![]()
![]()
![]()
ÅKE GUSTAFSSON Y FRANÇOIS MERGEN
La citogenética proporciona las bases de la variación inherente y es esencial para determinar la constitución cromosómica y la naturaleza de la variación genética de las especies arbóreas. ¿Qué papel desempeña la poliploidia y los cambios estructurales de cromosomas en variedad es y especies? ¿Qué diferencias genéticas resultan de cambios en el equilibrio cromosómico o de cambios estructurales y en qué medida se debe la variación a mutaciones «puntuales»?
Los géneros de gimnospermas con valor dasonómico tienen cromosomas grandes, la poliploidia es rara y los cariotipos dentro de los géneros son similares. En las Pinaceae los números básicos de cromosomas son 10, 11, 12 y 13, lo que ha inducido a Khoshoo a sacar en conclusión que el número básico original era 10 y que los números básicos mayores han surgido por fragmentación de cromosomas. Esto aparece claro en Pseudolarix y Podocarpus. El número básico de 12 pares de cromosomas es característico, sin embargo, de la mayoría de los géneros de las Pinaceae, siendo posible que los géneros con números básicos inferiores se hayan originado por una unión de cromosomas derivada de una translocación asimétrica. Se ha visto que la variación del número básico de cromosomas en ciertos géneros de angiospermas proviene de ambos fenómenos: fragmentación y unión de cromosomas.
Es rara la poliploidia en las Pinaceae, si bien los poliploides se dan en la naturaleza y pueden ser inducidos. Mergen descubrió que tanto los poliploides espontáneos como inducidos, se retrasan en crecimiento y tienen una baja supervivencia. Los prolijos experimentos de Hyun sobre poliploides inducidos en coníferas proporcionan nuevos datos necesarios.
ÅKE GUSTAFSSON es Director del Instituto de Genética Forestal, El Real Colegio de Montes, Estocolmo, Suecia. FRANÇOIS MERGEN es Profesor de Genética Forestal, Universidad de Yale, Escuela de Montes, New Haven, Connecticut, E.U.A. Otros miembros del equipo de trabajo fueron G. Sirén (FAO) y K. Sax (E.U.A.), Jefe de la Sección 1 de la Consulta quien preparó el resumen.
El gran tamaño de los cromosomas en las Pinaceae las hace muy sensibles a la radiación ionizante. Hace ya muchos años que Gustafsson averiguó que dosis relativamente bajas de rayos X dañaban las semillas de pino. Estos hechos deben ser tenidas en cuenta en la mejora genética por mutación y al comparar el valor relativo de la mutagénesis por radiación y química.
Contrastando con las gimnospermas, los géneros de angiospermas de valor forestal varían mucho en el número básico de cromosomas que va de 6 a 41 pares. Los cromosomas son relativamente pequeños y la poliploidia es frecuente. La poliploidia inducida puede ser interesante en las especies forestales de estos géneros como lo es para producir nuevos tipos de árboles y arbustos ornamentales.
Uno de los aspectos más prometedores de la genética forestal es la producción de híbridos F entre especies de habitats distintos. Especies de coníferas que han estado aisladas geográficamente por miles de generaciones pueden ser cruzadas y han producido híbridos vigorosos y relativamente fértiles, como lo demuestran los cruces entre el alerce europeo y el japonés (Larix decidua y L. leptolepis) y entre las especies de pinos blancos asiáticos y americanos. También se ha comprobado un vigor híbrido semejante entre especies americanas y europeas de Populus y Platanus.
El empleo de la radiación ionizante como agente mutagénico ha sido de gran valor en la mejora de las plantas agrícolas, a pesar de que las mutaciones deseables sean raras. Pero el largo ciclo vital de las especies forestales y el espacio necesario para probar el vigor y calidad de los mutantes deseados hace de la mejora por mutación un programa largo y caro. Quizá la selección en masa al tiempo de germinar la semilla pueda aplicarse cuando se seleccione por resistencia a la enfermedad, tal como se hizo con tanto acierto con los cereales tratando las semillas en germinación con la toxina del patógeno. La correlación entre la sensibilidad a los rayos X y el vigor inherente en las semillas de Zea, de reciente descubrimiento, puede servir para seleccionar los segregados genéticos superiores en poblaciones híbridas de árboles forestales.
Todos los organismos, aunque su estado de desarrollo sea inferior al cromosoma, están caracterizados por dos propiedades, a saber: repetición o constancia genética y mutación o cambio genético. A consecuencia de la mutación, los organismos están representados en la naturaleza por poblaciones de genotipos. Desde el comienzo de la vida existe una interacción entre la herencia y el medio ambiente. Las condiciones ambientales en que se desarrollan los organismos vivos están sujetas a cambio. Como diferentes poblaciones contienen individuos más o menos bien adaptados a sus ambientes específicos, la composición genética de las poblaciones se altera a consecuencia de la selección natural. Si la presión del ambiente rebasa las posibilidades genéticas de una población o especie, ésta desaparecerá y será reemplazada por otras.
La composición genética de una población de organismos superiores cambia a causa de la mutación y la recombinación. En la mutación, los genes o moléculas de la herencia experimentan cambios de distinta naturaleza y con frecuencia en su orden normal dentro del cromosoma. Los cromosomas forman grupos de ligamiento que limitan las posibilidades de recombinación pero, a causa del sobrecruzamiento, los grupos de ligamiento se rompen, aumentando así las posibilidades de recombinación durante la reproducción sexual.
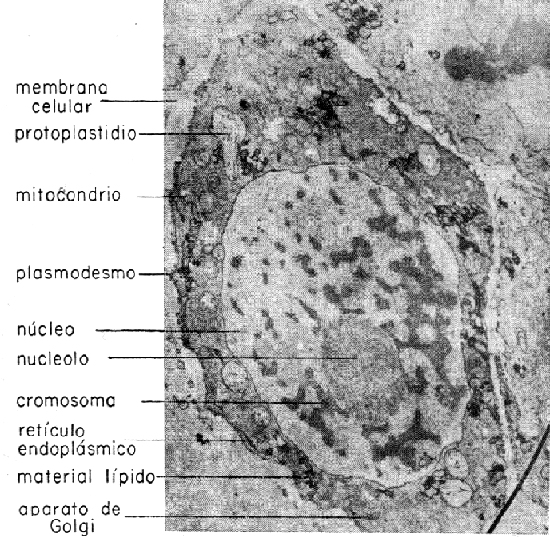
Al diferenciarse las células, el material hereditario quedó concentrado en los cromosomas, en el núcleo, mientras que las actividades fisiológicas y de desarrollo se centraron en el citoplasma (Figura 1). Esto no implica inactividad fisiológica por parte del núcleo, ni tampoco falta de información hereditaria en el citoplasma; en realidad los genes de los cromosomas son el origen de instrucciones bioquímicas para el citoplasma, existiendo en éste «plasmogenes» o «plastogenes» que actúan como unidades hereditarias independientes.
Este capítulo trata de los árboles forestales y su comportamiento citogenético. De lo dicho en la introducción se deduce que la historia en el pasado de una especie o género es de importancia básica para su constitución genética actual, su estructura poblacional, su polimorfismo y distribución. También hay diferencias fundamentales entre especies, géneros y familias en sus caracteres citológicos. Las diferencias en comportamiento cromosómico guardan relación con:
1. el número básico de cromosomas;
2. tamaño y estructura cromosómica; y
3. grado de poliploidia.
Estos problemas han sido discutidos en detalle por Gustafsson (1960a), con referencia a los trabajos que iniciaron la investigación sobre citología del árbol debidos a K. y H. J. Sax a principios de la década 1930-40.
Número básico de cromosomas
En las gimnospermas los números básicos (x) son 7 (Ephedra), 8 y 9 (varios géneros de Cycadaceae), 10 (Sciadopitys), 11, 12 y 13 (numerosos géneros); pueden también alcanzar valores más altos, como 19 y 20 (Podocarpus) y 22 (Pseudolarix). Estos números últimamente mencionados son sin duda secundarios, esto es, derivados, y pueden deberse en ciertos casos a alguna clase de rotura o translocación cromosómica.
Los géneros de angiospermas arbóreas pertenecen a varias familias diferentes del sistema natural. En consecuencia, las diferencias cromosómicas son complejas y variadas. Los números básicos van desde 6 y 7 (Cassia), 8 (Carpinus), 9 (Hevea), 10 (Cornus), 11 (Corylus y Eucalyptus), 12 (Quercus), 13 (Acer y Ficus), 14 (Alnus y Betula) hasta 19 (Populus), 21 (Platanus), 23 (Fraxinus) y 41 (Tilia).
Los géneros de gimnospermas y angiospermas difieren también con respecto al tamaño medio de cromosoma. Esto no significa que las angiospermas de cromosomas pequeños contengan menor número de genes que las gimnospermas de cromosomas largos. El hecho sólo indica que el material extragénico de los cromosomas de gimnospermas ha adquirido un desarrollo extraordinario. Hay factores selectivos que aparentemente pueden incrementar o disminuir el tamaño de los cromosomas, de acuerdo con el tipo de ambiente o, como indica el botánico danés Raunkiaer, con la forma de vida de la especie. Véase, por ejemplo, Babcock (1947), Stebbins (1950) y Gustafsson (1951, 1960a).
Stebbins ha señalado que muchas angiospermas leñosas poseen cromosomas menores que los géneros y especies herbáceas con ellas emparentados. Esta diferencia puede depender de que el leño de las angiospermas contiene fibrocélulas originadas por células iniciales del cambium de tamaño pequeño, o, de acuerdo con Stebbins, de la presunción (verdadera o falsa) de que las plantas leñosas requieren en general un sistema genético con una cantidad máxima de recombinación génica, lo que es favorecido por la presencia de muchos cromosomas pequeños. Las gimnospermas principales, las Coniferales, carecen de fibras en el leño y las células iniciales del cambium tienen tamaño igual. Como ya hemos visto, el número básico de cromosomas es más bien alto, aunque bastante menor aún que en las angiospermas, siendo los cromosomas de gran tamaño. Es de presumir, dice Stebbins, que la reducción en el tamaño de los cromosomas tuvo lugar desde muy pronto en la evolución de las angiospermas leñosas.
Hay una conexión interesante entre el tamaño de los cromosomas y la radiosensibilidad, puesta de manifiesto en años recientes por el trabajo de Sparrow y colaboradores. Sparrow y Miksche (1961) hallaron una correlación positiva buena entre la inhibición del crecimiento por radiación ionizante y el tamaño de los núcleos interfásicos en los meristemos de los brotes. Comparando la radiosensibilidad de Populus, Pinus y Picea, Gustafsson y Simak (1958) y Wettstein et al. (1959) comprobaron el efecto del tamaño de los cromosomas en la radiosensibilidad de especies arbóreas forestales. Los miembros de las Pinaceae tienen núcleos grandes comparados con los de las angiospermas y el volumen nuclear de los pinos es aproximadamente cinco veces el de los robles (Quercus spp.) a pesar de que ambos géneros tienen un número somático de 2n = 24. Esto podría explicar la mayor radiosensibilidad de las Pinaceae.
Desde que Gustafsson y Simak (1958) comunicaron la gran sensibilidad del pino y el abeto a la radiación, se han acumulado muchos datos. Pedigo (1960, 1962) y Platt (1963) describieron los efectos en Pinus taeda, Sparrow et al. (1963) en Pinus strobus, Mergen y Stairs (1962) en Pinus rígida, y Brandenburg et al. (1962) en Pinus monophylla.
Ofrecen un interés considerable los efectos en los pinos de la radiación crónica a bajo nivel. Arboles de Pinus rígida murieron tras una exposición de cinco años a rayos g en dosis no mayores de 8 r por día durante ocho meses cada año. Arboles de Quercus expuestos en las mismas condiciones sobrevivieron a esta radiación gamma crónica, probablemente debido a su volumen nuclear menor. Sin embargo la descendencia R1 de ambos géneros fue mucho más variable que la testigo, reflejando efectos genéticos que habían ocurrido. en los árboles irradiados crónicamente. Esto indica que puede darse en los árboles una cantidad de cambios genéticos y fisiológicos a consecuencia de una radiación acumulativa crónica de bajo nivel.
Poliploidia
Pocos géneros de gimnospermas contienen especies poliploides. Ejemplos son Ephedra, Gnetum y Welwitschia. El Juniperus chinensis es especie tetraploide y la Sequoia sempervirens es una especie exaploide de origen natural. Aunque la poliploidia no desempeña un papel importante entre las formas de variación de las Pinaceae se encuentran ocasionalmente plantas poliploides en los viveros y en la naturaleza. Se ha encontrado condición aneuploide y mixoploide en brinzales enanos de Picea abies (Kiellander, 1950; Illies, 1959) y tetraploide en un brinzal gemelo de Aojes firma (Kanezawa, 1949). Hay informes aislados sobre poliploidia natural en Larix. Christiansen (1950) localizó un tetraploide adulto de Larix decidua, y Chiba y Watanabe (1952) encontraron entre repicados de dos años de Larix leptolepis ocho plantas poliploides - dos tenían raíces diploides, mientras que las seis restantes eran todas tetraploides. Un ejemplar originado por cruzamiento entre Larix decidua y L. occidentalis era triploide (Larsen y Westergaard 1938). Se ha comunicado la existencia de ejemplares poliploides o mixoploides espontáneos en cuatro especies de pino. Zinnai (1953) localizó cinco plantitas tetraploides de Pinus densiflora; en el Pinus elliottii Mergen (1958) describió plantitas mixoploides con 2n, 3n y 4n complementos cromosómicos; la presencia de poliploidia en Pinus sylvestris en Suecia fue comunicada por Johnsson (1959); y Nishimura (1960) describió una planta tetraploide de Pinus thunbergii nacido de una semilla con dos embriones.
Por tratamiento con colchicina (Margen, 1959) se han formado poliploides en especies gimnospermas. En todas las especies en que se intentó, se consiguió inducir la poliploidia y los cambios en los brinzales o árboles fueron similares. En general, las acículas fueron más cortas y gruesas, el número de células se redujo, la ramificación fue más basta, la floración se inhibió y fue frecuente en los árboles el enanismo. Los tetraploides son poco deseables en la mayoría de las especies, pero tienen aplicación como paso intermedio en la producción de triploides.
Se ha intentado saltar la fase de esporofita tetraploide tratando estróbilos de microesporangios de Larix leptolepis (Illies, 1956) y Pinus nigra y Pinus mugo (Margen, 1959) durante la microesporogénesis. Se produjeron granos de polen diploides pero no se dispone de resultados relativos a la descendencia de este tipo de polen. El método es prometedor, sin embargo, y no cabe duda que recibirá más atención. Gustafsson (1960a), Mehra (1960) y Mergen (1963) dan más información sobre poliploidia en gimnospermas.
La poliploidia natural se origina a menudo después de la hibridación entre especies diferentes o poblaciones equivalentes a especies y subsiguiente doblado del número de cromosomas, debido a la formación de gametos no reducidos (condición conocida como anfiploidia o alopoliploidia). Tal tipo de poliploidia es común entre las angiospermas cultivadas (Nicotiana, Gossypium, Triticum, Brassica) aunque también en especies silvestres (Galeopsis, Rubus, Poa), pudiendo formarse artificialmente nuevas combinaciones como por ejemplo Triticale, que es un nuevo «género» combinación de Triticum y Secale. En otros muchos casos la poliploidia es de origen intraespecífico (el término empleado es autopoliploidia). Los límites entre la alo- y la autopoliploidia no son rígidos (Müntzing, 1936). El género Dactylis es interesante a este respecto pues los tetraploides naturales son considerados a menudo como poliploides interespecíficos, aunque los correspondientes diploides están sin duda muy emparentados (Müntzing, 1956).
Un caso interesante fue reseñado por Wright (1959a) en fresno blanco, Fraxinus americana, que se divide en tres «ecotipos», uno septentrional (2n = 46), uno intermedio (conteniendo poliploides) y uno meridional (con 2n = 46, 92, 138). El fresno Pumpkin, Fraxinus tomentosa, es una especie exaploide rara (&= 138), derivada probablemente de un cruce entre un fresno verde diploide y un fresno blanco tetraploide (Wright, 1959b). De acuerdo con estos hechos, en el Fraxinus se dan ambas formas, la autopoliploidia y la anfiploidia.
La característica más sobresaliente de las razas poliploides intraespecíficas consiste en los cambios del ritmo de desarrollo y de exigencias ecológicas, como fue explicado primeramente por Müntzing (1936) y luego, con más extensión, por Stebbins (1950, 1956) y por Müntzing (1956, 1959). La apariencia generalmente «gigas», el aumento del crecimiento vegetativo, la alteración de reacciones de incompatibilidad y los cambios en exigencias ecológicas permiten a los autopoliploides extender las áreas de cultivo de las variedades diploides. Tales poliploides tienen aplicación también en la mejora genética forestal, idea defendida por Nilsson-Ehle cuando hace ya tiempo estudió las reacciones y potencialidades del autotriploide gigante del Populus tremula, En las gimnospermas, así como en muchas frondosas, los alopoliploides pueden llegar a ser más adecuados para el uso directo en la práctica que los autopoliploides.
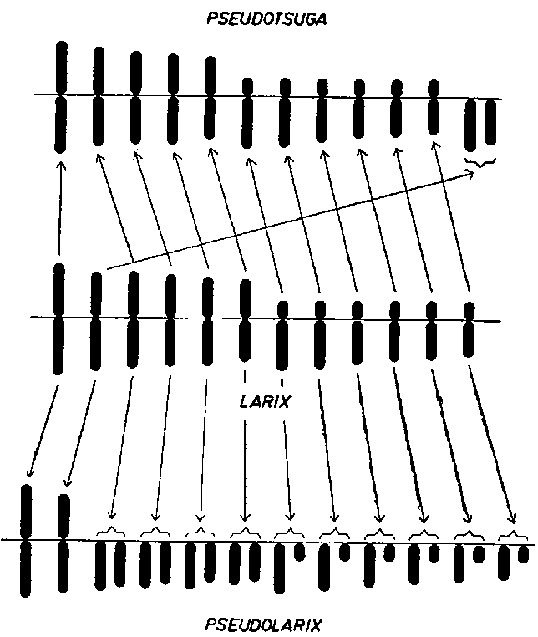
FIGURA 2. - Evolución cariotípica: cambios cromosómicos durante la evolución ilustrados por los cariotipos (idiogramas) de Pseudotsuga, Larix: y Pseudolarix.
Disploidia y poliploidia secundaria
El término disploidia (Chiarugi, 1932) significa que en un género se dan diferentes números básicos, relacionados filogenéticamente. Dicho término, aunque ha sido poco empleado, es muy útil. En géneros arbóreos no es rara la disploidia. Por ejemplo, en los géneros Cycas y Microcycas se encuentran números básicos de 11, 12 y 13 que probablemente deben derivar unos de otros. En Podocarpus con números básicos de 11, 12, 19 y 20 debe ocurrir alguna especie de rotura cromosómica que conduzca a la disploidia. El género australiano Casuarina muestra x= 8, 9, 10, 11, 12, 13, y así sucesivamente, según Barlow (1959). Aún más interesante desde un punto de vista evolucionista es el género Pseudolarix, relacionado con Larix, en el que parece posible determinar los acontecimientos filogenéticos por medio del análisis de cromosomas (Figura 2). Pseudolarix amabilis con x = 22 tiene dos pares de cromosomas con los centrómeros subcentrales o centrales y 20 pares con centrómeros casi terminales. En Larix todos los cromosomas tienen centrómeros centrales, subcentrales o subterminales. Es muy posible que el Pseudolarix se haya originado del Larix por medio de rotura de 10 cromosomas de la serie haploide en las proximidades del centrómero, mientras que dos permanecieron sin cambios (véase Mergen, 1961).
De acuerdo con Barner y Christiansen (1962) la Pseudotsuga taxifolia tiene 13 pares de cromosomas, de los cuales los dos más cortos son «telocéntricos» es decir, con centrómero terminal o casi terminal. Si consideramos estos dos cromosomas como derivados de un solo cromosoma largo, con centrómero central o subcentral, el idiograma resulta muy semejante al de Larix. Existen además otras peculiaridades genéricas que sugieren una relación filogenética entre ambos géneros relativamente estrecha.
Con seguridad, cuando los idiogramas de las especies y géneros de gimnospermas hayan sido determinados con más detalle, será posible sacar una serie de conclusiones relativas a la conexión entre las alteraciones cromosómicas y la filogenia.
Núcleo y citoplasma
En la introducción se señaló que también en el citoplasma existen constituyentes que pueden ser considerados como autorrepetidores y en cierto sentido independientes de los genes nucleares. A esto se debe la diferencia entre los términos genomio (suma de genes nucleares, Winkler, 1920) y plasmon (suma de factores hereditarios del citoplasma, Wettstein, 1926), así como el término menos claro plastom (factores hereditarios de los plastidios, Renner, 1934; Michaelis, 1957/58, y Gustafsson y Wettstein, 1957/58). Tiene un interés general la naturaleza plasmática de la esterilidad masculina en especies hermafroditas tales como el maíz, caña de azúcar, remolacha, cebollas y Dactylis, que hace de este factor plasmático un instrumento de mejora genética importante en relación con el vigor híbrido o heterosis. Lo mismo ocurre cuando hay diferenciación en sexos distintos y factores citoplasmáticos influyen en la formación de órganos femeninos o masculinos ((sirven de ejemplos Aquilegia, Godetia, Bryonia, Satureja, Cirsium y Streptocarpus). Son de gran importancia a este respecto los estudios clásicos de Correns (1908, 1916). Las coníferas son generalmente monoicas, pero en muchas especies, como Pinus sylvestris, se encuentran individuos predominantemente masculinos o femeninos. Una selección sistemática hacia la dioecia puede ser de gran utilidad en la mejora por heterosis o vigor híbrido así como para el empleo de individuos con esterilidad masculina en los trabajos de cruzamiento. Por el contrario para producir líneas consanguíneas puede ser ventajoso en ciertos casos el paso de la monoecia o dioecia (por ejemplo, en Salix o Populus) al hermafroditismo. Tales transformaciones han sido efectuadas en el cáñamo (Sisov, 1937; Sengbusch, 1950), con paso de dioecia a monoecia o hermafroditismo.
En estos últimos años se ha visto que parece posible introducir en una especie herencia plasmática vía mutaciones inducidas en genes nucleares (Figura 3). Por ejemplo, en relación con algunos letales clorofílicos, las mutaciones nucleares inducen cambios irreversibles en los plastidios o plastogenes. Si entonces se retiran los genes nucleares, la alteración de plastidios persiste y es transmitida plasmáticamente a través de la planta madre a la descendencia (Wettstein, 1961 y Gustafsson 1960b, no publicado). Sería de gran interés poder inducir o aislar en especies forestales la esterilidad masculina o la dioecia condicionadas plasmática o génicamente.
En relación con la resistencia a enfermedades, algunos resultados obtenidos por Langner (1952) indican una base citoplasmática en la susceptibilidad a la micosis defoliadora de los cruces entre Larix decidua y leptolepis.
FIGURA 3. - Mutación de plastón génico-dependiente: cambios citoplasmáticos, en este caso aberraciones de plastidios, condicionados por mutaciones de genes.
Estructura de especies y poblaciones
En cierto modo la especie es la unidad fundamental de la evolución. Sin embargo, admite definiciones diferentes, según los puntos de vista del observador y experimentador. Turesson intentó hacer en 1922 una división tajante, fundándose en caracteres biológicos en vez de morfológicos dentro de y entre especies o grupos de especies. Distinguía entre:
1. ecotipos, que eran subunidades interfértiles;2. ecoespecies, capaces de formar híbridos, si bien con una fertilidad híbrida disminuida y una variabilidad de la descendencia híbrida reducida;
3. coenospecies, constituidas por una o más ecoespecies que pueden intercambiar genes entre sí pero que no admiten recombinación con otros grupos de especies.
Las concepciones de Turesson fueron más tarde propugnadas por Clausen, Keck e Hiesey en sus magníficos estudios sobre la flora californiana (véase Clausen, 1951). Sin embargo, no fueron de aceptación general. Para los fines de este informe es suficiente considerar que la especie es en general heterogénea y polimorfa y que está dividida en poblaciones adaptadas o en vías de adaptación a varios climas, sitios y nichos. Si luego las: poblaciones dentro de una especie, son o no diferentes entre sí morfológicamente, tiene una importancia secundaria en esta discusión, lo mismo que si las poblaciones forman ecotipos delimitados con mayor o menor contraste o constituyen clines con cambio gradual en los caracteres fisiológicos y morfológicos (Huxley, 1938; Langlet, 1959a, b).
El grado de heterogeneidad y la intensidad de la recombinación génica depende en gran medida del tipo de fecundación, esto es, que sea autofecundación o fecundación cruzada, o también de la desaparición de la fecundación en las especies propagadas partenogenética o vegetativamente. En cierto número de especies arbóreas, incluyendo coníferas, se halla bien establecida la autofecundación natural. Tal es el caso, según Langner (1959), de la especie uniforme, Picea omorika Sin embargo, aun entonces se da el cruzamiento, a menudo seguido por una segregación pronunciada en generaciones posteriores. En poblaciones pequeñas o aisladas puede tener lugar la autofecundación, con la consiguiente homocigotización y disminución general de la viabilidad en las variantes homocigóticas. En las especies arbóreas hermafroditas o monoicas el grado de autofecundación natural puede variar de un año para otro, en dependencia con la abundancia de floración y con el grado de dispersión del polen dentro de la masa (sobre distancias de transporte aéreo del polen véase Andersson, 1955). Debe considerarse detenidamente este hecho cuando se utilice silvícolamente la reproducción natural. Se ha señalado que en especies en que la fecundación cruzada es normal, como el Picea abies, Pinus sylvestris y Pinus monticola puede darse autofecundación con relativa facilidad, habiéndose encontrado algunas variantes plenamente autocompatibles (Sylvén, 1910; Plym Forshell, 1953; Barnes et al., 1962; Eklundh-Ehrenberg, 1963). Los biotipos autofecundos aparecen espontáneamente o en la experimentación bajo la acción de agentes mutagénicos por mutación antimorfa (loss-mutation) o destrucción de los genes y alelos de incompatibilidad.
Sin embargo, en numerosas especies de plantas, incluyendo especies arbóreas angiospermas y coníferas, existe una serie de transiciones entre la autoincompatibilidad total y la aptitud pronunciada para la autofecundación. De todas formas, por razones ecológicas, históricas y migracionales, las especies están subdivididas en poblaciones que se intercruzan naturalmente y están más o menos adaptadas a sus habitats especiales, y en su papel de constituyentes de comunidades naturales de plantas, con competición y cooperación de biotipos individuales entre y dentro de las especies.
Se hizo un análisis de lo más interesante en relación con los efectos de la autofecundación en contraposición a la fecundación cruzada en cuatro árboles de Pinus monticola (Bornes et al., 1962), dos de los cuales eran autofecundos y dos parcialmente autoestériles. Se hicieron mezclas de polen de árboles y se emplearon para la fecundación. La proporción de semilla de cruzamiento y autógama varió, dependiendo del genotipo del genitor femenino así como del genotipo del árbol productor del polen. En algunos casos, el polen propio fue tan efectivo como el polen extraño, aunque en la mayoría de los casos lo fue menos.
Los híbridos fuertemente heterocigotos de especies apomícticas pueden propagarse ampliamente en la naturaleza conservando sus propiedades, debido a su capacidad para desarrollar semilla asexualmente o a formas de multiplicación vegetativa. Tales especies escasean entre los árboles de valor económico, si bien se han encontrado casos de apomixis en Alnus, Euonymus, Sorbas y otros géneros, propagándose vegetativamente muchas especies como los chopos temblones, chopos y sauces.
Las poblaciones de árboles y su adaptación
Con frecuencia se considera que las poblaciones naturales están bien adaptadas al medio ambiente. Por ejemplo, en una publicación reciente de Duffield (1962, p. 9), se dice lo siguiente: «Como método de mejora genética, por tanto, la inducción de mutaciones es sencillamente como un juego con escasísimas probabilidades de acierto, ya que en organismos tan bien adaptados como la mayoría de los árboles forestales, virtualmente cualquier cambio es de esperar sea a peor» (la bastardilla es del presente escritor). En contradicción con esta afirmación, las poblaciones arbóreas a menudo no consiguen adquirir una adaptación completa a sus habitats. Esto puede aplicarse a la capacidad para dar flores del Picea abies que es irregular en gran parte de su área de distribución, o a la producción de semilla, que falta en absoluto o es obstaculizada en varias formas, como sucede en el Pinus sylvestris y Picea abies a grandes latitudes o altitudes (Simak y Gustafsson, 1954). Incluso en relación con la supervivencia y desarrollo de la fase vegetativa, la resistencia a la helada y al frío de poblaciones que viven en condiciones duras es muchas veces imperfecta. Wibeck (1933) se ha apercibido plenamente de esto para las poblaciones escandinavas de Pinus sylvestris, siendo estudiado luego con más detalle por Eiche (1962 y no publ.). Véase también Gustafsson (1962). No debe olvidarse que el medio ambiente natural cambia continuamente, y que las poblaciones locales tienen que pasar por sucesivos procesos de adaptación, los cuales conducen también a cambios genéticos de la variabilidad potencial. Además, las contingencias históricas como la heterocigosis original, el modo de migración desde condiciones menos severas a más, ataques irregulares o catástrofes causados por insectos, hongos, roedores, fuegos y así sucesivamente, pueden influir mucho sobre la constitución de una población local.
Haciendo los cálculos por unidad de tiempo y quizá también por generación, las especies forestales muestran una adaptación lenta a las condiciones cambiantes del medio. El proceso es retrasado todavía más por condiciones de floración pobres o fallo en la producción de semilla. (En algunos distritos de Suecia, por ejemplo, las buenas cosechas de semilla de Pinus sylvestris se dan una vez cada 30 ó 40 años.) Generalmente, en las poblaciones septentrionales de pinos y abetos la etapa juvenil, hasta los 15 ó 20 años, es más sensible a tipos de clima extremado. Pasada esta etapa, la masa, así como los biotipos individuales, pueden considerarse vegetativamente adaptados. Por esto, dependiendo de las condiciones climáticas que siguieron a la germinación en el campo o a la plantación, las poblaciones locales pueden estar más o menos severamente seleccionadas para una localidad especial, implicando consecuencias de gran duración para las generaciones futuras.
En general, las poblaciones arbóreas se hallan imperfectamente adaptadas a muchas formas de intervención humana, tales como los métodos de aclareo, y no están, como especies silvestres que son, seleccionadas en forma adecuada para servir a las exigencias humanas en calidad de madera, producción de pasta o composición química. Tal estado de cosas hace más urgente todavía su mejora genética, que podría dar lugar a una domesticación intensa.
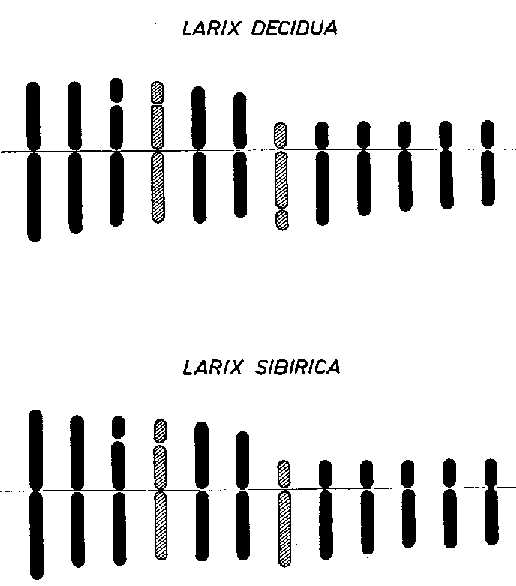
FIGURA 5. - Idiograrnas de especies de alerce: especies afines tienen a menudo cariotipos (idiogramas) diferentes como lo demuestran estos idiogramas de Larix decidua, y sibirica.
Composición genotípica de poblaciones de fecundación cruzada
Es de importancia fundamental en la mejora genética vegetal analizar la composición genotípica de la población: cómo se distribuyen los genes por los cromosomas sus efectos en heterocigosis y homocigosis, su recombinación libre o limitada. Los estudios sobre Drosophila por Mather (1943, 1960), Dobzhansky (1951) y sus colaboradores, han proporcionado un cuadro bastante bueno del comportamiento y constitución de las poblaciones tanto bajo condiciones naturales como experimentales, bajo presiones de selección intensas y suaves. Es un hecho bien comprobado la heterocigosis extrema de las poblaciones naturales, incluyendo la fragmentación y reagrupamiento de segmentos de cromosoma, así como la abundancia de mutaciones en estado heterocigótico, que cuando están en dosis homocigótica actúan como letales o semiletales y disminuyen la viabilidad. Muchas de tales mutaciones aumentan la viabilidad por encima de la «normal» cuando están en heterocigosis, siendo un ejemplo los letales clorofílicos (Gustafsson, 1954). Eiche (1955) ha mostrado que estos letales de clorofila son frecuentes también en poblaciones naturales de árboles forestales. En todo caso, existe un sistema complejo de equilibrio entre todo tipo de genes y mutaciones, tanto en heterocigosis como homocigosis. Debido a los fenómenos de ligamiento, se restringe la variabilidad potencial en las primeras fases de la selección, pero de nuevo es liberada una vez hayan tenido lugar el sobrecruzamiento y la recombinación de genes en segmentos próximos de cromosomas.
Desgraciadamente, no se conoce gran cosa sobre la ocurrencia de translocaciones o inversiones espontáneas en especies arbóreas. Los descubrimientos de Sugihara (1940) indican la existencia de translocaciones en poblaciones locales de Cephalotaxus drupacea, ya que se asegura la existencia de cuadrivalentes y hexavalentes en la meiosis de biotipos de esta especie. Pueden ocurrir pequeñas inversiones en todas las especies aunque no hayan sido observadas en los pocos estudios realizados hasta ahora sobre meiosis. El análisis cuidadoso de los idiogramas de especies emparentadas, por ejemplo, en Larix (Figura 5) o Pinus entre las coníferas, podría revelar si la diferenciación de especies se debe no sólo a mutación de genes, sino también a reordenamientos pequeños y grandes de los cromosomas. Del trabajo hecho de hibridación se desprende que pueden conseguirse resultados prometedores por el cruce de numerosas especies y géneros de coníferas, provocando la aparición del vigor híbrido (Righter, 1946, 1960; Hyun, 1960). En la naturaleza, la hibridación entre especies tiene gran conexión con la introgresión entre ellas (Andersson, 1949; Stebbins, 1950); esto es, los genes se transfieren de una especie a otra por hibridación y posterior retrocruzamiento del híbrido con una u otra de las especies parentales. Tal introgresión ha sido comunicada también para especies de Alnus, Quercus y Pinus.
El desarrollo armonioso de muchas poblaciones arbóreas está íntimamente asociado a fenómenos de simbiosis, por ejemplo la presencia de micorrizas. Esto conduce a una interacción mutua y a la continua adaptación de grupos de organismos enteramente diferentes. Además, numerosos investigadores están de acuerdo en que una masa de árboles forma a menudo o regularmente un enorme «convivium» resultante de una simbiosis de raíces de gran alcance, con transporte de nutrientes, hormonas, exudados, agua y otros materiales desde unos individuos a otros. Las consecuencias biológicas de tales influencias mutuas bióticas no han sido estudiadas por ahora desde un punto de vista genético o biométrico.
La herencia cuantitativa y su base genética
En los párrafos precedentes se hizo hincapié en la constitución compleja de las especies vegetales de fecundación cruzada. Las poblaciones reaccionan a la selección natural y artificial a la manera de sistemas poligénicos en los que numerosos genes, la mayoría con efectos individuales pequeños, cuando menos en estado heterocigótico, constituyen la base genética de los caracteres cuantitativos. La teoría cuantitativa de la herencia ha sido analizada por Mather en numerosos artículos (Mather, 1960). Ya en 1915 Nilsson-Ehle señaló la existencia de tales sistemas hereditarios complejos y escribió: «Es evidente por los análisis bastante completos hechos en diferentes países que los caracteres de importancia práctica, siempre que sean cuantitativos, tienen que ser concebidos en general como resultado de la acción de varios, y en ocasiones numerosos, factores genéticos, que obedecen las leyes de Mendel. Todas las propiedades hereditarias, no sólo los caracteres morfológicos externos sin trascendencia en un programa de mejora, sino también los caracteres fisiológicos y biológicos, tales resistencia a las inclemencias del invierno, resistencia genética a las enfermedades, precocidad, resistencia a los transplantes, facultad germinativa y demás, segregan claramente después de los cruzamientos y forman nuevas combinaciones. En general, las propiedades cuantitativas siguen este comportamiento sirviendo de ejemplo las características de tamaño y el contenido de proteína del trigo. El autor no halló en las investigaciones caracteres de importancia para la práctica que se comportasen de otra forma. La segregación puede ser más o menos compleja, lo que depende, de acuerdo con la teoría presentada aquí, del número y de la acción de los diferentes factores genéticos, pero no hay duda de que las diferentes propiedades se conducen en líneas generales de forma semejante. La composición de los caracteres de importancia práctica, dependiendo de un gran número de factores genéticos, tiene la máxima transcendencia para los principios y métodos de la mejora genética vegetal» (op. cit. página 57, traducido).
Sin embargo, cuando Nilsson-Ehle, East, Fisher, Mather y otros resaltan la importancia de los genes de efecto pequeño, no debe olvidarse que las plantas cultivadas también se separaron de sus antepasados silvestres por varios caracteres modificados drástica y bruscamente (Schwanitz, 1957). Las mutaciones de efecto grande han contribuido continuamente al proceso de domesticación. Alrededor de estos cambios mayores tienen lugar luego la recombinación y nuevas mutaciones que unen y ajustan los cambios grandes y pequeños en un todo equilibrado, en el que numerosos genes influyen en el mismo carácter cuantitativo. En las especies de fecundación cruzada también los genes y mutaciones deletéreas ejercen tales influencias modificadoras ligeras en un estado heterocigótico. La razón de que sean tan difíciles de analizar en la herencia cuantitativa de las especies forestales los cambios provocados por un solo gen, es sencilla y suficiente: que el diseño de los experimentos y los métodos de medición son muchas veces poco precisos e inadecuados, y las influencias ambientales encubren los efectos genéticos pequeños.
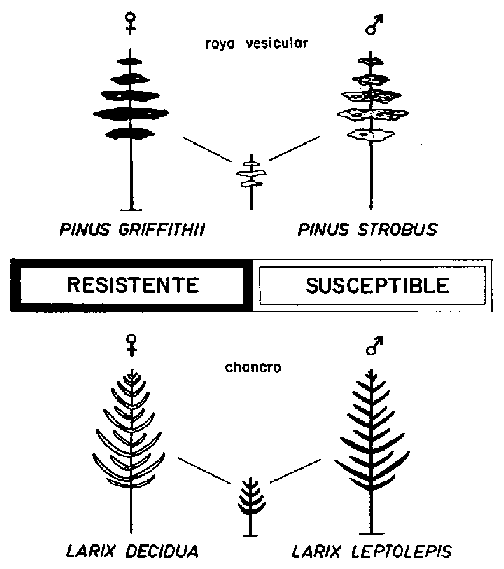
FIGURA 6. - Híbridos entre especies y resistencia a la enfermedad. En este caso de coníferas puede ser «recesiva» (figura superior) o «dominante» (figura inferior).
Resistencia a enfermedades y efectos génicos mayores
La genética de la resistencia a enfermedades en especies forestales ha ganado en importancia en años recientes. En un análisis publicado en 1962, Heimburger formulaba el problema de la siguiente forma: «La base genética de la resistencia en el hospedante puede ser poligénica o estar gobernada por un número pequeño de genes mayores; sin embargo, en la mayoría de los casos la resistencia de las plantas a la enfermedad se vio que dependía de una combinación de poligenes y genes mayores. Los genes mayores que gobiernan la resistencia a enfermedades constituyen la base de la mayoría de los avances espectaculares en la lucha genética contra las enfermedades en la agricultura y horticultura». Por otro lado, Heimburger dice a continuación: «Sin embargo, no debe olvidarse que la mayor parte de las plantas cultivadas fueron producidas por selección y mejora a base principalmente de poligenes y que muchas características nuevas y útiles, tales como resistencia a la enfermedad, pueden desarrollarse substancialmente de esta forma si no se dispone de otros materiales estrechamente emparentados con resistencia superior» (op. cit., p. 358). Los datos sobre resistencia de especies forestales a las enfermedades son vagos y contradictorios. Se debe esto en gran medida al hecho de haber sido estudiada la herencia de la resistencia principalmente en cruzamientos de especies, en los que la condición de híbridos implica la acción fisiológica de los genes, debiéndose la resistencia a causas muy diferentes. Quizá sea de esperar que los híbridos de especies entre biotipos resistentes resulten a menudo susceptibles. El mismo Heimburger cita híbridos entre Pinus griffithii y Pinus strobus en los que el «mecanismo inhibidor es alterado, la infección se extiende rápidamente desde las acicalas al tallo, siendo el resultado una fuerte mortalidad de las plantitas (op. cit, p. 360). Por otro lado, se sabe de casos en los que una especie hace que el híbrido sea casi completamente resistente, por ejemplo, en los cruces del Larix leptolepis, resistente al chancro, con el Larix europea (Figura 6).
Se dispone de numerosos datos sobre resistencia dependiente de genes mayores en plantas agrícolas y hortícolas. Knight (1946) presenta una lista de 33 plantas agrícolas en las cuales ha sido demostrada la existencia de genes mayores de resistencia a 84 plagas y enfermedades. Trabajos recientes de Briggs, Flor, Favret y otros han puesto de manifiesto la existencia de numerosos genes de resistencia dentro de una especie y que muchas veces no están distribuidos al azar por todo el genomio de la planta hospedante, sino que tienden a agruparse en segmentos genéticos, concentrados en unos pocos cromosomas. El primero en indicar este fenómeno fue Briggs en relación con la enfermedad de la caries del trigo. La genética de la resistencia de la cebada al mildiú constituye probablemente el ejemplo más destacado (Favret, 1960a, b). Dieciocho factores diferentes para la resistencia al mildiú se distribuyen en 14 loci; 17 están en el cromosoma 5 y uno en el cromosoma 4. Trece loci forman un largo «segmento isofénico» de 45 a 50 unidades de sobrecruzamiento. Este segmento isofénico puede ser dividido en cuatro secciones de acuerdo con la naturaleza de los alelos para la resistencia. Algunos loci contienen solamente un alelo para la resistencia, otros están constituidos por series de 2 a 5 alelos. Tres loci están ligados estrechamente formando un complejo génico en un segmento corto de una longitud aproximada de una unidad de sobrecruzamiento. La mayoría de los genes para la resistencia son dominantes o semidominantes, aunque también puede haber factores recesivos.
Es muy significativo el que este sistema complejo de resistencia haya sido encontrado en varias especies estudiadas con todo cuidado. Esto abona la hipótesis de una condición semejante también en las coníferas y frondosas arbóreas. Aunque actúen muchos genes, cada uno ejerce en general un efecto «mayor». Por esto, siguiendo a Mather, la herencia no es «poligénica» en el sentido de Heimburger, es decir, con efectos aditivos pequeños, sino que es «multifactorial» en el sentido de Nilsson-Ehle, basada quizá en una serie de pequeñas duplicaciones, como sugiere Favret en su trabajo sobre mutación, siendo muchos factores para la resistencia dominantes o semidominantes. Además, se debe hacer hincapié en que la resistencia a enfermedades y la tolerancia en el campo para una enfermedad pueden entrañar fenómenos diferentes.
FIGURA 7. - La resistencia a los hongos, en este caso al mildeu de la cebada, depende con frecuencia de muchos genes, dominantes o recesivos, que van en segmentos específicos de los cromosomas, llamados segmentos isofénicos.
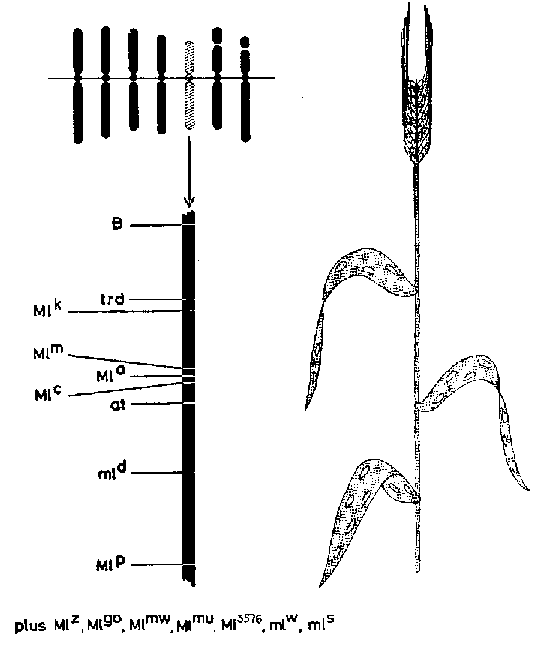
Concepto de heredabilidad
Al estudiar los caracteres cuantitativos de los árboles, tales como producción en volumen, crecimiento en altura o diámetro, forma del tronco y propiedades de la madera, las experiencias deberán establecerse de forma que puedan separarse las influencias genéticas de las del ambiente y estimar hasta qué punto un cierto fenotipo está condicionado por la herencia y hasta qué punto por el medio. En la mejora animal, debido al mucho tiempo entre generaciones y a los grandes costos del ensayo de descendencias, estos problemas son tan importantes como en la mejora genética de los árboles. El concepto de heredabilidad (h²) (véase Lush, 1948) fue ideado para representar aquella parte de la variancia fenotípica total (Vp) causada por factores genéticos con efectos aditivos (Va). Originariamente el término heredabilidad se refería sólo a la correlación entre genitores y descendencias, pero en la mejora genética arbórea la heredabilidad se emplea a menudo con dos significados, de acuerdo con Lush. La heredabilidad en sentido lato se refiere al funcionamiento de todo el genotipo y se emplea como contraste con la variancia ambiental. La definición de heredabilidad en sentido estricto incluye solamente los efectos promedios de los genes, transmitidos de los genitores a la descendencia a través de la meiosis (segregación cromosómica) y subsiguiente fecundación (recombinación cromosómica). «Esta acepción restringida de la heredabilidad se emplea cuando se tiene especial interés en expresar qué fracción de las diferencias fenotípicas entre genitores puede esperarse razonablemente sea recuperada en la descendencia (Lush, op. cit.). Véase también Toda, 1957; Zobel, 1961; y Eklundh Ehrenberg, 1963.
La heredabilidad en sentido lato es una confirmación del contraste de Johannsen entre genotipo y fenotipo. El concepto aparece muy claro en los experimentos sobre el color de la flor y la temperatura en Primula sinensis, descritos por Erwin Baur (1919) en su clásico libro de texto sobre genética (Figura 8). En las especies forestales, cuando la propagación vegetativa es fácil, el análisis clonal revela fácilmente el componente genotípico de la variación. Esto constituye un principio importante en la selección de árboles plus, pues numerosos factores, tales como las propiedades del suelo, humedad, densidad de la masa, aclareos irregulares y demás, pueden provocar un desarrollo plus en árboles inferiores genéticamente o un desarrollo minas en otros superiores genéticamente. Sin embargo, la forma del tronco, tipo de ramificación y especialmente ángulo de las ramas, mantienen frecuentemente una estrecha dependencia genética. En las características de la semilla de Pinus sylvestris se manifiesta de forma elegante la influencia relativa de la herencia y del ambiente (Simak y Gustafsson, 1954). Los caracteres que como el color y el tamaño de la semilla para su expresión dependen de la madurez son fácilmente influenciados por cambios ambientales. Por el contrario, dependen menos del ambiente los caracteres relativos a la forma de las semillas, es decir, cociente entre longitud y anchura, estructura superficial y forma del ala. Los detalles morfológicos de la semilla que son específicos de los biotipos individuales, como punta micropilar cónica o curveada, o hilum asimétrico muy marcado, son muy fijos e independientes de los cambios de clima o suelo. Con la ayuda de las características de naturaleza muy fija puede mantenerse una exacta identificación del material de pino manejado en el trabajo de injertado y en los huertos para semilla.
Las propiedades de la madera han suscitado en los últimos años gran interés (Zobel, 1961; Ericson, 1960, 1961; Wellwood y Smith, 1962). En algunas especies, la correlación del peso específico entre árboles plus seleccionados y sus clones en pruebas clonales es bastante pronunciada. Cuanto mayor es la densidad básica en un clon de Pinus sylvestris tanto mayor es su producción de pasta y mayor el índice de rasgado de la misma. Según Wellwood y Smith (op. cit.) para la Pseudotsuga taxifolia y Tsuga heterophylla, no existe relación entre las características externas de los árboles plus y las propiedades internas de su madera. En la mejora genética dirigida a incrementar la producción de pasta de estas especies deberá analizarse por separado las propiedades de la
FIGURA 8. - Respuesta del genotipo a diferentes ambientes. A. Muestra la respuesta fenotípica de un genotipo de Primula a ligeros cambios de temperatura. B. Indica cómo la forma de la semilla de un genotipo de Pinus es muy independiente de las condiciones climáticas, mientras el color de la semilla cambia mucho con el ambiente. C. Muestra que el ángulo de las rama, de Pinus sylvestris tiene una alta «heredabilidad» tanto en sentido lato como estricto.
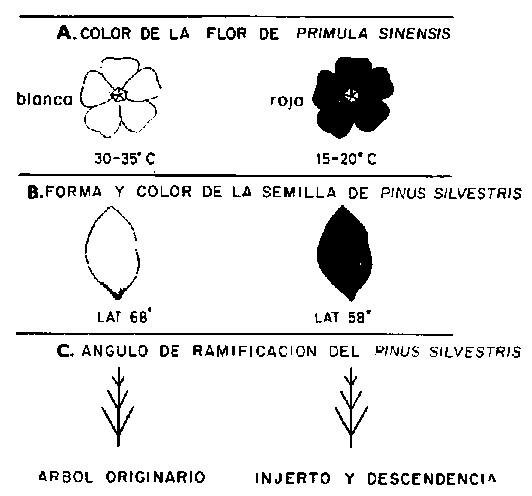
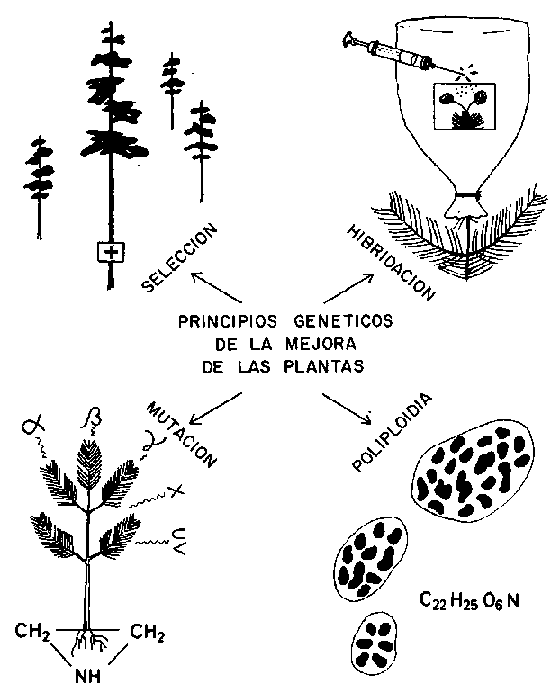
FIGURA 9. - Principios genéticos de la mejora de las plantas.
La heredabilidad en sentido estricto es con ciertas limitaciones un concepto útil. Cuando la experimentación es la apropiada, el análisis matemático indicará el valor del componente genético, por lo menos en descendencias de cruces intrapoblacionales. El problema importante es extraer las influencias ambientales no específicas. Si se seleccionan cuidadosamente los árboles plus empleados en la mejora genética, los biotipos con propiedades hereditarias minas serán excluidos automáticamente no participando en la producción de semilla. Esta forma de proceder, aunque negativa, es importante. Sin embargo, se reforzará el aspecto positivo de la selección si los árboles plus empleados son elegidos con todo cuidado, probados en ensayos de clones y cruzados con compañeros apropiados. Anima el ver que los ensayos de procedencias con semilla mezclada han revelado una relación clara entre la calidad fenotípica de la masa parental y la de su descendencia. Petrini (1959) describió un fenómeno de este tipo en un ensayo de procedencias de Pinus sylvestris de 50 años de edad. De sus estudios comparando la herencia de árboles plus con la de árboles minas, Eklundh Ehrenberg (1963) ha puesto de manifiesto que la descendencia de árboles plus seleccionados daba mejor desarrollo y producción que las descendencias de árboles minas.
Sin embargo, la mejora genética de las plantas no persigue sólo los efectos generales de la combinación, sino también los específicos. El establecimiento de huertos para semillas conteniendo 30, 40, o más clones derivados de árboles plus de una determinada región es una precaución que puede ser recomendada para zonas forestales de condiciones estacionales y climáticas rigurosas y variables. Por otro lado, en zonas más productivas con clima favorable se puede disminuir el número de clones y, al tiempo que se va adquiriendo más experiencia, atender más a la aptitud combinatoria específica. En los huertos de semillas formados para aprovechar el vigor híbrido por cruzamiento de especies y procedencias, el número de clones diferentes puede reducirse aún más tras cuidadosos ensayos de descendencia. Tiene esto aplicación cuando no se practica la repoblación natural y las generaciones futuras se establecerán por plantación. Los cálculos matemáticos sobre la relación de genitores a descendencia (heredabilidad en sentido estricto) presentarán con frecuencia fallos si no son tenidas en cuenta los efectos combinatorios específicos.
Principios genéticos de la mejora de plantas
A continuación se resumen los factores genéticos que intervienen en la mejora de las plantas, todos los cuales actúan también en la naturaleza. Son:
1. selección;2. hibridación;
a) heterosis o vigor híbrido de F1 en los cruces entre especies, poblaciones e individuos;b) recombinación y transgresión en F2 a Fn;
c) retrocruzamiento de individuos F1 a Fn con uno u otro progenitor;
3. mutaciones espontáneas o inducidas debidas a inestabilidades moleculares, radiaciones ionizantes, substancias químicas mutagénicas;
4. poliploidia natural o artificial resultante en alo- y autopoliploidia.
Hay numerosos artificios que pueden facilitar el trabajo de mejora genética a largo plazo. Tales son, por ejemplo, la propagación vegetativa, el empleo de la esterilidad masculina, producción de líneas consanguíneas u homocigóticas y su propagación masiva subsiguiente por injerto. El análisis con rayos X descubre el desarrollo del embrión y endosperma. La infección masiva, reveladora de la resistencia genética a las enfermedades, puede ejecutarse en invernaderos. Instalaciones climáticas de varias clases pueden facilitar el estudio de las relaciones genotipo-fenotipo y también las condiciones fisiológicas y ecológicas de la floración precoz y abundante. Las correlaciones de caracteres entre genitor y descendencia, en fases tempranas, simplifican la selección acertada de los árboles genitores en los trabajos de cruzamiento. Hay que hacer hincapié en la necesidad del análisis matemático. Otros muchos recursos de carácter científico y técnico, muchos aún no disponibles, permitirán en el próximo futuro acelerar la investigación y la puesta en práctica de los resultados.
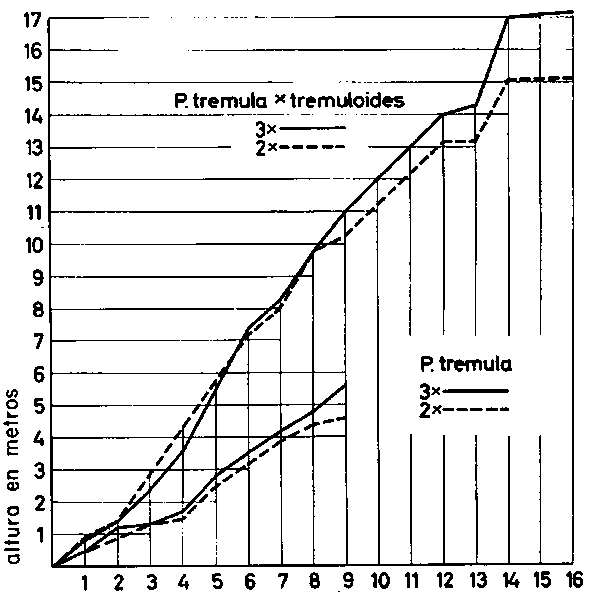
FIGURA 10. - Crecimiento en altura de híbridos de especies y triploides de Populus, ilustrando la combinación de dos métodos de mejora.
Uno de los aspectos más fascinadores de la biología moderna es la validez general de los principios genéticos aplicables a numerosas especies y géneros. Ello permite traspasar los resultados obtenidos en las bacterias, algas o moscas o poblaciones de árboles y al hombre. La aplicación de los descubrimientos puede estar limitada por el tamaño del organismo o la duración de su ciclo reproductivo. Existe un fatalismo en el punto de vista que el genetista tiene de la vida: el material hereditario constituido por unidades fijas con un grado de constancia muy elevado, se transmite en general sin cambios de generación en generación, si bien con la posibilidad de mutación. Sin embargo, el material hereditario no está aislado en el vacío; a través de mutaciones y recombinaciones los genotipos han sido seleccionados y modelados durante miles o millones de años en ambientes variantes. En otras palabras, es inconcebible un genotipo sin su medio ambiente.
Esta interacción genotipo-ambiente se ve en la práctica de la silvicultura. El valor económico de una masa puede incrementarse por medio de aclareos, podas y demás tratamientos, pero ningún forestal podrá convertir una masa fracasada por su origen inadecuado en una masa sobresaliente. Es imposible aplicar la mejora genética a las especies forestales sin prestar la debida consideración a las otras muchas disciplinas de la técnica forestal. A este respecto puede existir una cierta distinción en la mejora genética vegetal según se aplique a los montes o a la agricultura.
Las plantas agrícolas y hortícolas han experimentado diferentes cambios durante su desarrollo desde el estado original silvestre. Las diferencias implican cambios morfológicos, cambios fisiológicos dirigidos a aumentar la producción, y variaciones en el cariotipo que sirve de base. Los cambios cariotípicos afectan al número y estructura de los cromosomas y al ordenamiento de los genes en los cromosomas. Un ejemplo típico lo proporciona la comparación de las especies diploides silvestres de Triticum y Aegilops con la especie comercial hexaploide Triticum aestivum. Aun dentro del grupo de plantas cultivadas diploides, la modernización ha sido notable. Compárese, por ejemplo, el híbrido del maíz de la zona maicera de los Estados Unidos con las muestras de maíz primitivo descubiertas en las cuevas de Nuevo México y Sudamérica (Mangelsdorf, 1958). Hay también especies tales como el altramuz, que empezaron a cultivarse recientemente. Gracias a los métodos hábiles de Sengbusch se desarrollaron cultivares libres de alcaloides al final de la década 1920-30 y después han sido mejoradas continuamente con la ayuda de nuevas mutaciones y recombinaciones.
Ha comenzado un proceso similar de domesticación de algunas especies forestales con el descubrimiento por Nilsson-Ehle del chopo temblón triploide de crecimiento rápido (Nilsson-Ehle, 1936). Cruzando tetraploides con diploides es posible ahora producir grandes cantidades de triploides. Nuevos adelantos se deben a Johnsson (1953 y sin publicar) al demostrar (véase Gustafsson 1960b) que los híbridos entre Populus tremula y Populus tremuloides producen más en Suecia que el Populus tremula nativo (Figura 10). Johnsson consiguió aumentar ano más la producción introduciendo Populus tremula tetraploide en el programa de cruzamiento y al mismo tiempo logró transmitir la resistencia (o cuando menos la tolerancia) a Valsa nívea del tretraploide al híbrido. Por otro lado, numerosos cultivares de Populus son clones de alta calidad a nivel diploide, resultado también de varios cruces entre especies. Sin embargo, no puede evitarse la impresión de que la obra de hibridación hecha hasta ahora con el chopo ha tenido muchas veces un carácter casual y que en el futuro la planificación cuidadosa de los cruzamientos dará mejores resultados.
En las coníferas, igual que en las frondosas arbóreas y en las plantas agrícolas, la domesticación producirá cambios transcendentales en la estructura de la población. Son fáciles de producir híbridos en algunos géneros de árboles, especialmente en Pinus y Larix. Muchos combinan el vigor híbrido con la resistencia o tolerancia a las enfermedades. Nada se sabe sobre híbridos triploides de Pinus, pero posiblemente algunos habrá que tengan aplicación en el futuro. La labor de cruzamiento a nivel intraespecífico tendrá seguramente mayor importancia práctica inmediata. Por ejemplo, las procedencias de Picea abies oriundas de Europa central, Polonia o Rusia occidental producen a menudo en Escandinavia más que las poblaciones nativas. Se han importado grandes cantidades de semillas y plantas de aquellas procedencias, y en los huertos para semilla establecidos en Suecia figuran ya árboles plus seleccionados, de origen nativo o foráneo (Andersson y Andersson, 1962). Han sido hechos cruzamientos heterogéneos variados en experiencias con huertos de semillas clonales, lo que cambiará fundamentalmente la estructura de la población al generalizarse su empleo para semilla F1. Las masas, si bien heterogéneas, no cabe duda serán de alta productividad. En tales masas puede procederse otra vez a la selección de nuevos árboles plus. El proceso de domesticación en los árboles forestales será naturalmente más lento que en las plantas cultivadas anuales o bienales, variando según las especies y caracteres sometidos a selección. El producto final dependerá de las necesidades de utilización y de las condiciones ecológicas del monte. Las poblaciones expedidas para uso práctico por los mejoradores de plantas deben estar adaptadas plenamente a los climas y sitios donde se proyectan emplear. Supone esto la conformidad completa de la suma de genotipos de la población con el medio ambiente, principio fundamental que abre y cierra este capítulo.
ANDERSON, E. 1949. Introgressive hybridization. New York. 109 p.
ANDERSSON, E. 1955. Pollenspridning och avståndsisolering av skogsfröplantager [Pollen distribution and isolation by distance of seed orchards]. Norrlands Skogsv. Förb. Tidskr., p. 35-100.
ANDERSSON, E. 1955. Pollenverbreitung und Abstandsisolierung von Forstsamenplantage. Z. Forstgenet., 4: 150-152.
ANDERSSON, E. s; ANDERSSON, S. 1962. Förteckning över Domänverkets och Skogevårdsstyrelsernas skogsfröplantager [List of seed orchards owned by the Swedish Forest Service and the Local Forest Conservation Boards]. Stockholm, Samarbetsnämnden för skoglig växtförädling och genetik. Limited distribution. 158 p.
BABCOC, E. B. 1947. The genus Crepis. Part One. The taxonomy, phylogeny, distribution and evolution of Crepis. Berkeley, University of California Press. 197 p.
BARLOW, B. A. 1959. Chromosome numbers in the Casuarinaceae. Aust. J. Bot., 7 (3): 230-237.
BARNER, H. & CHRISTIANSEN, E. 1962. The formation of pollen, the pollination mechanism, and the determination of the most favourable time for controlled pollination in Pesudotsuga menziesii. Silvae Genet., 11: 89-102.
BARNES, B. V., BINGHAM, R. T., & SQUILLACE, A. E. 1962. Selective fertilization in Pinus monticola Dougl. II. Results of additional tests. Silvae Genet., 11: 103-111.
BAUR, E. 1919 .Einführung in die experimentelle Vererbungslehre. Berlin. 410 p.
BRANDENBURG, M. K., MILLS, E. L., RICKARD, W. K. & SHIELDS, L. M. 1962 .Effects of acute gamma radiation on growth and morphology in Pinus monophylla Torr. and Frem. (Pinyon pine). Radiation Botany, 2; 251-264.
CHIARUGI, A. 1962 .La cariologia nelle sue applicazioni a problemi di botanica. Atti della Società Italiana Progresso delle Science, XXI Riunione Roma, 1932, III: 1-38.
CHIBA, S. & WATANABE, M. 1952. Tetraploids of Larix kaempferi appeared in the nurseries. J. Jap. For. Soc., 34: 276-279.
CHRISTIANSEN, H. 1950. A tetraploid Larix decidua Miller. Det Kgl. Danske Videnskap. Selsk., Biol. Meddel., 18 (9). 9 p.
CLAUSEN, J. 1951. Stages in the evolution of plant species. New York. 206 p.
CORRENS, C. 1908. Die Rolle der männlichen Keimzellen bei der Geschlechtsbestimmung der gynodioecischen Pflanzen. Ber. disch. bot. Ges., 26: 686-701.
CORRENS, C. 1916. Untersuchungen über Geschlechtsbestimmung bei Distelarten. Sitz. Ber. Kön. Preuss. Akad. Wiss., 20: 448-477.
DOBZHANSKI, TH. 1951. Genetics and the origin of species. 3rd ed. New York, Columbia University Press. 364 p.
DUFFIELD, J. W. 1962. Forest tree improvement: old techniques and the new science of genetics. H. R. MacMillan Lectureship, University of British Columbia, Columbia, 12. 16 p.
EICHE, V. 1955. Spontaneous chlorophyll mutations in Scots pine (Pinus silvestris L.). Medd. Statens skogsforshn. Inst. Stockh., 45: 1-69.
EICHE, V. 1962. Nya aspekter på plantavgång och bristande vinterhärdighet i norrländska tallkulturer [New aspects on plant mortality and deficient winter hardiness in North-Swedish pine plantations]. Skogen: 423-426, 432.
EKLUNDH EHRENBERG, C. 1963. Genetic variation in progeny tests of Scots pine (Pinus silvestris L.). Studia Forestalia Suecica, 10: 1.-135.
ERICSON, B. 1960. Studies of the genetical wood density variation in Scots pine and Norway spruce. Statens skogsforshn. Inst. Stockh., Avdel. skogsproduktion, Rappt. 4, 1-52.
ERICSON, B. 1961. Skogsträdsförädling med sikte på ökat massautbyte. Tek. Vetenskaplig Forskning, 32: 194-203. (Statens skogsforskn. Inst. Ser. Uppsatser 81).
FAVRET, E. A. 1960a. Spontaneous and induced mutations of barley for the reaction to mildew. Hereditas, 46: 20-28.
FAVRET, E. A. 1960b. Induced mutations for resistance to diseases. Genet. Agr., 13: 1-26.
GUSTAFSSON, Å. 1951. Some aspects on variation and evolution in plants. Evolution, 5: 181-184.
GUSTAFSSON, Å. 1954. Mutations, viability and population structure. Acta Agr. Scand., 4: 601-632.
GUSTAFSSON, Å. 1960a. Polyploidy and mutagenesis in forest tree breeding. Proc. 5th World For. Congr., Seattle, U.S.A., p. 793805.
GUSTAFSSON, Å. 1960b. Chemical mutagenesis in higher plants. Chemische Mutagenese Erwin-Baur-Gedächtnisvorlesungen I (1959). Abhandl. Deut. Akad. Wiss. Berlin, Kl. Med 1, 14-29.
GUSTAFSSON, Å. 1962. Genetick och växtförädling i skogsbrokets tjänst [Genetics and plant breeding as applied to Swedish forestry]. Svenska Skogsv. Fören Tidskr., 111-150.
GUSTAFSSON, Å. & SIMAK, M. 1958. Effect of X and y-rays on conifer seed. Medd. Statens. skogsforskn. Inst. Stockh., 48 (5): 120.
GUSTAFSSON, Å. s; WETTSTEIN, D. von. 1957-58. Mutationen und Mutationszüchtung. Handb. Pflanzenzücht., 1: 612-699.
HEIMBURGER, C. 1962. Breeding for disease resistance in forest trees. For. Chron., 38: 356-362.
HUXLEY, J. 1938. Clines: an auxiliary taxonomic principle. Nature, Lond., 142: 219-220.
HYUN, S. K. 1960. Mass production of control-pollinated seed of conifers. Proc. 5th World For. Congr., Seattle, U.S.A., p. 787-792.
ILLIES, Z. M. 1952. Colchicineversuche on Larix decidua Miller und Picea abies (L). Karst. Z. Forstgenet., 1: 36-39.
ILLIES, Z. M. 1956. Veränderungen der Pollengrösse bei Lärche nach Blütenbehandlung mit Colchicin. E. Forstgenet., 5: 112-115
ILLIES, Z. M. 1959. Polysomatie im Meristem von Einzelbaumabsaaten bei Picea abies. Silvae Genet., 7: 94-97.
JOHNSSON, H. 1953. Hybridaspens ungdomsutveckling och ett fõrsök till framtidsprognos [The early development of hybrid aspen and an attempt to predict its future production]. Svenska Skogsv. Fören. Tidskr., p. 73-96.
JOHNSSON, H. 1959. Föreningen för Växtförädling av Skogsträd 1936-1958 [The Association for Forest Tree Breeding 1936-1958]. Norrtelje Tidn. Boktr. AB. 68 p.
KANEZAWA, R. 1949. Polyploids appeared in twin seedlings of forest trees. J. Jap. For. Soc., 31: 22-24.
KIELLANDER, C. L. 1950. Polyploidy in Picea abies. Hereditas, 36: 613-516.
KNIGHT, R. L. 1946. The role of major genes in the evolution of economic characters. J. Genet., 48: 370-387.
LANGLET, O. 1959a. Norrlandstallens praktiska och systematiska avgränsning. Svenska Skogsv. Fören. Tidskr., 1959: 425-436.
LANGLET, O. 1959b. A cline or not a cline-a question of Scots pine. Silvae Genet., 8: 13-22.
LANGNER, W. 1952. Kreuzungaversuche mit Larix europaea D.C. und Larix leptolepis Gord. Z. Forstgenet., 1: 40-56.
LANGNER, W. 1959. Selbstfertilität und Insucht bei Picea omorika (Pancic) Purkyne. Silvae Genet., 8: 84-93.
LARSEN, C. S. s; WESTERGAARD, M. 1938. Contributions to the cytogenetics of forest trees. I. A triploid hybrid between Larix decidua Miller and Larix occidentalis. Nutt. J. Genet., 36: 523-530.
LUSH, J. L. 1948. Heritability of quantitative characters in farm animals. Proc. 8th Intern. Congr. Genet., p. 356-375.
MANGELSDORF, P. C. 1958. Reconstructing the ancestor of corn. Proc. Am. Phil. Soc., 102: 454-463.
MATHER, K. 1943. Polygenic inheritance and natural selection. Biol. Rev., 18: 32-64.
MATHER, K. 1960. Evolution in polygenic system. Accad. Naz. dei Lincei. Evoluzione e Genetica 1960 Colloquio Internazionale, Roma, 1959), p. 131-152.
MEHRA, P. N. 1960. Cytology and breeding of conifers. Proc. 5th World For. Congr., Seattle, U.S.A., II: 728-733.
MERGEN, F. 1958. Natural polyploidy in slash pine. For. Sci., 4: 283-295.
MERGEN, F. 1959. Colchicine-induced polyploidy in pines. J. For., 57: 180-190.
MERGEN, F. 1961. The chromosomes of Pseudolarix amabilis. Cytologia, 23: 213-216.
MERGEN, F. 1963. Evaluation of spontaneous, chemical, and radiation-induced mutations in the Pinaceae. FAO/FORGEN 631/1.
MERGEN, F. s; STAIRS, G. R. 1962. Low level chronic gamma irradiation of Pitch-Pine-Oak forest -its physiological and genetical effects on sexual reproduction. Radiation Botany, 2: 206-216.
MERICLE, L. W., MERICLE, R. P. & SPARROW, A. H. 1962. Cumulative radiation-damage in oak trees. Radiation Botany, 2: 265-272.
MICHAELIS, P. 1957-58. Plasma-Vererbung. Hand. Pflanzensücht., I: 140-175.
MÜNTZING, A. 1936. The evolutionary significance of auto polyploidy. Hereditas, 21: 263-378.
MÜNTZING, A. 1956. Chromosomes in relation to species differentiation and plant breeding. Wageningen, Conference on Chromosomes, p. 1 -37.
MÜNTZING, A. 1959. Darwin's views on variation under domestication in the light of present-day knowledge. Proc. Am. Phil. Soc., 103 (2): 190-220.
NILSSON-EHLE, H. 1915. Den modärna ärftlighetsläran och dess betydelse för växtodlingen [Modern genetics and its importance for plant husbandry]. Stockholm, Nordiska Bokhandeln. 83 p.
NILSSON-EHLE, H. 1936. Über eine in der Natur gefundene Gigantsform van Populus tremula. Hereditas, 21: 379-382.
NISHIMURA, S. 1960. Chromosome numbers of poly-embryony seedlings of Pinus thunbergii Parl. J. Jap. For. Soc., 42: 263-264.
PEDIGO, R. A. 1960. The effects of ionizing radiation on the ecology of Pinus taeda. Bull. Ecol. Soc. Am., 41: 94.
PEDIGO, R. A. 1962. The effects of ionizing radiation on the ecology of Pinus taeda L. Unpublished Ph. D. dissertation, Emory Univ.
PETRINI, S. 1959. De två äldsta svenska tallproveniensförsöken [The two oldest pine provenance trials in Sweden]. Medd. Statens skogsforsk. Inst. Stock., 48 (11): 1-49.
PRATT, R. B. 1963. Ecological effects of ionizing radiation on organisms, communities, and ecosystems. Proc. 1st Nat. Symp. Radioecol. AIBS and Reinhold Publ. (In press)
PLYM FORSHELL, C. 1953. Kottens och fröets utbilding efter själv- och krosbefroktning hos tall (Pinus silvestris) [The development of cones and seeds in the case of self- and cross-pollination in Pinus silvestris L.]. Medd. Statens skogsforskn. Inst. Stockh., 43 (10): 1-42.
PENNER, O. 1934. Die pflanzlichen Plastiden als selbstständige Elements der genetischen Konstitution. Ber. Math. - Phys. Kl. Sächs. Akad. Wiss. (Leipzig), 86: 241-266.
RIGHTER, F. I. 1946. New perspectives in forest tree breeding. Science, 104: 1-3.
RIGHTER, F. I. 1960. Forest tree improvement through inbreeding and interspecific hybridization. Proc. 5th World For. Congr., Seattle, U.S.A., p. 783-787.
SCHWANITZ, F. 1957. Die Entstehung der Kulturpflanzen. Berlin. 151 p.
SENGBUSCH, R. VON. 1952. Ein weiterer Beitrag zur Vererbung des Geschlechts bei Hanf als Grundlage für die Züchtung eines monözischen Hanfes. Z. Pflanzenzücht, 31: 319-338.
SIMAK, M. & GUSTAFSSON, A. 1954. Fröbeskaffenheten hos moderträd och ympar av tall [Seed properties in mother trees and grafts of Scots pine]. Medd. Statens skogsforshn. Inst. Stockh., 44: 2, 1-73.
SISOV, L. A. 1937. Breeding of hemp. In Vavilov, N. I. Handbook of plant breeding. Leningrad, 1937. (Cited from R. von Sengbusch, 1952)
SPARROW, A. H. & MIKSCHE, J. P. 1961. Correlation of nuclear volume and DNA content with higher plant tolerance to chronic radiation. Science, 134: 282-283.
SPARROW, A. H. et al. 1963. The radiosensitivity of gymnosperms. I. The effect of dormancy on the response of Pinus strobus seedlings to acute gamma irradiation. Radiation Botany. (In press)
STEBBINS, G. L. 1950. Variation and evolution in plants. New York, Columbia University Press. 643 p.
STEBBINS, G. L. 1956. Artificial polyploidy as a tool in plant breeding. In Genetics in plant breeding, Brookhaven Symp. Biol. No. 9, p. 37-52.
SUGIHARA, Y. 1940. On the multipartite chromosome-ring in Cephalotaxus drupacea Sieb. et Zucc. Sci. Rep. Of Tohoku Imp. Univ. Sendai, 4: 13-18.
SYLVÉN, N. 1910. Om pollineringsförsök med tall och gran [Experiments in self-pollination with pine and spruce]. Medd. Statens skagsförsökeanstalt, 7: 219-228.
TODA, R. 1957. Variation and heritability of some quantitative characters in Cryptomeria. Silvae Genet., 7: 87 -93.
TURRESON, G. 1922. The genotypical response of the plant species to the habitat. Hereditas, 3: 211-350.
WELLWOOD, R. W. & SMITH, J. G. H. 1962. Variation in some important qualities of wood from young Douglas fir and hemlock trees. Res. Publ. Faculty of Forestry, Univ. Brit. Columbia, No. 50, p. 1- 15.
WETTSTEIN, D. VON. 1961. Nuclear and cytoplasmic factors in development of chloroplast structure and function. Can. J. Bot., 39: 1537-1545.
WETTSTEIN, D. VON, GUSTAFSSON, A. & EHRENBERG, L. 1959. Mutationsforschung und Züchtung. Arbeitsgemeinschaft für Forschung Landes Nordrhein-Westfalen, 73: 7-48, 59:60.
WETTSTEIN, F. VON. 1926. Über plasmatische Vererbung, sowie Plasma- und Genwirkung. Nachr. Ges. Wiss. Göttingen. Math.-Phys. Kl., p. 250-281.
WIBECK, E. 1933. Till skogens raefrågå på lokala kallorter [Race problems of Scots pine in frigid localities]. Norrlands Skogvs. Förb. Tidskr., p. 227-250.
WINKLER, H. 1920. Vererbung und Ursache der Parthenogenese im Pflanzen- und Tierreich. Jena. 166 p.
WRIGHT, J. W. 1959a. Silvical characteristics of White Ash (Fraxinus americana). Northeast. For. Expt. Sta., Upper Darby, Pa. Sta. Paper No. 123, p. 1-19.
WRIGHT, J. W. 1959b. Silvical characteristics of Green Ash (Fraxinus pennsylvanica). Northeast For. Expt. Sta., Upper Darby, Pa. Sta. Paper No. 126, p. 1-18.
ZINNAI, I. 1953. The morphological characters and the fertillity of the pollen of a tetraploid Japanese red pine induced by the colchicine method. J. Jap. For. Soc., 35: 245-248.
ZOBEL, B. 1961. Inheritance of wood properties in conifers. Silvae Genet., 10: 65-70.
![]()
![]()
![]()
{kind=link}
{kind=link}