![]()
![]()
![]()
por
Alexandre Magno Sebbenn
Antonio Carlos Scatena Zanatto
Lígia de Castro Ettori
Omar Jorge Di Dio
Junior[23]
INTRODUCCIóN
El Instituto Forestal de Sao Paulo, Brasil, (IFSP) ha venido conservando desde 1979 una serie de especies brasileñas de árboles forestales que están amenazadas o sometidas a una reducción genética ocasionada por la creciente industrialización y los desarrollos agrícolas y urbanos no planificados. Vitor (1975) da una idea de la preocupante situación de los bosques naturales en el estado de Sao Paulo, y relata que a principios del siglo XIX, el 70,5% de la superficie del Estado estaba cubierta por bosques naturales mientras que en 1975 sólo quedaba el 7% de estos bosques. El Cuadro de Expertos de la FAO en Recursos Genéticos Forestales, inscribe las siguientes especies de Brasil como necesitadas de atención: Aspidosperma polyneuron, Araucaria angustifolia, Balfourodendron riedelianum, Cariniana legalis, Dipterix alata, Hymenaea coubaryl, Myracrodruon urundeuva (FAO, 1998, 2000). Una referencia histórica interesante es la realizada a mediados del siglo XX por el naturalista sueco Albert Löfgren: “La devastación de los bosques está destruyendo lo que tardó la naturaleza millones de años en crear; el precio de esta destrucción será muy elevado. En nombre de la vida misma y de nuestro futuro necesitamos detener esta devastación” (IFSP, 1994).
Hoy en día se siente vivamente la falta de una política forestal para estimular la conservación genética de los bosques remanentes, la reforestación y recuperación de las áreas degradadas y la ordenación forestal sostenible. Mientras en 1886, quedaba más del 70% del bosque atlántico, este área es actualmente sólo el 7%.
El programa de conservación genética ex situ de las especies brasileñas nativas en el IFSP se propone conservar la variabilidad genética existente entre distintas poblaciones de especies forestales y dentro de ellas, para su utilización actual o futura, a fin de atender las necesidades económicas, sociales y ambientales. Los objetivos del programa incluyen también el estudio de la estructura genética, el comportamiento selvícola, la transmisión hereditaria de los rasgos de interés económico y la mejora de la producción de semilla para la reforestación de áreas degradadas y alteradas.
EL INSTITUTO FORESTAL DE SAO PAULO
El IFSP, que forma parte actualmente de la Agencia Ambiental del Gobierno del Estado de Sao Paulo, fue fundado en 1888 por el naturalista sueco Alberto Löfgren y conocido originalmente como el Jardín Botánico. En 1911 el Jardín Botánico se transformó en el Servicio Forestal del Estado de Sao Paulo y en 1970 se convirtió en el Instituto Forestal de Sao Paulo (IFSP, 1994). Cuenta este Instituto con varias estaciones ecológicas, parques, bosques, estaciones experimentales, viveros y reservas forestales estatales, distribuidos por todo el estado de Sao Paulo, ocupando aproximadamente el 3,4% de su superficie[24]. Del área total administrada por el IFSP, el 94% (0.8 millones de hectáreas) se utilizan para actividades de conservación. Los mayores de los últimos restos del bosque atlántico del Estado de Sao Paulo se encuentran en estas áreas, que sirven para la conservación in situ y como fuentes de semilla y propágulos para la rehabilitación futura de los bosques naturales del Estado. Por ello, el valor de estas áreas es incalculable. El IFSP asegura también la conservación ex situ de las especies forestales brasileñas nativas y exóticas. En la actualidad el programa de conservación ex situ incluye 24 especies arbóreas forestales nativas y 45 especies arbóreas introducidas. Las últimas incluyen 23 especies de Eucalyptus, 14 de Pinus y 8 de otros géneros: Liriodendron, Toona, Grevillea, Cupressus, Cryptomeria, Cordia y Araucaria (véase Gurgel Garrido, 1997).
CONSERVACIÓN EX SITU
Las actividades de conservación ex situ fueron iniciadas por la Dra. Ana Cristina Machado de Franco Siqueira, que fue uno de los primeros científicos en reconocer los problemas de la pérdida acelerada de recursos genéticos forestales en el Estado de Sao Paulo y en Brasil, para adoptar las acciones correspondientes.
El Cuadro 1 contiene información sobre las especies forestales nativas incluídas en las áreas de conservación ex situ. Estas áreas se han establecido desde 1979. Sin embargo, incluyen también dos ensayos de procedencias de Araucaria angustifolia establecidos en 1973, que fueron posteriormente incorporados al programa de conservación ex situ. En la actualidad el programa incluye 24 especies y 41 áreas experimentales. Algunos experimentos están repetidos en dos estaciones. Las áreas ex situ incluyen ensayos de progenies y de progenies y poblaciones, establecidas inicialmente como plantaciones puras, siguiendo diseños experimentales como bloques con distribución al azar, bloques completos al azar y el diseño compacto en bloques por familias, siempre con 6 bloques y 5 plantas por muestra, lo que totaliza 30 plantas por progenie o estación. Desde 1990, reconociendo la importancia de respetar las etapas sucesionales para la mejor adaptación y desarrollo de la mayoría de las especies arbóreas forestales nativas de Brasil, los experimentos se han establecido mezclando varias especies de fases sucesionales en los bloques, incluyendo especies pioneras, secundarias y climácicas. El paréntesis con un número en el Cuadro 1 indica que esta especie está mezclada con especies con el mismo número del paréntesis.
Cuadro 1: Experimentos de conservación genética ex situ con especies forestales nativas en el IFSP, Brasil. Recopilado por Gurgel Garrido (1997)[25]
|
Especie |
Experimento |
Establecimiento |
Número de procedencias |
Número total de plantas |
||
|
Tipo |
Diseño |
Año |
Estac. |
|||
|
Anadenathera falcata |
Pt |
IC |
1994 |
1 |
1 |
20 |
|
Araucaria angustifolia |
Pt |
BC |
1974 |
2 |
1 |
32 |
|
Araucaria angustifolia |
Or |
BC |
1985 |
1 |
1 |
19 |
|
Araucaria angustifolia |
p/p |
BFC |
1973 |
2 |
5 |
115 |
|
Araucaria angustifolia |
Pt |
BC |
1981 |
1 |
7 |
70 |
|
Araucaria angustifolia |
p/p |
BFC |
1980 |
1 |
15 |
121 |
|
Aspidosperma polyneuron |
Pt |
BC (1) |
1992 |
1 |
1 |
23 |
|
Astronium graveolens |
Pt |
BC |
1990 |
1 |
1 |
23 |
|
Balfourodendron riedelianum |
p/p |
BFC |
1984 |
1 |
3 |
48 |
|
Cariniana legalis |
p/p |
BFC |
1992 |
2 |
3 |
50 |
|
Cordia trichotoma |
Pt |
BC |
1986 |
1 |
1 |
32 |
|
Cordia trichotoma |
Pt |
BC |
1986 |
1 |
1 |
25 |
|
Dipteryx alata |
p/p |
BFC |
1980 |
1 |
2 |
38 |
|
Dipteryx alata |
p/p |
BFC |
1986 |
1 |
2 |
42 |
|
Dipteryx alata |
Pt |
IC |
1981 |
1 |
1 |
25 |
|
Dipteryx alata |
Pt |
BC |
1986 |
1 |
1 |
25 |
|
Enterolobium contortisiliquum |
Pt |
BC |
1990 |
1 |
1 |
23 |
|
Esenbeckia leiocarpa |
Pt |
BC (2) |
1992 |
1 |
1 |
25 |
|
Gallesia gorarema |
Pt |
BC |
1985 |
1 |
1 |
25 |
|
Gallesia gorarema |
p/p |
CFB |
1982 |
2 |
3 |
65 |
|
Genipa americana |
Pt |
BC (3) |
1997 |
1 |
1 |
21 |
|
Guazuma ulmifolia |
Pt |
BC (3) |
1997 |
1 |
1 |
20 |
|
Hymenaea courbaryl |
p/p |
IC |
1985 |
1 |
3 |
68 |
|
Machaerium villosum |
p/p |
BC (1) |
1992 |
1 |
1 |
13 |
|
Machaerium villosum |
p/p |
CFB |
1981 |
1 |
4 |
26 |
|
Myracrodruon urundeuva |
Pt |
BC (2) |
1993 |
1 |
1 |
23 |
|
Myracrodruon urundeuva |
p/p |
BFC |
1981 |
1 |
5 |
91 |
|
Myracrodruon urundeuva |
Pt |
BC |
1986 |
1 |
1 |
42 |
|
Myracrodruon urundeuva |
Pt |
BC |
1988 |
1 |
1 |
28 |
|
Myroxylon peruiferum |
p/p |
IC |
1984 |
1 |
4 |
45 |
|
Parapiptadenia rigida |
Pt |
BC (1) |
1991 |
1 |
1 |
25 |
|
Peltophorum dubium |
Pt |
BC (2) |
1992 |
1 |
1 |
26 |
|
Peltophorum dubium |
p/p |
BFC |
1985 |
2 |
2 |
28 |
|
Peltophorum dubium |
Pt |
BC |
1985 |
1 |
1 |
18 |
|
Persea condata |
Pt |
BC (1) |
1992 |
1 |
1 |
9 |
|
Prunus sellowii |
pt |
IC |
1989 |
1 |
1 |
17 |
|
Pterogyne nitens |
p/p |
BFC |
1989 |
1 |
4 |
36 |
|
Pterogyne nitens |
p/p |
BFC |
1989 |
1 |
5 |
93 |
|
Tabebuia heptaphylla |
pt |
BC |
1984 |
1 |
1 |
19 |
|
Tabebuia heptaphylla |
pt |
BC |
1985 |
1 |
1 |
15 |
|
Tabebuia vellosoi |
p/p |
BFC |
1986 |
1 |
1 |
18 |
RESULTADOS DE LOS ESTUDIOS GENÉTICOS
El Cuadro 2 presenta los resultados de los análisis de la estructura genética de las poblaciones conservadas, estimados empleando la metodología desarrollada por Hamrick (1976). La mayor parte de la variabilidad genética en todas las especies se encontró dentro de las poblaciones, especialmente entre individuos dentro de las progenies. Esto está de acuerdo con la mayoría de los estudios de estructura de poblaciones realizados basándose en los rasgos cuantitativos de especies forestales tropicales y templadas (Hamrick 1976; Khalil, 1985; Li et al., 1993; Moraes, 1993; Buliuckas et al. 1999, Rehfeldt, 1999; Sebbenn et al. 1999), y también en los datos con isoenzimas (Hamrick & Godt, 1990). Se cree que la escasa variación genética encontrada entre poblaciones obedece al pequeño número de poblaciones de cada especie e incluidas en los experimentos e indica la necesidad de incluir un mayor número de poblaciones en los bancos de conservación ex situ para conservar la variabilidad entre poblaciones. La falta de variabilidad genética entre poblaciones observada a nivel molecular parece indicar que, para fines de reproducción de árboles, la ventaja de seleccionar una u otra población puede ser limitada. Si se confirma esta observación, la selección del material a incluir en los huertos semilleros puede realizarse basándose en el resultado de las progenies a través de las poblaciones.
Se evaluó también la variabilidad genética dentro de las poblaciones, estimando la heredabilidad, el coeficiente de variación genética y la variación fenotípica entre la progenie. La heredabilidad en cuanto a la altura y el diámetro era generalmente reducida. Las anomalías encontradas en las estimaciones de los parámetros genéticos obedecen posiblemente al uso de un escaso número de progenies por población y a unos supuestos básicos defectuosos sobre genética cuantitativa para la estimación de los parámetros genéticos (véase p.ej. Nogueira et al., 1986b, Siqueira et al., 1993, Sebbenn et al., 2000). A este respecto, Moraes (1993), al estudiar los sistemas reproductivos en poblaciones de Myracrodruon urundeuva basándose en las isoenzimas, observó que la tasa de cruzamiento lejano de la especie era de 0,49. Posteriormente se encontró que esta especie tiene un comportamiento reproductivo mixto (exogamia y endogamia) lo que subraya la necesidad de información básica sobre los sistemas reproductivos, como sugiere Ritland (1989) o Surles et al. (1990). Sebbenn (2000), al comparar la estimación de los parámetros genéticos en poblaciones de Cariniana legalis utilizando (i) el modelo clásico, y (ii) incorporando los principios de Ritland, observó unas estimaciones exageradas en los coeficientes de capacidad hereditaria hasta del 33% cuando se usan los modelos clásicos. Hay pruebas muy evidentes, según la bibliografía, de que la mayoría de las especies forestales nativas de Brasil tienen un comportamiento reproductivo mixto (véase p.ej. Paiva, 1993; Lepsch-Cunha, 1996; Sebbenn et al., 1998; Souza, 1997; Moraes, 1997; Oliveira, 2000). Por lo tanto, está claro que los parámetros genéticos en las especies forestales nativas no pueden estimarse sin sentido crítico basándose en los modelos genéticos clásicos de carácter cuantitativo.
Cuadro 2. Proporción de variación
fenotípica dentro de progenies
( ), variación genética
entre progenies dentro de poblaciones
(
), variación genética
entre progenies dentro de poblaciones
(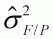 ) y variación genética
entre poblaciones (
) y variación genética
entre poblaciones ( ) para algunas
especies de árboles tropicales de conservación ex situ en
el IFSP, Brasil.
) para algunas
especies de árboles tropicales de conservación ex situ en
el IFSP, Brasil.
|
Especie |
Rasgo |
Pob. |
|
|
|
Referencia |
|
Balfourodendron riedelianum |
DAP |
3 |
84,1 |
12,7 |
3,1 |
Siqueira et al. (2000) |
|
ALT |
3 |
81,5 |
11,8 |
6,7 |
|
|
|
|
|
|
|
|
|
|
|
Pterogyne nitens
|
DAP |
4 |
91,9 |
7,7 |
0,4 |
Sebbenn et al. (2000) |
|
ALT |
4 |
84,7 |
8,7 |
6,7 |
|
|
|
|
|
|
|
|
|
|
|
Peltophorum dubium
|
DAP |
2 |
80,9 |
17,8 |
1,2 |
Sebbenn et al. (2000) |
|
ALT |
2 |
74,4 |
24,2 |
1,5 |
|
|
|
|
|
|
|
|
|
|
|
Cariniana legalis
|
FF |
3 |
91,6 |
2,4 |
6,0 |
Sebbenn (2000) |
|
DAP |
3 |
96,5 |
3,4 |
0,1 |
|
|
|
ALT |
3 |
97,3 |
2,6 |
0,1 |
|
|
|
VC |
3 |
85,7 |
1,5 |
12,8 |
|
El Bosque Atlántico de Brasil es uno de los mayores patrimonios de diversidad biológica forestal del planeta, conteniendo muchas áreas más de 400 especies arbóreas por hectárea; el 90% de las especies se encuentran con frecuencias de un solo individuo por hectárea (Kageyama et al, 1997). Sin embargo, como se indicó antes, las áreas de conservación ex situ del IFSP incluyen sólo 24 especies, habiéndose incluido menos de cinco poblaciones para la mayoría de ellas. Todas las áreas y experimentos ex situ fueron establecidos con recursos procedentes del IFSP pero ha habido una reducción gradual en las inversiones financieras del Gobierno del Estado en la institución, habiéndose reducido drásticamente en los últimos años la inclusión de nuevas especies: el 73% de los 41 experimentos se establecieron antes de 1990.
Es necesario aumentar el número de poblaciones de las especies ya establecidas en los rodales ex situ para mejorar la cobertura de la variación genética conservada. Existe además la necesidad de continuar evaluando periódicamente el material contenido en los bancos de conservación ex situ, para lograr un mejor conocimiento de las características genéticas y del comportamiento selvícola de las especies. En tercer lugar, es necesario considerar la transformación de los experimentos en huertos semilleros, utilizando una selección de baja intensidad a fin de mantener poblaciones efectivas de gran dimensión. Finalmente, existe la necesidad de aumentar el número de especies bajo conservación ex situ e incluir una variedad de poblaciones de cada nueva especie incluida. Una muestra de 30 poblaciones, representada cada una por 25 progenies y 30 plantas por progenie establecidas como mínimo en tres localidades, cumpliría probablemente los objetivos de conservación y garantizaría la conservación también de alelos raros (frecuencia < 5%).
Se han identificado nuevas especies candidatas a incluir, basándose en dos criterios: (i) existencia en el estado de Sao Paulo, y (ii) valor económico. Estas especies están incluidas en el Cuadro 3 de acuerdo con los grupos sucesionales a que pertenecen.
Cuadro 3: Lista de nuevas especies candidatas a incluir en el nuevo programa de conservación ex situ.
|
Especie |
Nombre común |
Familia |
|
Especies pioneras |
|
|
|
Cecropia glaziovi Snethl. |
Embaúbavermelha |
Cacropiaceae |
|
Cecropia hololeuca Miquel |
Imbaúba prateada |
Cacropiaceae |
|
Clethra scabra Pers. |
Carne de vaca |
Clethraceae |
|
Croton floribundus Spreng |
Capixingui |
Euphorbiaceae |
|
Piptadenia paniculata Bentham |
Angico |
Mimosaceae |
|
Especies secundarias |
|
|
|
Cabraela canjerana |
Canjarana |
Meliaceae |
|
Cariniana estrellensis |
Jequitibá branco |
Lecithidacea |
|
Cedrela fissilis Vellozo |
Cedro |
Meliaceae |
|
Centrolobium robustum (Vell.) Martius ex Bentham
|
Araribá rosa |
Fabaceae |
|
Centrolobium tomentosum Guillem ex Bentham
|
Arauva |
Fabaceae |
|
Chlorophora tinctoria (Linnaeus) Gaudichaud
|
Taiúva |
Moraceae |
|
Chorisia speciosa (A. S. Hil.) Dawson |
Paineira |
Bombacaceae |
|
Cinnamomum glaziovii (Mez) Kosterm |
Garuva |
Lauraceae |
|
Copaifera langsdorffii Desf. |
Bálsamo |
Caesalpinaceae |
|
Copaifera trapezifolia Hayne |
Copaíba |
Caesalpinaceae |
|
Cryptocarya moschata Nees & Martius ex Nees
|
Canela branca |
Lauraceae |
|
Dalbergia brasiliensis Vogel |
Jacaranda |
Fabaceae |
|
Erytrina falcata Benth. |
Corticeira |
Fabaceae |
|
Inga striata |
Inga |
Mimosoidae |
|
Joannesia princeps Vell. |
Boleira |
Euphorbiaceae |
|
Lafoensia pacari A. Saint- Hilaire |
Dedaleiro |
Lythraceae |
|
Luehea divaricata Martius & Zuccarini
|
Açoito a cavalo |
Tiliaceae |
|
Machaerium scleroxylon Tulasne |
Caviuna |
Fabaceae |
|
Nectandra lanceolata Nees et Matius ex Nees.
|
Canela-amarela |
Lauraceae |
|
Platypodium elegans Vog. |
Amendoin do campo |
Fabaceae |
|
Podocarpus lambertii Klotzsch ex Endl.
|
Pinheiro bravo |
Podocarpaceae |
|
Talauma ovata A. Saint- Hilaire |
Baguaçu |
Magnoliaceae |
|
Especies climácicas |
|
|
|
Calophyllum brasiliensis Combessedes |
Guanandi |
Clusiaceae |
|
Casealpinia leiostachya Ducke |
Pauferro |
Caesalpiniaceae |
|
Dalbergia nigra (Vell.) Freire Allemão ex
Bantham |
Jacaranda da Bahia |
Fabaceae |
|
Holocalyx balansae Micheli |
Alecrim |
Fabaceae |
|
Laplacea fruticosa (Schrader) Kobuski |
Santa Rira |
Theaceae |
|
Ocotea catharinensis Mez |
Canela-preta |
Lauraceae |
|
Ocotea odorifera (Vell.) Rohwer |
Canela-sassafrás |
Lauraceae |
|
Ocotea porosa (Nees ex Martius) Libarato Barroso
|
Imbuia |
Lauraceae |
|
Plathymenia reticulata Benth. |
Amarelinho |
Mimosaceae |
|
Tabebuia impetiginosa |
Ipê-rosa |
Bignoniaceae |
Los autores agradecen a los jefes y directores de las Estaciones Experimentales del IFSP que han estado a cargo del mantenimiento de los bancos de conservación; y a los equipos de evaluación de campo que han garantizado la gran calidad y eficiencia de la recolección de datos. Agradecemos también a Girlei Costa da Cunha, Ingeniero Forestal, por la lectura del manuscrito y las valiosas sugerencias de nuevas especies a incluir en el banco de conservación del IFSP.
REFERENCIAS
Buliuckas, V.; Ekberg, I.; Eriksson, G. & Norell, L. Genetic variation among and within populations of four Swedish hardwood species assessed in a nursery trial. Silvae Genetica, Frankfurt, 48(1): 17-25, 1999.
Ettori, L.C.; Siqueira, A.C.M.F.; Nogueira, J.C.B.; Ferreira, A.B. & Zanatto, A.C.S. Conservação ex situ dos recursos genéticos do Ipê-amarelo (Tabebuia vellosoi Tol.) através de testes de procedências e progênies. Rev. Inst. Flor., São Paulo, 7(2): 157-168., 1995.
Ettori, L.C.; Siqueira, A.C.M.F.; Sato, A.S. & Campos, O.R. Variabilidade genética em populações de Ipé-roxo - Tabebuia heptaphylla (Vell.) Tol. - para conservação ex situ. Rev. Inst. Flor., São Paulo, 8(1): 61-70, 1996.
Ettori, L.C.; Siqueira, A.C.M.F.; Zanatto, A.C.S. & Boas, O.V. Variabilidade genética em duas populações de Cordia trichotoma. Rev. Inst. Flor., São Paulo, 11(2): 179-187, 1999.
FAO. 1998. FAO Panel of Experts on Forest Gene Resources: Tenth Session, Rome, Italy, 9-11 September 1997. FAO, Rome, 68 p.
FAO. 2000. FAO Panel of Experts on Forest Gene Resources: Eleventh Session, Rome, Italy, 29 September to 1 October 1999. FAO, Rome, 90 p.
Giannotti, E.; Timoni, J.L.; Mariano, G.; Coelho, L.C.C.; Fontes, M.A. & Kageyama, P.Y. Variação genética entre procedências e progênies de Araucaria angustifolia (Bert.) O. Ktze. In: Congresso National sobre Essëncias Nativas, Campos do Jordão, 1982. Anais... Silvicultura em São Paulo, São Paulo, 16A: 970-975. Pt. 2. (Edição especial).
Gurgel Garrido, L.M.A.; Siqueira, A.C.M.F.; Cruz, S.F.; Romanelli, R.C.; Ettori, L.C.; Crestana, C.S.M.; Silva, A.A.; Morais, E.; Zanatto, A.C.S. & Sato, A.S. Programa de melhoramento genético florestal do Instituto Florestal de São Paulo (Acervo). IF Série Registros, São Paulo, 18: 1-53, 1997.
HamricK, J.L. & Godt, M.J.W. Allozyme diversity in plant species. In: Brown, A.H. D.; Clegg, M.T.; Kahler, A.L. & Weir, B.S. (Eds) Plant population genetics, breeding and genetic resources. Sinauer, Sunderland, Massachusetts, p. 43-63, 1990.
HamricK, J.L. Variation and selection in western montane species II. Variation withing and between populations of White Firon an elevation transect. Theor. Appl. Genet, 47: 27-34, 1976.
IFSP. Intituto Florestal de São Paulo. São Paulo, Brasil, IF-Série Registros No. 12, 1994, 31 p.
Kageyama, P.Y. Gandara, F. & Borges, A. Pesquisa e recuperação de áreas degradadas na Mata Atlântica - Avaliação bibliográfica. In: Recuperação de áreas degradadas na Mata Atlântica. CNBR, São Paulo, p. 11 a 16. 1997.
Khalil, M.A. Genetics of wood characteres of black spruce [Picea mariana (Mill.) B.S.P.] in Newfoundland, Canada. Silvae Genetica, Frankfurt, 34(6): 221-230, 1985.
Lepsch-Cunha, N. Variabilidade Genética Intrapopulacional de Duas Espécies de Baixa Densidade na Amazônia Central: Couratari multiflora (J.E. Smith) Eyma e Couratari guianensis Aublet. Item 1. de Resultados da Dissertação “Estrutura Genética e Fenologia de Espécies Raras de Couratari spp. (Lecythidaceae) na Amazônia Central”. ESALQ/USP, Piracicaba, SP. 1996, 121p. (Dissertação de mestrado)
Li, P.; Beaulieu, J.; Corriveau, A. & Bousquet, J. Genetic variation in juvenile growth and phenology in a White Spruce provenance-progeny test. Silvae Genetica, Frankfurt, 42 (1): 52-60, 1993.
Moraes, M.L.T. Variabilidade genéica por isoenzimas e caracteres quantitativos em duas populações naturais de aroeira Myracrodruon urundeuva F.F. & M.F. Allemão Anacardiaceae (Syn: Astronium urundeuva (Fr. Allemão) Engler. ESALQ/USP, Piracicaba, SP, 1993, 139 p. (Tese de doutorado)
Moraes, P.L.R. Estrutura genética de populações de Cryptocarya moschata Nees & Martius ex Nees (Lauraceae). UNESP, Rio Claro, SP, 1997, 190 p. (Tese de doutorado)
Mori, E.S. Estrutura genética de populações narurais de canafístula Peltophorum dubium (Spreng.) Taub. UNESP, Botucatu, São Paulo, 145 p. 2000. (Tese de livre docência).
Nogueira, J.C.B.; Siqueira, A.C.M.F. & Zanatto, A.C.S. Teste de progênies e procedências do pau d”alho Gallesia gorarema Vell. Moq. Boletim Técnico do IF, São Paulo, 40A: 344-356, 1986a.
Nogueira, J.C.B.; Siqueira, A.C.M.F.; Moares, E. & Ywane, M.S.S. Teste de progênies e procedências da aroeira Astronium urundeuva (Fr. All.) Engl. Boletim Técnico IF, São Paulo, 40A: 367-377, 1986b.
Oliveira, A.F. Estrutura genética de populações naturais de Copaifera langsdorffii Desf. a partir de isoenzimas. UFLA, Lavras, MG, 2000, 115 P. (Dissertação de mestrado)
Paiva, J.R. Variabilidade isoenzimática em populações naturais de seringueira [Hevea brasiliensis (Willd) ex Adr. de Juss.] Müell. Arg. ESALQ/USP, Piracicaba, 1992, 144 p. (Tese de doutorado)
Rehfeldt, G.E. Systematics and genetic structure of Washoe Pine: Applications in Conservation genetics. Silvae Genetica, Frankfurt, 48(3-4): 167-173, 1999.
Ritland, K. Correlated matings in the partial selfer Mimulus guttatus. Evolution, 43(4): 848-859, 1989.
Sebbenn, A. M.; Durigan, J.; Siqueira, A. C. M. F.; Pontinha, A. A. S. Teste de procedências de Grevilha robusta A. Cunn. . Rev. do Inst. Flor, SP, 11(1): 65-73, 1999.
Sebbenn, A. M.; Kageyama, P. Y. & Vencosvky, R. Variabilidade genética, sistema reprodutivo e estrutura genética espacial em Genipa americana L. através de marcadores isoenzimáticos. Scientia Forestalis, 53: 15-30, 1998b.
Sebbenn, A. M.; Siqueira, A. C. M. F.; Kageyama, P. Y. & Di Dio J., O. P. Estrutura genética de Pterogine nitens Tul. (Caesalpinaceae) através de caracteres quantitativos. Scientia Forestalis, Piracicaba, São Paulo, 2000. (no prelo)
Sebbenn, A.M. Estudo de populações de Cariniana legalis (Mart.) O. Ktze, por caracteres quantitatívos e isoenzimas. ESALQ/USP, Piracicaba, SP, 209 p. 2000. (Tese de doutorado)
Sebbenn, A.M.; Siqueira, A.C.M. F.& Gurgel Garrido, L. M. A. Interação progênies x locais e variabilidade genética em Jequitibá-Rosa - Cariniana legalis (Mart.) O. Ktze. Rev. do Inst. Flor. SP, 12(1): 13-23, 2000.
Sebbenn, A.M.; Siqueira, A.C.M.F. Vencovsky, R. & Machado, J.A.R. Interação genótipo ambiente na conservação ex situ de Peltophorum dubium, em duas regiões do Estado de São Paulo. Rev. Inst. Flor, SP, 11(1): 75-89, 1999.
Siqueira, A.C.M.F.; Nogueira, J.C.B. & Kageyama, P.Y. Conservação de recursos genéticos ex situ de cumbaru (Dipteryx alata Vog.) - Leguminosae. Rev. do Inst. Flor., São Paulo, 5(2): 231-43, 1993.
Siqueira, A.C.M.F.; Sebbenn, A.M.; Garrido, L.M.A.G.; Castollen, M.S. & Zanatto, A.C.E. Comportamento silvicultural e genético de duas espécies arbóreas tropicais secundárias. Rev. do Inst. Flor., SP, 11(1): 53-64, 1999.
Siqueira, A.C.M.F.; Sebbenn, A.M.; Kageyama, P.Y. & Ettori, L.C. Distribuição da variação genética entre e dentro de populações de Balfourodendron riedelianum (Engler) Engler para a conservação ex situ. Rev. do Inst. Flor, São Paulo, 2000. (no prelo)
Souza, L.M.F.I. Estrutura genética de populações naturais de Chorisia speciosa St. Hil. (Bombacaceae) em fragmentos florestais na região de Bauru (SP) - Brasil. ESALQ/USP, Piracicaba, SP, 1997, 76 p. (Dissertação de mestrado).
Surles, S.E.; Arnold, J.; Schnabel, A.; Hamrick, J.L. & Bongarten, B.C. Genetic relatedness in open-pollinated families of two leguminous tree species, Robinia pseudoacacia L. and Gleditsia triacanthos L. Theor. Appl. Genet, 80: 49-56, 1990.
Victor, M. A devastação florestal. Sociedade Brasileira de Silvicultura, São Paulo, 28-32, 1975.
![]()
![]()
![]()
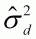 (%)
(%)