C.D. Allen
Craig D. Allen trabaja para el Estudio Geológico de los Estados Unidos, Centro de Ciencia de Fort Collins, Estación de Campo Montañas de Jemez, Los Álamos, Nuevo México (Estados Unidos de América).
Una introducción a las pautas mundiales emergentes relativas a la muerte regresiva del bosque inducida por el clima.
La expansión demográfica y económica está causando, en muchas zonas de bosque de diversas regiones, transformaciones rápidas y directas. Menos evidentes resultan los efectos generalizados del cambio climático en curso en la situación de los bosques a través del mundo. Los bosques cubren en conjunto el 30 por ciento de la superficie de la Tierra (FAO, 2006). Se han documentado recientemente casos de muerte regresiva (definida aquí como la muerte de árboles en cantidades muy por encima de los normales niveles de mortalidad) relacionados con la sequía y el calor que llevan a constatar que este fenómeno obedece a pautas mundiales. El presente artículo ofrece una introducción a la comprensión de estas pautas y examina la posibilidad de que la muerte regresiva inducida por agentes climáticos suponga un factor de riesgo para muchos bosques y terrenos boscosos. Un artículo más exhaustivo (Allen et al., 2009) estudia este tema con mayor detalle.
Si bien los fenómenos climáticos –desde tempestades de hielo hasta tornados y huracanes– pueden ocasionar múltiples daños en los bosques, aquí se hace hincapié en el estrés hídrico impulsado por la sequía y las altas temperaturas.
Se reconoce que el clima de la Tierra está siendo afectado por importantes cambios de origen humano, y que las temperaturas mundiales medias sobrepasan en la actualidad los máximos históricos de los últimos 1 300 años (IPCC, 2007). En muchas regiones se pronostican cambios aún más pronunciados en los patrones climáticos durante las próximas décadas: temperaturas más elevadas y pautas de precipitación alteradas que harán variar la disponibilidad de agua para los vegetales.
El recalentamiento mundial, los cambios en la composición atmosférica (por ejemplo, el incremento de concentraciones de compuestos nitrogenados y de CO2 originados por el enorme volumen de emisiones provenientes de las actividades humanas) y los aumentos locales de luz solar y precipitaciones han favorecido el crecimiento de muchos bosques en el últimos decenios. Esto ha ocurrido en lugares y períodos en que había agua en abundancia, y se ha debido a que la mayor parte de los bosques del mundo se encuentra en zonas donde el crecimiento y la productividad de los árboles están limitados por la temperatura, la luz o los nutrientes (Boisvenue y Running, 2006).
Por otra parte, alrededor de un tercio de las tierras del mundo son en la actualidad demasiado secas para el crecimiento de los árboles. Muchas zonas arboladas y terrenos boscosos están localizados en zonas climáticas marginales, en las que la productividad neta de la vegetación primaria está muy limitada por el agua (Boisvenue y Running, 2006). En respuesta a la sequía o a las temperaturas más cálidas (por ejemplo, Peñuelas, Lloret y Montoya, 2001), los bosques de estas regiones semiáridas suelen registrar disminuciones marcadas del crecimiento y un aumento de la mortalidad; así también sucede con las especies de árboles que crecen en las márgenes más secas de su área de distribución (por ejemplo, Jump, Hunt y Peñuelas, 2006).
Sin embargo, en los bosques más húmedos del mundo, desde los húmedos tropicales hasta los boreales, el crecimiento y la mortalidad también son procesos biológicos muy sensibles a los efectos de la sequía (Clark, 2004; Nepstad et al., 2007; Soja et al., 2007). Tal y como se observó durante la sequía y ola de calor que asoló Europa en 2003, los bosques templados que crecen en lugares productivos pueden evidenciar marcadas disminuciones de crecimiento, altos niveles de mortalidad y efectos retardados debidos a estrés por sequía y calor extremos que se manifiestan a lo largo de muchos años (Ciais et al., 2005; Breda et al., 2006). Las altas temperaturas pueden, por sí solas e independientemente de las precipitaciones, aumentar el estrés hídrico del bosque (Barber, Juday y Finney, 2000; Angert et al., 2005), y no pareciera que ningún bosque del mundo está a salvo de los impactos de la sequía.
Como la mortalidad de los árboles se debe por lo general a factores múltiples e interrelacionados, que van de la sequía a las plagas de insectos y enfermedades, la determinación de una causa de mortalidad única sería poco realista. No obstante, los factores de estrés abiótico son frecuentemente la razón fundamental de las enfermedades de los bosques; y se considera que los factores de estrés ligados al clima juegan un rol preponderante en la aparición de muchos brotes graves de insectos y enfermedades forestales (Desprez-Loustau et al., 2006; Raffa et al., 2008).
El estrés hídrico inducido por el clima es causa indirecta de mortalidad de árboles debido a efectos agudos y de breve duración tales como la interrupción irreversible de la columna de agua presente en el interior del tronco y las hojas (cavitación). La resistencia y vulnerabilidad de las especies de árboles a la cavitación, factor clave de la resistencia a la sequía, es sumamente variable. Los árboles sometidos a estrés hídrico minimizan el riesgo de cavitación mediante la oclusión de los estomas; consiguen así reducir la pérdida de agua y la consiguiente tensión en el xilema. La oclusión estomática tiene su precio, ya que impide la difusión del CO2 hacia el follaje y limita la fotosíntesis. El estrés hídrico crónico y prolongado debilita y termina matando los árboles, ya sea directamente por privación de carbono, o indirectamente debido a los ataques de plagas, como el escarabajo del pino de montaña, que aniquilan las defensas ya disminuidas de estos árboles (McDowell et al., 2008). El clima afecta también directamente a la dinámica de las poblaciones de insectos forestales y patógenos fúngicos (por ejemplo, Hicke et al., 2006). Por consiguiente, algunos brotes masivos de insectos que matan árboles son atribuibles al cambio climático (Raffa et al., 2008). Independientemente del mecanismo que la determine, la muerte regresiva es a menudo un proceso no lineal; puede manifestarse abruptamente a escala regional, cuando las condiciones climáticas exceden el umbral fisiológico de tolerancia de la especie, o puede desencadenar brotes de plagas de insectos (Allen, 2007).
No pocos informes establecen un nexo entre el aumento de la mortalidad forestal y una combinación de condiciones climáticas muy secas y/o cálidas, tales como las sequías observadas en el trópico tras fenómenos de El Niño en 1988 y 1997-1998 particularmente intensos, el recalentamiento constante y la sequía generalizada que se han registrado en la zona occidental de América del Norte desde el decenio de 1990, y la ola de calor y la sequía excepcionales del verano de 2003 en la zona occidental de Europa.
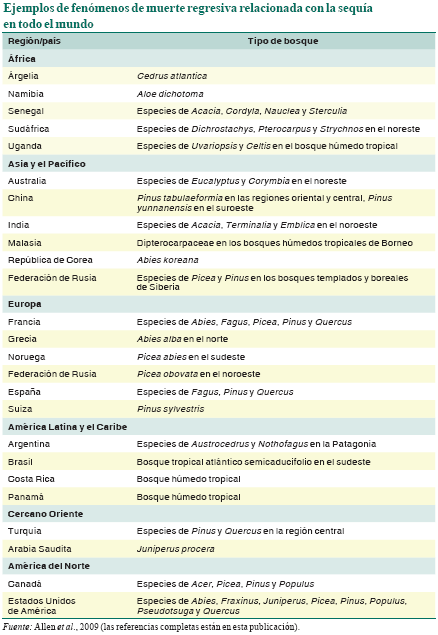
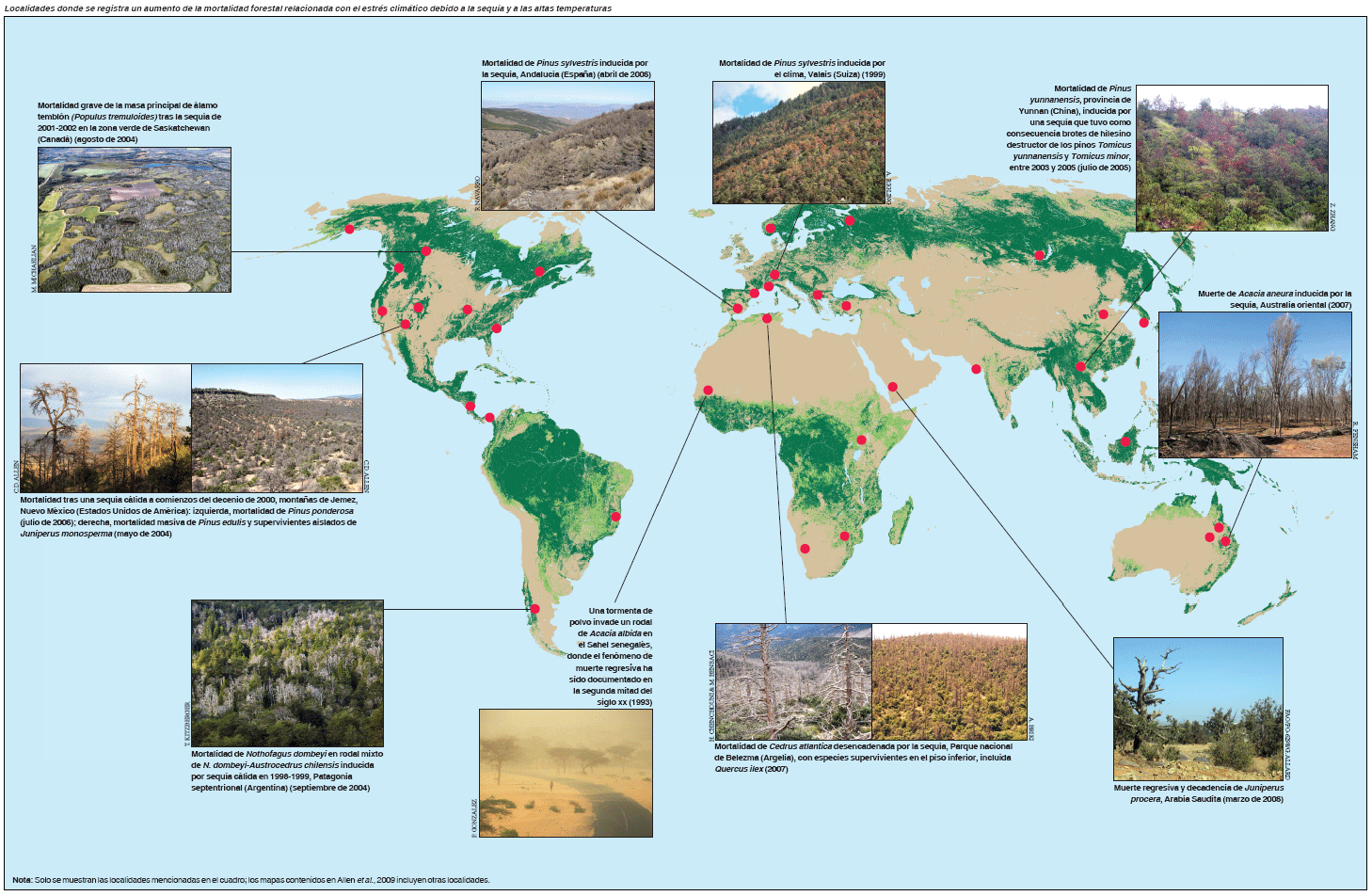
En los últimos tiempos, la mortalidad forestal asociada con la sequía se ha documentado en todos los continentes en los que hay tierras arboladas (véase la figura, pág. 46), y en diversos tipos de bosque y zonas climáticas. Por lo general, la muerte regresiva se observa en las cercanías del margen geográfico o altitudinal de un tipo de bosque o especie de árbol (Jump, Hunt y Peñuelas, 2006) y, probablemente, no lejos del umbral histórico de idoneidad climática donde cabría esperar la respuesta más sensible a las fluctuaciones climáticas.
En el cuadro se presentan algunos ejemplos de episodios de muerte regresiva impulsada por estrés hídrico y térmico derivado del clima desde 1970; los ejemplos se basan en un estudio mundial de más de 120 casos documentados (Allen et al., 2009). Aunque la muerte regresiva se observa comúnmente en regiones semiáridas donde el índice de crecimiento leñoso vegetal bordea el límite fisiológico de sequedad de las plantas leñosas (por ejemplo, Fensham, Fairfax y Ward, 2009), es evidente que la sequía inducida por el clima y el estrés térmico pueden ser causa potencial de muerte regresiva en muchos tipos de bosque y zonas arboladas en todo el mundo. Estos casos se han documentado con frecuencia en algunas zonas meridionales de Europa (Peñuelas, Lloret y Montoya, 2001; Breda et al., 2006) y en bosques templados o boreales de la zona occidental de América del Norte, donde, en los últimos decenios, las tasas de mortalidad inevitable han aumentado rápidamente (van Mantgem et al., 2009) y donde la muerte profusa de muchas especies en numerosos tipos de bosque ha afectado a más de 10 millones de hectáreas desde 1997 (Breshears et al., 2005; Raffa et al., 2008).
 |
Puesto que los árboles crecen de forma relativamente lenta pero pueden morir rápidamente, es sumamente importante evaluar el potencial y las consecuencias de un fenómeno de muerte regresiva generalizada inducido por el clima. Una sequía intensa puede matar en pocos meses o años árboles centenarios. Por consiguiente, la mortalidad forestal desencadenada por la sequía puede determinar en el ecosistema cambios veloces en zonas muy extensas. Dichos cambios tardan mucho menos en manifestarse que los que provienen de transiciones graduales debidas a la regeneración y al crecimiento. En el cuenca del Amazonas, por ejemplo, los impactos ligados al uso de la tierra, como las quemas antropógenas y la fragmentación forestal y los factores de estrés inducidos por el clima, que se influyen mutuamente, pueden verosímilmente, en algunas regiones, intensificar la muerte regresiva (Nepstad et al., 2008). Si la muerte regresiva masiva obligase a los actuales ecosistemas forestales a un reajuste abrupto, se registrarían efectos ecológicos y sociales omnipresentes y persistentes que se traducirían en pérdidas de productos forestales y servicios del ecosistema, y en especial modo la capacidad de absorción del carbono atmosférico.
Entre las consecuencias de la muerte regresiva intensa cabe mencionar la redistribución de los depósitos de carbono dentro de un ecosistema y una rápida liberación de carbono a la atmósfera. Por ejemplo, los efectos climáticos derivados de la muerte regresiva, de la mortalidad por insectos y enfermedades y del impacto del fuego han convertido en los últimos tiempos los bosques templados y boreales de Canadá, antes sumideros netos de carbono, en fuentes netas de carbono (Kurz et al., 2008). Análogamente, no sería imposible que, debido al colapso forestal generalizado por sequía, durante el presente siglo los bosques húmedos tropicales del mundo, hoy sumideros netos de carbono, se transformasen en una gran fuente neta de carbono (Lewis, 2005).
Ante los riesgos potenciales de muerte regresiva inducida por el clima, cabe desear que en las actuaciones de ordenación se prestase mayor atención a las opciones de adaptación destinadas a reforzar la resistencia y resiliencia de los bosques a los efectos pronosticados de estrés, por ejemplo mediante el raleo de los rodales con el objeto de reducir la competencia entre especies, la selección de genotipos diferentes (por ejemplo, de resistencia a la sequía) o la translocación de especies con el fin de su adaptación a los cambios climáticos augurados.
Forestales y expertos en ecología saben ya desde hace tiempo que el estrés climático tiene efectos considerables en la salud forestal. El interés que despierta la muerte regresiva inducida por el clima y las acciones de sensibilización que se han llevado a cabo al respecto no son recientes (Auclair, 1993; Ciesla y Donaubauer, 1994). Se conoce que la variación climática natural ha impulsado a lo largo de la historia episodios de mortalidad forestal generalizada (Swetnam y Betancourt, 1998). Entonces, ¿tienen lugar hoy sucesos diferentes? La Tierra experimenta sin lugar a dudas en nuestra época un cambio climático substancial y rápido, que tiene una orientación precisa. Este cambio ha sido avivado por grandes alteraciones antropógenas que afectan a la atmósfera, la superficie terrestre y las aguas (IPCC, 2007). Al mismo tiempo, el fenómeno de muerte regresiva inducida por el clima, va, según parece, en aumento en muchas partes del mundo. Aunque no se dispone aún de datos probatorios definitivos, es posible que los numerosos informes del fenómeno indiquen un incremento mundial de los problemas relacionados con la salud forestal y la muerte regresiva del bosque. A la luz de los problemas ya constatados bajo condiciones de un aumento relativamente contenido de la temperatura media mundial en tiempos recientes (unos 0,5 oC desde 1970) y un clima más seco en determinadas zonas (por ejemplo, Seager et al., 2007); y dado que se esperan temperaturas medias aún mayores (de alrededor de 2 a 4 oC a nivel mundial, y más en algunas regiones), y condiciones de secado durante períodos prolongados en ciertos lugares, cabe esperar, según las proyecciones, a partir de 2100, una intensificación de los factores de estrés forestal crónico y de riesgo de mortalidad (IPCC, 2007). Más allá de las variaciones en las condiciones climáticas promedio, otros cambios climáticos tales como sequías extremas, temperaturas máximas muy altas y olas de calor prolongadas –siempre más frecuentes y graves– (IPCC, 2007), podrían, según las proyecciones, exacerbar los episodios de muerte regresiva.
Las conclusiones acerca de las tendencias de mortalidad forestal y las predicciones relativas al fenómeno de muerte regresiva inducida por el clima en el futuro se ven limitadas por vacíos de información e investigaciones inciertas. En primer término, y pese a que se han realizado seguimientos forestales nacionales e incluso regionales, se carece aún de datos mundiales adecuados sobre salud forestal (FAO, 2006). Para determinar con exactitud la situación y tendencias mundiales del estrés forestal y la mortalidad, así como para entender las respuestas del ecosistema tras los acontecimientos de muerte regresiva, se necesita llevar a cabo un seguimiento de la salud forestal preciso, a largo plazo y a escala mundial, en el que datos de telepercepción y mediciones en el terreno se combinen.
En segundo lugar, solo se dispone hoy de datos cuantitativos del umbral fisiológico de mortalidad causada por estrés hídrico agudo para unas pocas especies de árboles (McDowell et al., 2008); en cambio, los datos conexos sobre sensibilidad térmica se desconocen casi por completo. Más aún, poco se sabe de las secuencias específicas del lugar y de los rangos de condiciones climáticas promedio y extremas capaces de desencadenar un fenómeno de mortalidad de árboles específicos en bosques situados en paisajes reales que pueda conducir potencialmente a la muerte regresiva generalizada de los individuos.
En tercer lugar, los investigadores ignoran aún los procesos de retroalimentación y las interacciones no lineales entre estrés forestal inducido por el clima y otros fenómenos de perturbación relacionados con el clima, tales como los brotes de insectos y el fuego, que son causa de mortalidad forestal muy difundida (Allen, 2007).
La incertidumbre científica acerca de los procesos fundamentales de mortalidad en árboles constituye una limitación básica para la realización de modelos cuantitativos más exactos de muerte regresiva futura inducida por el clima (por ejemplo, Huntingford et al., 2008). Así pues, estas carencias acortan también la posibilidad de predecir las consecuencias de la muerte regresiva en el potencial de secuestro de carbono atmosférico excedente por los bosques del mundo; o, en cambio, de predecir que los bosques se conviertan en fuentes de carbono y contribuyan a un cambio climático ampliado (Lucht et al., 2006).
En resumen, para aumentar la certidumbre científica de los riesgos de mortalidad futura inducida por el clima, y con el objeto de fundamentar las decisiones normativas y la ordenación forestal en todo el mundo, es indispensable intensificar el seguimiento de la salud forestal mundial y llevar a cabo nuevas investigaciones.
![]() Bibliografía
Bibliografía
Allen, C.D. 2007. Cross-scale interactions among forest dieback, fire, and erosion in northern New Mexico landscapes. Ecosystems, 10: 797–808.
Allen, C.D., Macalady, A., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., Gonzales, P., Hogg, T., Rigling, A., Breshears, D.D., Fensham, R., Zhang, Z., Kitzberger, T., Lim, J.-H., Castro, J., Running, S.W., Allard, G., Semerci, A. y Cobb, N. 2009. Climate-induced forest mortality: a global overview of emerging risks. Forest Ecology and Management (en revisión).
Angert, A., Biraud, S., Bonfils, C., Henning, C.C., Buermann, W., Pinzon, J., Tucker, C.J. y Fung. I. 2005. Drier summers cancel out the CO2 uptake enhancement induced by warmer springs. Proceedings of the NationalAcademy of Sciences of the United States of America, 102(31): 10823–10827.
Auclair, A.N.D. 1993. Extreme climatic fluctuations as a cause of forest dieback in the Pacific Rim. Water, Air and Soil Pollution, 66(3–4): 207–229.
Barber, V.A., Juday, G.P. y Finney, B.P. 2000. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature, 405: 668–673.
Boisvenue, C. y Running, S.W. 2006. Impacts of climate change on natural forest productivity – evidence since the middle of the 20th century. Global Change Biology, 12(5): 862–882.
Breda, N., Huc, R., Granier, A. y Dreyer, E. 2006. Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Annals of Forest Science, 63: 625–644.
Breshears, D.D., Cobb, N.S., Rich, P.M., Price, K.P., Allen, C.D., Balice, R.G., Romme, W.H., Kastens, J.H., Floyd, M.L., Belnap, J., Anderson, J.J., Myers, O.B. y Meyer, C.W. 2005. Regional vegetation die-off in response to global-change-type drought. Proceedings of the NationalAcademy of Sciences of the United States of America, 102(42): 15144–15148.
Ciais, P., Reichstein, M., Viovy, N., Granier, A., Ogee, J., Allard, V., Aubinet, M., Buchmann, N., Bernhofer, C., Carrara, A., Chevallier, F., De Noblet, N., Friend, A.D., Friedlingstein, P., Grunwald, T., Heinesch, B., Keronen, P., Knohl, A., Krinner, G., Loustau, D., Manca, G., Matteucci, G., Miglietta, F., Ourcival, J.M., Papale, D., Pilegaard, K., Rambal, S., Seufert, G., Soussana, J.F., Sanz, M.J., Schulze, E.D., Vesala, T. y Valentini, R. 2005. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature,437(7058): 529–533.
Ciesla, W.M. y Donaubauer, M.E. 1994. Decline and dieback of trees and forests: a global overview. FAO Forestry Paper No. 120. Roma, FAO.
Clark, D.A. 2004. Sources or sinks? The responses of tropical forests to current and future climate and atmospheric composition. Philosophical Transcripts of the Royal Society of London, Series B, 359: 477–491.
Desprez-Loustau, M.-L., Marcais, B., Nageleisen, L.-M., Piou, D. y Vannini, A. 2006. Interactive effects of drought and pathogens in forest trees. Annals of Forest Science, 63: 597–612.
FAO. 2006. Evaluación de los Recursos Forestales Mundiales 2005. Hacia la ordenación forestal sostenible. Estudio FAO: Montes 147. Roma.
Fensham, R.J., Fairfax, R.J. y Ward, D.P. 2009. Drought-induced tree death in savanna. Global Change Biology (en prensa).
Hicke, J.A., Logan, J.A., Powell, J. y Ojima, D.S. 2006. Changing temperatures influence suitability for modeled mountain pine beetle (Dendroctonus ponderosae) outbreaks in the western United States. Journal of Geophysical Research, 111: G02019.
Huntingford, C., Fisher, R.A., Mercado, L., Booth, B.B.B., Sitch, S., Harris, P.P., Cox, P.M., Jones, C.D., Betts, R.A., Malhi, Y., Harris, G., Collins, M. y Moorcroft, P. 2008. Towards quantifying uncertainty in predictions of Amazon “die-back”. Philosophical Transcripts of the Royal Society of London, Series B, 363(1498): 1857–1864.
Intergovernmental Panel on Climate Change (IPCC). 2007. Climate change 2007 –the physical science basis. Contribution of Working Group I to the Fourth Assessment Report of the IPCC. Cambridge, Reino Unido, Cambridge University Press.
Jump, A., Hunt, J.M. y Peñuelas, J. 2006. Rapid climate change-related growth decline at the southern range edge of Fagus sylvatica. Global Change Biology,12: 2163–2174.
Kurz, W.A., Stinson, G., Rampley, G.J., Dymond, C.C. y Neilson, E.T. 2008. Risk of natural disturbances makes future contribution of Canada’s forests to the global carbon cycle highly uncertain. Proceedings of the NationalAcademy of Sciences of the United States of America, 105: 1551–1555.
Lewis, S.L. 2005. Tropical forests and the changing earth system. Philosophical Transcripts of the Royal Society of London, Series B, 361: 195–210.
Lucht, W., Schaphoff, S., Erbrecht, T., Heyder, U. y Cramer, W. 2006. Terrestrial vegetation redistribution and carbon balance under climate change. Carbon Balance and Management, 1: 6.
McDowell, N., Pockman, W.T., Allen, C.D., Breshears, D.D., Cobb, N., Kolb, T., Sperry, J., West, A., Williams, D. y Yepez, E.A. 2008. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? Tansley Review. New Phytologist, 178: 719–739.
Nepstad, D.C., Tohver, I.M., Ray, D., Moutinho, P. y Cardinot, G. 2007. Mortality of large trees and lianas following experimental drought in an Amazon forest. Ecology,88(9): 2259–2269.
Nepstad, D.C., Stickler, C.M., Soares-Filho, B. y Merry, F. 2008. Interactions among Amazon land use, forests and climate: prospects for a near-term forest tipping point. Philosophical Transcripts of the Royal Society of London, Series B, 363(1498): 1737–1746.
Peñuelas, J., Lloret, F. y Montoya, R. 2001. Severe drought effects on Mediterranean woody flora in Spain. Forest Science, 47: 214–218.
Raffa, K.F., Aukema, B.H., Bentz, B.J., Carroll, A.L., Hicke, J.A., Turner, M.G. y Romme, W.H. 2008. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. Bioscience,58(6): 501–517.
Seager, R., Ting, M., Held, I., Kushnir, Y., Lu, J., Vecchi, G., Huang, H.-P., Harnik, N., Leetmaa, A., Lau, N.-C., Li, C., Velez, J. y Naik, N. 2007. Model projections of an imminent transition to a more arid climate in southwestern North America. Science, 316: 1181–1184.
Soja, A.J., Tchebakova, N.M., French, N.H.F., Flannigan, M.D., Shugart, H.H., Stocks, B.J., Sukhinin, A.I., Varfenova, E.I., Chapin F.S. y Stackhouse, P.W. Jr. 2007. Climate-induced boreal forest change: Predictions versus current observations. Global Planet. Change,56(3-4):274–296.
Swetnam, T.W. y Betancourt, J.L. 1998. Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest. Journal of Climate, 11: 3128–3147.
van Mantgem, P.J., Stephenson, N.L., Byrne, J.C., Daniels, L.D., Franklin, J.F., Fulé, P.Z., Harmon, M.E., Larson, A.J., Smith, J.M., Taylor, A.H. y Veblen, T.T. 2009. Widespread increase of tree mortality rates in the western United States. Science, 323: 521–524.