![]()
![]()
![]()
4.1 Land, people and domestic and wild herbivores
4.2 Landscapes, soils and vegetation
4.3 Climate
4.4 Rangeland production
4.5 Water resources
References
A knowledge of the land, livestock and people of the study area is needed to understand their interplay in shaping the livestock production system. Hence, this chapter deals with the biophysical environment (climate, grazing and water resources) of the study area as observed between 1981 and 1983. This is discussed further in the context of long-term trends in Chapter 10 (The long-term productivity of the Maasai livestock production system). Chapter 5 (The study area: Socio-spatial organisation and land use) centres on how people and their livestock use these resources.
This chapter begins with a general outline of the assets of the three group ranches in terms of land, people and livestock. This is followed by sections describing the landscape, soils and vegetation. The discussion of the characteristics of the climate, particularly rainfall, emphasises the differences between the semi-arid north and the arid south and the implications of these for the fodder resources and carrying capacity of the rangelands.
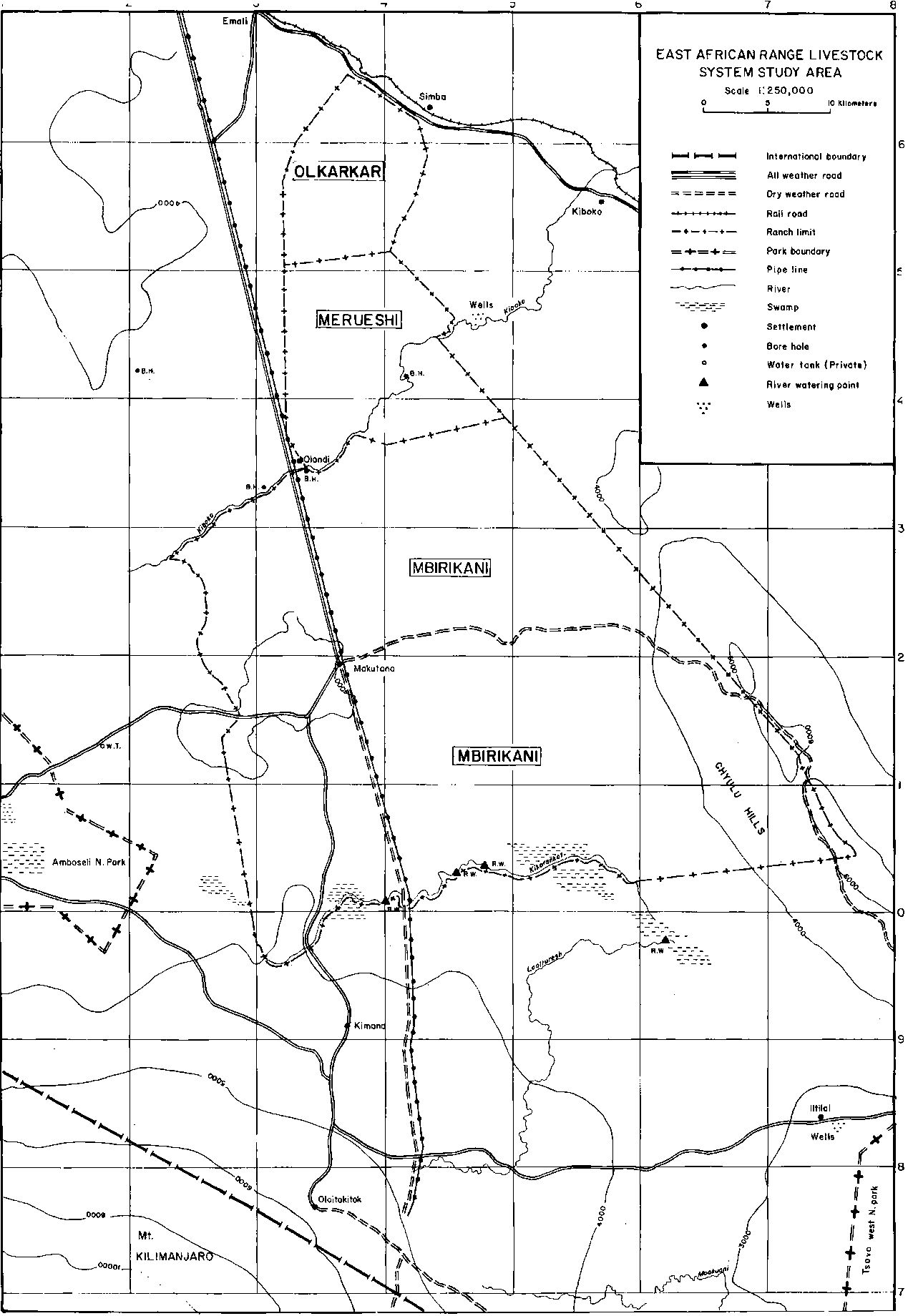
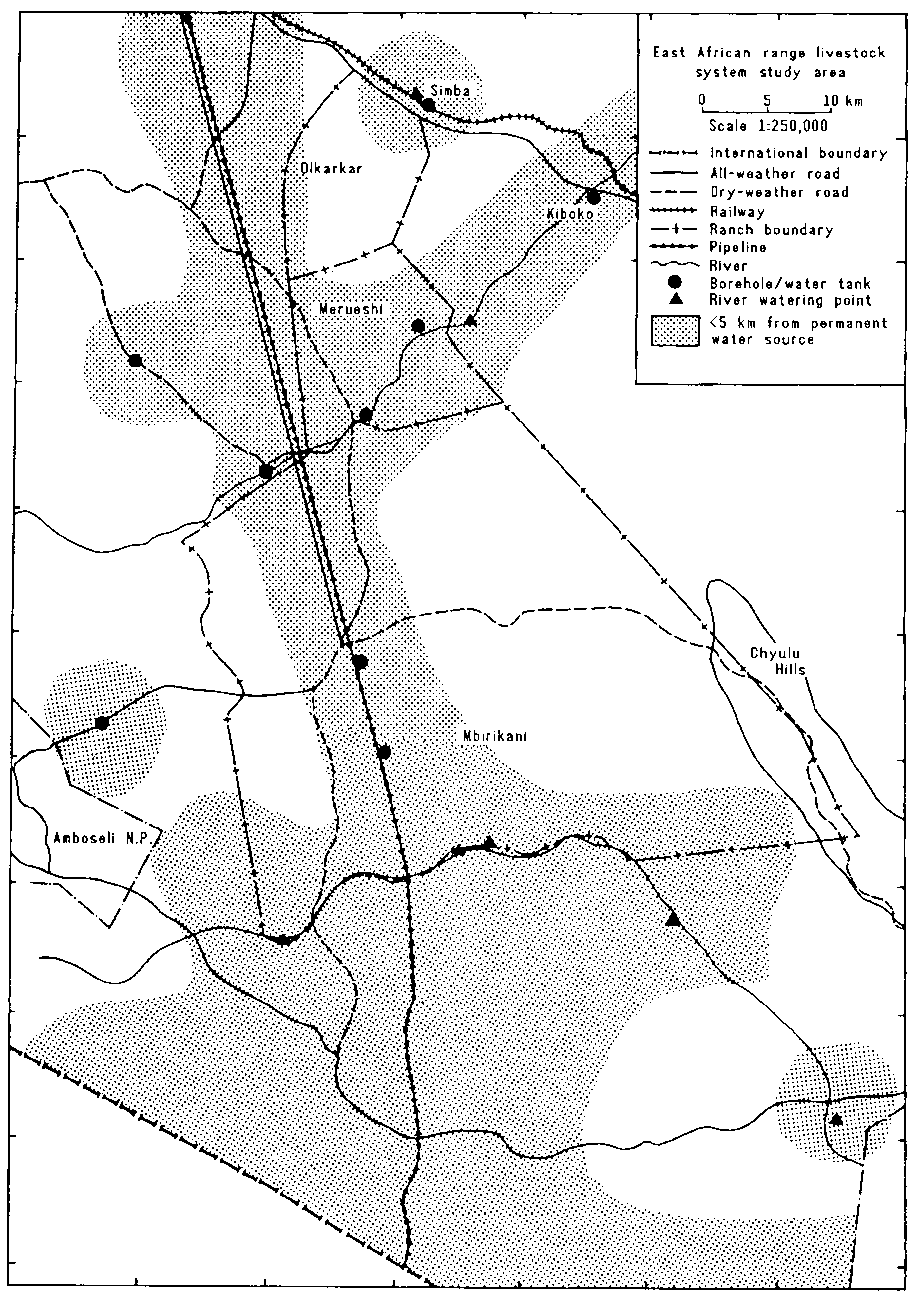
The study area comprised three group ranches, Olkarkar, Merueshi and Mbirikani, in eastern Kajiado District (Figure 4.1).
Olkarkar had higher densities of both people and livestock than the other two ranches (Table 4.1)1. As a result, the amount of land available per person and per livestock unit increased from north to south in the study area.
1 The human and livestock populations shown in Table 4.1 apply to years of average rainfall only and not to drought years. During the minor drought in 1982, most of the people and livestock left Mbirikani (for details, see Section 5.3: Water utilisation, grazing patterns and stocking rates). Even the northern ranches, which in normal years have a fairly sedentary population, experienced emigration during the severe drought of 1984 (Grandin et al, 1989).
Wild herbivores add roughly 25 to 30% to the livestock biomass in the study area. Grazers, e.g. wildebeest and zebra, account for some 40% of the wild herbivore biomass, or some 10% of total livestock biomass (see Section 2.3.5: Herbivore population).
Landscapes
The distribution of different landscape units in the study area is shown in Figure 4.2. The characteristics of the units (their land-form, geology and vegetation physiognomy) are listed in Table 4.2.
The physiography of the whole study area is influenced by the Chyulu Hills, which bound the area to the east. The Chyulu Hills consist of an upper-level plateau rising to an altitude of 2000 m (unit 2), which is surrounded by lava flows (unit 3) and a mixture of smaller lava ridges, uplands and footslopes (unit 8).
To the north-west of the Hills volcanic uplands are prominent, rising to an altitude of about 1200 m (unit 6) with cones and hills another 100 m above the rolling surface (unit 4). These uplands are the principal landscape in Olkarkar and continue into the northern and eastern part of Merueshi Ranch. To the east of this area is another upland unit (unit 5), with soils developed over basement complex; this unit covers most of the adjacent Kiboko Ranch.
Table 4.1. Land, people and livestock in three group ranches.
|
|
Olkarkar |
Merueshi |
Mbirikani | |
|
Size (km2) |
102 |
183 |
1350 | |
|
No. of households |
40 |
36 |
250 | |
|
People |
400 |
414 |
2700 | |
|
Cattle |
6500 |
5270 |
41500 | |
|
Smallstock |
6720 |
3170 |
19500 | |
|
Land availability |
|
|
| |
|
|
ha/person |
25 |
44 |
50 |
|
|
ha/household |
255 |
508 |
540 |
|
|
ha/TLU1 |
1.7 |
3.9 |
4.3 |
1 Tropical Livestock Unit (TLU) weighs 250 kg (cattle = 0.83 TLU in the north and 0.70 in the south, smallstock = 0.1 TLU; date from mid-June 1982).Stock populations refer to census estimates of resident populations in June 1982 for Olkarkar and Merueshi, and to aerial survey counts of all stock in December 1982 for Mbirikani (King et al, 1985). The latter were used because census estimates for this ranch as a whole were not available.
Figure 4.1. Map of the study area.
Table 4.2. Landscape units and their characteristics in eastern Kajiado District.
|
Map unit |
Land-form |
Geology |
Vegetation |
Location1 |
Area |
|
|
% |
km2 |
|||||
|
1 |
Hills |
Basement complex (gneiss) |
Bushland, woodland and thickets |
NW, Olgumtus mountain |
4.1 |
150 |
|
2 |
Upper slopes |
Volcanic (Recent) |
Sub-montane grasslands |
Chyulu Hills |
5.4 |
200 |
|
3 |
Lava flows |
Volcanic (Recent) |
Lava forest |
Chyulu Hills |
4.3 |
160 |
|
4 |
Cones |
Volcanic (Pleistocene) |
Bush- and grassland |
North and Central |
2.2 |
80 |
|
5 |
Uplands |
Basement complex |
Bushed grassland |
NE, Kiboko |
2.7 |
100 |
|
6 |
Uplands |
Volcanic (Pleistocene) |
Open and bushed grasslands |
Olkarkar and Merueshi |
6.5 |
240 |
|
7 |
Uplands |
Volcanic (Pleistocene) |
Bush- and woodland |
South |
10.8 |
400 |
|
8 |
Plains |
Volcanic (Recent) |
Open and bushed grasslands, patches of lava forest |
Chyulu Hills, SE Mbirikani |
9.5 |
350 |
|
9 |
Plains |
Volcanic (Recent) |
Grassland |
SE, Mbirikani |
3.3 |
120 |
|
10 |
Plains |
Volcanic (Pleistocene) |
Grassland |
NW, Poka |
1.6 |
60 |
|
11 |
Erosional plains |
Basement complex (with volcanic ash) |
Grasslands |
Central (Kiboko, Merueshi, Mbuko) |
5.4 |
200 |
|
12 |
Erosional plains |
Basement complex |
Bush- and woodland |
NW |
8.7 |
320 |
|
13 |
Erosional plains |
Basement complex |
Bushed and wooded grassland |
SW |
6.5 |
240 |
|
14 |
Erosional plains |
Basement complex (with volcanic ash) |
Open and bushed grassland |
E Mbirikani |
4.9 |
180 |
|
15 |
Erosional and piedmont plains |
Basement complex and colluvium |
Bushed grassland |
S Mbirikani |
10.3 |
380 |
|
16 |
Piedmont and lacustrine plains |
Colluvial and alluvial deposits |
Wooded grassland and woodland |
South |
3.8 |
140 |
|
17 |
Floodplains and bottomland |
Alluvial deposits |
Grassland |
Various |
8.9 |
330 |
|
18 |
Swamps |
Alluvial deposits |
Grassland |
South |
1.1 |
40 |
1 See Figure 4.2 for location of landscape units.
Source: Touber (1983).
To the south and west of these uplands are erosional plains over gneissic basement complex (units 11 and 12); these extend south along the western boundary of Mbirikani (unit 13). These flat or slightly undulating plains are bounded in the south by another series of volcanic uplands (unit 7), which are studded with small irregular outcrops of basaltic boulders. This unit forms the southern boundary of Mbirikani and extends south to the foothills of Kilimanjaro.
In the central part of Mbirikani, erosional plains form a lower-lying trough (1100-1150 m; units 14 and 15), merging with the Chyulu foothills to the east. The Kiboko river flows north-eastwards through these plains. In the south, the plains are broken by the Kikarankot River and its associated swamps (unit 18) and lacustrine plains (unit 16). The floodplains and bottomlands that flank these river systems are shown as unit 17.
Soils
The diverse physiography of the study area has resulted in a wide range of soils, most of which are deep and fine-textured. On the volcanic uplands and plains the soils range from stony Cambisols on the upper slopes to dark, cracking Vertisols in bottomlands and valleys. In the Chyulu Hills the main soils are Lithosols on lava flows, Andosols on coarse ash deposits and deep Luvisols on the flatter plains. Soils overlying gneissic basement complex are generally sandy, well drained and susceptible to erosion. The plains in the central, driest part of Mbirikani feature dark clays with vertic and saline-sodic properties (Touber, 1983).
Vegetation
Treeless grassland covers more than 40% of the study area (Table 4.3), including large parts of Olkarkar and Merueshi (unit 6) and almost all of the eastern part of Mbirikani (units 9,14 and large parts of unit 15).
Table 4.3. Density of woody cover in the study area.
|
Physiognomy |
Woody cover (%) |
Per cent of total area |
|
Open grassland |
0-2 |
42 |
|
Wooded and bushed grassland |
2-20 |
20 |
|
Bush- and woodland |
20-40 |
30 |
|
Dense woodland and forest |
> 40 |
8 |
Derived from Touber (1983).
Woody cover is found on units over basement complex, such as the northern plains and uplands (units 5 and 12). The southern fringe of Mbirikani is also somewhat more wooded; bushland is largely confined to the basalt outcrops on the volcanic upland (unit 7), but there are extensive Acacia tortilis woodlands on the lacustrine plains (unit 16). There are also patches of acacia woodlands along the Kiboko river. Dense forest occurs only on lava flows in the Chyulu Hills.
Many woody species have been identified in the area (see, for instance, de Leeuw and Chara, 1985; Touber, 1983; Kemei, 1982), but the predominant species in most parts of zones V and VI are Acacia mellifera, A. tortilis, A. nubica, A. ancistroclada, A. nilotica, Commiphora riparia, C. africana and Balanites aegyptiaca. Less drought-tolerant species (e.g. Combretum, Grewia and Premna) are confined to zone IV and occur mostly in unit 1.
The species composition of the herbaceous layer is fairly uniform across the study area, despite the diversity of the landscapes and soils. Four principal grassland communities were distinguished, based on the dominant genera (Chloris, Digitaria, Pennisetum and Sporobolus), but many species occurred widely (Table 4.4). Data from permanent transects in the Kaputiei area showed the same tendency of uniform species composition across sites (Njoka, 1984).
Although perennial grass species made up most of the grazable biomass in most landscape units, annual grasses and fortes were important but variable components of the herbaceous layer. Eragrostis cilianensis, E. tenuifolia, Dacryloctenium aegyptiaca, Aristida adscensionis and A. adoensis contributed substantially to the biomass in good rainy seasons, as did a plethora of annual herbs (de Leeuw and Chara, 1985; Njoka, 1984).
Several grassland types also included dwarf shrubs and perennial herbs, many of which are important browse plants for sheep and goats (de Leeuw and Chara, 1985; Kamau, 1986). These shrubs and herbs were more common on sandy soils over basement complex than on heavy soils and were more abundant in intensively grazed areas. Thus, such perennials were commonest in units 5, 11, 12 and 13 and in overgrazed portions of unit 15 along the pipeline in Mbirikani.
It is difficult to assess the extent to which the species composition of the herbaceous layer affects the grazing potential of the different landscape units. The productivity of the different grassland types was much confounded with rainfall events (i.e. localised showers or storms) and with past use (see Section 4.5: Water resources and Section 5.3: Water utilisation, grazing patterns and stocking rates). However, species composition influenced the grazing habits of domestic stock. This was demonstrated by Semenye (1987) who, over three seasons in 1983, recorded the forage species selected by grazing cattle in five locations in the study area. He found that, across seasons and locations, Chloris roxburghiana, Digitaria macroblephara and Pennisetum mezianum together made up 50 to 70% of the animals' diet. This appeared to be related to the abundance of the species in the study area (Table 4.4). Kibet (1986) made similar observations at the National Range Research Station, Kiboko.
The study area straddles the semi-arid and arid zones (zones V and VI: see Section 2.1: Agroclimatic zones and livestock-carrying capacity). The northern ranches and the eastern part of Mbirikani are situated in the semi-arid zone; the remainder of Mbirikani is in the arid zone. Most of the study area is classified as "Lower Midland Ranching Zone", where rainfed cropping will succeed only in seasons in which rainfall is exceptionally good, i.e. above average and/or well distributed (Jaetzold and Schmidt, 1983).
Rainfall is distinctly bimodal throughout eastern Kajiado The "first rains" fall from October to December and the "second rains" fall from March to May. There is a short dry period during January and February and a long dry season from June to early October. The growing season in the study area thus runs from October to May, with a hiatus of variable duration in January and February.
Table 4.4. Perennial grass composition of four grassland types.
|
Species |
North |
Bottomlands (7) |
South |
Per cent selected in grazing dietb |
||
|
Plains/uplands |
Uplands |
Uplands (9) |
Plains (15) |
|||
|
Aristida keniensis |
x |
x |
x |
- |
x |
6 |
|
Bothriochloa insculpta |
- |
x |
x |
- |
x |
2 |
|
Cenchrus ciliaris |
xx |
xx |
- |
- |
xx |
2 |
|
Chloris roxburghiana |
xx |
xxx |
x |
x |
xx |
20 |
|
Chrysopogon aucheri |
x |
- |
- |
xxx |
x |
3 |
|
Cynodon dactylon |
x |
x |
x |
- |
x |
- |
|
C. plectostachyus |
x |
x |
x |
- |
x |
7 |
|
Digitaria macroblephara |
xxx |
xx |
x |
x |
x |
16 |
|
D. scalarum |
- |
- |
- |
x |
- |
- |
|
Enneapogon macrostachys |
x |
- |
- |
- |
- |
- |
|
Eragrostis superba |
x |
- |
- |
xx |
- |
- |
|
Eustachys paspaloides |
x |
- |
- |
xx |
- |
- |
|
Ischaemum afrum |
- |
- |
xx |
- |
- |
1 |
|
Lintonia nutans |
- |
- |
xx |
- |
- |
- |
|
Pennisetum massaicum |
x |
- |
- |
- |
- |
- |
|
P. mezianum |
xx |
- |
xxx |
- |
xx |
19 |
|
P. stramineum |
- |
- |
- |
- |
xx |
- |
|
Sporobolus fimbriatus |
xxx |
x |
- |
xx |
- |
6 |
|
S. ioclados |
- |
- |
- |
- |
xx |
- |
|
Themeda triandra |
x |
- |
- |
x |
- |
- |
- = rare or absent; x = occasional; xx = common; xxx = abundant.
a Landscape units: see Table 4.2 and Figure 4.2.
b Adapted from Semenye (1987).
Annual potential evaporation is about 1950 mm, giving a moisture index of 0.31 for the mean annual rainfall of 616 mm at Makindu, a meteorological station in zone V near the study area. Daily potential evaporation ranges from a peak of about 6.0 mm in January and February to a low of 4.5 mm for June and July in the long dry season. Mean maximum temperatures for these two periods are 30°C and 26°C respectively and the mean minima are 19°C and 14°C. Absolute minimum temperature can go as low as 8°C (Musembi, 1986).
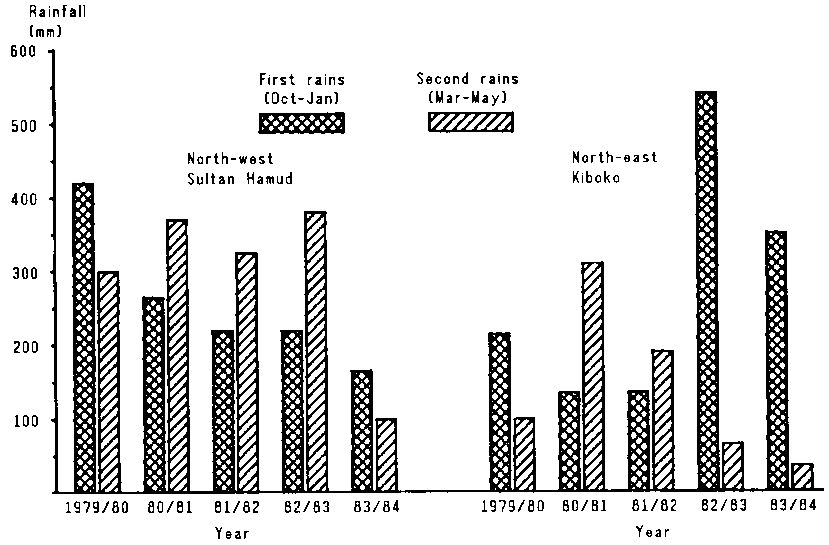
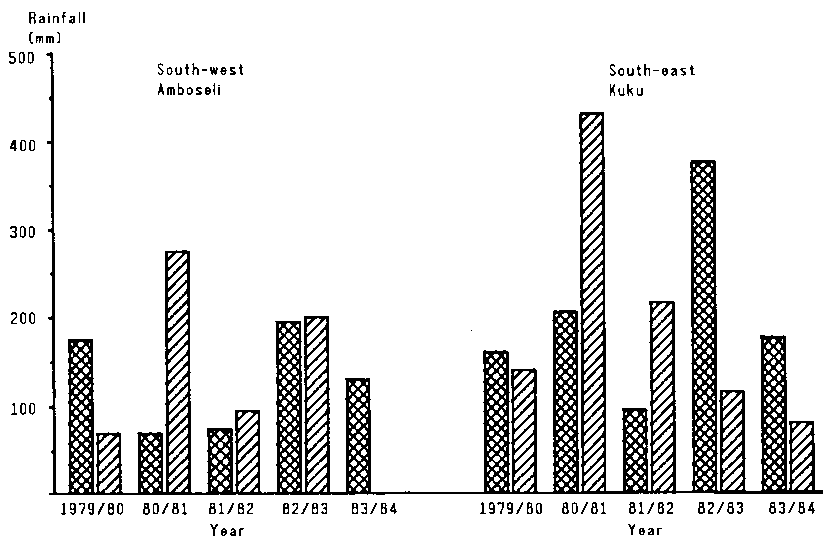
Figure 4.3 illustrates the patterns of rainfall in the study area between 1979 and 1984. The steep north-south rainfall gradient is evident.
Rainfall was above average in much of the study area for most of the late 1970s. In the northern part, climatic conditions remained close to normal through 1983, while the south experienced a minor drought for most of 1982; this was terminated by good rains in late 1982. A second, major drought affecting the whole study area started early in 1984 after a very short rainy season in late 1983 and very little or no rain in early 1984.
4.4.1 Biomass yield, rainfall and growing season
4.4.2 Forage quality
4.4.3 Carrying capacity
The general relationships between herbage production and carrying capacity were discussed briefly in Chapter 2 (see Section 2.1: Agroclimatic zones and livestock-carrying capacity). It was noted that amount and distribution of rainfall are the prime factors determining herbage production, but that availability of herbage is strongly influenced by grazing pressure in previous seasons. The following sections elaborate on these relationships, in particular as they apply to the forage supply of the study area.
Figure 4.3. Seasonal rainfall at four sites in eastern Kajiado District, 1979-84. (A)
Figure 4.3. Seasonal rainfall at four sites in eastern Kajiado District, 1979-84. (B)
Relationships between seasonal rainfall and biomass yield have often been used to predict forage availability. Deshmukh (1984) calculated an average ungrazed yield of 8 kg DM/ha per mm of rainfall for some major grassland types in eastern and southern Africa. Braun (1973) and Sinclair (1979) recorded average yields of 4 to 6 kg DM/ha per mm in the Serengeti Plains. Data given by Potter (1985) indicate yields of 4 to 7 kg DM/ha per mm, increasing with rainfall, for Themeda grasslands of the Athi Plains (de Leeuw and Nyambaka, 1988). Van Wijngaarden (1985) recorded similar yields in Tsavo National Park. The Tsavo study also demonstrated the importance of soil type and plant cover. Yields were 30 to 55% greater on deep, well drained sandy clays than on shallow gravely soils, and increased threefold as grass cover increased from 20 to 80% (van Wijngaarden, 1985; de Leeuw and Nyambaka, 1988)
Other workers have related biomass production to estimates of daily growth during the growing season. Bille and Heemstra (1979) estimated a growth rate of 30 kg DM/ha per day in the ILCA study area, while Braun (1973) in the Serengeti Plains found that daily growth rate increased from 15 kg DM/ha in short grasslands to 32 kg DM/ha in vegetation types with tall grasses. Data from Potter (1985) showed daily growth rates of 20-30 kg DM/ha for rainfall of 300-400 mm per season, decreasing to 10-15 kg DM/ha per day for rainfall of 150-250 mm per season. Daily growth rates based on Potter's (1985) data have been used to estimate long-term forage supplies (see Section 10.1.1: Fodder resources). Primary production in the three group ranches was surveyed several times between 1980 and 1984.
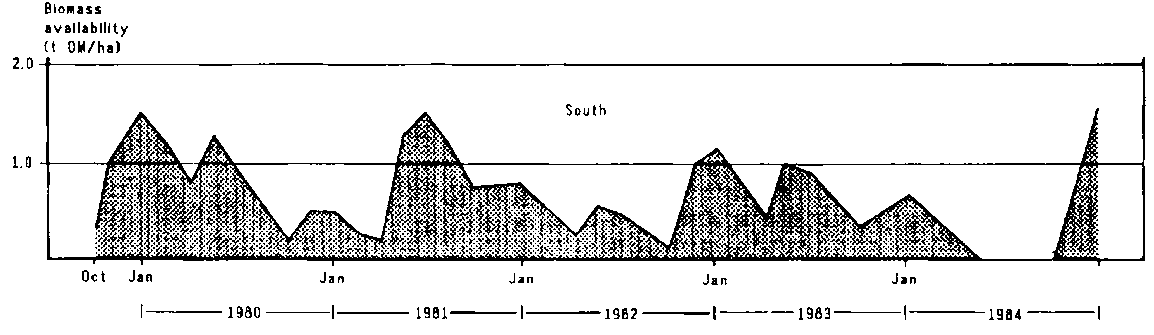
In 1980-81 standing biomass was measured on all three group ranches at the end of three dry seasons (October 1980, March 1981 and October 1981) and at the end of one rainy season, in June 1981 (Bille and Chara, 1981). Standing biomass was generally less than 0.5 t DM/ha at the end of the dry seasons except in river valleys and for grassland over Vertisols, where yields reached 1 t/ha. The response to the rains in early 1981 was low. In the northern ranches, standing biomass averaged about 0.7 t/ha in June 1981, ranging from 0.4 to 1.1 t depending on the level of overgrazing and soil type. The lower rainfall in the south was reflected in less standing biomass on Mbirikani than on the northern ranches.
Allowing for the amount of herbage removed by grazing, Bille and Chara (1981) estimated net primary productivity at 800-900 kg DM/ha, about 2-3 kg/ha per mm of rain or 15 kg/ha per day for a growing season receiving 250-350 mm rainfall. These growth rates are lower than those quoted above, which the authors attributed to the high grazing pressure on the ranches: at least one third of Olkarkar and half of Merueshi was seriously overgrazed, which resulted in low plant cover and consequent poor response to rainfall.
The good rains in late 1982 (first rains of 1982/83) resulted in considerable herbage growth throughout the study area. In the north, standing biomass in ungrazed swards increased in November 1982 from about 1.0 to 1.7 t DM/ha and reached 3.4 t in late January 1983 (Table 4.5). Similarly, in the south (eastern Mbirikani), standing biomass rose from about 1 t DM/ha in early November to 1.9 t DM/ha in early 1983 (Table 4.5); showers in February pushed yields up to nearly 3 t/ha in April. In response to these rains, plant cover in Mbirikani increased quickly. Dense cover (over 60%) was recorded in the north-east along the foothills of the Chyulu Hills, along the river valleys in the south and south-east and along the Kiboko River in the north-west. Regeneration of plant cover was much poorer in central Mbirikani because it had been overgrazed and because of the prevalence in that area of sodic and saline soils.
Table 4.5. Grazable standing biomass in northern and southern parts of eastern Kajiado District 1982/83.
|
Period |
North |
South |
||||
|
n |
Grazable biomass |
n |
Grazable biomass |
|||
|
Mean |
SE |
Mean |
SE |
|||
|
Early November |
14 |
960 |
170 |
9 |
940 |
100 |
|
Late November 1982 |
6 |
1710 |
210 |
|
|
|
|
Late January 1983 |
16 |
3370 |
210 |
13 |
1850 |
270 |
|
Late April 1983 |
13 |
2870 |
360 |
|
|
|
SE = standard error.
n = number of samples.
Similar patterns of herbage growth were recorded after the rains in late 1984 that broke the 1983/84 drought. More than one third of the 180 plots sampled had more than 2.5 t DM of standing biomass per hectare. Regression of standing biomass on plant cover indicated yields of 3 t DM/ha at 80% plant cover, similar to values observed by van Wijngaarden (1985) for a seasonal rainfall of 250 mm.
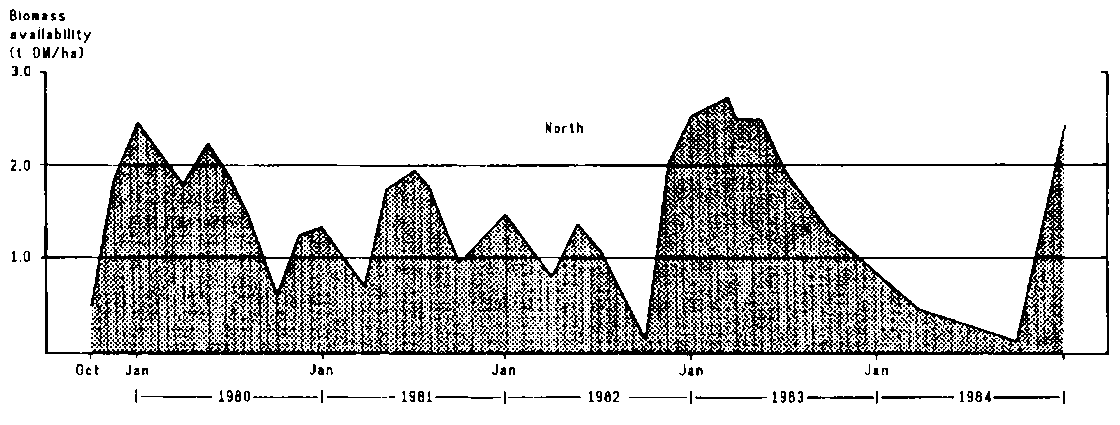
A profile of herbage availability was constructed for the northern and southern parts of the study area. The amount of forage available in the north (northern Olkarkar) rarely fell below 1 t DM/ha except towards the end of the long dry season of 1982 and during the 1983/84 drought(Figure 4.4). Yields were generally much lower in Mbirikani than in Olkarkar (Figure 4.4); the high yields shown in Table 4.5 were mainly confined to the eastern part of the ranch, where soils were more fertile and grazing pressure was low.
Forage supply was monitored using aerial surveys and ground sampling. The latter, carried out between January 1982 and June 1983, involved determination of both amount and quality of standing biomass. These studies were complemented by comprehensive analyses of the nutrient content of extrusa from oesophagally fistulated cows grazing with local herds over three seasons in 1983, between February and October, in five grazing locations covering all three group ranches (Semenye, 1988).
Crude protein content
During the growing season, mean crude protein content of clipped and grazed herbage ranged from 11% in the first rains to 7.5% during the second rains. Contents of up to 16% were measured in new regrowth with yields of less than 0.5 t DM/ha (Figure 4.5). Crude protein content fell by about 1% a month as the herbage matured and bulked up, falling to 46% in dry grass and litter.
In all seasons the crude protein content of leaves was higher than that of stems and leaf sheaths (Semenye, 1987). Leaves formed 70-80% of the diet of grazing cows during the growing season and 40% of the diet during the dry season.
Protein content was closely linked with the amount of standing biomass. During growing seasons in which rainfall was good (e.g. 1982/83) leaf protein content fell from about 10% when there was 1 tonne DM of standing biomass per hectare to about 5% in mature stands of 2.5 t DM/ha. Thus, the good rains in late 1982 (first rains of 1982/83 growing season) resulted in there being in January-February 1983 a large amount of standing biomass that contained only 4.5% crude protein.
The crude protein content of extrusa from fistulated cows differed markedly between ranches. During the second rains of 1982/83, cows in northeast Olkarkar consumed a diet containing 13.0% crude protein compared with 8.5% crude protein in the diet of cows in south-west Mbirikani. This difference was due in part to better rainfall in the north of the study area leading to a flush of herbage growth, but was also related to differences in soils and species composition of the herbaceous layer. On average, herbage growing on deep basalt soils contained 40% more crude protein than herbage growing on soils over basement complex; unfortunately, interactions with season and differences in sampled standing biomass do not allow a firm conclusion (Semenye, 1988).
Figure 4.5. Nitrogen content of green and dry herbage over five seasons, January 1982 to June 1983.
Digestibility
In vitro digestibility changed much less between seasons than did crude protein content. During growing seasons, ingested leaf herbage had an average digestibility of 54%, with short-lived peaks of up to 65% in very young growth. Late in the dry seasons digestibility fell to 46%. At other times, when both mature, dry herbage and green herbage were present, extrusa were between 45 and 50% digestible, depending on the degree of selection animals practised (Semenye, 1988). As expected, in vitro digestibility was closely correlated with crude protein content (r=0.86). Thus, since these two factors are the main determinants of the nutritive value of range forage, subsequent discussion refers to crude protein content alone as an indicator of nutritive value.
Mineral content
Mineral contents of extrusa from fistulated cows were generally above minimum required levels for cattle (Semenye, 1988), although copper content was marginal at one sample site at 4.2 ppm. In whole-plant samples phosphorus content ranged from 0.25% in dry herbage to 0.50% in green growth (de Leeuw, unpublished data), well above the minimum of 0.18% required by cattle for growth. As with other nutritive characters, P content was highest in plants grown on volcanic soils.
Seasonal trends
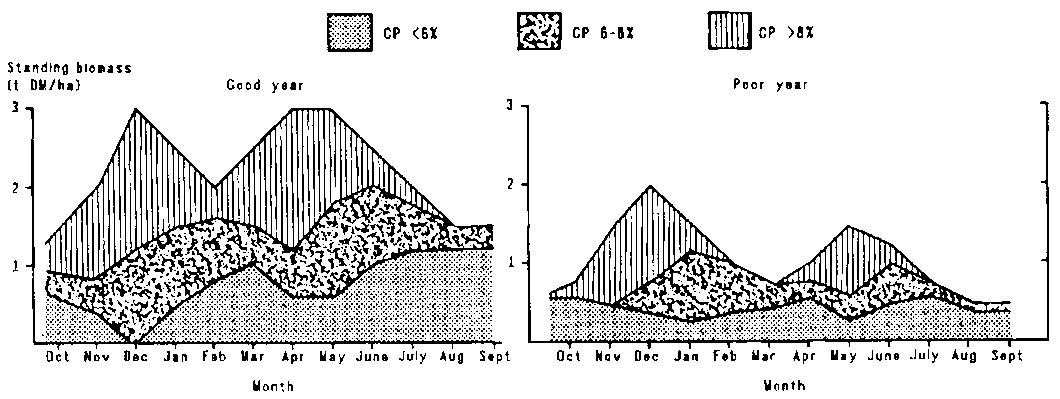
These data on nutritive values of forage were used to analyse the forage supply situation further. Two additional data sets were compiled by estimating monthly quantity and quality for good and bad years. The first set provided average digestibility and crude protein content by month. To illustrate the variability between year-types, the parameters are given for a fairly good and a poor rainfall year, roughly indicative of the grazing conditions in the north and in the south of the study area (Figure 4.6). Although differences between years are pronounced, the annual curves follow similar trends. In a good year the herbage contains an average of 8% or more crude protein for 8 months, compared with only 5-6 months in a poor year.
At the onset of each rainy season there is a rapid increase in the amount of high-quality biomass concomitant with the rapid disappearance of old standing herbage left over from the previous season (Figure 4.6). As the rainy season progresses the crude protein content of the herbage declines and old standing forage continues to disappear. At the end of the rains only medium and poor-quality forage remains.
Figure 4.6. Crude protein content of standing biomass in a good and a poor rainfall year.
The supply of crude protein clearly differs markedly between year-types. In a good year supplies of good-quality forage exceed 1 t DM/ha for 6 months (November-January and March-May), compared with only 2 to 3 months in a poor year.
In conclusion, the nutritional status of Maasai cattle is strongly influenced by the duration of the alternating dry and wet seasons and the resultant fluctuations in forage quality and supply. Due to the relatively high fertility of the predominantly volcanic soils, mineral content of forage was quite high. Hence, shortage of forage seems to be more limiting than the quality of the forage available (see Section 10.2.1: Stocking rate and herd size).
The long-term carrying capacity of agroclimatic zones V and VI, within which the group ranches are located, has been estimated to be between 3 and 7 ha per 250 kg tropical livestock unit (Section 2.1: Agroclimatic zones and livestock-carrying capacity). However, such average estimates may not be very useful given the large between-year differences in grazing resources.
The carrying capacity of grazing land is determined from:
· the amount of forage available per unit area within a specified time period;· forage requirements of the herbivore population by species;
· forage allowances in relation to animal requirements and to safeguards aimed at ensuring sustained range productivity;
· availability of forage to the herbivore population as determined by location or distance.
Forage availability
The amount of standing biomass at the end of a growing season was estimated for Olkarkar and Merueshi (Table 4.6). These estimates are lower than the yields given in Table 4.5 and Figure 4.4 mainly because the estimated yields were adjusted to allow for average plant cover on each ranch.
Table 4.6. End-of-season standing biomass (kg DM/ha) in relation to growing season and rainfall, Olkarkar and Merueshi.
|
Rainfall (mm) |
End-of-season standing biomass |
|
|
Olkarkar1 |
Merueshi2 |
|
|
100 |
500 |
240 |
|
200 |
1000 |
480 |
|
300 |
1500 |
720 |
1 Based on 50% plant cover and a rainfall Use efficiency (RUE) of 10 kg DM/ha.
2 Based on 30% plant cover and an RUE of 8 kg DM/ha.
Forage requirements
Assuming an average daily dry-matter intake of 2.5% of bodyweight (Boudet and Riviere, 1968; Minson and McDonald, 1987), each tropical livestock Unit (TLU) will consume 6.25 kg of forage dry matter daily or 2.3 t DM annually.
Forage allowance
The rate at which herbage disappears is higher than animal intake because of wastage and trampling. Thus, forage allowance was set at 10 kg DM/TLU per day (3.6 t DM/TLU per year), i.e. a utilisation rate of 62.5%. The increase in daily allowance over forage intake is related to the 'proper-use factor', i.e. the maximum rate of utilisation for sustainable rangeland use. The most common 'proper-use factor' is a utilisation rate of 50% of standing herbage yield; this gives a herbage allowance of 12.5 kg DM/TLU per day or 4.6 t DM/TLU per year. Applying this 'safe' allowance to the stocking rate for Kajiado District as a whole (3.3 ha/TLU in 1983; see Section 2.3.5: Herbivore population) indicates the need for an average annual forage yield of 1.4 t DM/ha. A dry-matter disappearance rate of 10 kg DM/TLU per day gives a required yield of 1.1 t DM/ha per year.2
2. The 'proper-use factor' is based on the concept that there is a certain rate of defoliation above which the sustained productivity of range vegetation is impaired. Van Wijngaarden (1985) in Tsavo National Park (850-500 mm rainfall) demonstrated that when more than 45% of the dry-season biomass was removed, perennial plant cover during the following rainy season was reduced, while below this level of removal, plant cover increased. In contrast Potter (1985), working in the somewhat higher rainfall area of the Themeda grasslands of the Athi Plains, showed that long-term productivity was not reduced even at a very high defoliation rate (cutting every 8 weeks at a height of 5 cm) or when grasslands were continuously grazed at a stocking rate of 2 ha/TLU. These contrasting observations have implications for assessing the long-term carrying capacity of the Maasai group ranches (see Section 10.2.1: Stocking rate and herd size, for long-term implications).
Accessibility of forage supplies
Factors that modify the actual amount of forage that is accessible to livestock include distance from water, disease hazards, palatability and type of species present (e.g. the proportion of woody species in the biomass) (see Section 5.3: Water utilisation, grazing patterns and stocking rates).
Safe stocking rate
The safe stocking rate was calculated from forage requirements over the long dry season as this is the most critical period in terms of forage supply. It was assumed that the amount of standing forage available for dry-season use is determined solely by the second rains (March-May), i.e. no forage is carried over from the first rains. The dry season usually lasts 5 to 7 months.
For a herbage allowance of 10 kg DM/TLU per day, safe stocking rate for Olkarkar varies from 1.0 ha/TLU when a good rainy season (300 mm) is followed by a 5-month dry season to 4.2 ha/TLU when a poor rainy season (100 mm) is followed by a 7-month dry season (Table 4.7). The total amount of stock that can be safely carried on the 10000 ha Olkarkar ranch thus varies from 10000 to 2400 TLU. The predicted yields of herbage for Merueshi ranch are about half those for Olkarkar (Table 4.6); the safe stocking rate for this 18300 ha ranch thus ranges from 2.1 to 8.75 ha/TLU, or 8700 to 2100 TLU.
Table 4.7. Minimum land requirement (ha/TLU) for Olkarkar in relation to seasonal rainfall and duration of subsequent dry season.
|
Rainfall (mm) |
Dry-season length (months) |
||
|
5 |
6 |
7 |
|
|
100 |
3.0 |
3.6 |
4.2 |
|
200 |
1.5 |
1.8 |
2.1 |
|
300 |
1.0 |
1.2 |
1.4 |
In Olkarkar the long dry season lasts, on average, about 6 months and the second rains average about 200 mm. Thus, a dry-season stocking rate of 2 ha/TLU could be maintained in most years, but would have led to shortage of forage in 1980/81 (Bille and Chara, 1981) and towards the end of the long dry season in 19823. In Merueshi the long dry season commonly lasts up to 1 month longer than in Olkarkar and rainfall in the second season averages 150 mm. Thus, a stocking rate of 4 ha/TLU would be safe in most years, but would have led to serious shortages of forage in 1980/81 and during the dry spell in 1982.
3 Sloane (1986) used the length of the growing season to estimate carrying capacity of rangelands in Kenya, but arrived at much lower values. For instance, for a growing period of 8 months, a stocking rate of 6 ha/TLU was allowed. This translates to a conservative utilisation of only 25% of the standing biomass as compared to 62.5% allowed in the present study. It appears that Sloane chose conservative values as long-term averages to provide sufficient margins for seasons of below-normal rainfall, to allow for the often large proportion of unpalatable species in the available biomass and for extensive areas of low herbage productivity (see Section 2.1: Agroclimatic zones and livestock-carrying capacity).
It is difficult to estimate the safe stocking rate for Mbirikani because much of the ranch is too far from the water pipeline (the main source of water) to be grazed (see Section 5.3.3: Grazing patterns and stocking rates in the southern ranch). Areas within reach of the pipeline have been seriously overgrazed. Rainfall during the second rains rarely exceeds 200 mm and was much less in 1982 and 1984. Thus, while in good years a stocking rate of 46 ha/TLU may be safe, more than 10 ha/TLU may be needed after poor rains.
The most important structure supplying water to the study area is the pipeline that cuts through western Mbirikani and skirts Merueshi on the west (Figure 4.7). There are several public water outlets from the pipeline, some with storage tanks, but private outlets are increasingly important. During 1983-84 at least 15 private connections, some with storage tanks and most with water troughs, were installed between Makutano and Olandi over less than 15 km. In Merueshi at least three private installations had been completed by 1984. There is a second, smaller pipeline system around the Amboseli National Park. Other man-made water sources include boreholes along the Kiboko River and in the north-west of the study area.
There are several other permanent water sources serving the study area. Some have been improved by man. The most important of these is Simba Springs, which provides water to most of the stock on Olkarkar and Kiboko group ranches, similar springs near Kiboko town, and the manmade shallow wells in the Kiboko River on the border between Kiboko and Merueshi group ranches. The swamps that form the southern boundary of Mbirikani Group Ranch are also a permanent source of water. Those farthest west drain into Lake Amboseli; the rest drain into the Looltureshi River and thence into the Tsavo River.
Most of Merueshi lies within 5 km of a permanent water source, compared with only a quarter of Olkarkar. There is no permanent source of water in eastern Mbirikani and more than 60% of this ranch is more than 5 km from a permanent water source (Figure 4.7).
Two types of seasonal water source are important in the study area: pools in riverbeds and streams, and ponds. Pools are found in riverbeds following the flash floods that occur after heavy rainfall. Ponds are common in areas underlain by basement complex, such as Kiboko Ranch, Mbuko Ranch (west of the pipeline) and along the northern end of the Chyulu foothills. Six ponds were used in Merueshi in good rainfall seasons during the study period. Olkarkar has few ponds because of its volcanic geology. Ponds along the Chyulu Hills rarely fill up because the volcanic ash deposits are very porous and only very heavy storms leave standing water.
Figure 4.7. Distribution of permanent water sources in the study area.
The availability of water is an important facet of the Maasai production system and water facilities and watering management are discussed further in subsequent chapters: in Chapter 6 (Labour and livestock management), the effect of the location of water points on residence and land utilisation patterns; in Chapter 7 (Productivity of cattle and smallstock), the relationship to labour and herding management; and in Chapter 11 (The potential for improving the livestock production and welfare of the pastoral Maasai), the possible improvements to the existing facilities.
Bille J C and Chara P 1981. Annual evolution of the grass biomass in Olkarkar, Merueshi and Mbirikani areas. Internal Report, ILCA/Kenya. 30 pp. ILCA (International Livestock Centre for Africa), Addis Ababa, Ethiopia. [ILCA Microfiche no. 18767]
Bille J C and Heemstra H. 1979. An illustrated introduction to the rainfall patterns in Kenya. ILCA/Kenya Working Document 12. 57 pp. ILCA (International Livestock Centre for Africa), Addis Ababa, Ethiopia. [ILCA Microfiche no. 16878]
Boudet G and Riviere R. 1968. Emploi pratique des analyses fourragères pour l'appréciation des paturages tropicaux. Revue d'élevage et de médecine vétérinaire des pays tropicaux 21 (2):227-266.
Braun H M H. 1973. Primary production in the Serengeti: Purpose, methods and some results of research. Annales de l'Université d'Abidjan Série E: Ecologie 6:171 -188.
Deshmukh I K. 1984. A common relationship between precipitation and grassland peak biomass for East and southern Africa. African Journal of Ecology 22:181 -186.
Grandin B E, de Leeuw P N and Lembuya P 1989. Drought, resource distribution and mobility in two Maasai group ranches in southeastern Kajiado district. In: Downing T E, Gitu K W and Kamau C M (eds), Coping with drought in Kenya. National and local strategies. Lynne Rienner Publishers, Boulder, Colorado, USA. pp. 245-263.
Jaetzold R and Schmidt H. 1983. Farm management of Kenya. Vol. IIC. East Kenya. Ministry of Agriculture, Nairobi, Kenya. 410 pp.
Kamau P N. 1986. Effects of available forage on goat dietary selection and nutrition. In: Hansen R M, Woie B M and Child R D (eds). Range development and research in Kenya. Proceedings of a conference held at the Agricultural Resources Centre, Egerton College, Njoro, Kenya, 1-5 April 1986. Winrock International Institute for Agricultural Development, Morrilton, Arkansas, USA. pp. 103-106.
Kemei I K. 1982. Vegetation environmental relationships on the Kiboko Range Research Station, Kenya. PhD thesis, Texas A & M University, Texas, USA. 87 pp.
Kibet P F K. 1986. Botanical composition and selection of diets by range cattle. In: Hansen R M, Woie B M and Child R D (eds). Range development and research in Kenya. Proceedings of a conference held at the Agricultural Resources Centre, Egerton College, Njoro, Kenya, 1-5 April 1986. Winrock International Institute for Agricultural Development, Morrilton, Arkansas, USA. pp. 93-102.
King J M, Sayers A R. Chara P. de Leeuw P N. Peacock C P, Fillo F and Maehl J H H. 1985. Improving aerial counts of Maasai livestock. Agricultural Systems 16:231-256.
de Leeuw P N and Chara P. 1985. Species preferences in browse of sheep and goats in mixed Maasai flocks in eastern Kajiado, Kenya. In: Proceedings of the Fourth Small Ruminant CRSP Workshop, Kakamega, March 1985. Small Ruminant Collaborative Research Support Program, Nairobi, Kenya. pp. 252-261.
de Leeuw P N and Nyambaka R. 1988. The prediction of rangeland production from rainfall data in arid and semi-arid eastern Africa. In: Pasture Network for Eastern and Southern Africa (PANESA), African forage plant genetic resources, evaluation of forage germplasm and extensive livestock production systems. Proceedings of the Third Workshop held at the International Conference Centre, Arusha, Tanzania, 27-30 April 1987. ILCA (International Livestock Centre for Africa), Addis Ababa, Ethiopia. pp. 260-268.
Minson D J and McDonald C K.1987. Estimating forage intake from the growth of beef cattle. Tropical Grasslands 21:116-122.
Musembi D K. 1986. The seasonal climate of rangelands. In: Hansen R M, Woie B M and Child R D (eds), Range development and research in Kenya. Proceedings of a conference held at the Agricultural Resources Centre, Egerton College, Njoro, Kenya, 1-5 April 1986. Winrock International Institute for Agricultural Development, Morrilton, Arkansas, USA pp. 183-200.
Njoka T J. 1984. Evaluation of seasonal and drought effects on range trend analysis in Kajiado District, Kenya. In: Siderius W (ed), Proceedings of the workshop on land evaluation for extensive grazing (LEEG). ILRI Publication 36. International Institute for Land Reclamation and Improvement, Wageningen, The Netherlands. pp. 195-206.
Potter H L. 1985. Aspects of climate, herbage growth and animal production in a semi-arid area of Kenya. PhD thesis, University of Nairobi, Nairobi, Kenya. 195 pp.
Semenye P P. 1987. Factors influencing Maasai came productivity and nutrition in Kajiado District, Kenya. PhD thesis, University of Nairobi, Nairobi, Kenya. 326 pp.
Semenye P P 1988. Grazing behaviour of Maasai cattle. in: Pasture Network for Eastern and Southern Africa (PANESA), African forage plant genetic resources, evaluation of forage germplasm and extensive livestock production systems. Proceedings of the Third Workshop held at the International Conference Centre, Arusha, Tanzania, 27-30 April 1987. ILCA (International Livestock Centre for Africa), Addis Ababa, Ethiopia. pp. 325-330.
Sinclair A R E. 1979. The Serengeti environment. In: Sinclair A R E and Norton-Griffiths M (eds). Serengeti, dynamics of an ecosystem. University of Chicago Press, Chicago, USA. pp. 3145.
Sloane P. 1986. Population supporting capacity study for Kenya. FAO Internal Report INT/75/P 13. FAO (Food and Agriculture Organization of the United Nations), Rome, Italy. 170 pp.
Touber L. 1983. Soils and vegetation of the Amboseli-Kibwezi area. Kenya Soil Survey R6, Ministry of Agriculture, Nairobi, Kenya. 125 pp.
van Wijngaarden W. 1985. Elephants-trees-grass-grazers: Relationships between climate, soil, vegetation and large herbivores in a semi-arid savanna ecosystem. ITC Publication 4. ITC (International Institute for Aerospace Survey and Earth Sciences), Enschede, The Netherlands. 165 pp.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
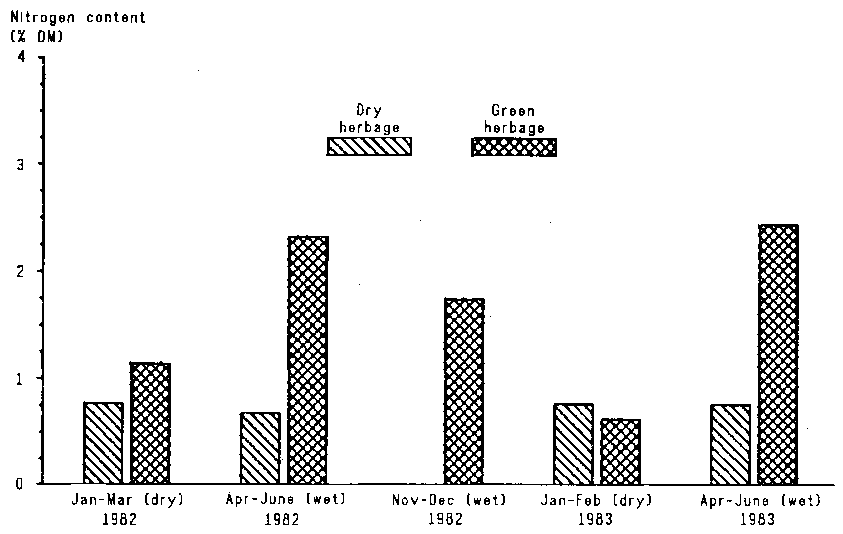{kind=link}
{kind=link}
{kind=link}