![]()
![]()
![]()
JONATHAN W. WRIGHT
La hibridación tiene una importancia variable en los programas de mejora genética de árboles. En las especies de Castanea y Populus (excepción hecha de los temblones) ha recibido la máxima prioridad a causa de la facilidad de los cruzamientos y de que los híbridos presentan propiedades que no se pueden obtener por selección dentro de una especie. En algunas especies de Pinus y Picea la hibridación deberá practicarse a la vez que el ensayo de procedencias y la selección por individuos, pues esta combinación de métodos será en ellos mucho más eficaz que aplicar un solo método. También hay algunas especies con las que ha fracasado todo intento de cruzamiento o que son claramente superiores a los híbridos hasta ahora producidos y ensayados.
Son frecuentes los híbridos naturales, pero probablemente no representan más que una pequeña proporción de las combinaciones posibles. El estudio de los datos disponibles sobre cruzamientos naturales es siempre un punto de partida bueno para un programa serio de hibridación. A veces el mejorador tendrá que guiarse sólo por los cruzamientos naturales, cuando las polinizaciones dirigidas sean difíciles de lograr en cantidad razonable. En otros casos, en cambio, las polinizaciones dirigidas serán tan fáciles de ejecutar que serán claramente las más indicadas.
La introgresión natural en gran escala es relativamente poco común, pero es interesante cuando se da, pues proporciona una visión teórica introspectiva de la naturaleza de la diferenciación genética y de la selección natural. Además, la gran variabilidad genética de un enjambre de híbridos es conveniente como punto de partida en programas de mejora selectiva.
Se conocen pocos híbridos intergenéricos. El término «híbrido», sin especificar, suele referirse al cruce entre dos especies. Populus y Castanea responden a un tipo de cruzabilidad relativamente sencillo, pudiendo combinarse entre si casi todas las especies del mismo género. El comportamiento es un poco más complejo en Picea, en el que las especies semejantes morfológicamente y vecinas se pueden cruzar mucho más fácilmente que las que están separadas geográficamente o son menos parecidas. En el otro extremo está el Pinus, que se subdivide en gran número de grupos intracruzables que guardan gran correspondencia con las series y secciones taxonómicas. Algunos grupos contienen varias especies mientras en otros sólo entra una, que se halla tan bien diferenciada que no se cruza con ninguna otra. El conocimiento de la cruzabilidad tiene gran interés pues permite al mejorados concentrarse en los cruces que ofrecen mayores posibilidades de éxito.
JONATHAN W. WRIGHT es Profesor de Genética Forestal, Departamento Forestal, Universidad del Estado de Michigan, East Lansing, Michigan, E.U.A. Fueron otros miembros del equipo de trabajo, C. Heimburger (Canadá), E. W. Jones (Reino Unido) y O. Fugalli (FAO)
El empleo reciente de agentes inductores de mutación y de polinizaciones mixtas abre nuevas posibilidades de aumentar el número de cruces con éxito.
Es relativamente frecuente que aparezca vigor híbrido en la generación F1, sobre todo cuando se trata de especies exóticas o cuando se ensayan híbridos en habitats perturbados. La aparición del vigor híbrido no puede ser predicha de antemano; sólo puede ser determinado empíricamente ensayando las plantas en cierto número de habitats. Aún no se sabe si el desarrollo superior de un cruce es debido a dominancia, superdominancia o a interacción entre parejas de genes que tengan efectos aditivos por si mismos. Este conocimiento no es necesario cuando se trata de emplear en la práctica los F1 pero de él depende si el vigor extra puede ser o no fijado en generaciones posteriores.
Algunas combinaciones híbridas pueden ser útiles aunque la F1 sea intermedia. El género Castanea proporciona un ejemplo. El Castanea dentaba es aceptable por casi todos los conceptos, pero le falta resistencia al chancro Endothia parasítica. Esta puede lograrse por cruce con especies asiáticas que dejan que desear en cuanto a calidad de madera y rapidez de crecimiento. El retrocruzamiento a lo largo de varias generaciones permite incorporar la resistencia a la enfermedad y calidad de la madera en los mismos árboles.
En un programa destinado a la producción de un cultivar F1 heterótico habrá que prestar gran atención a la cruzabilidad, técnicas de producción masiva y ensayos a largo plazo en las localidades de uso probable. Si el objetivo es un producto de segregación en generaciones posteriores, habrá que modificar algo la importancia de las técnicas. Consideraciones teóricas de genética de poblaciones indican que el objetivo puede alcanzarse mejor comenzando la selección de familias en F3. De ahí que se debe abreviar en lo posible las etapas requeridas por las generaciones F1 y F2.
Para obtener vigor híbrido o un segregado de generaciones posteriores no es preciso comenzar con especies distintas, sino con razas diferentes o porciones de clines dentro de una especie. Los investigadores suecos se han adelantado en este terreno cruzando razas nativas con otras continentales. Sus resultados preliminares muestran la conveniencia de una gran expansión de esta hibridación intraespecífica.
La hibridación entre especies es tema muy interesante. El investigador afortunado que llega a conseguir una combinación no conocida hasta el presente, tiene la sensación de que ha contribuido a algo nuevo que, con toda probabilidad, la naturaleza no hubiera producido por sí sola, con la posibilidad de que esta combinación conseguida por él tenga un día gran utilidad práctica. Muchas de las plantas de mayor importancia comercial son de origen híbrido y de los árboles híbridos nuevos debemos esperar aprovechamientos más ventajosos.
El estudio completo de la hibridación con un solo género, es empresa que puede ocupar toda una vida. El trabajo de los taxónomos sobre cruzamientos naturales raros o sobre introgresión proporciona una buena orientación sobre los cruzamientos que pueden emprenderse; por ejemplo, los híbridos naturales nos informan sobre la cruzabilidad en el género Eucalyptus.
Para conseguir cruzamientos satisfactorios, esa orientación debe completarse con numerosos trabajos de polinización dirigida, ya que una gran parte de las especies más fácilmente cruzadas no aparecen juntas en la naturaleza. Hay aún mucho que hacer en los estudios de cruzamiento para llegar a conocer que dos especies A y B pueden cruzarse y que existen posibilidades de que los resultados sean prometedores. Cuestiones fundamentales, cuya solución puede requerir muchos años, son: cómo pueden multiplicarse en masa los híbridos interesantes, y qué razas concretas, o individuos de las especies parentales son los que nos han de dar los mejores resultados.
El papel de la hibridación varía de unas regiones a otras y de unos géneros a otros. Existen grupos tales como Castanea en los cuales la mejor especie para madera es deficiente en un carácter solo (en este caso, resistencia a la enfermedad), que puede ser mejorado por cruzamiento con otra especie. La mayor parte de los híbridos son intermedios morfológicamente y en exigencias de estación entre ambas especies parentales.
Por eso, puede ser fácil formar híbridos, pero sin gran valor para el monte cuando no se dispone de zonas extensas de habitats intermedios. La perturbación causada por el hombre ha creado a veces habitats intermedios, a los que se adaptan mejor tipos intermedios de árboles, que exhiben vigor híbrido, ejemplos de lo cual se encuentran en Eucalyptus y Aesculus. También sucede a veces que una especie tiene su máxima aplicación fuera de su área natural, en cuyo caso los híbridos pueden resultar mejor adaptados, por ejemplo el híbrido Larix decidua X leptolepis. La hibridación ofrece relativamente poca esperanza en la mejora del Pinus resinosa americano, con el cual han fracasado casi todos los intentos de cruce con otras especies. En la región sudeste de Estados Unidos los pinos del Sur ofrecen pocas perspectivas de ser mejorados por hibridación interespecífica; se reproducen fácilmente y crecen aprisa, mientras ninguna otra especie de Pinus los sobrepasa en peso específico de la madera, una de las propiedades más importantes que tiene. En cualquier caso los híbridos deben sufrir una comprobación empírica antes de dar por buenas sus ventajas.
Son frecuentes los informes sobre híbridos naturales entre especies del mismo género. Existe también un informe fehaciente de hibridación Cupressus X Chamaecyparis. No es tan convincente, en cambio, el híbrido presunto Tsuga X Picea.
Hay gran probabilidad de obtener híbridos naturales cuando se siembra semilla recogida de individuos aislados que han crecido en arboretos o en parques. Cuando la especie no se autofecunda normalmente, la mayor parte de la semilla debe provenir del cruce con otras especies. Los trabajos en los viveros suelen ser también lo suficientemente intensivos para poder reconocer la aparición de tipos raros. La mayor parte de los híbridos naturales de Abies y Acer se encontraron por ese procedimiento, y también muchos de los conocidos de Picea, Pinus, Populus y Quercus.
En la mayor parte de los casos, la hibridación natural produce sólo uno o unos pocos árboles F1. El grupo de híbridos subsiste por una generación, después desaparece. La próxima repetición de la misma combinación puede tener lugar en localidad completamente diferente. Así ha sucedido, por ejemplo, con la mayor parte de los híbridos de Quercus reseñados en América y también con algunos híbridos interesantes de pino, tales como Pinus sylvestris X nigra y el P. palustris X taeda.
El estudio de unos pocos casos aislados de hibridación natural de Populus, Pinus y Picea provocó el ímpetu inicial del trabajo intensivo posterior sobre hibridación artificial, pero ha sido el estudio de los miles de hibridaciones artificiales realizados lo que ha dado más luz sobre el problema. En cambio, en la hibridación de Quercus y Aesculus se ha aprendido más de casos esporádicos que de las hibridaciones artificiales difíciles de conseguir en cantidad.
Los enjambres de híbridos naturales no son frecuentes, pero sí muy instructivos. Se presentan generalmente en habitats «híbridos» y especialmente en áreas perturbadas. Uno de los mejores ejemplos estudiados sobre este asunto son los robles del sudoeste de los Estados Unidos (Muller, 1952). En la parte central y norte de Texas, el Quercus havardii y el Q. stellata ocupan típica y respectivamente suelos arenosos y arcillosos. Los híbridos son frecuentes en la zona de contacto entre los dos suelos, pero no lejos de ella. La forma «pura» de cada especie se encuentra en su habitat puro, típico de la especie. En la misma región citada, el Q. mohriana y el Q. havardii aparecen en suelos de origen calizo y arenoso, respectivamente. Los híbridos se presentan en los sitios en que el suelo está compuesto por una mezcla de caliza y arena, pero no en los habitats puros. En California, la especie híbrida Quercus X alvordiana es realmente un grupo de enjambres de híbridos entre Q. douglasii y Q. turbinella y no se encuentra más que en habitats intermedios donde los dos padres establecen estrecho contacto. Algunos de estos enjambres de Quercus X alvordiana han sobrevivido a través de muchas generaciones desde el fin del Período Pleistoceno.
En Australia, el Eucalyptus dalrympleana era uno de los componentes de la selva virgen en áreas no perturbadas. Al intervenir el hombre, muchas de ellas fueron perjudicadas grandemente por los incendios. Hoy en día esas superficies están siendo invadidas por enjambres de híbridos de E. pauciflora X divas (Pryor, 1957), mientras que bajo condiciones naturales se excluirían mutuamente a causa de la competición. Recientemente se han encontrado otros ejemplos de enjambres de híbridos de Eucalyptus en habitats perturbados.
El Aesculus, sec. Paria, incluye cuatro especies nativas de la parte central y sudeste de Estados Unidos. En el monte original las especies permanecían bien distintas. Sólo se formaron algunos híbridos aisladamente o en grupos muy pequeños. Hoy en día hay muchas trochas abiertas para caminos y superficies cortadas, en las cuales los híbridos sobrepasan a las especies parentales. En habitats intermedios de ese tipo existen accidentalmente enjambres híbridos de A. glabra X pavia, A. glabra X octandra y A. octandra X sylvatica. En estos dos últimos casos la hibridación ha provocado un considerable flujo de genes del A. octandra a las otras dos especies, y en la combinación A. pavia X sylvatica la abundancia del flujo de genes ha tenido lugar en ambas direcciones entre las respectivas especies parentales.
Por introgresión se comprende un período largo de hibridación seguido de un flujo considerable de genes de una especie a otra. Muchas veces es difícil distinguir entre la convergencia de especies por introgresión y la divergencia de especies o razas. En Picea, por ejemplo es probable que la introgresión sea un factor importante en la adaptabilidad de las especies de este género de la zona circumpolar norte, si bien es verdad que no se ha comprobado más que en dos casos (Wright, 1955). El cruce de Picea engelmannii y P. glauca aparece fácilmente en el interior de Columbia británica y en Alberta. Se han encontrado caracteres de una especie en la otra, lejos de la zona de contacto de ambas. Las masas de Picea del norte de Escandinavia y zonas adyacentes de la U.R.S.S. son una mezcla de grados intermediosentre P. abies y P. obovata. En otros casos se sabe que especies muy difundidas con áreas adyacentes se hibridan artificialmente y se presume que lo habrán hecho naturalmente en el pasado cuando el clima haya favorecido el solape de sus áreas. En cambio, son características de las muy dispersas especies meridionales de Picea la carencia de variabilidad genética y de oportunidades recientes de hibridación.
Después de la introducción por el hombre de Populus deltoides de Norteamérica en Europa meridional, ha habido una gran introgresión de aquella especie en el P. nigra. Hoy en día, en muchas localidades, es raro encontrar individuos puros de P. nigra, y el material de viveros es en su mayoría de híbridos (Schreiner, 1959).
Entre los robles es poco frecuente la introgresión a pesar del gran número de especies registradas. Aparecen tipos intermedios entre las dos especies del norte de Europa Q. robar y Q. petrea allí donde se sobreponen sus áreas naturales y pueden ser híbridos. En el sudoeste de Estados Unidos, Quercus X drummondii parece ser una derivación introgresiva de Q. stellata con una instilación de genes de Q. gambelii. En la mayoría de los demás casos las especies han mantenido su identidad incluso creciendo mezcladas con varias otras clases de robles (Palmer, 1948).
Hay tres razones que ayudan a explicar por qué algunas veces la hibridación da lugar sólo a unos pocos tipos intermedios aislados, y otras, en cambio, a enjambres de híbridos o introgresiones. La primera, que puede haber sólo limitadas oportunidades para el cruce. Se han encontrado en arboretos algunos híbridos naturales del cruce entre P. strobus de América oriental y el P. griffithii del Himalaya. Estos híbridos suelen aparecer en el norte de Italia donde ambas especies están a menudo juntas en jardines y parques. El cruzamiento artificial es fácil; es lógico que hubiera habido introgresión si las especies parentales hubieran ocupado la misma zona. El que Acer rubrum y A. saccharinum no se conviertan en una sola especie; es debido indudablemente a su diferente época de floración. Esto explica que no se hayan observado sus híbridos naturales, aunque vivan mezcladas generalmente y sea fácil la hibridación artificial de ambas especies.
La segunda razón es la posible esterilidad de los híbridos, aunque por ahora se sabe de pocos casos en que suceda esto.
La tercera razón es la naturaleza e intensidad de la presión selectiva. Supongamos que los genes A y a son alelomorfos recíprocos, con frecuencias respectivas p = 0,9 y q = 0,1. Con apareamientos al azar los cigotos estarán presentes con frecuencias relativas de (p + q)² = p² + 2pq + q² = 0,81AA + 0,18Aa + 0,01aa. Si la selección favorece a los heterocigotos por un período de una generación, la proporción de árboles Aa se elevará a un máximo (siendo el máximo teórico 0,50). Pero si entonces cambian las condiciones del medio y dan lugar a que la selección favorezca a los homocigotos, los árboles Aa tenderán a desaparecer. Con eliminación completa de heterocigotos, un grupo híbrido en que la frecuencia de cigotos fuera 0,81 AA: 0,18 Aa, 0,01 aa producirá gametos (polen y ovocélulas) en los cuales la frecuencia de genes será 0,988 A: 0,012 a. Estos gametos producirán una nueva generación de árboles en los cuales la frecuencia de cigotos será 0,9757 AA: 0,0242 Aa: 0,0001 aa. Nótese que la selección contra los heterocigotos ha reducido la frecuencia de los árboles Aa: en un 90 por ciento y la del homocigoto menos común (aa) en 99,9 por ciento. Es decir, que quedan muy pocos genes a con que poder producir árboles Aa para la generación siguiente. Si el sentido de la selección vuelve a invertirse otra vez favoreciendo los árboles Aa, la formación de un enjambre de híbridos empezaría de nuevo.
Este ejemplo matemático tiene dos corolarios. Uno, que la existencia de un enjambre de híbridos persistente es prueba de que la selección favorece a los hoterocigotos; es decir, que existe vigor híbrido aprovechable. Y otro, que la ausencia de un enjambre de híbridos no significa necesariamente que no se puedan conseguir híbridos útiles. Artificialmente se puede aprovechar el vigor híbrido manifestado en dos de cada tres generaciones, aun cuando la naturaleza por sí sola no lo pueda hacer.
Si el interés de un mejorador está en sacar ventaja de la superdominancia¹ constatada por la existencia de un enjambre de híbridos, debe intentar la multiplicación masiva por medios artificiales de poblaciones F1 o quizás de F2, porque en estas generaciones es donde se da más pronunciada aquella superdominancia. Sin embargo, si el investigador no está en condiciones de emprender un programa tan intenso, no deberá intentar procedimientos intermedios, tales claras de carácter genético. Por otro teorema sobre genética de poblaciones, se puede demostrar que tales poblaciones híbridas llegan a alcanzar un equilibrio natural cuando la frecuencia del gene a que denominaremos q, sea
¹ So dice que hay superdominancia cuando el hoterocigoto es superior al mejor de los homocigotos.
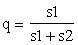
siendo s1 y s2 la presión selectiva contra los árboles AA y aa, respectivamente. Se puede también demostrar que la población llega a su máxima producción cuando alcanza ese equilibrio natural (si es que no puede mantenerse en un estado de 100 por ciento de heterocigotos por cruzamientos artificiales de árboles AA y aa, Citaremos un ejemplo: si los árboles AA, Aa y aa, tienen clasificaciones de idoneidad de 0,60, 1,00 y 0,85, respectivamente, la población híbrida, si está en equilibrio, tendrá una idoneidad media si se halla en equilibrio de 0,89. La selección artificial que favoreciese a los árboles mejores (que serían los heterocigotos) aumentaría el número de árboles improductivos AA y disminuiría la idoneidad media a 0,862, mientras que empleando la polinización artificial para asegurarse de que sólo se han plantado árboles Aa, se aumentaría la idoneidad media a 1,00.
Si el interés primordial del investigador se dirige hacia la variancia genética aditiva, un enjambre de híbridos nativos será un excelente punto de partida para un programa de selección. En el sur de Michigán existe un gran grupo de híbridos entre Pinus thunbergii y P. densiflora. En varios caracteres la variancia genética aditiva dentro de este grupo es probablemente dos o tres veces mayor que la que pueda encontrarse en cualquier población natural pura de dichas especies, y en cuanto a la heredabilidad y réplica a la selección serían también dos o tres veces mayores que en las masas puras.
Con la excepción antes mencionada de Cupressus x Chamaecyparis pueden dejarse a un lado los híbridos intergenéricos en zonas templadas y fijar la atención mejor en los cruces dentro de los géneros. Los tipos de cruzabilidad más conocidos son en Populus, Pinus, Picea, Quercus, Castanea y Larix. Dentro de estos grupos se han hecho suficientes intentos de cruce para poder hoy formular reglas de tipo general que permiten prever cuáles son las combinaciones de especies con más probabilidades de {éxito. Para Acer, Aesculus, Betula, Fraxinus y Ulmus, la información hoy existente no es tan completa y sólo permite las generalizaciones más amplias. Para la mayor parte de los géneros (incluyendo todos los tropicales) es aún menos completa y consiste casi exclusivamente en listas de híbridos naturales.
La cantidad de diferenciación genética dentro de lo que se considera como un género, varía mucho. En un extremo están los Populus. Hay dentro de él tan poca diferenciación genética que cada una de sus especies puede a primera vista ser cruzada con cualquiera de las demás. Castanea puede tener un tipo análogo, pues han resultado bien todos los cruzamientos intentados. En Picea la diferenciación genética es algo más pronunciada, pues la mayor parte de sus especies pueden cruzarse con sus vecinas geográficas, pero no con los procedentes de localidades alejadas. No existe una división marcada en distintos grupos cruzables y es posible (aunque no se haya comprobado aún) que cada una de sus especies pueda ligarse con cualquier otra, recurriendo al cruce con tipos intermedios. Los Eucalyptus presentan un tipo similar en el que las especies de la misma sección se cruzan con relativa facilidad. Los Quercus se subdividen en tres subgéneros (de los cuales sólo dos son conocidos genéticamente). Se conoce un gran número de híbridos entre especies pertenecientes al subgénero roble blanco Lepidobalanus y tal vez pudieran hibridarse dos cualesquiera robles blancos. Lo mismo puede decirse del subgénero de roble rojo Erythrobalanus. En cambio, no hay pruebas convincentes de hibridación entre subgéneros de roble.
En el otro extremo, podemos considerar a los pinos. El género Pinus se divide en dos subgéneros y en un gran número de series. Salvo tres excepciones, los cruces dentro de la misma serie han sido siempre conseguidos. El caso de los Acer es análogo al de los pinos. De 50 cruces artificiales intentados sólo se consiguieron cinco. Y éstos fueron siempre dentro de una de las 13 secciones reconocidas. Desde el punto de vista genético, las series o secciones en cualquiera de estos dos géneros puede admitirse tienen aproximadamente la misma cantidad de diferenciación que algunos géneros completos. Esto podría inducir a considerar a otros pequeños grupos desde el punto de vista biológico (pero no del práctico) como géneros independientes, como hicieron ya algunos autores tiempo atrás.
Las semejanzas morfológicas o fisiológicas y la distribución geográfica dan una orientación excelente sobre los tipos de cruzabilidad en un género. Las combinaciones más fáciles son las realizadas entre especies similares de zonas próximas que casi llegan a solaparse. Sucede a menudo que la distancia constituye un obstáculo suficiente, de modo que no fue necesario ningún otro dispositivo de aislamiento. Si ese obstáculo llega a salvarse por medio de plantaciones intermedias o por la polinización artificial, los híbridos surgen con facilidad. Por otra parte, es corriente que especies que viven mezcladas en la misma masa no se crucen. Algunas veces, diferencias en la época de floración o en las exigencias de su microestación, constituyen esa barrera de aislamiento que puede salvarse fácilmente por cruzamiento artificial Otras veces ese aislamiento es producido por diferenciaciones génicas o cromosómicas. La irradiación del polen y el empleo de mezclas polínicas quizá pudieran servir para superar este obstáculo. También es difícil conseguir cruces entre especies distintas morfológicamente o especies de áreas de distribución muy separadas. En tales casos su gran separación suele ir acompañada de grandes diferenciaciones de tipo génico o cromosómico.
Cruces que fallan en un determinado ambiente pueden conseguirse en otro sitio o momento. El conocimiento de la naturaleza del impedimento ayuda muchas veces a anularlo.
Existen numerosos ejemplos de la relación entre la cruzabilidad, la semejanza y la distribución geográfica. Para explicar mejor el asunto puede citarse el género Pinus. La serie Strobi abarca varias especies semejantes: todas ellas, excepto una, tienen áreas de distribución separadas (aunque próximas si se consideran bajo el punto de vista geológico), y pueden cruzarse unas con otras con relativa facilidad. La única excepción (Pinus lambertiana) no es típica de la serie y no se cruza con las demás. Se han hecho intentos de cruzar especies de la serie Strobi con individuos de otras series, pero no hubo éxito más que en un solo caso.
El Pinus ser. Sylvestres abarca unos doce pinos duros. Los asiáticos son semejantes morfológicamente y se cruzan unos con otros en casi todas las combinaciones posibles. La semejanza entre ellos y los representantes de la serie en Europa es menor y sólo se han conseguido unos pocos cruzamientos europeos x asiáticos, que dieron escasa producción de semilla. El único representante de los Estados Unidos en la serie Sylvestres (P. resinosa) tiene varios rasgos de diferenciación morfológica y cromosómica. Tras un esfuerzo de 20 años, participando muchos investigadores, sólo se consiguieron cuatro híbridos entre P. resinosa y otra especie.
El Pinus ser. Australes abarca grupos de especies del sudeste y sudoeste de Estados Unidos. Los individuos de cada grupo son más semejantes entre sí que con los miembros del otro grupo. Se han conseguido híbridos auténticos de todas las combinaciones posibles de especies sudeste x sudeste y de una gran parte de las sudoeste x sudoeste. En cambio, no se ha conseguido ninguno de las sudoeste x sudeste (Duffield, 1952).
Los abetos (Picea) muestran también la relación entre cruzabilidad, semejanza y distribución geográfica. En el norte de Estados Unidos, Canadá y nordeste de Asia, existe un grupo de cinco especies, cada una de ellas con área de distribución casi separada y todas con cierta semejanza a las especies vecinas. Todos los intentos de hibridación dentro de ese grupo fueron conseguidos, y en cambio no lo fueron los de aquellos abetos con otros de fuera. Hay otra «cadena» de especies similares que cruza el norte de Europa, Siberia y el norte de China, en la cual los cruzamientos se dan con suma facilidad. Las cinco especies que cohabitan en la misma montaña del Japón no se cruzan entre sí y, sin embargo, una de ellas lo hace con abetos del continente. Una de las excepciones notables al tipo general de cruzabilidad lo constituye el cruce P. omorika X sitchensis, logrado en Europa.
El género Larix tiene unas 10 especies, cada una con su zona geográfica propia, separada. Su diferenciación parece ser exclusivamente de carácter geográfico, siendo fácil el cruzamiento de las especies.
Con el Fraxinus de siete intentos de hibridación artificial sólo se ha conseguido uno y fue entre una especie del este de los Estados Unidos y otra de la misma sección del oeste de los Estados Unidos. Las dos especies pueden considerarse próximas desde un punto de vista geológico y son tan semejantes que cualquier persona puede tomar un ejemplar aberrante de una de ellas por la otra. En el sudoeste de Europa se tiene conocimiento de enjambres híbridos entre miembros de una pareja semejante de especies.
Se conocen casos de varios cruzamientos entre especies con diferente número de cromosomas que han producido buena semilla. La relación entre el número de cromosomas y la facultad de hibridación aún necesita ser más estudiada.
Todas estas generalizaciones podemos resumirlas haciendo observar que para proyectar un programa de cruzamiento de especies es de gran ayuda disponer de una buena monografía y una detallada serie de mapas de distribución geográfica. Si admitimos la existencia de 90 especies en el género Pinus, el número de combinaciones diferentes de especie x especie sería de 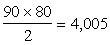 . Y por la labor hecha hasta hoy se podría esperar que el 10 por ciento de ellas, o sea 400, tendrían éxito. Cruzando dos especies al azar, habría un 10 por ciento de probabilidad de éxito; pero estudiando la taxonomía y distribución podría elevarse esa probabilidad hasta un 80 por ciento.
. Y por la labor hecha hasta hoy se podría esperar que el 10 por ciento de ellas, o sea 400, tendrían éxito. Cruzando dos especies al azar, habría un 10 por ciento de probabilidad de éxito; pero estudiando la taxonomía y distribución podría elevarse esa probabilidad hasta un 80 por ciento.
Lo que más se ha buscado con las hibridaciones ha sido la obtención de híbridos F1 vigorosos. La labor hecha hasta ahora ha demostrado que puede llegarse a resultados muy prometedores, pues aunque la mayoría de los híbridos carecen de utilidad, hay un número suficiente que muestra vigor híbrido compensando los trabajos empleados. Puede ocurrir que el vigor híbrido sea patente en una parte del área natural de una especie y en cambio no aparezca en otra.
El proceso de introducir una combinación híbrida acertada F1 en la práctica comercial comprende cuatro etapas. La primera es estudiar las posibilidades de cruce para determinar qué combinaciones pueden hacerse. Como ya hemos dicho, estos estudios están bastante avanzados en algunos géneros y apenas iniciados en otros. La segunda es producir unos pocos híbridos de cada una de las combinaciones más prometedoras y hacer su ensayo comparativo con los individuos no híbridos a quienes deben reemplazar. En este sentido, se ha hecho bastante labor, pero en casi ningún caso los estudios han sido lo suficientemente extensos para conocer la verdadera potencialidad del híbrido. La mayor parte de los híbridos de Pinus obtenidos en California y Pensylvania apenas son conocidos fuera de unos pocos lugares de experimentación y la potencialidad de Larix decidua X leptolepis, tan útil en Europa, es casi desconocida en América. La tercera etapa es la determinación de las posibilidades de su producción en masa. Esto no es problema en las especies de Populus que pueden propagarse perfectamente por estaquilla, pero sí lo es en otros géneros. El Instituto de Genética Forestal de Placerville, California, hizo bastantes trabajos con varias combinaciones de Pinus para demostrar que es posible la producción en masa. En los últimos años, el Instituto Coreano de Genética Forestal ha hecho también una gran labor, con resultados muy esperanzadores, en unas pocas combinaciones de Pinus. La cuarta etapa para introducir con éxito un híbrido F1 es la de determinar cuáles son los árboles que deben ser preferidos como genitores. Este estudio apenas se ha iniciado para algunas combinaciones de especies. Tomemos, por ejemplo, el cruce de Pinus sylvestriis X densiflora. Se obtuvo primero en Connecticut y después en Pensylvania. En los dos casos se tomaron como genitores los primeros árboles disponibles de origen completamente desconocido. Los híbridos crecieron con extraordinaria rapidez, pero no han podido utilizarse por su mala forma y escasa producción de semilla.
Las estaciones experimentales forestales del sudoeste y del nordeste del Servicio Forestal de los Estados Unidos han obtenido un gran número de combinaciones con especies de Pinus. Aquí sólo pueden mencionarse unas pocas. Pinus monticola X strobus es una de las más prometedoras. Su producción de semilla es lo suficientemente buena para augurar grandes posibilidades en la producción en masa. Su crecimiento en California y en otras muchas partes de los Estados Unidos iguala o excede al de la especie parental de mayor desarrollo. Actualmente se trabaja intensamente con ambas especies parentales para desarrollar cultivares resistentes a la enfermedad y cuando esto se haya conseguido, será posible contar con híbridos F1 verdaderamente notables. El híbrido P. attenuata X radiata es también prometedor. Podría producirse en cantidad. Aunque no sobrepasa la rapidez de crecimiento de la especie parental más rápida (P. radiata) en localidades templadas, parece que llegará a ser uno de los pinos más productivos en sitios demasiado fríos para aquella especie.
El híbrido P. densiflora X thunbergii da grandes cantidades de semilla. Esta combinación muestra un gran vigor híbrido en Pensylvania, pero no en el Japón de donde son nativas las especies parentales. En cambio, la utilidad potencial de una cuarta combinación, P. nigra X resinosa, se ve limitada por dificultades de multiplicación. Al cabo de muchos años de trabajos sólo se obtuvieron 4 híbridos que son de buen crecimiento, pero que han quedado sin más provecho que el de curiosidades botánicas.
En Corea es donde se ha explotado al máximo la potencialidad de los híbridos F1 de pino. Se han destinado cada año a cruces de P. rígida X taeda miles de bolsas aislantes. El proyecto parece ser económico. Los híbridos producen mayor renta aunque su obtención sea más cara que las especies puras.
El cruce Larix decidua X leptolepis ha sido objeto de más de 27 publicaciones de otras tantas regiones del norte de Europa, todas ellas ensalzando su mayor crecimiento sobre el de las especies parentales (para referencias véase Wright, 1962). Su producción de semilla es alta, pero el Larix no se presta tan bien a la polinización dirigida en masa como lo hace el Pinus. Por ello Syrach Larsen, Dinamarca, ha ideado un huerto para semilla constituido por un clon autoincompatible de una especie y planta de semilla o clonal de la otra. El clon autoincompatible hace de genitor femenino y produce la semilla híbrida.
En América y aún más en Rusia se ha trabajado mucho en la obtención de varias combinaciones híbridas artificiales de Quercus. Algunas de ellas han resultado intermedias entre los genitores, pero otras han demostrado un vigor híbrido utilizable (Piatnitsky, 1960). Sin embargo, actualmente parecen ser de poca utilidad incluso las últimas combinaciones. La polinización dirigida es extremadamente enojosa y probablemente antieconómica en escala comercial. La multiplicación vegetativa no es practicable. Podrían ser una solución los huertos de cruzamiento natural, pero los esfuerzos hechos para encontrar árboles que den grandes porcentajes de semilla híbrida en polinización cruzada natural no han sido compensadores.
La silvicultura de Populus se basa más en los híbridos que en las especies puras. En Europa comúnmente se plantan cultivares derivados de cruzamientos entre P. deltoides, P. nigra, P. trichocarpa y otras especies. Muchos de ellos no han sido ensayados comparativamente con sus genitores, pero se da generalmente por descontado la existencia de vigor híbrido. Como resultado de los trabajos de Schreiner, en el nordeste de los Estados Unidos, en los que fue un iniciador, se han probado muchos híbridos prometedores para América y pronto podrán ser plantados en escala comercial.
Los Populus, fuera de los temblones, pueden multiplicarse fácilmente por estaquilla y su propagación masiva por tanto no constituye problema. Su facilidad de enraizamiento ha permitido además a los mejoradores realizar un gran número de ensayos sobre diferencias clonales entre plantas hermanas F1 que tienen comunes los dos genitores. Como era de esperar, se han hallado raramente diferencias clonales significativas, excepto cuando los genitores mismos eran híbridos.
En ensayos a corto plazo, Acer platanoides x mayrii y A. platanoides X cappadocicum han dado vigor híbrido. Los cruzamientos a escala experimental eran fáciles de conseguir con un rendimiento de algunos cientos al día. Este rendimiento, sin embargo, no se pudo aumentar mucho por la estructura de la flor; la polinización artificial en masa no parece factible. Ni sirven los huertos de cruzamiento natural por la diferencia en las épocas de floración. La solución pudiera ser la poliploidia. Quizás de los pocos híbridos disponibles podrían originarse formas nuevas estables duplicando el número de sus cromosomas. Pero ¿sería el tetraploide tan ventajoso como el diploide? Esto es lo que hay que averiguar.
Teóricamente se puede esperar que persista una cierta cantidad de vigor híbrido en la generación F2. Esto aumenta la posibilidad de aprovechar la generación F2 cuando la F1 no se puede obtener económicamente. Por lo menos, en algunos de los pinos los F1 se sabe que son lo suficientemente fértiles para que se produzca semilla de los F2 en cantidad. El Instituto de Genética Forestal de California ha buscado esta posibilidad en varias combinaciones de Pinus. En el Reino Unido se proyecta conseguir abundantes descendencias Fa de alerce europeo X japonés (Larix decidua X leptolepis) con la esperanza de que persistirá algún vigor híbrido en esta generación.
En la mayor parte de las ilustraciones citadas aquí, los híbridos son jóvenes todavía, ya que no hay ensayos que alcancen edades adecuadas en las conbinaciones interesantes. Además, muchos de los ensayos preliminares se han realizado en una o dos estaciones solamente, y es de todos sabido que el vigor híbrido puede manifestarse en una localidad y no en otra. Por eso queda todavía por hacer una labor considerable a este respecto.
Prácticamente todos los híbridos obtenidos hasta hoy de Pinus, Picea y Populus lo han sido aprovechando cualquier genitor disponible, sin selección previa alguna. Este tema debe preocupar en lo sucesivo a los mejoradores. Los primeros Pinus densiflora X P. thunbergii obtenidos en Filadelfia parecen muy útiles, pero es inconcebible que puedan ser los mejores. Lo probable es que sean superados cruzando razas selectas de cada especie o quizás individuos escogidos dentro de esas mismas razas destacadas.
Casi todos los hibridadores tienen la esperanza de producir generaciones F1 ó F2 que posean el suficiente vigor híbrido para que merezca la pena su plantación en escala comercial. Pero esto no siempre es posible. Algunas veces se obtendrá el producto más apreciado por segregación en generaciones posteriores. Esto puede ser cierto si se cumplen tres circunstancias:
1. si la polinización dirigida es tan difícil que los F1 y F2 no se puedan producir en cantidad (Juglans Castanea, Acer, Quercus y posiblemente Abies);2. si, existiendo vigor híbrido, no es lo suficientemente destacado para que merezca la pena de crear un cultivar utilizable en la práctica (Castanea dentaba X mollisima);
3. si la variancia genética es principalmente aditiva.
Si lo que se desea es incorporar un carácter de la especie A con muchos caracteres de la especie B, lo que procede es el retrocruzamiento. Habrá que producir artificialmente el híbrido F1 entre A y B y después retrocruzarlo con la especie B, y si fuera necesario, volver a repetir la misma operación. En las plantas de cultivo suelen ser necesarias hasta cuatro o cinco generaciones retrocruzadas. Si se desea incorporar la misma cantidad aproximadamente de las especies A y B, se produce una generación F1; se cruzan entre los F1 para producir una F2, éstos se cruzan entre sí para formar una F3, etc.
La necesidad de hacerlo así se ha venido reconociendo desde hace tiempo, pero los investigadores se resisten a seguir este procedimiento por el tiempo que requiere. La espera puede ser indispensable, pero no indefinida. Hay muchas especies importantes de árboles que producen flor a la edad de uno a cinco años. Si desde el principio el mejorador conociese su bondad, en 20 anos podría llegar a generaciones F4 en plena producción.
Actualmente los proyectos de mejora en generaciones múltiples son consecuencias improvisadas de experiencias sobre hibridación F1. Esta situación debe enmendarse si se pretende progresar más rápidamente. Los ensayos de campo proyectados para obtener formas segregadas en generaciones posteriores deben ser distintos de los que se proyecten para probar la utilidad de las generaciones F1 y F2 per se.
El objetivo de un proyecto con generaciones múltiples debe ser producir y probar una F3 o un retrocruce por segunda generación, lo antes posible. ¿Por qué? La generación F1 es una generación uniforme y por lo tanto no da oportunidades de poder seleccionar.² Supongamos que tenemos 25 brinzales F1 y que 10 de ellos fructifican a muy corta edad. El componente genético de la variación en la población F1 es casi nulo y, por tanto, hay poca diferencia en usar esos 10 como reproductores para una generación Fa o en esperar varios años hasta ver si las otras 15 plantas llegan a tener mejores fenotipos. La población Fa es variable desde el punto de vista genético y ofrece oportunidades para seleccionar genéticamente. Pero una gran proporción de la variancia Fa no es aditiva y por tanto imposible de fijar por cruzamiento de los mejores árboles entre sí. Esto justifica que el mejorados no haga más que una valoración moderadamente buena sin excesivo detenimiento de los F2, puesto que por mucho que mire a los árboles no podrá conocer su potencial de mejora genética como lo conocerá esperando a las generaciones F3 y F4.³ Es preferible observar a F2 unos pocos años y hacer una selección moderada. Puede escogerse, por ejemplo, el 10 ó 20 por ciento de los árboles mejores y utilizarse como genitores para un ensayo con repeticiones de F3 en el cual se mantendrá la identidad de todas las familias.4
² Es cierto que el árbol en estado natural es variable. El híbrido F1 entre la raza I de la especie A y la raza VI de la especie B no será absolutamente isogénico, pero lo será relativamente en comparación con la mezcla entre las dos. La selección dentro de tal F1 ofrece tan pocas posibilidades de mejora (en comparación con las que ofrece la selección en posteriores generaciones) que puede no tomarse en consideración.Esta ineficacia de selección en El se puede aplicar también a la selección dentro de las descendencias. Supongamos que la raza I de la especie A se cruza con la raza VI de la especie B y que la raza IV de la especie A se cruza con la II de la B. Hay que esperar que las dos descendencias F1 difieran y proporcionen la base para dos proyectos de selección que podrán unirse más tarde.
³ La cuestión de hasta cuando se debe valorar la F2 antes de proceder a la producción de F3 no se puede contestar categóricamente. Probablemente en una población F2 existe suficiente variancia genética aditiva que un error sería emplear el primer I por ciento de los árboles que florezcan si se notasen francamente deficientes en cualquier característica importante de su desarrollo. El vigor híbrido pronunciado en los F1 y F2 indica la presencia de una variancia no aditiva. Se deduce, por tanto, que cuanto mayor es la desviación de F3 y de F2 de la semisuma de las medias parentales, más corto podrá ser el período que se necesita para los ensayos. Si los ensayos de crecimiento demuestran que F1 iguala los promedios de las medias parentales en todos los aspectos, podrá empezarse la selección de F2 en las mismas condiciones que en F3.
4 El término familia se emplea para designar la descendencia de un individuo por polinización libre o la obtenido por un solo cruzamiento entre dos individuos.
Ha sido mencionado el hecho de que la selección es más provechosa en la generación F3 que en la F2 debido al valor menor de la variancia genética no aditiva. Otra razón para preferir la F3 es el hecho de que ofrece oportunidad para hacer la selección de familias. Esto significa seleccionar el material de mejora fijándose en el fenotipo de un árbol y en el comportamiento de otros que tienen el mismo o los mismos genitores que él, lo cual es mucho más eficaz que seleccionar sólo a base de fenotipos. (Para una buena discusión comparativa sobre las ganancias genéticas por selección en masa o de familias, véase Lerner, 1958.)
Cuando se procede a un programa de mejora selectiva, como el que acabamos de explicar ¿será más conveniente emplear descendencias de polinización artificial (los dos genitores conocidos) o, por el contrario, descendencias de libre polinización (conocido sólo el árbol que dio la semilla)? Cuando las poblaciones F1 y F2 pueden ser relativamente pequeñas, de unas pocas docenas o centenas de árboles, pueden obtenerse bien por la polinización artificial. Cuando se trabaje con múltiples factores de herencia (que es el caso más frecuente), la población F3 debe ser bastante abundante y contendrá lotes de cientos de descendencias y miles de plantitas individuales. Las probabilidades de éxito con ensayos F3 pequeños son escasas para garantizar la labor que requieren. En Populus, Picea y en muchas especies de Pinus puede polinizarse artificialmente en la escala que se desee con relativa facilidad. En cambio, para los que trabajen con géneros cuya polinización artificial sea dificultosa, como sucede en Abies, cuyas flores femeninas son casi inaccesibles, o con Castanea Juglans y Acer, en los que cada polinización produce poca semilla, es un buen consejo que lo hagan con descendencias de polinización libre que les permitirá establecer ensayos de tamaño adecuado.
La multiplicación vegetativa es comercialmente posible en muchas especies de Populus. Por lo tanto, en este género es preferible hacer ensayos de clones en las generaciones F2 y F3 a fin de seleccionar clones de buenas propiedades. En otros géneros como Pinus y Quercus, los ensayos de clones son caros y sin aplicación, pues comercialmente se emplearán familias de brinzales.
La selección de familias en generaciones segregantes debe jugar un papel importante en los programas de mejora del Castanea. La especie americana C. dentaba es de crecimiento bastante rápido, con porte y características de madera satisfactorias, pero no resiste al chancro (Endothia parasitica). Esta resistencia se la pueden dar los castaños asiáticos C. mollissima o C. crenata, ninguno de los cuales alcanza buenas dimensiones. Se ha obtenido un corto número de híbridos F1 más o menos intermedios entre los genitores. También se han obtenido unos pocos F2 y algunos retrocruzamiéntos. Varios de los segregados F2 son vigorosos y mucho más resistentes que el genitor C. dentaba. No es bastante, sin embargo, y parece necesario llegar a la producción de generaciones numerosas, F3 y F4, para tener un cultivar que sea comercialmente aceptable, desde el punto de vista maderero.
Una gran parte de la cría de Populus que se viene haciendo en Europa meridional tiende probablemente a producir segregados valiosos en generaciones posteriores. Sin embargo, no se ha iniciado todavía un programa formal de generaciones múltiples y la selección se hace entre árboles de generación desconocida.
En el Istituto de Genética Forestal de California, algunos híbridos F1 de Pinus han llegado ya a edad de fructificación. La mayoría son fértiles. Uno de los más prometedores es el cruce de P. jeffreyi X P. coulteri. Algunos de los F2 y de los retrocruces muestran los buenos caracteres de desarrollo del primero y la resistencia del segundo a los ataques del Cylindrocopturus eatoni. El grado de recombinación es suficiente para deducir que un grupo de familias F3 puede presentar algún cultivar aceptable comercialmente.
FIGURA 14. - Flores femeninas aisladas de Pinus sylvestris.
El cruce de razas diferentes dentro de una especie. ofrece las mismas posibilidades que el de variedades distintas de las plantas de cultivo o el de diferentes razas de animales. Los híbridos F1 pueden mostrar vigor híbrido, lo mismo que los obtenidos por cruces entre líneas consanguíneas de maíz. O pueden presentar una combinación de caracteres satisfactorios que sirvan de fundamento para un programa de selección en varias generaciones.
Dengler, iniciador de la mejora arbórea en Alemania, comenzó en 1926 una hibridación interracial cruzando pinos silvestres (Pinus sylvestris), oriundos de Alemania, Francia y Escocia. Creyó que algunas combinaciones mostraban vigor híbrido a la edad de 10 años, pero Scamoni más tarde, cuando los árboles tenían 20 años de edad, informaba que los híbridos eran intermedios entre sus padres.
En 1948, Johnsson y sus colaboradores en Ekebo (Suecia) hicieron una serie sistemática de cruzamientos entre temblones europeos (Populus tremula) originarios de tres diferentes latitudes de Suecia. A los 10 años todos los híbridos eran considerablemente superiores a la semisuma de las medias parentales y una de las combinaciones (meridional x central) desarrolló más que la parental de crecimiento más rápido.
Otros investigadores suecos, especialmente Nilsson y Langlet han conseguido resultados prometedores de la hibridación interracial en Pinus sylvestris y Picea abies vistos durante los viajes de estudio que precedieron la consulta mundial. Siguiendo sus pasos es indudable que los investigadores de otras regiones aumentarán su interés por este tema.
El primer requisito para una labor productiva de hibridación es disponer de buenas colecciones de material de mejora. Si lo que interesa es determinar las aptitudes para el cruce, se necesitarán varios individuos capaces de florecer de varias razas o especies. Si interesa la multiplicación en cantidad de un híbrido concreto destacado, se necesitarán muchos ejemplares seleccionados de cada una de las especies parentales (Figuras 11 y 12). No bastará disponer sólo de uno o dos ejemplares de cada tipo parental. Podrían no fructificar o no producir suficiente semilla sin hibridar para emplearla como testigo.
En los climas templados, antes de comprometerse en serio en un programa importante de hibridación, debiera dedicársele algún tiempo en las primaveras durante un período de cinco años. Ese tiempo habría que emplearlo en embolsar, polinizar, desembolsar y observar el desarrollo de las flores (Figuras 13 y 14). Las cosechas de semilla suelen ser tan variables que el mismo cruzamiento requiere ser repetido varias veces para poder establecer una pauta media que sea de confianza.
Un hibridador necesita un vivero de experimentación pequeño pero bueno y varios sitios de ensayo. Corrientemente, se dispone de pocas semillas de las combinaciones hechas por primera vez. Esto quiere decir que los ensayos han de hacerse con mucha más precisión que la corriente en las investigaciones ordinarias con plantaciones. Por eso, tanto el vivero como los campos de experiencia deben estar equipados para dar a los ensayos cuidados especiales.
Si se tiene interés en desarrollar cultivares de las dos F1 y F3 será preciso llevar a cabo por duplicado las plantaciones de ensayo. El diseño empleado para determinar el desarrollo comparativo de las F1 con sus ascendientes no debe ser el mismo que para conseguir una fructificación rápida. Como tampoco servirá necesariamente el mismo diseño para probar el vigor híbrido que para obtener híbridos F2 y F3 por polinización natural libre.
DUFFIELD, J. W. 1952. Relationships and species hybridization in the genus Pinus. Z. Forstgenet., 1: 93-100.
LERNER. I. M. 1958. The genetic basis of selection. New York, Wiley. 298 p.
MULLER, C. H. 1952. Ecological control of hybridization in Quercus: a factor in the mechanism of evolution. Evolution, 6: 147-161.
PALMER, E. J. 1948. Hybrid oaks of North America. J. Arnold Arbor., 29: 1-48.
PIATNISKY, S. S. 1960. Evolving new form of oak by hybridization. In Questions of forestry and forest management. Proc. 5th World For. Congr., Seattle, U.S.A., p. 231-244.
PRYOR, L. D. 1957. Selecting and breeding for cold resistance in Eucalyptus. Silvae Genet., 6: 98-109.
SCHREINER, E. J. 1969. Production of poplar timber in Europe and its significance and application in the United States. U.S. Dept. Agric. Handbook 150. 124 p.
WRIGHT, J. M. 1955. Species crossability in spruce in relation to distribution and taxonomy. For. Sci., 1: 319-349.
WRIGHT, J. W. Mejoramiento genético de los árboles forestales. Roma, FAO: Estudios de silvicultura y productos forestales N° 16, 436p.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}