper 54. Rome. [also available at http://www.fao.org/3/ad654e/ad654e00.htm].
Kareiva, P., Watts, S., McDonald, R. & Boucher, T. 2007. Domesticated nature: Shaping landscapes and ecosystems for human welfare. Science, 316(5833): 1866–1869.
Katila, P., Pierce Colfer, C., De Jong, W., Galloway, G., Pacheco, P., & Winkel, G., eds. 2019. Sustainable Development Goals: their impacts on forests and people. Cambridge, UK, Cambridge University Press.
Kawarazuka, N. & Béné, C. 2011. The potential role of small fish species in improving micronutrient deficiencies in developing countries: building evidence. Public Health Nutrition, 14(11): 1927–1938.
Kay, C.E. 2018. The Condition and Trend of Aspen, Willows, and Associated Species on the Northern Yellowstone Range. Rangelands, 40(6): 202–211. [also available at https://www.sciencedirect.com/science/article/pii/S0190052818300774?via%3Dihub]
Keenan, R.J., Reams, G.A., Achard, F., de Freitas, J.V., Grainger, A. & Lindquist, E. 2015. Dynamics of global forest area: Results from the FAO Global Forest Resources Assessment 2015. Forest Ecology and Management, 352: 9–20.
Kelleher, C.T., de Vries, S.M.G., Baliuckas, V., Bozzano, M., Frydl, J., Gonzalez Goicoechea, P., Ivankovic, M. et al. 2015. Approaches to the conservation of forest genetic resources in Europe in the context of climate change. European Forest Genetic Resources Programme (EUFORGEN). Rome, Bioversity International.
Kerr, J., Pender, J. & Suyanto, B.L. 2008. Property rights, environmental services and poverty alleviation in Indonesia. BASIS Brief 2008-03. Madison, WI, University of Wisconsin.
King, L., Lala, F., Nzumu, H., Mwambingu, E. & Douglas-Hamilton, I. 2017. Beehive fences as a multidimensional conflict-mitigation tool for farmers coexisting with elephants. Conservation Biology, 31(4): 743–752.
Klein, A.M., Vaissiere, B.E., Cane, J.H., Steffan-Dewenter, I., Cunningham, S.A., Kremen, C. & Tscharntke, T. 2007. Importance of pollinators in changing landscapes for world crops. Proceedings of the Royal Society B – Biological Sciences, 274: 303–313.
Koskela, J., Lefèvre, F., Schueler, S., Kraigher, H., Olrik, D.C., Hubert, J., Longauer, R. et al. 2013. Translating conservation genetics into management: Pan-European minimum requirements for dynamic conservation units of forest tree genetic diversity. Biological Conservation, 157: 39–49.
Koskela, J., Vinceti, B., Dvorak, W., Bush, D., Dawson, I., Loo, J., Kjær, E.D. et al. 2014. Use and transfer of forest genetic resources: A global review. Forest Ecology and Management, 333: 22–34.
Krishnan, S., Wiederkehr Guerra, G., Bertrand, D., Wertz-Kanounnikoff, S. & Kettle, C. forthcoming. Enhancing the cross-sectoral benefits from forests for pollination services at landscape scales: a review of management interventions. [tentative title]. FAO working paper. Rome, FAO and Bioversity International.
Lambin, E.F., & Meyfroidt, P. 2011. Global land use change, economic globalization, and the looming land scarcity. PNAS, 108(9): 3465–3472.
Lambin, E.F., Turner, B.L., Geist, H.J., Agbola, S.B., Angelsen, A., Bruce, J.W., Coomes, O.T. et al. 2001. The causes of land-use and land-cover change: moving beyond the myths. Global Environmental Change, 11(4): 261–269.
Laurance, W.F., Nascimento, H.E.M., Laurance, S.G., Andrade, A.C., Fearnside, P.M., Ribeiro, J.E.L. & Capretz, R.L. 2006. Rain forest fragmentation and the proliferation of successional trees. Ecology, 87(2): 469–482.
Le Bel, S., Mapuivre, G. & Czudek, R. 2010. Human–wildlife conflict toolkit: comprehensive solutions for farmers and communities. Unasylva, 236: 12–13.
Lefèvre, F., Koskela, J., Hubert, J., Kraigher, H., Longauer, R., Olrik, D.C., Schüler, S. et al. 2013. Dynamic conservation of forest genetic resources in 33 European countries. Conservation Biology, 27(2): 373–384.
Lele, S., Wilshusen, P., Brockington, D., Seidler, R. & Bawa, K. 2010. Beyond exclusion: alternative approaches to biodiversity conservation in the developing tropics. Current Opinion in Environmental Sustainability, 2(1): 94–100.
Leticia Pact. 2019. Leticia Pact for the Amazon. [Cited 2 January 2020]. https://id.presidencia.gov.co/Documents/190906-Pacto-Leticia-Amazonia-Ingles.pdf
Leverington, F., Lemos Costa, K., Pavese, H., Lisle, A. & Hockings, M. 2010. A global analysis of protected area management effectiveness. Environmental Management, 46(5): 685–698.
Levis, C., Costa, F.R., Bongers, F., Peña-Claros, M., Clement, C.R., Junqueira, A.B., Neves, E.G. et al. 2017. Persistent effects of pre-Columbian plant domestication on Amazonian forest composition. Science, 355(6328): 925–931.
Lham, D., Wangchuk, S., Stolton, S. & Dudley, N. 2019. Assessing the effectiveness of a protected area network: a case study of Bhutan. Oryx, 53(1): 63–70.
Li, Q., Morimoto, K., Kobayashi, M., Inagaki, H., Katsumata, M., Hirata, Y., Hirata, K. et al. 2008. Visiting a forest, but not a city, increases human natural killer activity and expression of anti-cancer proteins. International Journal of Immunopathology and Pharmacology, 21: 117–128.
Lindenmayer, D.B. & Fischer, J. 2006. Habitat fragmentation and landscape change: An ecological and conservation synthesis. Washington, DC, Island Press.
Linnell, J.D. & Alleau, J. 2016. Predators that kill humans: myth, reality, context and the politics of wolf attacks on people. In F.M. Angelici, ed. Problematic wildlife: A cross-disciplinary approach, pp. 357–371. Cham, Switzerland, Springer.
Liu, X., Li, Y., Guasch-Ferré, M., Willett, W.C., Drouin-Chartier, J.-P., Bhupathiraju, S.N. & Tobias, D.K. 2019. Changes in nut consumption influence long-term weight change in US men and women. BMJ Nutrition, Prevention & Health, 2(2) [online] [Cited 13 January 2020]. http://dx.doi.org/10.1136/bmjnph-2019-000034
Lo, M., Narulita, S. & Ickowitz, A. 2019. The relationship between forests and freshwater fish consumption in rural Nigeria. PLOS ONE, 14(6): e0218038 [online]. [Cited 4 January 2020]. https://doi.org/10.1371/journal.pone.0218038
Lobón-Cerviá, J., Hess, L.L., Melack, J.M. & Araujo-Lima, C.A. 2015. The importance of forest cover for fish richness and abundance on the Amazon floodplain. Hydrobiologia, 750(1): 245–255.
Lompo, D., Vinceti, B., Gaisberger, H., Konrad, H., Duminil, J., Quedraogo, M., Sina, S. & Geburek, T. 2017. Genetic conservation in Parkia biglobosa (Fabaceae: Mimosoideae) – what do we know? Silvae Genetica, 66(1): 1–8.
Lompo, D., Vinceti, B., Konrad, H., Gaisberger, H. & Geburek, T. 2018. Phylogeography of African locust bean (Parkia biglobosa) reveals genetic divergence and spatially structured populations in West and Central Africa. Journal of Heredity, 109(7): 811–824.
Luke (Natural Resources Institute Finland). 2018. 5+1 Steps towards a functioning insect economy. In: Luke, Natural Resources Institute Finland [online]. Helsinki. [Cited 4 January 2020]. https://www.luke.fi/en/51-steps-towards-functioning-insect-economy/
Lung, T. & Schaab, G. 2010. A comparative assessment of land cover dynamics of three protected forest areas in tropical eastern Africa. Environmental Monitoring and Assessment, 161(1): 531–548.
Lupala, Z.J., Lusambo, L.P., Ngaga, Y.M. & Makatta, A.A. 2015. The land use and cover change in Miombo woodlands under community based forest management and its implication to climate change mitigation: a case of southern highlands of Tanzania. International Journal of Forestry Research, Volume 2015: Article ID 459102 [online]. [Cited 4 January 4, 2020]. http://dx.doi.org/10.1155/2015/459102
Maas, J., Verheij, R.A., Groenewegen, P.P., de Vries, S. & Spreeuwenberg, P. 2006. Green space, urbanity, and health: how strong is the relation? Journal of Epidemiology and Community Health, 60(7): 587–592.
Mace, G.M. 2014. Whose conservation? Science, 345(6204): 1558–1560.
Mahoney, S.P. & Geist, V., eds. 2019. The North American model of wildlife conservation. Baltimore, MD, USA, Johns Hopkins University Press.
Maisels, F., Strindberg, S., Blake, S., Wittemyer, G., Hart, J., Williamson, E.A., Aba’a, R. et al. 2013. Devastating decline of forest elephants in central Africa. PLOS ONE, 8(3): e59469 [online]. [Cited 4 January 2020]. https://doi.org/10.1371/journal.pone.0059469
Maxwell, S.L., Fuller, R.A., Brooks, T.M. & Watson, J.E.M. 2016. The ravages of guns, nets and bulldozers. Nature, 536(7615): 143–145.
May, R. 2010. Tropical arthropod species, more or less? Science, 329(5987): 41–42.
Mbora A., Jamnadass R. & Lillesø J.-P.B. 2008. Growing high priority fruits and nuts in Kenya: Uses and management. Nairobi, The World Agroforestry Centre.
McDonell, E. 2019. Creating the culinary frontier. A critical examination of Peruvian chefs’ narratives of lost/discovered foods. Anthropology of Food, 14 [online]. [Cited 4 January 2020]. http://journals.openedition.org/aof/10183
McFarlane, R.A., Barry, J., Cissé, G., Gislason, M., Gruca, M., Higgs, K., Horwitz, P. et al. 2019. SDG 3: Good health and well-being – framing targets to maximise co-benefits for forests and people. In P. Katila, C.J. Pierce Colfer, W. de Jong, G. Gallowa, P. Pacheco & G. Winkel, eds. Sustainable Development Goals: their impacts on forests and people, pp. 72–107. Cambridge, UK, Cambridge University Press.
McKeown, R. 2002. Education for sustainable development toolkit. Version 2. [Cited 4 January 2020]. http://esdtoolkit.org/esd_toolkit_v2.pdf
McShane, T.O., Hirsch, P.D., Trung, T.C., Songorwa, A.N., Kinzig, A., Monteferri, B., Mutekanga, D. et al. 2011. Hard choices: Making trade-offs between biodiversity conservation and human well-being. Biological Conservation, 144: 966–972.
MEA. 2005. Ecosystems and human well-being: current state and trends. Washington, DC, Island Press.
Medaglia, J.C., Phillips, L.-K. & Perron-Welch, F. 2014. Biodiversity legislation study: a review of biodiversity legislation in 8 countries. London, The Global Legislators’ Organisation, Hamburg, Germany, the World Future Council, and Montreal, Canada, the Centre for International Sustainable Development Law. [also available at http://www.cisdl.org/wp-content/uploads/2018/04/Biodiversity-Legislation-Study.pdf]
MEF (Ministry of Environment and Forestry). 2018. The state of Indonesia’s forests 2018. Jakarta.
MERECP. 2007. Mount Elgon Regional Ecosystem Conservation Programme (MERECP), Work Plan (version March 2007). Nairobi, Kenya.
Min, Q. 2017. Learning from the past for the future: experiences of Hani Rice Terraces in coping with extreme drought. Presentation at a side event on Globally Important Agricultural Heritage Systems and Climate Change, 23rd session of the Conference of the Parties to UNFCCC, Bonn, Germany, 10 November.
MINEF. 1998. Décision No. 0108/D/MINEF/CAB du 9 février 1998: “Portant application des normes d’intervention en milieu forestier en République du Cameroun.” Chapitre VI, Articles 28, 29 et 30 – “Protection de la faune.” Yaoundé.
MINEF. 2001. Order No. 0222/A/MINEF of May 25, 2002 on “procedures for developing, approval, monitoring and control of the implementation of forest management plans for the production forests in the permanent forest estate.” Article 11(1) and (3). Yaoundé.
MINEPDED. 2013. “Readiness Preparation Proposal (R-PP) submitted to the World Bank’s Forest Carbon Partnership Facility (FCPF)” (unpublished).
MIPAAF. 2017. Comunicati stampa – Creato primo elenco alberi monumentali d’Italia [Press release – First list of monumental trees of Italy created]. In: Ministero delle politiche agricole alimentari e forestali [online]. Rome. [Cited 4 January 2020]. https://www.politicheagricole.it/flex/cm/pages/ServeBLOB.php/L/IT/IDPagina/12052
MIPAAF. 2019. Elenco degli alberi monumentali d’Italia ai sensi della Legge n. 10/2013 e del Decreto 23 ottobre 2014 [List of the monumental trees of Italy under Law No. 10/2013 and the Decree of 23 October 2014]. In: Ministero delle politiche agricole alimentari e forestali [online]. Rome. [Cited 4 January 2020]. www.politicheagricole.it/flex/cm/pages/ServeBLOB.php/L/IT/IDPagina/11260
Mitchell, R. & Popham, F. 2008. Effect of exposure to natural environment on health inequalities: an observational population study. Lancet, 372(9650): 1655–1660.
Mittermeier, R.A., Myers, N., Thomsen, J.B., da Fonseca, G.A.B. & Olivieri, S. 1998. Biodiversity hotspots and major tropical wilderness areas: approaches to setting conservation priorities. Conservation Biology, 12(3): 516–520.
Mittermeier, R.A., Gil, P.R., Hoffman, M., Pilgrim, J., Brooks, T., Mittermeier, C.G., Lamoreux, J. & da Fonseca, G.A.B. 2004. Hotspots revisited: Earth’s biologically richest and most endangered terrestrial ecoregions. Colonia Centro, Monterrey, Mexico, Cemex.
Mittermeier, R.A., Turner, W.R., Larsen, F.W., Brooks, T.M. & Gascon, C. 2011. Global biodiversity conservation: The critical role of hotspots. In F.E. Zachos & J.C. Habel, eds. Biodiversity hotspots: Distribution and protection of conservation priority areas, pp. 3–22. Berlin, Springer, cited by IPBES. 2019b. Chapter 2.2 Status and Trends – Nature. Unedited draft chapter for IPBES Global Assessment on Biodiversity and Ecosystem Services [online]. Bonn, Germany. [Cited 13 January 2020]. https://ipbes.net/sites/default/files/ipbes_global_assessment_chapter_2_2_nature_unedited_31may.pdf
MNRT. 2015. National Forest Resources Monitoring and Assessment of Tanzania mainland (NAFORMA). Main results. Dar es Salaam, MNRT.
Molinario, G., Hansen, M., Potapov, P., Tyukavina, A. & Stehman, S. 2020. Contextualizing Landscape-Scale Forest Cover Loss in the Democratic Republic of Congo (DRC) between 2000 and 2015. Land 9(1), 23. [also available at https://doi.org/10.3390/land9010023]
Monbiot, G. 2013. Feral: Rewilding the Land, Sea and Human Life. Penguin.
Mongbo, R., Floquet, A., Choden, S. & Moreno Diaz, M.L. 2011. Protected areas – Not just for biodiversity conservation. The contributions of protected areas to the economic and social development in Bhutan, Costa Rica and Benin. Costa Rica, Universidad Nacional.
MoP (Ministry of Planning and International Cooperation) & MoE (Ministry of Environment Jordan). 2008. Integrated financing strategy for sustainable land management in Jordan. Final report. Amman. [also available at http://extwprlegs1.fao.org/docs/pdf/jor169877.pdf].
Mora, C., Tittensor, D.P., Adl, S., Simpson, A.G.B. & Worm, B. 2011. How many species are there on Earth and in the ocean? PLOS Biology, 9(8): e1001127 [online]. [Cited 5 January 2020]. https://doi.org/10.1371/journal.pbio.1001127
Mulenga, B.P., Tembo, S.T. & Richardson, R.B. 2019. Electricity access and charcoal consumption among urban households in Zambia. Development Southern Africa, 36(5): 585–599.
Myers, N. 1990. The biodiversity challenge: Expanded hot-spots analysis. Environmentalist, 10(4): 243–256.
Myers, N., Mittermeier, R.A., Mittermeier, C.G., da Fonseca, G.A.B. & Kent, J. 2000. Biodiversity hotspots for conservation priorities. Nature, 403: 853–858.
NACSO. 2017a. Human wildlife conflict – the hot potato. In: Namibian Association of CBNRM Support Organizations [online]. Windhoek. [Cited 5 March 2019]. http://www.nacso.org.na/news/2017/03/human-wildlife-conflict-%E2%80%93-the-hot-potato
NACSO. 2017b. Resources & publications: State of Community Conservation figures and tables. In: Namibian Association of CBNRM Support Organizations [online]. [Cited 18 December 2019]. http://www.nacso.org.na/resources/state-of-community-conservation-figures-and-tables
Nadkarni, N. 2004. Not preaching to the choir: Communicating the importance of forest conservation to nontraditional audiences. Conservation Biology, 18(3): 602–606.
Nasi, R., Taber, A. & Van Vliet, N. 2011. Empty forests, empty stomachs? Bushmeat and livelihoods in the Congo and Amazon Basins. International Forestry Review, 13(3): 355–368.
Nasi, R., Brown, D., Wilkie, D., Bennett, E., Tutin, C., van Tol, G. & Christophersen, T. 2008. Conservation and use of wildlife-based resources: the bushmeat crisis. Technical Series No. 33. Montreal, Canada, Secretariat of the Convention on Biodiversity, and Bogor, Indonesia, CIFOR.
Nature4Climate. 2019. Nature-based solutions: a summary of announcements and developments during the UN Climate Action Summit and Climate Week. In: Nature4Climate [online]. [Cited 5 January 2020]. https://nature4climate.org/news/nature-based-solutions-a-summary-of-announcements-and-developments-during-the-un-climate-action-summit-and-climate-week
NCED. 2019. What is a conservation easement? In: NCED, National Conservation Easement Database [online]. Greenville, SC, USA. [Cited 5 January 2020]. www.conservationeasement.us/what-is-a-conservation-easement
Nel, A. & Hill, D. 2013. Constructing walls of carbon–the complexities of community, carbon sequestration and protected areas in Uganda. Journal of Contemporary African Studies, 31(3): 421–440.
Nellemann, C., Henriksen, R., Kreilhuber, A., Stewart, D., Kotsovou, M., Raxter, P., Mrema, E. & Barrat, S., eds. 2016. The rise of environmental crime: A growing threat to natural resources peace, development and security. Nairobi, UNEP, and Oslo, Norwegian Center for Global Analyses (RHIPTO).
Nelson F. & Sinandei, M. 2018. Building stronger grassroots organizations that can take community land rights to scale. In: Land portal [online]. Amersfoort, The Netherlands. [Cited 5 January 2020]. https://landportal.org/blog-post/2018/03/building-stronger-grassroots-organizations-can-take-community-land-rights-scale
New Generation Plantations. 2018. Rainforest restoration in Brazil’s Atlantic Forest [online]. [Cited 13 December 2019]. https://newgenerationplantations.exposure.co/rainforest-restoration-in-brazils-atlantic-fores
Newton, P., Miller, D.C., Byenkya, M.A.A. & Agrawal, A. 2016. Who are forest-dependent people? A taxonomy to aid livelihood and land use decision-making in forested regions. Land Use Policy, 57: 388–395.
Nguinguiri, J.C., Czudek, R., Larrubia, C.J., Ilama, L., Le Bel, S., Angoran, E.J., Trebuchon, J.F. & Cornelis, D. 2017. Managing human–wildlife conflicts in central and southern Africa. Unasylva, 249: 39–44.
Nielsen, M.R., Meilby, H., Smith-Hall, C., Pouliot, M. & Treue, T. 2018. The importance of wild meat in the global south. Ecological Economics, 146: 696–705.
Nilsson M., Griggs D. & Visbeck M. 2016. Policy: Map the interactions between Sustainable Development Goals. Nature, 534: 320–322.
Nilsson, K., Sangster, M., Gallis, C., Hartig, T., De Vries, S., Seeland, K. & Schipperijn, J., eds. 2010. Forests, trees and human health. New York, USA, Springer Science + Business Media.
Nirmal, S.A., Pal, S.C., Otimenyin, S.O., Aye, T., Elachouri, M., Kundu, S.K., Thandavarayan, R.A. & Mandal, S.C. 2013. Contribution of herbal products in the global market. The Pharma Review, November–December 2013: 95–104.
Norgrove, L. & Hulme, D. 2006. Confronting conservation at Mount Elgon, Uganda. Development and Change, 37(5): 1093–1116.
Nowak, D.J., Crane, D.E. & Stevens, J.C. 2006. Air pollution removal by urban trees and shrubs in the United States. Urban Forestry & Urban Greening, 4(3–4): 115–123.
Nwaokoro, N. & Kwon-Ndung, E. 2010. Exploiting the potentials of Parkia biglobosa in Nigeria. Paper presented at Plant Biology 2010, Joint Annual Meeting of the American Society of Plant Biologists and the Canadian Society of Plant Physiologists– La Société Canadienne de Physiologie Végétale, Montreal, Canada, 31 July–4 August 2010.
NYDF. 2019. Protecting and restoring forests: A story of large commitments yet limited progress. New York Declaration on Forests Five-year assessment report. Amsterdam, Climate Focus.
Nyhus, P.J. 2016. Human–wildlife conflict and coexistence. Annual Review of Environment and Resources, 41: 143–171.
O’Brien, L. 2009. Learning outdoors: The Forest School approach. Education 3–13, 37(1): 45–60.
O’Brien, L. & Murray, R. 2007. Forest school and its impacts on young children: case studies in Britain. Urban Forestry & Urban Greening, 6(4): 249–265.
Ødegaard, F. 2000. How many species of arthropods? Erwin’s estimate revised. Biological Journal of the Linnean Society, 71(4): 583–597.
Odetokun, S.M. 1996. The nutritive value of baobab fruit (Adansonia digitata). Rivista Italiana delle Sostanze Grasse, 73: 371–373, cited by Manfredini, S., Vertuani, S. & Buzzoni, V. 2002. Adansonia digitata. Il baobab farmacista. L’integratore nutrizionale, 5: 25–29.
OECD. 2019a. Agricultural policy monitoring and evaluation 2019. Paris.
OECD. 2019b. Biodiversity: Finance and the economic and business case for action. Paris.
Olival, K.J., Hosseini, P.R., Zambrana-Torrelio, C., Ross, N., Bogich, T.L. & Daszak, P. 2017. Host and viral traits predict zoonotic spillover from mammals. Nature, 546: 646–650.
Ollerton, J., Winfree, R. & Tarrant, S. 2011. How many flowering plants are pollinated by animals? Oikos, 120(3): 321–326.
Olson, D.M., Dinerstein, E., Wikramanayake, E.D., Burgess, N.D., Powell, G.V.N., Underwood, E.C., D’Amico, J.A. et al. 2015. Terrestrial ecoregions of the world: A new map of life on Earth. BioScience, 51(11): 933–938.
Onana, J.-M., Cheek, M. & Pollard, B. 2011. Red Data Book of the Flowering Plants of Cameroon: IUCN global assessments. Richmond, Surrey, UK, Kew Publishing.
Ong, S. & Carver, E. 2019. The rosewood trade: An illicit trail from forest to furniture. In: YaleEnvironment360 [online]. New Haven, CT, USA. [Cited 5 January 2020]. https://e360.yale.edu/features/the-rosewood-trade-the-illicit-trail-from-forest-to-furniture
Oregon Fish and Wildlife Office. n.d. Northern spotted owl. In: U.S. Fish& Wildlife Service, Oregon Fish and Wildlife Office [online]. Washington, DC. [Cited 5 January 2020]. www.fws.gov/oregonfwo/articles.cfm?id=149489595
Orgiazzi, A., Bardgett, R., Barrios, E., Behan Pelletier, V., Briones, M.J.I., Chotte, J-L., De Deyn, G. et al. eds. 2016. Global Soil Biodiversity Atlas. European Commission, Publications Office of the European Union, Luxembourg.
Ostrom, E. & Nagendra, H. 2006. Insights on linking forests, trees, and people from the air, on the ground, and in the laboratory. PNAS, 103(51): 19224–19231.
Osuri, A.M., Ratnam, J., Varma, V., Alvarez-Loayza, P., Hurtado Astaiza, J., Bradford, M., Fletcher, C. et al. 2016. Contrasting effects of defaunation on aboveground carbon storage across the global tropics. Nature Communications, 7: 11351 [online]. [Cited 5 January 2020]. https://doi.org/10.1038/ncomms11351
Park, B.J., Tsunetsugu, Y., Kasetani, T., Kagawa, T. & Miyazaki, Y. 2010. The physiological effects of Shinrin-yoku (taking in the forest atmosphere or forest bathing): evidence from field experiments in 24 forests across Japan. Environmental Health and Preventive Medicine, 15: 18 [online]. [Cited 5 January 2020]. https://doi.org/10.1007/s12199-009-0086-9
Patenaude, G. & Lewis, K. 2014. The impacts of Tanzania’s natural resource management programmes for ecosystem services and poverty alleviation. International Forestry Review, 16(4): 459−473.
Paumgarten, F., Locatelli, B. & Witkowski, E.T.F. 2018. Wild foods: safety net or poverty trap? A South African case study. Human Ecology, 46(2): 183–195.
Payn, T., Carnus, J.M., Freer-Smith, P., Kimberley, M., Kollert, W., Liu, S., Orazio, C., Rodriguez, L., Neves Silva, L. & Wingfield, M. 2015. Changes in planted forests and future global implications. Forest Ecology and Management, 352: 57–67.
Pereira, H.M., Leadley, P.W., Proenca, V., Alkemada, R., Scharlemann, J.P.W., Fernandez-Manjarres, J.F., Araujo. M.B. et al. 2010. Scenarios for global biodiversity in the 21st century. Science, 330(6010): 1496–1501.
Peres, C.A., Thaise, E., Schietti, J., Desmoulieres, S.J.M. & Levi, T. 2016. Dispersal limitation induces long-term biomass collapse in overhunted Amazonian forests. PNAS, 113: 892–897.
Persha, L., Agrawal, A. & Chhatre, A. 2011. Social and ecological synergy: Local rulemaking, forest livelihoods, and biodiversity conservation. Science, 331(6024): 1606–1608.
Peters, C.M. 2000. Pre-Columbian silviculture and indigenous management of neotropical forests. In D.L. Lentz, ed. Imperfect balance: landscape transformations in the Pre-Columbian Americas, pp. 203–223. New York, USA, Columbia University Press.
Phalan, B., Onial, M., Balmford, A. & Green, R. 2011. Reconciling food production and biodiversity conservation: land sharing and land sparing compared. Science, 333(6047): 1289–1291.
Plumptre, A.J., Kayitare, A., Rainer, H., Gray, M., Munanura, I., Barakabuye, N., Asuma, S., Sivha, M. & Namara, A. 2004. The socio-economic status of people living near protected areas in the Central Albertine Rift. Albertine Rift Technical Reports, 4. Kampala, Albertine Rift Programme.
Polisar, J., de Thoisy, B., Rumiz, D., Dıaz Santos, F., Balas McNab, R., Garcia-Anleu, R., Ponce-Santizo, G., Arispe, R. & Venega, C. 2016. Using certified timber extraction to benefit jaguar and ecosystem conservation. Ambio, 46: 588–603.
Porter-Bolland, L., Ellis, E.A., Guariguata, M.R., Ruiz-Mallén, I., Negrete-Yankelevich, S. & Reyes-García, V. 2012. Community managed forests and forest protected areas: An assessment of their conservation effectiveness across the tropics. Forest Ecology and Management, 268: 6–17.
Potapov, P., Hansen, M.C., Laestadius, L, Turubanova, S., Yaroshenko, A., Thies, C., Smith, W. et al. 2017. The last frontiers of wilderness: Tracking loss of intact forest landscapes from 2000 to 2013. Science Advances, 3(1): e1600821 [online]. [Cited 5 January 2020]. DOI: 10.1126/sciadv.1600821
Poudel, J., Zhang, D. & Simon, B. 2019. Habitat conservation banking trends in the United States. Biodiversity and Conservation, 28(6): 1629–1646.
Poulsen, J.R., Clark, C.J. & Palmer, T.M. 2013. Ecological erosion of an Afrotropical forest and potential consequences for tree recruitment and forest biomass. Biological Conservation, 163: 122–130.
Powell, B., Hall, J. & Johns, T. 2011. Forest cover, use and dietary intake in the East Usambara Mountains, Tanzania. International Forestry Review, 13(3): 305–317.
Premauer J. & Berkes F. 2012. Makuira, Colombia: the cosmological centre of origin for the Wayúu people. In N. Dudley & S. Stolton, eds. Protected landscapes and wild biodiversity, p. 53–60. Gland, Switzerland, IUCN.
Premauer, J. & Berkes, F. 2015. A Pluralistic approach to protected area governance: Indigenous peoples and Makuira National Park. Ethnobiology and Conservation 4: 1–16.
Pretty, J. & Smith, D. 2004. Social capital in biodiversity conservation and management. Conservation Biology, 18(3): 631–638.
Price, R. 2017. Economic drivers and effects of the illegal wildlife trade in sub-Saharan Africa. K4D Helpdesk Report. Brighton, UK, IDS.
Rasolofoson, R.A., Hanauer, M.M., Pappinen, A., Fisher, B. & Ricketts, T.H. 2018. Impacts of forests on children’s diet in rural areas across 27 developing countries. Science Advances, 4(8): eaat2853 [online]. [Cited 5 January 2020]. DOI: 10.1126/sciadv.aat2853
Ratnam, W., Rajora, O.P., Finkeldey, R., Aravanopoulos, F., Bouvet, J.-M., Vaillancourt, R.E., Kanashiro, M., Fady, B., Tomita, M. & Vinson, C. 2014. Genetic effects of forest management practices: Global synthesis and perspectives. Forest Ecology and Management, 333: 52–65.
Redford, K.H. 1992. The empty forest. BioScience, 42: 412–422.
Redmond, I., Aldred, T., Jedamzik, K. & Westwood, M. 2006. Recipes for survival: controlling the bushmeat trade. London, Ape Alliance & World Society for the Protection of Animals.
Reed, J, van Vianen, J., Foli, S., Clendenning, J., Yang, K., MacDonald, M., Petrokofsky, G., Padoch, Ch., Sunderland, T. 2017. Trees for life: The ecosystem service contribution of trees to food production and livelihoods in the tropics. Forest Policy and Economics, 84: 62–71.
Reid, H. & Huq, S. 2005. Climate change-biodiversity and livelihood impacts. In C. Robledo, M. Kanninen & L. Pedroni, eds. Tropical forests and adaptation to climate change, pp. 57–70. Bogor, Indonesia, CIFOR.
Reij, C., Tappan, G. & Smale, M. 2009. Agroenvironmental transformation in the Sahel. Another kind of “Green Revolution”. IFPRI Discussion Paper 00914. Washington, DC, IFPRI.
Reimchen T.E. & Arbellay, E. 2019. Influence of spawning salmon on tree-ring width, isotopic nitrogen, and total nitrogen in old-growth Sitka spruce from coastal British Columbia. Canadian Journal of Forest Research, 49: 1078–1086.
Reimoser, F. 2000. Income from hunting in mountain forests of the Alps. In M.F. Price & N. Butt, eds. Forests in sustainable mountain development: a state of knowledge report for 2000, pp. 346–353. IUFRO Research Series No. 5. New York, CABI Publishing.
Repetto, R. 1992. Accounting for environmental assets. Scientific American, 266(6): 94–101.
Reyes-Garcia, V., Guèze, M., Luz, A.C., Paneque-Gálvez, J., Macía, M.J., Orta-Martínez, M., Pino, J. & Rubio-Campillo, X. 2013. Evidence of traditional knowledge loss among a contemporary indigenous society. Evolution and Human Behavior, 34(4): 249–257.
Ribeiro, M.C., Metzger, J.P., Martensen, A.C., Ponzoni, F.J. & Hirota, M.M. 2009. The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biological Conservation, 142(6): 1141–1153.
Ribot, J.C. 2002. Democratic decentralization of natural resources: institutionalizing popular participation. Washington, DC, WRI.
Ripple, W.J., Newsome, T.M., Wolf, C., Dirzo, R., Everatt, K.T., Galetti, M., Hayward, M.W. et al. 2015. Collapse of the world’s largest herbivores. Science Advances, 1: e1400103 [online]. [Cited 5 January 2020]. https://doi.org/10.1126/sciadv.1400103
Ripple, W.J., Abernethy, K., Betts, M.G., Chapron, G., Dirzo, R., Galetti, M., Levi, T. et al. 2016. Bushmeat hunting and extinction risk to the world’s mammals. Royal Society Open Science, 3: 160498 [online]. [Cited 5 January 2020]. https://doi.org/10.1098/rsos.160498
Ritchie, H., Roser, M., Mispy, J. & Ortiz-Ospina, E. 2018. SDG Tracker: Indicator 15.1.2. In: SDG Tracker [online]. Oxford, UK. [Cited 19 December 2019]. https://sdg-tracker.org/biodiversity#15.1.2
Rivers, M.C., Beech, E., Bazos, I., Bogunić, F., Buira, A., Caković, D., Carapeto, A. et al. 2019. European red list of trees. Cambridge, UK, IUCN.
RNZ. 2019. Calls to train a million UK volunteers to tackle invasive species. In: RNZ [online]. Wellington, New Zealand. [Cited 5 January 2020]. www.rnz.co.nz/news/world/401840/calls-to-train-a-million-uk-volunteers-to-tackle-invasive-species
Roberts, P. 2019. Tropical forests in prehistory, history, and modernity. Oxford, UK, Oxford University Press.
Rodas, A. & Stoian, D. 2015. Determinación de los beneficios socioeconómicos del aprovechamiento forestal percibidos por tres comunidades con concesiones comunitarias en el Petén, Guatemala. Report of the ADA Community Forestry Project in Mesoamerica. Petén, Guatemala, Bioversity International.
Rohr, J.R., Civitello, D.J., Halliday, F.W., Hudson, P.J., Lafferty, K.D., Wood, C.L. & Mordecai, E.A. 2019. Towards common ground in the biodiversity–disease debate. Nature Ecology & Evolution, 4: 24–33.
Rook, G.A. 2013. Regulation of the immune system by biodiversity from the natural environment: an ecosystem service essential to health. PNAS, 110(46): 18360–18367.
Roosevelt, A.C., Lima da Costa, M., Lopes Machado, C., Michab, M., Mercier, N., Valladas, H., Feathers, J. et al. 1996. Paleoindian cave dwellers in the Amazon: the peopling of the Americas. Science, 272(5260): 373–384.
Roper, B.B., Saunders, W.C. & Ojala, J.V. 2019. Did changes in western federal land management policies improve salmonid habitat in streams on public lands within the Interior Columbia River Basin? Environmental Monitoring and Assessment, 191:574 [online]. [Cited 5 January 2020]. https://doi.org/10.1007/s10661-019-7716-5
Rowland, D., Blackie, R.R., Powell, B., Djoudi, H., Vergles, E., Vinceti, B. & Ickowitz, A. 2015. Direct contributions of dry forests to nutrition: a review. International Forestry Review, 17(S2): 45–53.
Rowland, D., Ickowitz, A., Powell, B., Nasi, R. & Sunderland, T. 2017. Forest foods and healthy diets: quantifying the contributions. Environmental Conservation, 44(2): 102–114.
RRI. 2015. Protected areas and the land rights of indigenous peoples and local communities: current issues and future agenda. Washington, DC, RRI.
RSCN. 2018. Report on the benefits generated by local communities from DBR [in Arabic]. Amman, RSCN.
RSCN and Wild Jordan. 2017. Explore Dana: Jordan’s rift valley spectacular. Brochure. Amman, RSCN.
Ruf, F. & Zadi, H. 1998. Cocoa: from deforestation to reforestation. In: Smithsonian’s National Zoo & Biology Institute [online]. Washington, DC. [Cited 5 January 2020]. https://nationalzoo.si.edu/scbi/migratorybirds/research/cacao/ruf.cfm
Ruijs, A. & Vardon, M. 2019. Natural capital accounting for mainstreaming biodiversity in public policy making. In Vardon, M., Bass, S., and Ahlroth, S. eds. Natural Capital Accounting for Better Policy Decisions: Climate change and Biodiversity. Proceedings and Highlights of the 3rd Forum on Natural Capital Accounting for Better Policy Decisions, pp. 73–100. World Bank WAVES, Washington D.C.
Ruokolainen, L., Von Hertzen, L., Fyhrquist, N., Laatikainen, T., Lehtomäki, J., Auvinen, P. & Knip, M. 2015. Green areas around homes reduce atopic sensitization in children. Allergy, 70(2): 195–202.
Sabogal, C., Besacier, C. & McGuire, D. 2015. Forest and landscape restoration: concepts, approaches and challenges for implementation. Unasylva, 245: 3–10.
Sacande, M., Jøker, D., Dulloo, M.E. & Thomsen, K.A. eds. 2004. Comparative storage biology of tropical tree seeds. Rome, IPGRI.
Sachedina, H. & Nelson, F. 2012. The development of payments for ecosystem services as a community-based conservation strategy in East Africa. In J. Ingram, F. DeClerck & C. Rumbaitis del Rio, eds. Integrating ecology and poverty reduction: the application of ecology in development solutions, pp. 149–171. New York, USA, Springer.
Sarkar, D., Walker-Swaney, J. & Shetty, K. 2019. Food diversity and indigenous food systems to combat diet-linked chronic diseases. Current Developments in Nutrition, nzz099 [online]. [Cited 5 January 2020]. https://doi.org/10.1093/cdn/nzz099
Sassen, M. 2014. Conservation in a crowded place: forest and people on Mount Elgon, Uganda. Wageningen University. (PhD thesis)
Sassen, M., Arnell, A.P. & van Soesbergen, A. forthcoming. Mapping risks to biodiversity and ecosystem services from cocoa-driven deforestation in West Africa.
Sassen, M., Sheil, D., Giller, K.E. & ter Braak, C.J. 2013. Complex contexts and dynamic drivers: understanding four decades of forest loss and recovery in an East African protected area. Biological Conservation, 159: 257–268.
Saunders, C.D., Brook, A.T. & Meyers, O.E. 2006. Using psychology to save biodiversity and human well-being. Conservation Biology, 20: 702–705.
Save the Elephants. 2019. Welcome to The Elephants and Bees Project [online]. Nairobi. [Cited 5 January 2020]. https://elephantsandbees.com
Sayer, J.A., Campbell, B., Petheram, L., Aldrich, M., Perez, M., Endamana, D., Nzooh Dongmo, Z.-L. et al. 2007. Assessing environment and development outcomes in conservation landscapes. Biodiversity and Conservation, 16(9), 2677–2694.
Sayer, J.A., Margules, C., Boedhihartono, A.K., Sunderland, T., Langston, J.D., Reed, J., Riggs, R. et al. 2017. Measuring the effectiveness of landscape approaches to conservation and development. Sustainability Science, 12: 465–476.
Schelley, C., Cross, J.E., Franzen, W.S., Hall, P. & Reeve, S. 2012. How to go green: creating a conservation culture in a public high school through education, modelling, and communication. Journal of Environmental Education, 43(3): 143–161.
Schroth, G., Harvey, C.A., da Fonseca, G.A., Vasconcelos, H.L., Gascon, C. & Izac, A.M.N. eds. 2004. Agroforestry and biodiversity conservation in tropical landscapes. Washington, DC, Island Press.
Schroth, G., Läderach, P., Martinez-Valle, A.I., Bunn, C. & Jassogne, L. 2016. Vulnerability to climate change of cocoa in West Africa: Patterns, opportunities and limits to adaptation. Science of the Total Environment, 556: 231–241.
Schueler, S., Falk, W., Koskela, J., Lefèvre, F., Bozzano, M., Hubert, J., Kraigher, H., Longauer, R. & Olrik, D.C. 2014. Vulnerability of dynamic genetic conservation units of forest trees in Europe to climate change. Global Change Biology, 20: 1498–1511.
Schulp, C.J., Thuiller, W. & Verburg, P.H. 2014. Wild food in Europe: A synthesis of knowledge and data of terrestrial wild food as an ecosystem service. Ecological Economics, 105: 292–305.
Schuster, R., Germain, R.R., Bennett, J.R., Reo, N.J. & Arcese, P. 2019. Vertebrate biodiversity on indigenous-managed lands in Australia, Brazil, and Canada equals that in protected areas. Environmental Science and Policy, 101: 1–6.
Schweik, C.M. 2000. Optimal foraging, institutions and forest change: A case from Nepal. Environmental Monitoring and Assessment, 62: 231–260.
SEGeF. 2018. Suivi de la gestion de la faune dans les forêts de production [online]. Yaoundé. [Cited 13 January 2020]. http://151.236.37.239/segef/public/
Shackleton, S., Paumgarten, F., Kassa, H., Husselman, M. & Zida, M. 2011. Opportunities for enhancing poor women’s socioeconomic empowerment in the value chains of three African non-timber forest products (NTFPs). International Forestry Review, 13(2): 136–151.
Shaffer, L.J., Khadka, K.K., Van Den Hoek, J. & Naithani, K.J. 2019. Human-elephant conflict: a review of current management strategies and future directions. Frontiers in Ecology and Evolution, 6: 235 [online]. [Cited 5 January 2020]. https://doi.org/10.3389/fevo.2018.00235
Shanahan, D.F., Lin, B.B., Bush, R., Gaston, K.J., Dean, J.H., Barber, E. & Fuller, R.A. 2015. Towards improved public health outcomes from urban nature. American Journal of Public Health, 105: 470–477.
Sharpe, B. 1998. First the forest: conservation, community and participation in south-west Cameroon. Africa, 68(1): 25–45.
Shisegar, N. 2014. The impact of green areas on mitigating urban heat island effect: a review. International Journal of Environmental Sustainability, 9: 119–130.
Sichuan Forestry Department. 2015. The Pandas of Sichuan: The 4th Survey Report on Giant Panda in Sichuan Province. Chengdu, China, Sichuan Science and Technology Press. Cited by Brinckmann, J.A., Luo W., Xu Q., He X., Wu J., & Cunningham A.B. 2018. Sustainable harvest, people and pandas: Assessing a decade of managed wild harvest and trade in Schisandra sphenanthera. Journal of Ethnopharmacology, 224: 522–534.
Silva, L.N., Freer-Smith, P. & Madsen, P. 2019. Production, restoration, mitigation: a new generation of plantations. New Forests, 50(2): 153–168.
Sinovas, P., Price, B., King, E., Hinsley, A. & Pavitt, A. 2017. Wildlife trade in the Amazon countries: an analysis of trade in CITES listed species. Technical report prepared for the Amazon Regional Program (BMZ/DGIS/GIZ). Cambridge, UK, UNEP-WCMC.
Sirén, A. & Machoa, J. 2008. Fish, wildlife, and human nutrition in tropical forests: a fat gap? Interciencia, 33: 186–193.
Skole, D. & Tucker, C.J. 1993. Tropical deforestation and habitat fragmentation in the Amazon: satellite data from 1978 to 1988. Science, 260(5116): 1905–1910.
Soares-Filho, B., Moutinho, P., Nepstad, D., Anderson, A., Rodrigues, H., Garcia, R., Dietzsch,L. et al. 2010. Role of Brazilian Amazon protected areas in climate change mitigation. PNAS, 107(24): 10821–10826.
Solecki, R. 1975. Shanidar IV, a Neanderthal flower burial in northern Iraq. Science, 190(4217): 880–881.
Song, X.P., Hansen, M.C., Stehman, S.V., Potapov, P.V., Tyukavina, A., Vermote, E.F. & Townshend, J.R. 2018. Global land change from 1982 to 2016. Nature, 560: 639–643.
Southworth, J., Nagendra, H. & Munroe, D.K. 2006. Introduction to the Special Issue: Are parks working? Exploring human-environment tradeoffs in protected area conservation. Applied Geography, 26(2): 87–95.
Spies, T.A., Stine, P.A., Gravenmier, R., Long, J.W., Reilly, M.J., tech. coords. 2018. Synthesis of science to inform land management within the Northwest Forest Plan area. 3 volumes. General Technical Report PNW-GTR-966. Portland, OR, USA, US Department of Agriculture, Forest Service, Pacific Northwest Research Station.
Stanturf, J., Mansourian, S. & Kleine, M., eds. 2017. Implementing forest landscape restoration: A practitioner’s guide. Vienna, IUFRO-SPDC.
Stanturf, J.A., Palik, B.J. & Dumroese, R.K. 2014. Contemporary forest restoration: a review emphasizing function. Forest Ecology and Management, 331: 292–323.
St. John, F.A.V, Edwards-Jones, G. & Jones, J.P.G. 2010. Conservation and human behaviour: lessons from social psychology. Wildlife Research, 37: 658–667.
Stattersfield, A.J., Crosby, M.J., Long, A.J., and Wege, D.C. 1998. Endemic bird areas of the world: priorities for biodiversity conservation. Cambridge, UK, BirdLife International.
Stavert, J.R., Pattemore, D.E., Gaskett, A.C., Beggs, J.R. & Bartomeus, I. 2007. Exotic species enhance response diversity to land-use change but modify functional composition. Proceedings of the Royal Society B – Biological Sciences, 284(1860): 20170788 [online]. [Cited 13 January 2020]. https://doi.org/10.1098/rspb.2017.0788
Steffen, W., Richardson, K., Rockström, J., Cornell, S.E., Fetzer, I., Bennett, E.M., Biggs, R. et al. 2015. Planetary boundaries: Guiding human development on a changing planet. Science, 347(6223): 1259855 [online]. [Cited 5 January 2020]. https://doi.org/10.1126/science.1259855
Stevens, C., Winterbottom, R., Springer, J. & Reytar, K. 2014. Securing rights, combating climate change: How strengthening community forest rights mitigates climate change. Washington, DC, WRI.
Stoian, D. & Rodas, A. 2018. Successful community stewardship of tropical forests: evidence from community forest concessions in Petén, Guatemala. Paper presented at the 19th Annual Conference on Land and Poverty held by the World Bank in Washington DC on March 19–23, 2018 [online]. [Cited 5 January 2020]. https://cgspace.cgiar.org/bitstream/handle/10568/93439/Successful_Stoian_2018.pdf?sequence=1
Stoian, D., Rodas, A., Butler, M., Monterroso, I. & Hodgdon, B. 2018. Forest concessions in Petén, Guatemala: A systematic analysis of the socioeconomic performance of community enterprises in the Maya Biosphere Reserve. Bogor, Indonesia, CIFOR.
Stolton, S., Redford, K.H., Dudley, N., Bill, W., Corcuera, E. & Mitchel, B.A. 2014. The futures of privately protected areas. Gland, Switzerland, IUCN.
Strassburg, B., Beyer, H.L., Crouzeilles, R. Iribarrem, A., Barros, F., Siqueira, M., Sánchez-Tapia, A. et al. 2019. Strategic approaches to restoring ecosystems can triple conservation gains and halve costs. Nature Ecology & Evolution, 3: 62–70.
Sunderland, T., Sunderland-Groves, J., Shanley, P. & Campbell, B. 2009. Bridging the gap: how can information access and exchange between conservation biologists and field practitioners be improved for better conservation outcomes? Biotropica, 41(5): 549–554.
Sunderlin, W.D., Angelsen, A., Belcher, B., Burgers, P., Nasi, R., Santoso, L. & Wunder, S. 2005. Livelihoods, forest, and conservation in developing countries: an overview. World Development, 33(9): 1383–1402.
Tamosiunas, A., Gražulevičienė, R., Luksiene, D., Dedele, A., Reklaitiene, R., Baceviciene, M., & Milinaviciene, E. 2014. Accessibility and use of urban green spaces, and cardiovascular health: findings from a Kaunas cohort study. Environmental Health, 13: 20 [online]. [Cited 5 January 2020]. https://doi.org/10.1186/1476-069X-13-20
Taniwaki, R.H., Leal, C.G., de Barros Ferraz, S.F., Henrikson, L., Jägrud, L. & de Paula, F.R. 2018. Blue Targeting Tool: a simple forestry planning for riparian buffer zones adapted to Brazilian streams. Poster presented at the Joint Conference on Forests and Water, 2018, Valdivia, Chile. [also available at https://www.researchgate.net/publication/329102135_Blue_Targeting_Tool_a_simple_forestry_planning_for_riparian_buffer_zones_adapted_to_Brazilian_streams].
Tauli-Corpuz, V., Alcorn, J. & Molnar, A. 2018. Cornered by protected areas: replacing ‘fortress’ conservation with rights-based approaches helps bring justice for indigenous peoples and local communities, reduces conflict, and enables cost-effective conservation and climate change. Washington, DC, RRI.
The Guardian. 2020. A rewilding triumph: wolves help to reverse Yellowstone degradation. In: The Guardian [online]. [Cited 15 January 2020]. https://www.theguardian.com/environment/2020/jan/25/yellowstone-wolf-project-25th-anniversary
Tibesigwa, B., Siikamäki, J., Lokina R. & Alvsilver J. 2019. Naturally available wild pollination services have economic value for nature dependent smallholder crop farms in Tanzania. Scientific Reports, 9: 3434 [online]. [Cited 5 January 2020]. https://doi.org/10.1038/s41598-019-39745-7
TNC. 2019. Tropical Forest Conservation Act. Benefits for Natural Resources and the American People. In: The Nature Conservancy [online]. [Cited 15 February 2020]. https://www.nature.org/en-us/about-us/who-we-are/how-we-work/policy/tropical-forest-conservation-act/
Tracewski, Ł., Butchart, S.H.M., Donald, P.F., Evans, M., Fishpool, L.D.C. & Buchanan, G.M. 2016. Patterns of twenty-first century forest loss across a global network of important sites for biodiversity. Remote Sensing in Ecology and Conservation, 2(1): 37–44.
TRAFFIC. 2019. African elephants: elephant conservation and the global trade in ivory. In: TRAFFIC [online]. Cambridge, UK. [Cited 5 January 2020]. www.traffic.org/what-we-do/species/elephants-ivory
Triguero-Mas, M., Dadvand, P., Cirach, M., Martínez, D., Medina, A., Mompart, A., Basagaña, X., Gražulevičienė, R. & Nieuwenhuijsen, M.J. 2015. Natural outdoor environments and mental and physical health: relationships and mechanisms. Environment International, 77, 35–41.
Tropical Forest Alliance. 2017. The Role of the Financial Sector in Deforestation-Free Supply Chains. Tropical Forest Alliance and World Economic Forum, Geneva. [also available at https://www.vivideconomics.com/wp-content/uploads/2019/08/TFA2020_Framing_Paper_030117.pdf].
Turner, I. 1996. Species loss in fragments of tropical rain forest: a review of the evidence. Journal of Applied Ecology, 33: 200–209.
Turner, B.L. & Sabloff, J.A. 2012. Classic Period collapse of the Central Maya Lowlands: Insights about human–environment relationships for sustainability. PNAS, 109(35):13908–13914.
UAESPNN. 2005. Plan de manejo Parque Nacional Natural Macuira 2005–2009 [National Natural Park Makuira, Management Plan 2005–2009]. Bogota, Parques Nacionales Naturales de Colombia.
Udawatta, R.P., Rankoth, L.M. & Jose, S. 2019. Agroforestry and biodiversity. Sustainability, 11(10): 2879 [online]. [Cited 5 January 2020]. https://doi.org/10.3390/su11102879
UN. 1992a. United Nations Convention on Biological Diversity. New York, USA. [also available at https://www.cbd.int/doc/legal/cbd-en.pdf].
UN. 1992b. United Nations Framework Convention on Climate Change. New York, USA. [also available at https://unfccc.int/resource/docs/convkp/conveng.pdf].
UN. 1992c. United Nations Convention on to Combat Desertification. New York, USA. [also available at https://www.unccd.int/sites/default/files/relevant-links/2017-01/UNCCD_Convention_ENG_0.pdf]
UN. 2008a. United Nations Declaration on the Rights of Indigenous Peoples. New York, USA. [also available at https://www.un.org/esa/socdev/unpfii/documents/DRIPS_en.pdf].
UN. 2008b. World urbanization prospects: The 2007 revision. New York, USA.
UN. 2015. Paris Agreement. New York, USA. [also available at https://unfccc.int/files/essential_background/convention/application/pdf/english_paris_agreement.pdf].
UN. 2017a. United Nations Strategic Plan for Forests 2017–2030. In: United Nations Department of Economic and Social Affairs – Forests [online]. New York, USA. [Cited 5 January 2020]. www.un.org/esa/forests/documents/un-strategic-plan-for-forests-2030/index.html
UN. 2017b. New York Declaration on Forests (list of endorsers updated in July 2017). New York, USA. [also available at https://www.undp.org/content/dam/undp/library/Environment%20and%20Energy/Forests/New%20York%20Declaration%20on%20Forests_DAA.pdf].
UN. 2019a. The Sustainable Development Goals Report 2019. New York, USA.
UN. 2019b. General Assembly. Seventy-third Session. 107th plenary meeting, Monday, 16 September 2019, 10 a.m., New York. A/73/PV.107. New York, USA. [also available at https://undocs.org/en/A/73/PV.107].
UN. 2020. SDG indicators: Metadata repository. In: United Nations, Department of Economic and Social Affairs, Statistics Division [online]. New York, USA. [Cited 5 January 2020]. https://unstats.un.org/sdgs/metadata
UN, European Commission, FAO, IMF, et al. 2014a. System of Environmental Economic Accounting 2012 — Central Framework. New York. [also available at http://unstats.un.org/unsd/envaccounting/seeaRev/SEEA_CF_Final_en.pdf]
UN, European Commission, FAO, OECD, et al. 2014b. System of Environmental Economic Accounting 2012 — Experimental Ecosystem Accounting. New York. [also available at http://unstats.un.org/unsd/envaccounting/seeaRev/eea_final_en.pdf]
United Nations General Assembly. 2008. 62/98. Non-legally binding instrument on all types of forests. Resolution adopted by the General Assembly on 17 December 2007. A/RES/62/98. New York, USA. [also available at https://undocs.org/en/A/RES/62/98].
United Nations General Assembly. 2015a. Transforming our world: the 2030 Agenda for Sustainable Development. Resolution adopted by the General Assembly on 25 September 2015. A/RES/70/1. New York, USA. [also available at https://undocs.org/en/A/RES/70/1].
United Nations General Assembly. 2015b. Tackling illicit trafficking in wildlife. Resolution adopted by the General Assembly on 30 July 2015. A/RES/69/314. New York, USA. [also available at https://undocs.org/en/A/RES/69/314].
UNCCD. n.d. The LDN Fund – An impact investment fund for land degradation neutrality. In: United Nations Convention to Combat Desertification [online]. Bonn, Germany. [Cited 2 January 2020]. https://www.unccd.int/actions/impact-investment-fund-land-degradation-neutrality
UNCCD. 2018. Decision 7/COP.13. The future strategic framework of the Convention. Bonn, Germany. [also available at https://www.unccd.int/sites/default/files/relevant-links/2018-08/cop21add1_SF_EN.pdf].
UNCCD. 2019a. The LDN target setting programme. In: United Nations Convention to Combat Desertification [online]. Bonn, Germany. [Cited 5 January 2020]. www.unccd.int/actions/ldn-target-setting-programme
UNCCD. 2019b. The GGW aims to restore Africa’s degraded landscapes and transform millions of lives in one of the world’s poorest regions. In: United Nations Convention to Combat Desertification [online]. Bonn, Germany. [Cited 5 January 2020]. https://knowledge.unccd.int/publications/ggw-aims-restore-africas-degraded-landscapes-and-transform-millions-lives-one-worlds
UNCTAD. 2006. International Tropical Timber Agreement, 2006. TD/TIMBER.3/12. Geneva, Switzerland. [also available at https://treaties.un.org/doc/source/docs/tdtimber3d12_en.pdf].
UNDESA. 2016. Documents – UN forest instrument. In: United Nations Department of Economic and Social Affairs [online]. [Cited 18 December 2019]. https://www.un.org/esa/forests/documents/un-forest-instrument/index.html
UNDP. 2017. What is biodiversity finance? In: United Nations Development Programme – BIOFIN – The Biodiversity Finance Initiative [online]. New York, USA. [Cited 5 January 2020]. www.biodiversityfinance.net/about-biofin/what-biodiversity-finance
UNECE & FAO. 2018. Forests and Water. Valuation and payments for forest ecosystem services. Geneva. [also available at https://www.unece.org/fileadmin/DAM/timber/publications/sp-44-forests-water-web.pdf]
UNEP. 1979. Convention on the Conservation of Migratory Species of Wild Animals. Nairobi. [also available at https://www.cms.int/sites/default/files/instrument/CMS-text.en_.PDF].
UNEP. 2019. Global environment outlook GEO-6. Summary for policy makers. Cambridge, UK, Cambridge University Press.
UNEP-WCMC. 2007. A spatial analysis approach to the global delineation of dryland areas of relevance to the CBD Programme of Work on Dry and Subhumid Lands. Cambridge, UK.
UNEP-WCMC. 2020. Welcome to the global ICCA Registry [online]. Cambridge, UK. [Cited 5 January 2020]. http://www.iccaregistry.org/
UNEP-WCMC & IUCN. 2019. World Database on Protected Areas. In: Protected Planet [online]. Cambridge, UK. [Cited 31 December 2019]. https://www.protectedplanet.net/c/world-database-on-protected-areas
UNEP-WCMC, IUCN & NGS. 2020. Protected Planet Digital Report [online]. Cambridge, UK, Gland, Switzerland and Washington, DC. [Cited 18 December 2019] https://livereport.protectedplanet.net
UNEP-WCMC & UNSD. 2019. Assessing the linkages between global indicator initiatives, SEEA Modules and the SDG Targets. Working Document. [also available at https://seea.un.org/sites/seea.un.org/files/seea_global_indicator_review_methodological_note_post_workshop_0.pdf]
UNESCO. 1971. Convention on wetlands of international importance especially as waterfowl habitat. Paris. [also available at https://treaties.un.org/doc/Publication/UNTS/Volume%20996/volume-996-I-14583-English.pdf].
UNFCCC. 2011. Report of the Conference of the Parties on its 16th session, Cancun, Mexico, 29 November – 10 December 2010. Addendum: Part Two: Action taken by the Conference of the Parties at its sixteenth session. Decision 1/CP.16. The Cancun Agreements: Outcome of the work of the Ad Hoc Working Group on Long-term Cooperative Action under the Convention. FCCC/CP/2010/7/Add.1. Bonn, Germany. [also available at https://unfccc.int/resource/docs/2010/cop16/eng/07a01.pdf].
UNODC. 2016. World wildlife crime report: Trafficking in protected species 2016. Vienna.
USAID. 2017. Countries with TFCA Programs. In: UNAID [online]. Washington, DC. [Cited 2 January 2020]. https://www.usaid.gov/biodiversity/TFCA/programs-by-country
USDA. n.d.a. Northwest Forest Plan. In: United States Department of Agriculture, Forest Service [online]. Washington, DC. [Cited 2 January 2020]. https://www.fs.usda.gov/detail/r6/landmanagement/planning/?cid=fsbdev2_026990
USDA. n.d.b. 5022: Wild crop harvesting. In: United States Department of Agriculture, Agricultural Marketing Service [online]. Washington, DC. [Cited 2 January 2020]. https://www.ams.usda.gov/rules-regulations/organic/handbook/5022
US Fish & Wildlife Service. 1998. Recovery plan for the Oregon chub (Oregonichthys crameri). Portland, OR, USA.
US Fish & Wildlife Service. 2018. North American Model of Wildlife Conservation. In: US Fish & Wildlife Service, Hunting [online]. Washington, DC. [Cited 1 January 2020]. https://www.fws.gov/hunting/north-american-model-of-wildlife-conservation.html
US/ICOMOS. 2019. Heritage trees: international research and registries. In: US/ICOMOS [online]. Washington, DC. [Cited 5 January 2020]. https://usicomos.org/heritage-trees-international-research-and-registries/
Uusivuori, J., Lehto, E. & Palo, M. 2002. Population, income and ecological conditions as determinants of forest area variation in the tropics. Global Environmental Change, 12: 313–323.
Valencia, R., Balslev, H. & Paz y Miño, G.C. 1994. High tree alpha-diversity in Amazonian Ecuador. Biodiversity & Conservation, 3:21– 28, cited by Dirzo, R. & Raven, P. H. 2003. Global state of biodiversity and loss. Annual Review of Environment and Resources, 28: 137–167.
van Lierop, P., Lindquist, E., Sathyapala, S. & Franceschini, G. 2015. Global forest area disturbance from fire, insect pests, disease and severe weather events. Forest Ecology and Management 352: 78–88.
Van Vliet, N., Muhindo, J., Nyumu, J.K. & Nasi, R. 2019. From the forest to the dish: A comprehensive study of the wildmeat value chain in Yangambi, Democratic Republic of Congo. Frontiers in Ecology and Evolution, 7: 132 [online]. [Cited 5 January 2020]. https://doi.org/10.3389/fevo.2019.00132
Verdone, M. & Seidl, A. 2017. Time, space, place, and the Bonn Challenge global forest restoration target. Restoration Ecology, 25(6): 903–911. [also available at http://dx.doi.org/10.1111/rec.12512]
Verissimo, D. 2013. Influencing human behaviour: an underutilised tool for biodiversity management. Conservation Evidence, 10: 29–31.
Verschuuren, B. & Brown, S. eds. 2018. Cultural and spiritual significance of nature in protected areas: Governance, management and policy. Abingdon, UK, Routledge.
Vié, J.-C., Hilton-Taylor, C. & Stuart, S.N. eds. 2009. Wildlife in a changing world: an analysis of the 2008 IUCN Red List of Threatened Species. Gland, Switzerland, IUCN.
Vlam, M., van der Sleen, P., Groenendijk, P. & Zuidema, P.A. 2017. Tree age distributions reveal large-scale disturbance-recovery cycles in three tropical forests. Frontiers in Plant Science, 7: 1984 [online]. [Cited 5 January 2020]. https://doi.org/10.3389/fpls.2016.01984
Vogt, P. 2019a. Image object Accounting (Available in the free JRC software GuildosToolbox). Ispra, Italy, European Commission Joint Research Center, Directorate for Sustainable Resources. [also available at https://ies-ows.jrc.ec.europa.eu/gtb/GTB/psheets/GTB-Objects-Accounting.pdf].
Vogt, P. 2019b. Measuring Forest Area Density to quantify forest fragmentation. (Available in the free JRC software GuidosToolbox). Ispra, Italy, European Commission Joint Research Center, Directorate for Sustainable Resources. [also available at https://ies-ows.jrc.ec.europa.eu/gtb/GTB/psheets/GTB-Fragmentation-FADFOS.pdf.
Vogt, P., Riitters, K.H., Caudullo, G., Eckhardt, B. 2019. FAO – State of the World’s Forests: Forest fragmentation. JRC Technical Report, EUR 29972 EN. Luxembourg, Publications Office of the European Union. [also available at https://publications.jrc.ec.europa.eu/repository/bitstream/JRC118594/technicalreport_fao_frag.pdf].
Vorontsova, M.S., Clark, L.G., Dransfield, J., Govaerts, R. & Baker, W.J. 2016. World checklist of bamboos and rattans. INBAR Technical Report No. 37, Beijing, INBAR.
Walker, X.J., Baltzer, J.L., Cumming, S.G., Day, N.J., Ebert, C., Goetz, S., Johnstone, J.F. et al. 2019. Increasing wildfires threaten historic carbon sink of boreal forest soils. Nature, 572: 520–523.
Watson, E.E. 2005. Gender-sensitive natural resource management (NRM) research-for-development. DFID NRSP Programme Development Report PD123: Gender Sensitive Research for Development. Cambridge, UK, University of Cambridge, Department of Geography.
Watson, J.E.M., Dudley, N., Segan, D.B. & Hockins, M. 2014. The performance and potential of protected areas. Nature, 515: 67–73.
Watson, J.E.M., Evans, T., Venter, O., Williams, B., Tulloch, A., Stewart, C., Thompson, I. et al. 2018. The exceptional value of intact forest ecosystems. Nature Ecology & Evolution 2: 599–610.
WEF. 2020. One trillion trees – World Economic Forum launches plan to help nature and the climate. In: World Economic Forum [online]. Geneva, Switzerland. [Cited 15 February 2020]. https://www.weforum.org/agenda/2020/01/one-trillion-trees-world-economic-forum-launches-plan-to-help-nature-and-the-climate/
West, P., Igoe, J., & Brockington, D. 2006. Parks and peoples: the social impact of protected areas. Annual Review of Anthropology, 35: 251–277.
White, M.P., Alcock, I., Wheeler, B.W. & Depledge, M.H. 2013. Would you be happier living in a greener urban area? A fixed-effects analysis of panel data. Psychological Science, 24(6): 920–928.
WHO. 2002. WHO Traditional medicine strategy: 2002–2005. Geneva, Switzerland.
WHO. 2016. Ambient air pollution: a global assessment of exposure and burden of disease. Geneva, Switzerland.
WHO. 2017. 5. Annex 5. Guidelines for the production, control and regulation of snake antivenom immunoglobulins. Replacement of Annex 2 of WHO Technical Report Series, No. 964. Geneva, Switzerland.
WHO. 2018a. Household air pollution and health. In: World Health Organization [online]. Geneva, Switzerland. [Cited 5 January 2020]. www.who.int/news-room/fact-sheets/detail/household-air-pollution-and-health
WHO. 2018b. Air pollution: Maps and databases. In: World Health Organization [online]. Geneva, Switzerland. [Cited 5 January 2020]. www.who.int/airpollution/data/en
WHO. 2019. Traditional, complementary and integrative medicine: About us. In: World Health Organization [online]. Geneva, Switzerland. [Cited 5 January 2020]. www.who.int/traditional-complementary-integrative-medicine/about
WHO. 2020. Q&A on coronaviruses (COVID-19). In: World Health Organization [online]. Geneva, Switzerland. [Cited 1 April 2020]. www.who.int/news-room/q-a-detail/q-a-coronaviruses.
WHO/UNICEF. 2000. Global water supply and sanitation assessment 2000 report. Geneva, Switzerland, WHO/UNICEF Joint Monitoring Programme for Water Supply and Sanitation.
Wilcox, B.A. & Ellis, B. 2006. Forests and emerging infectious diseases of humans. Unasylva, 224: 11–18.
Wilkie, D.S., Wieland, M., Boulet, H., Le Bel, S., van Vliet, N., Cornelis, D., BriacWarnon, V., Nasi, R. & Fa, J.E. 2016. Eating and conserving bushmeat in Africa. African Journal of Ecology, 54: 402–414.
Willett, W., Rockström, J., Loken, B., Springmann, M., Lang, T., Vermeulen, S., Garnett, T. et al. 2019. Food in the Anthropocene: the EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet, 393(10170): 447–492.
Willis, K.J. ed. 2017. State of the World’s Plants 2017. Richmond, Surrey, Kew Publishing.
Willis, K.J. ed. 2018. State of the World’s Fungi 2018. Richmond, Surrey, Kew Publishing.
Winfree, R., Williams, N.M., Dushoff, J. & Kremen, C. 2007. Native bees provide insurance against ongoing honey bee losses. Ecology Letters, 10: 1105–1113.
Winfree, R., Aguilar, R., Vazquez, D.P., LeBuhn, G. & Aizen, M.A. 2009. A meta-analysis of bees’ responses to anthropogenic disturbance. Ecology, 90: 2068–2076.
Witt, K.A. 2013. The nutrient content of Moringa oleifera leaves. In: ECHO Community [online]. North Fort Myers, FL, USA. [Cited 5 January 2020]. https://www.echocommunity.org/resources/a7ee06e3-40f2-4ef0-859e-4e64b90a56c8
World Agroforestry. 2009. The Agroforestree Database. In: World Agroforestry [online]. Nairobi. [Cited 13 January 2020]. http://www.worldagroforestry.org/output/agroforestree-database
World Bank. 2002. A revised forest strategy for the World Bank Group. Washington, DC.
World Bank. 2017. Guidebook on Ecosystem Accounting. Washington, DC. [also available at https://elibrary.worldbank.org/doi/pdf/10.1596/29829].
World Bank. 2019. Global Wildlife Programme Phase 2: Summarized version of child projects [online]. [Cited 5 January 2020]. https://www.thegef.org/sites/default/files/webdocuments/10200_PFD_Wildlife_Annex_ChildProjects.pdf
World Cocoa Foundation. 2017. Cocoa & Forests Initiative: Statement of intent. In: World Cocoa Foundation [online]. Washington, DC. [Cited 5 January 2020]. www.worldcocoafoundation.org/cocoa-forests-initiative-statement-of-intent/
World Land Trust. n.d. Golden-headed lion tamarin. In: World Land Trust [online]. Halesworth, Suffolk, UK. [Cited 5 January 2020]. www.worldlandtrust.org/species/mammals/golden-headed-lion-tamarin
WWF. 2018. WWF Tanzania set to implement debt for nature swap programme. In: WWF [online]. [Cited 5 January 2020]. wwf.panda.org/?324230/WWF-Tanzania-Set-to-implement-Debt-for-Nature-Swap-Programme
WWF China. 2012. Standards for Giant Panda friendly products. Chengdu, China.
Yearsley. 2019. FairWild project in India is a win-win-win for Terminalia trees, people, and hornbills. HerbalEGram, 16(6) [online]. [Cited 5 January 2020]. http://cms.herbalgram.org/heg/volume16/06June/FairWildTerminalia.html
Zhang, D. & Pearse, P.H. 2011. Forest economics. Vancouver, UBC Press.
Zhu, H., Xu, Z.F., Wang, H. & Li, B.G. 2004. Tropical rain forest fragmentation and its ecological and species diversity changes in southern Yunnan. Biodiversity and Conservation, 13(7): 1355–1372.
Zomer, R.J., Trabucco, A., Coe, R. & Place, F. 2009. Trees on farm: analysis of global extent and geographical patterns of agroforestry. ICRAF Working Paper 89. Nairobi, Kenya, World Agroforestry Centre.
ZSL & WWF. 2014. Living Planet Index [online]. [Cited 26 December 2019]. http://www.livingplanetindex.org/home/index