![]()
![]()
![]()
A. Criterios para valoración selección de arboles plus
B. Algunos métodos de injertado
C. Anatomía de la zona de unión en injertos de pino silvestre y abeto rojo
D. Lista de borradores de capítulos y contribuciones
E. Miembros de las mesas de la consulta y otros participantes
En Suecia la selección de árboles plus se basa en la apreciación de caracteres fenotípicos, concretamente en el grado de crecimiento, porte, tipo de ramificación y calidad de la madera. Cuando se trata de árboles que viven a grandes altitudes, también se consideran la aptitud para la floración y la calidad de la semilla.
Arbol plus es aquél claramente superior a determinados árboles de las proximidades y a la media del rodal. Los árboles empleados para la comparación con el árbol plus son los cuatro mayores pertenecientes a la misma especie situados en las proximidades inmediatas, esto es, a menos de 25 a 50 metros (23 a 46 yardas) del árbol plus. Dichos pies deberán ser todos dominantes.
Criterios para seleccionar árboles plus de Pinus sylvestris y Picea abies
El árbol plus y los árboles a que se compara deben ser de la misma edad, medida a la altura del pecho. La diferencia de edad no ha de exceder normalmente de 10 años. Sin embargo, cuando la diferencia entre el árbol plus y los cuatro vecinos dominantes exceda de 10 años, las dimensiones de aquél deberán ser marcadamente superiores a lo que sea normal para las condiciones de edad y estación correspondientes.
Las condiciones de estación, tales como la fertilidad y humedad del suelo, la pendiente y espesura del rodal deben ser en apariencia los mismos para todos los árboles.
El árbol plus debe ser un árbol sano, libre de ataques por insectos o enfermedades; debe ser considerablemente más alto y poseer un diámetro claramente mayor que los árboles de comparación. Sus troncos han de ser de buena calidad con ramificación horizontal o ligeramente hacia arriba.
Para clasificar los árboles plus se emplea un sistema de puntuación en el que se considera el volumen y la calidad. El volumen se mide sin corteza. El volumen de los árboles de comparación se ajusta a la fase de desarrollo correspondiente a la edad del árbol plus. La puntuación por volumen se saca del cociente entre el volumen del árbol plus y el volumen medio de los árboles comparativos. Cada décima parte en exceso del valor 1,0 recibe un punto por volumen.
La puntuación por calidad se basa en la apreciación de la ramosidad, ángulo de ramificación y densidad de la madera. También se tiene en cuenta el número de ramas por verticilo. En las los Cuadros A1 y A2 se indica con claridad cómo se determinan los puntos de calidad.
La puntuación total del árbol plus es la suma de la puntuación por volumen y por calidad. De estas cifras
Publicados por el Comité de Coordinación para la Mejora C Genética Forestal, de acuerdo con una proposición de Börge Häggström, Servicio Forestal del Estado, Suecia.
CUADRO A1. - CLASES EMPLEADAS PARA LA DENSIDAD BÁSICA RELATIVA DE LA MADERA¹
|
Clases de densidad |
Puntuación |
|
|
|
Porcentaje |
|
|
Extremadamente alta |
115,5 |
+ 4,0 |
|
Muy alta |
107,5-111,4 |
+ 2,5 |
|
Alta |
103,5-107,4 |
+ 1,0 |
|
Media |
96,5-103,4 |
0,0 |
|
Baja |
92,5- 96,4 |
- 1,0 |
|
Muy baja |
88,5- 92,4 |
- 2,4 |
|
Extremadamente baja. |
88,4 |
- 4,0 |
¹ La densidad básica relativa expresa la relación entre la densidad básica de la madera observada y la calculada.
CUADRO A2. - CLASES SEGÚN EL GROSOR Y ÁNGULO DE LAS RAMAS
|
Grosor |
Angulo¹ | ||
|
Clase |
Puntuación |
Clase |
Puntuación |
|
Ramas-muy gruesas |
- 4 |
50° |
- 2 |
|
Ramas gruesas |
- 2,5 |
50 a 60° |
- 1 |
|
Ramas moderadamente gruesas |
- 1 |
60 a 70° |
0 |
|
Ramas normales |
0 |
70 a 80° |
+ 1 |
|
Ramas moderadamente delgadas |
+ 1 |
80 a 90° |
+ 2 |
|
Ramas delgadas |
+ 2,5 |
|
|
|
Ramas muy delgadas |
+ 4 |
|
|
¹Sólo se emplea para el pino silvestre.NOTA: La puntuación por calidad se reduce en 1 ó 2 puntos si el número de ramas por piso es anormalmente grande.
CUADRO A3. - CONVERSIÓN DE LA PUNTUACIÓN TOTAL EN UNA CLASIFICACIÓN DE ÁRBOLES PLUS
|
Puntuación total |
Clasificación |
|
- 7,5 a - 1,5 |
Bc¹ |
|
o a + 6 |
B |
|
7,5 a 13,5 |
Ba |
|
15 a 21 |
AB |
|
22,5 a 28,5 |
a |
|
> 30 |
A |
¹ Desaprobada.NOTA: En el caso del abeto rojo la puntuación más alta posible es 8, ya que no se puntúa el ángulo de las ramas. se pasa a una escala de clases: Be (desaprobada), B, Ba, AB, a y A de acuerdo con Cuadro A3.
El árbol plus y los de comparación son revisados por el jefe científico del Comité de Coordinación para la Mejora Genética Forestal. Durante esta inspección se valoran variaciones en calidad de estación, densidad del rodal, influencia de árboles próximos y sobre los cuidados culturales dados a la masa en el pasado. Este segundo examen puede dar lugar a cambios en la primitiva clasificación.
En el Cuadro A4 se muestra la influencia que tienen las puntuaciones por volumen y calidad sobre la puntuación total.
Esta estimación preliminar de los árboles plus peca desde luego de superficial y es poco satisfactoria. Interesa, por tanto, definir criterios más lógicos y seguros, basados en resultados de las pruebas de descendencia. Estos criterios para seleccionar los árboles plus y para la selección en masa deberán tener en cuenta la heredabilidad de diferentes caracteres y las correlaciones genéticas y fenotípicas entre ellos. También habrá de considerarse el valor económico relativo de las diferentes propiedades.
CUADRO A4. - RELACIÓN ENTRE LAS PUNCTUACIONES POR VOLUMEN Y CALIDAD EMPLEADAS EN LA SELECCIÓN DE ÁRBOLES PLUS DE PINO SILVESTRE Y EL ABETO BOJO
|
Puntuación volumétrica |
20 |
20 |
22,5 |
25 |
27,5 |
30 |
|
|
15 |
15 |
17,5 |
20 |
22,5 |
25 |
|
|
10 |
10 |
12,5 |
15 |
17,5 |
20 |
|
|
5 |
5 |
7,5 |
10 |
12,5 |
15 |
|
|
0 |
0 |
2,5 |
5 |
7,5 |
10 |
|
Puntuación cualitativa |
0 |
2,5 |
5 |
7,5 |
10 | |
INGEGERD DORMLING
Epoca de injertado
En Suecia las coníferas se injertan en invernadero en febrero y marzo o al aire libre en marzo o junio. Se emplean patrones de tres a cuatro savias (planta repicada 2 + 1 ó 2 + 2). Los destinados a injertado de invernadero colócanse en tiestos la primavera o verano anterior a la operación de injertado y se meten en el invernadero a principios de enero. Se ha visto que el mejor momento para injertar es cuando las yemas de los patrones empiezan a reventar. Como púas se emplean ramitas del último año en completo letargo.
El material procedente del norte de Suecia tiene con frecuencia brotes anuales demasiado cortos y delgados por lo que hay que emplear para púas remitas de dos a tres años. Las frondosas pueden injertarse en invernadero en marzo o abril o al aire libre en abril. En ambos casos no debe haber comenzado el movimiento de la savia.
Con frecuencia es necesario almacenar las púas por cierto tiempo, particularmente para el injertado al aire libre, cuando las ramitas se recogen en letargo durante el invierno. El almacenamiento en bolsas de plástico perforadas, a temperatura de 0°C (32°F) ha sido satisfactorio. Pueden emplearse temperaturas más bajas si las púas se recogen en tiempo frío. Sin embargo, no es aconsejable la recogida cuando la temperatura es inferior a - 5°C (23°F).
Métodos de injertado
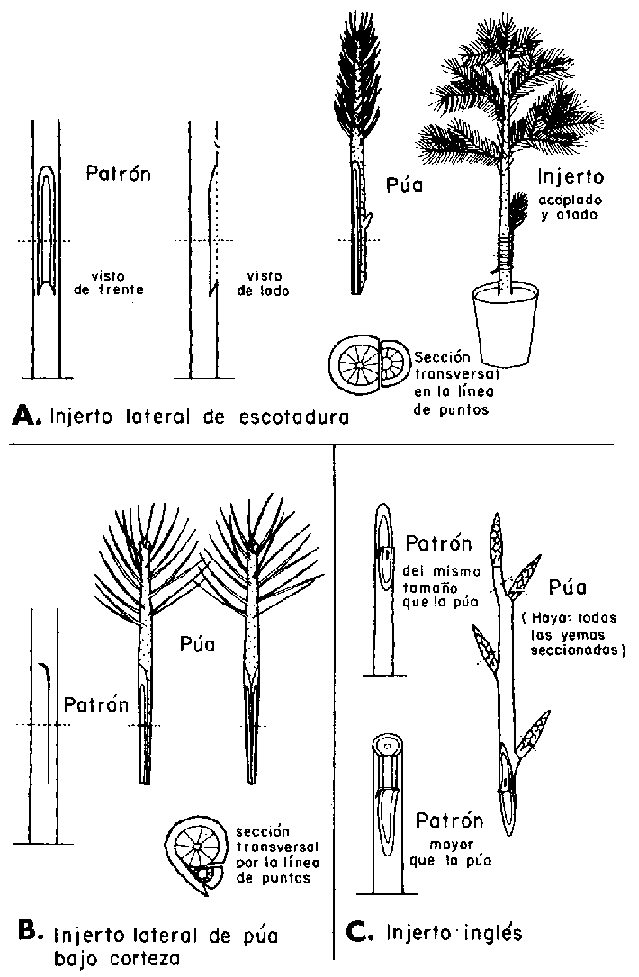
Los métodos de injertado más usados en Suecia se muestran en la Figura 28 A, B y C. El injerto lateral de escotadura (vencer side graft) es el método más satisfactorio para coníferas, en especial para el abeto rojo (Picea abies). La púa debe tener de 5 a 8 cm. (2 a 3 pulgadas) de largo. Se quitan todas las acículas de la parte inferior y la púa se prepara dándole dos cortes, uno de 3 a 4,5 cm. (1,2 a 1,7 pulgadas) de largo y el otro corte en el lado opuesto, de manera que la base queda en forma de cuña. El patrón se limpia en la región del injerto y por un corte superficial se extrae una tira de corteza con trazas de madera. En la base se da un corte hacia abajo y adentro que desprenda la tira del anterior corte. Los cortes en el patrón y púa deben corresponderse de forma tal que permita acoplar perfectamente las zonas del cambium.
El injerto lateral de púa bajo corteza (side-slit graft) se aplica principalmente al pino silvestre (Pinus sylvestris) cuando es necesario injertar púas delgadas sobre patrones robustos. Se da a la púa dos cortes iguales de 3 a 4 cm. (1,2 a 1,5 pulgadas) de largo para formar una cuña. La incisión en el patrón se hace tangencialmente al cambium, se levanta la corteza e inserta la púa de modo que las zonas cámbiales se ajusten bien en la cara de incisión.
INGEGERD DORMLING es Asistente en el Departamento de Genética Forestal del Real Colegio de Montes, Estocolmo. Véase también el Apéndice C.
FIGURA 28. - Métodos de injertado de árboles.
El injerto inglés (whip-and-tongue). se practica especialmente con las frondosas, aunque también ha tenido algún éxito con el pino silvestre y el alerce europeo (Larix decidua). La última especie puede ser injertada por cualquiera de los métodos aquí descritos. El extremo de la base se prepara por un corte sesgado que sea aproximadamente seis veces más largo que ancho. En la superficie de este corte se hace una lengüeta con la punta para abajo. El corte en el patrón varía según el tamaño del patrón y la púa. Si tienen igual diámetro, se da un corte sesgado en el patrón de la misma longitud y se hace una lengüeta con la punta hacia arriba para acoplarla a la de la púa. Si el patrón es considerablemente mayor que la púa, el corte debe ser comparativamente superficial. Es importante dejar en contacto las regiones cámbiales de púa y patrón en las lengüetas y en la parte inferior de la superficie de unión. Se quitan siempre las yemas terminales de las púas y, si se trata de Fagus sylvatica, el resto de las yemas se recortan a un tercio de su longitud. Si no se hace así, la púa desarrolla una gran masa de follaje antes de que se establezca la unión del injerto y morirá por desecación.
Los injertos se atan con cinta de plástico o rafia y embadurnan con mástic. En el sur de Suecia (Ekebo) no se emplea mástic en el injertado de coníferas, pero en Sundmo, Brunsberg y Bogesund en el norte y centro de Suecia se ha visto era necesario para conseguir buenos resultados.
Cuando el injerto ha prendido, los patrones se van recortando gradualmente. Esta operación debe hacerse con gran cuidado y no muy de prisa. En injertos hechos en Ekebo con pino silvestre y alerce europeo se terminó el recorte de los patrones en el primer verano con buenos resultados. Fuera de estos casos ha sido aconsejable esperar hasta el próximo verano.
Otros métodos de injertado
Existen otras muchas técnicas que pueden ser empleadas con pleno éxito. En Finlandia y Estados Unidos se ha practicado el injerto de verano en coníferas, con material semisuculento, recogiéndose las púas inmediatamente antes de la operación.
Para algunas especies de coníferas (especialmente Larix) se ha ensayado con éxito el llamado injerto a mano (hand grafting), es decir, empleando el método usual, pero sobre patrones de dos savias extraídos directamente del semillero y trasplantados bajo sombra en vivero inmediatamente después de la operación.
A voces se recomienda el injerto terminal sobre plantas relativamente grandes de coníferas cultivadas en huertos para semilla, pero es aconsejable sólo con especies de fácil injerto, tales como Larix y posiblemente Pinus. En Dinamarca se emplea con Pseudotsuga taxifolia. En Suecia se ha practicado en pequeña escala el injerto terminal sobre patrones enmacetados de Picea abies en invernadero con buenos resultados. La técnica de injertado comprende la decapitación del patrón y cortes en patrón y púa similares al injerto lateral de escotadura; este método se denomina injerto terminal de escotadura (vencer crown or top veneer graft).
No cabe hacer una generalización universal sobre la mejor época para injertar. Varía con el clima, método y especie. Puede obtenerse alguna información de las referencias citadas abajo.
Para injertar al aire libre son útiles los invernaderos transportables de polietileno, como los empleados en Finlandia. En Canadá y Estados Unidos han sido utilizadas con éxito bolsas de polietileno, cubriendo individualmente los injertos.
BRAUN, H. J. 1962. Zur Frage günstiger Pfropftermine bei Holzpflanzen-Pfropfungen. Allgem. Forst. Jagdz., 133: 266-259.
DORMLING, I. 1962. Ympningsmetoder för tall och gran [Grafting methods for Scots pine and Norway spruce]. Medd. Statens Skogsforskninginst., Stockh. 5 1 (2): 23.
FOWLER, D. P. 1959. A summer field grafting technique for pine. For. Chron., 35: 30-35.
GARNER, R. J. 1959. The grafters handbook. London. 260 p.
GUINAUDEAU, J. 1961. Note sur les precédés de greffage du pin maritime. Rev. forest. franç., 13: 152-160.
HOLST, M. J. 1956. Phenology of rootstocks and grafts in a timing experiment with autumn and winter grafting of Norway and white spruce. Can. Dep. Northern Affairs Nat. Resources, Forestry Branch, Forest Res. Div. Tech. Note 48. 17 p.
KEILLANDER, C. L. 1946. Om bärrträdsforädling och bärrträdsympning [On the breeding and grafting of conifers]. Svensk Papperstid., 49: 556-563, 586-593.
LESKINEN, U. 1960. Kokemuksia männyn vartamisesta Suomessa [Experiences in the grafting of pine in Finland]. Eripainos Metsätaloudellisesta Aihakauslehdestä, 11 6 p.
NAESS-SCHMIDT, K. & SØEGAARD, B. 1960. Podehöjdens inflydelse på podekvistens vaekstrytme og form [The influence of the grafting height on the development of the scion]. Det Forst. Forsogsvaesen i Danmark, 26: 315-324.
MERGEN, F. 1955. Grafting slash pine in the field and in the greenhouse. J. For., 53: 836-842.
MERGEN, F. & ROSSOL, E. 1954. How to root and graft slash pine. Southeast. Forest. Expt. Sta., Asheville, Sta. Paper 46. 22 p.
ORR-EWING, A. L. & PRIDEAUX, D. C. 1959. Grafting methods for the Douglas fir. For. Chron., 35: 192202.
SCHRÖCK, O. & HOFFMANN, K. 1957. Sommerpfropfung im Freiland für die Anlage von Samenplantagen. Züchter, 27: 246-250.
SCHRÖCK, O., KOOTZ, F. W. & HOFFMANN, K. 1954. Forstliche Samenplantagen. Ein Beitrag zu ihrer Anlage. Berlin. 79 p.
WEBB, C. D. 1961. Field grafting Loblolly pine. N. Carolina State Coll., School Forestry, Raleigh, Tech. Rept. 10. 33 p.
ZAK, B. 1955 The grafting of short-leaf and other pine species. Southeast. Forest. Expt. Sta., Paper 59. 13p.
INGEGERD DORMLING
Preparación del material
Se ha investigado la anatomía de las uniones de injertos hechos en invernadero por el método lateral de escotadura y lateral de púa bajo corteza. Las uniones fueron cortadas en trozos de 5 milímetros (0,2 pulgadas) de largo, fijadas en «Craf» embebido en parafina y cortando sendas secciones transversal y longitudinal de 15m de grosor y teñidas con safranina y verde rápido.
En la Figura 29 se presentan los detalles anatómicos observados en sección transversal, tangencial y radial. La estructura del tallo es similar en el pino silvestre y abeto rojo, pero existen algunas diferencias notables en la formación del callo y la unión del injerto. El parenquima del cambium para adentro (médula, radios medulares, canales resiníferos) está formado en Pinus sylvestris de células con paredes delgadas, mientras que en Picea abies la mayoría de estas células tienen paredes lignificadas. Los radios medulares multiseriados son más numerosos en pino silvestre que en abeto rojo y faltan en los brotes de un año del último. La parte suberificada del peridermo en brotes jóvenes es mayor en el abeto rojo que en el pino silvestre y los rastros foliares son más numerosos en el abeto que en el pino.
INGEGERD DORMLING es Asistente en el Departamento de Genética Forestal del Real Colegio de Montes, Estocolmo.
Formación del tejido calloso
Al injertar se produce inmediatamente una excreción de resina en la superficie de la herida, manando de los canales resiníferos cortados. Nada indica que la resina pueda constituir una barrera entre los componentes del injerto; por el contrario, sirve para soldar. En las superficies del corte, en los sitios donde está expuesto el parenquima, se forman capas de contacto consistentes en células dañadas directa o indirectamente. La primera reacción perceptible de las células vivas es el alargamiento de las células adyacentes a las superficies de la herida. Las células epiteliales de los canales resiníferos verticales de la corteza primaria seccionados reaccionan particularmente pronto. La división celular aparece cerca de las superficies de la herida, tanto en el patrón como en la púa, tres o cuatro días después del injerto. Con frecuencia la púa presenta las primeras divisiones y mayor actividad en los días que siguen inmediatamente al injertado.
Las células que rodean los rastros de hojas y ramas y las células de los radios que emergen de los intersticios foliares son especialmente activas en la formación del callo. En conjunto el patrón produce un volumen mayor de callo que la púa, pero los rastros foliares adyacentes a las superficies de corte de las púas originan una formación local de callo mayor. Los radios medulares que han sido cortados hacia afuera en el floema suelen formar más callo que los seccionados cerca o dentro del cambium. El límite entre el floema y la corteza primaria es una zona de crecimiento muy activa.
La región cambial desempeña una función subordinada como productora de callo en el injerto de escotadura ordinario. Sin embargo, en los injertos laterales de púa bajo corteza, ésta ha sido despegada del leño en la región cambial y las células originan en el pino silvestre una vigorosa formación de callo. Este proceso se inicia por las células de los radios medulares, pero pronto participa la mayor parte de las células cámbiales si no han sido dañadas durante el injertado.
La formación de callo por el parenquima que queda al interior del cambium sólo se da en el pino silvestre, pudiendo a veces ser considerable. La médula de la púa es especialmente activa al ser seccionada.
En el Cuadro C1 se compara la predisposición de varios tejidos a producir callo en ambas especies. Una gran actividad en uno de los componentes del injerto parece
CUADRO C1. - INTENSIDAD DE DIVISIÓN CELULAR EN DIFERENTES TEJIDOS DE INJERTOS DE PINO SILVESTRE Y ABETO BOJO
|
Tejido |
Pino silvestre |
Abeto rojo |
|
Peridermis |
- |
- |
|
Corteza primaria, tejido fundamental |
+++ (1) |
+++ (1) |
|
Corteza primaria, canales resiníferos |
++++ |
++++ |
|
Floema, radios medulares |
+ + + + (1) |
+ + + + (1) |
|
Floema, parenquima vertical |
+ + (1) |
+ + (1) |
|
Floema, células cribosos completamente diferenciadas |
- |
- |
|
Región cambial, radios |
+ + + (1) |
+ + + (1) |
|
Región cambial, otras células no diferenciadas o incompletamente diferenciadas |
+ + |
+ + |
|
Xilema, radios medulares |
+ |
- (+) |
|
Xilema, canales resiníferos |
+ |
- |
|
Xilema, traqueidas completa mente diferenciadas |
- |
- |
|
Médula, intersticios de hojas y ramas |
+ + |
- |
(1) = Los tejidos influenciados por rastros foliares y de ramas muestran mayor actividad que otros++++ = Intensidad de división celular muy grande
+++ = Intensidad de división celular grande
++ = Intensidad de división celular variable: en médula e intersticios foliares y de ramas del pino silvestre las divisiones acaecen en fase temprana, en otros casos no hasta que han empezado a dividirse otros tejidos próximos
+ = Puede haber divisiones en cantidad variable, pero a menudo no se dan
(+) = En injertos tardíos, las células de los radios del xilema del año en curso pueden dividirse
- = No hay divisiones celulares estimular en los tejidos adyacentes del otro componente un incremento de actividad antes de que se haya establecido una conexión directa entre los tejidos.
Soldadura del injerto
Han sido confirmadas soldaduras entre células parenquimatosas ocho o diez días después del injerto. Quince días después de éste existen uniones de parenquima en todos los injertos de invernadero que ofrecen perspectivas de ulterior desarrollo. Sólo son capaces de soldarse células de nueva formación, posteriores al injerto. En los injertos de escotadura, las primeras uniones tienen lugar entre células derivadas de tejidos exteriores a la región cambial. En los injertos de púa bajo corteza con incisión radial en la corteza del patrón (Figura 29E), las primeras uniones se establecen entre el callo de la región cambial del labio de corteza y el callo de la corteza primaria, radios medulares liberianos y médula de la púa. En injertos de púa bajo corteza con incisión tangencial sobre el patrón (Figura 29F) las soldaduras se dan también en la cara de incisión en forma muy similar al injerto de escotadura. Pueden verificarse soldaduras entre células de nueva formación independientemente de la clase de tejido original.
La soldadura entre cámbiumes corticales tiene lugar de 15 a 20 días después del injertado. Los felógenos viejos más exteriores que envuelven los troncos no participan en la formación de los nuevos, teniendo lugar sólo gradualmente uniones entre felógeno nuevo y viejo.
Ha sido confirmada la soldadura de tejidos vasculares a las tres semanas aproximadamente en injertos bien acoplados. Cuando el acople no es tan perfecto la soldadura puede requerir cinco o seis semanas. En los injertos bien acoplados es corriente que las divisiones en la región cambial puedan servir desde el principio para la soldadura de los tejidos vasculares. En tal caso sólo se forman en corto número células pequeñas irregulares. Cuando las regiones cámbiales queden algo lejos, se extienden a través de los tejidos parenquimatosos intermedios en dirección recíproca, por inducción de una célula a la siguiente.
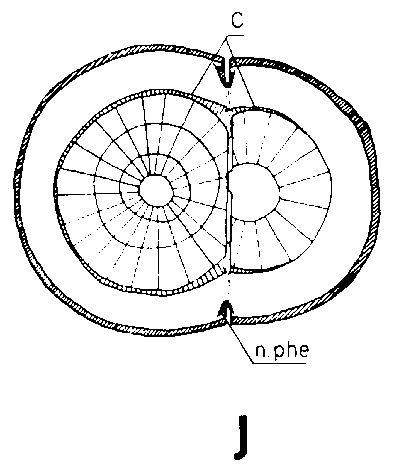
Debido a que el cambium del patrón ya se encuentra en estado activo al tiempo de injertar y a que está mejor provisto que el de la púa de agua y nutrientes, resulta que es capaz de poner varias capas nuevas de traqueidas hasta el momento en que es posible la soldadura. Esto da lugar a que el cambium sea impelido hacia afuera, separándose del cambium de la púa que haya sido colocado enfrente, pero emparejándose si hubiese sido colocado hacia afuera (Figura 29G, H, I y J).
Las primeras soldaduras de los tejidos vasculares en el injerto lateral de escotadura se dan a menudo entre la muesca del patrón y los tejidos situados en la superficie del corte pequeño de la púa. Parece que los rastros foliares desempeñan un importante papel en el establecimiento de conexiones vasculares, especialmente en el abeto rojo donde se dan en gran número. Las uniones cámbiales en la arista más interna del injerto lateral de púa bajo corteza son complejas, dependiendo de la posición mutua de los tejidos de los componentes del injerto.
No es preciso que los espacios entre las superficies de ambos componentes del injerto se llenen con teijdo calloso. Por el contrario, con frecuencia queda vacío cuando los tejidos se sueldan rápidamente por la parte de afuera.
En el verano del año siguiente al injerto el patrón es desmochado dando un corte oblicuo inmediatamente por encima del extremo superior de contacto con la púa. La herida así causada se recubre de callosidad a partir de todos los lados del mismo modo que en una rama cortada.
Injerto lateral de escotadura
Hay dos formas de hacer la muesca del patrón en el injerto lateral de escotadura, como se indica en la Figura 29, C y D. De las dos es preferible el corte hacia abajo de la Figura 29, D. De la otra forma una tira de leño irá adherida todo a lo largo del labio y obstruirá una unión lisa entre el labio y la superficie del corte pequeño de la púa.
Una buena colocación de las regiones cámbiales en los injertos de escotadura es esencial para la soldadura rápida de los componentes del injerto. Esto no significa sin embargo, que las regiones cámbiales de la púa y patrón deban colocarse exactamente enfrente una de otra. Es mejor que el cambium de la púa quede a una corta distancia hacia afuera en relación con el del patrón para compensar el crecimiento de éste, más vigoroso (Figura 29, G, H, I, y J).
Se ha visto que es mucho más difícil ajustar bien los componentes del injerto en el abeto rojo que en el pino silvestre. Las dificultades en el injerto del abeto rojo dependen del hecho de ser frecuentemente los patrones considerablemente más gruesos que las púas. Se ha visto, sin embargo, que era posible un buen ajuste dando un corte superficial en el patrón, corte que apenas alcanza al leño y que permite acoplar la superficie del corte de la púa a la del patrón. En el pino silvestre en cambio, no es aconsejable hacer tal incisión superficial en el patrón pues da lugar a la formación de un vigoroso callo sobre la superficie entera de corte lo cual retrasa la unión de las capas de cambium y hasta puede provocar la expulsión de la púa. Esto no ocurre en el abeto rojo. Por supuesto los componentes del injerto deben ir atados fuertemente para evitar que el patrón tape la superficie del corte antes de que se establezca la conexión con la púa.
Comparaciones entre brotes de un año de pino silvestre y abeto rojo mostraron que la corteza de la última especie contiene una proporción mucho menor de tejidos vivos. Los brotes de abeto rojo se hallan rodeados de una gran capa de células muertas siendo muy delgada la corteza primaria entre las crestas que forman los rastros foliares. Siendo comparativamente limitados en extensión los tejidos capaces de proliferación en las púas de abeto rojo, es de gran importancia lograr un buen ajuste cambial en esta especie. La investigación ha demostrado claramente que son de preferir los patrones jóvenes y pequeños cuando se trate del injerto lateral de escotadura.
Injerto lateral de púa bajo corteza
La incisión en el patrón en los injertos bajo corteza se hace de dos formas: radial o tangencialmente (Figura 29 E y F). Cuando la incisión es radial la púa entera deberá colocarse por dentro de la cara de incisión(Figura 29E) quedando muy separados en este punto los bordes de los cámbiumes. Con incisión tangencial (Figura 29 F) existe la posibilidad de ajustar las capas cambiales de modo que se obtengan las mismas uniones que en el caso de injerto lateral de escotadura. De los injertos bajo corteza de abeto rojo practicados en una investigación sólo tuvieron éxito los de incisión tangencial en el patrón. La mayor parte de las soldaduras tuvieron lugar en la cara de incisión mientras que la corteza solapada falló mucho. Un corte en el patrón afectando a una tira de leño da una superficie de contacto para la púa aún mejor y menos espacio vacío en la esquina del ángulo.
FIGURA 29A
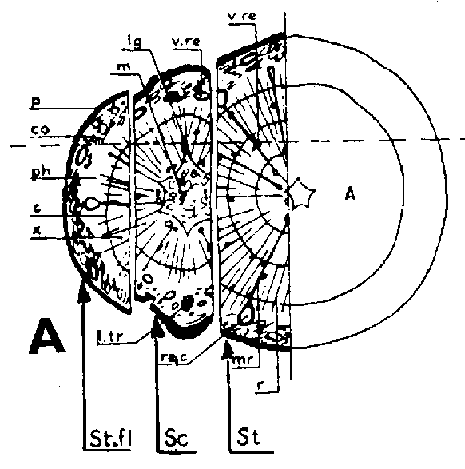
FIGURA 29B
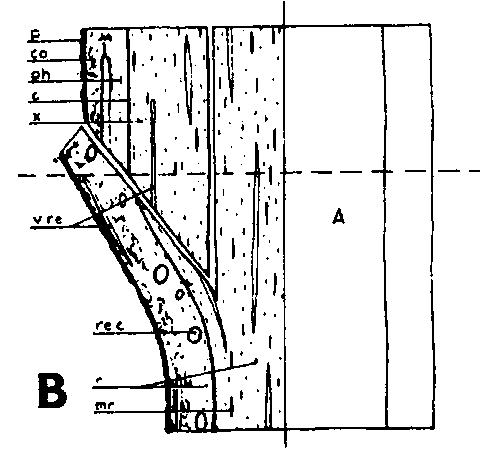
FIGURA 29C
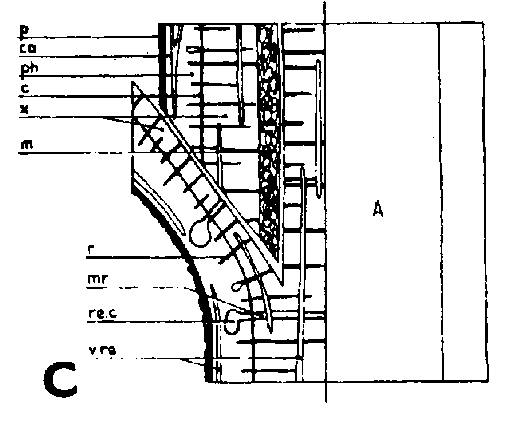
FIGURA 29D
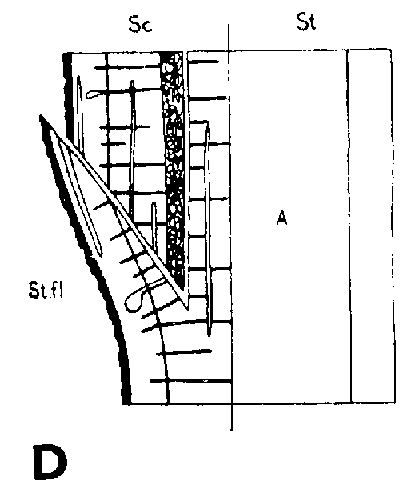
FIGURA 29E
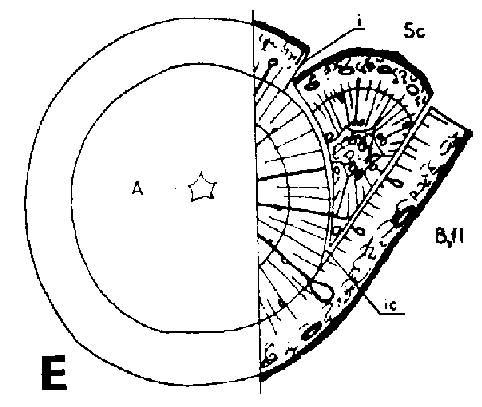
FIGURA 29F
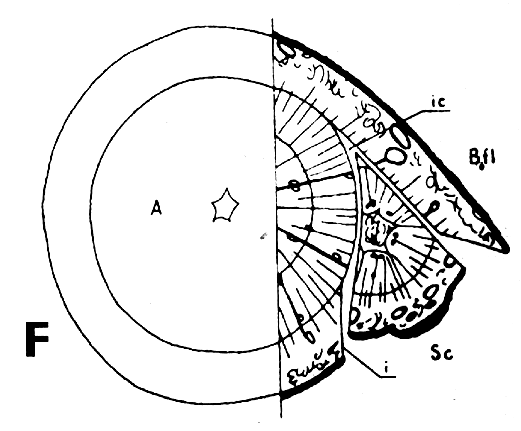
FIGURA 29G
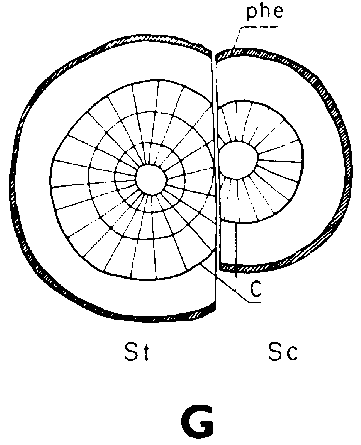
FIGURA 29H
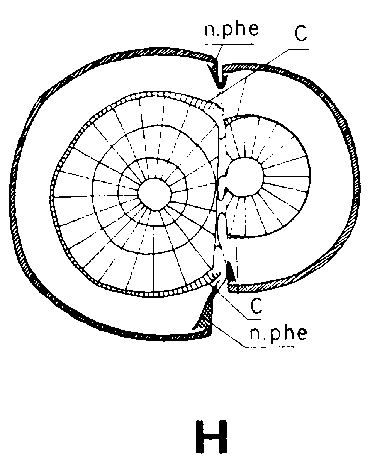
FIGURA 29I
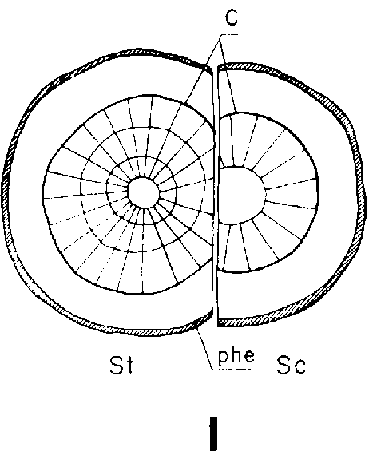
FIGURA 29J
Símbolos de la Figura 29. - Sc = púa, St = patrón, St. fl = muesca en el patrón (en injertos laterales de escotadura), B. fl = labio de corteza (en injertos laterales de púa bajo corteza), i = cara de incisión (en injertos laterales de púa bajo corteza), ic = arista más interior (en injertos laterales de púa bajo corteza), m = médula, x = xilema, c = cambium, fl = floema, co = corteza primaria, p = peridermis, phe = felógeno (cambium del corcho), n.phe = felógeno nuevo, r = radio medular, mr = radio modular multiseriado (radio conteniendo un canal resinífero), re. c = vesícula resinífera, v.re = canal resinífero vertical, l.tr = rastro foliar, lg = intersticio foliar.
FIGURA 29. - A y B. Secciones transversal y tangencial respectivamente de la parte superior de un injerto lateral de escotadura en pino silvestre. La línea a trazas señala dónde se tocan las dos secciones. A parte del patrón cortada antes de fijar las secciones. - C y D. Secciones radiales de la parte inferior de injertos laterales de escotadura con la muesca del patrón cortada hacia arriba y hacia abajo respectivamente. - E y F. Secciones transversales de injertos laterales de púa bajo corteza. Incisión radial y tangencial en la corteza del patrón. - G, H, I y J. Ilustraciones simplificadas del crecimiento cambial en los injertos laterales de escotadura. Secciones transversales: G y H. Patrón grande, púa pequeña: G. Emplazamiento corriente inmediatamente después del injerto. Las capas cambiales acopladas en un lado; el cambium de la púa por dentro del cambium del patrón en el otro lado. H. Después de tres semanas aproximadamente. De crecimiento más rápido, el cambium del patrón ha dejado atrás al de la púa en el lado superior. En el lado inferior desarrollo vigoroso de callo; los componentes del injerto son empujados aparte. Cámbiumes corticales unidos en el lado superior. - I. Patrón y púa de tamaño aproximadamente igual: I. Emplazamiento ideal. El cambium de la púa por fuera del cambium del patrón en ambos lados. J. Después de tres semanas aproximadamente. Cámbiumes listos para unirse. Cámbiumes corticales unidos.
De ambos métodos ensayados es claramente superior el injerto lateral de escotadura que es el recomendado para el abeto rojo. El injerto lateral bajo corteza puede ser aplicado al pino silvestre cuando se precisa injertar púas pequeñas sobre patrones gruesos. Las incisiones sobre éstos deberán ser tangenciales.
Injertos heteroplásticos
Los injertos heteroplásticos o interespecíficos se emplean en la mejora de árboles frutales para producir árboles pequeños de floración temprana y rica. El mismo efecto se persigue cuando se injertan árboles forestales para huertos de semillas.
En Suecia se han hecho algunas experiencias pero con resultados poco satisfactorios. El Pinus mago se empleó como patrón para el Pinus sylvestris. Estas especies se sueldan bien si la primera es de raza de buen crecimiento, pero entonces el injerto tiene los mismos caracteres que si fuese intraespecífico. Si el patrón es de raza de crecimiento lento la púa crecerá más aprisa que él. Se producirá un engrosamiento por encima de la unión que queda debilitada y puede romperse fácilmente con el viento. Excesivo crecimiento del patrón puede originar la expulsión de la púa. La experiencia ha enseñado que los mejores patrones para pino silvestre y abeto rojo son brinzales de la misma especie y de procedencia tan próxima a la de la púa como sea posible. De algunos árboles reproductores es muy difícil obtener injertos y a veces hay que contar con incompatibilidad entre los componentes de injerto de una misma especie.
En países donde se disponga de varias especies aumentan mucho las posibilidades de combinaciones acertadas de injertos interespecíficos. Mergen (1954 b), Pitcher (1960) y Ahlgren (1962) han hecho estudios con especies forestales.
La incompatibilidad entre el patrón y la púa ha sido objeto de numerosos estudios (Proebsting, 1926, 1928; Bradford y Sitton, 1929; McClintock, 1948; Herrero, 1951; Thiel, 1954; Stigter, 1956; Pitcher, 1960 y Mosse, 1962). Ocurre frecuentemente que las púas persistan uno o más años y entonces se mueran. Schönbach (1960) da cuenta de algunos injertos de Populus cuyas púas vivieron hasta siete años. En estos injertos las soldaduras del xilema son casi completas pero las del floema son poco satisfactorias. Intervienen capas de corcho en la región del injerto. La continuidad del cambium parece romperse al final de la estación de crecimiento. Puede establecerse una nueva soldadura en la primavera, pero a menudo no aparece. Algunas combinaciones de injerto han resultado compatibles cuando se dejaron hojas en el patrón (Stigter, 1956; Pitcher, 1960).
A menudo se ha discutido la influencia mutua entre patrón y púa, pero no se conoce plenamente su naturaleza (Brix, 1952; Thiel, 1954; Rogers y Beakbane, 1957). Sin embargo, es evidente que los caracteres hereditarios nunca pueden ser influidos por esta vía.
AHLGREN, C. E. 1962. Some factors influencing survival, growth, and flowering of intraspecific and interspecific pine grafts. J. For., 60: 785-789. (H) 1
BRADFORD, F. C. & SITTON, B. G. 1929. Defective graft union in the apple and the pear. Mich. Agric. Exp. Sta. Tech. Bull. 99. 106 p. (A, H).1
BRAUN, H. J. 1958. Die normalen Verwachaungavorgänge nach Pfropfung von Laubbäumen. I. Des Verfahren des seitlichen Einspitzens. Z. Botan., 46: 309338. (A) 1
BRAUN, H. J. 1959. Die normalen Verwaohaungavorgänge nac Pfropfung von Laubbäumen. II. Die Verfahren des seitlichen Anplattens und der Kopulation. Z. Botan., 47: 145-166. (A) 1
BRAUN, H. J. 1960a. Die normalen Verwachsungevorgänge nach Pfropfung von Laubbäumen. III. Das seitliche T-Schnitt-Verfahren, die Geissfuss- und Keilpfropfung, sowie das Ablaktieren. Z. Botan., 48: 58-65. (A) 1
BRAUN, H. J. 1960b. Neuere Erkenntnisse über die Vorgänge beim Pfropfen von Bäumen. Mitt. Deut. Dendrol. Ges., 61: 32-44. (A) 1
BRIX, KÄTHE. 1952 .Untersuchungen über den Einfluss der Pfropfung auf Reis und Unterlage und die Möglichkeit einer Übertragung eventueller Veränderungen auf die Nachkommen. Z. Pflanzenzucht., 31: 261-288. (H) 1
DORMLING, I. 1964. Anatomical and histological examinations of the union of scion and stock in grafts of Scots pine (Pinus silvestris L.) and Norway spruce (Picea abies [L] Karst.). (To be published in Sludia Forestalia Soecica) (A) 1
EVANS, G., WATSON, D. P. & DAVIDSON, H. 1961. Initial evolution of grafting some species of the Rosaceae. Proc. Am. Soc. Hort. Sci., 78: 580-585. (A, H) 1
HERRERO, J. 1951. Studies of compatible and incompatible graft combinations. J. Hort. Sci., 26: 212-237. (H) 1
JULIANO, J. B. 1941. Callus development in graft union. Philippine J. Sci., 75: 245-251. (A)¹
LAUNAY, J. 1961 .Phénomènes d'histogenèse produits lors du greffage du pin maritime. Rec. Trav. Lab. Vég. Fac. Sc., Bordeaux 2. (A) 1
MCCLINTOCK, J. A. 1948. A study of uncongeniality between peaches as scions and the Marianna plum as a stock. J. Agric. Res., 77: 253-260. (H) 1
MERGEN, F. 1954a. Anatomical study of slash pine graft unions. Quart. J. Florida Acad. Sci., 17: 237-245. (A) 1
MERGEN, F. 1954b. Heteroplastic micrografting of Slash pine. Southeast. Forest Expt. Sta., Sta. Paper 47, 17 p. (H) 1
PITCHER, J. A. 1960. Heteroplastic grafting in the genera Acer, Fraxinus, Picea and Abies. Proc. 7th Northeast. For. Tree Impr. Conf., p. 52-57. (H) 1
PROEBSTING, E. L. 1926. Structural weakness in interspecific grafts of Pyrus. Bot. Gaz., 82: 336. (A, H) 1
PROEBSTING, E. L. 1928. Further observations on structural defects of the graft union. Bot. Gaz., 84: 82-92. (A, H) 1
ROGERS, W. S. & BEAKBANE, A. B. 1957. Stock and scion relations. Ann. Rev. Plant Physiology., 8: 217-236. (H)¹
SASS, J. E. 1932. Formation of callus knots on apple grafts as related to the histology of the graft union. Bot. Gaz., 94: 364-380. (A, H) 1
SCHÖNBACH, H. 1960. Beiträge zur Pappelforsohung nr V. Beobachtungen an heteroplastischen Pfropfungen innerhalb der Gattung Populus. Wiss. Abh. Landw.-wiss, Berlin, 47: 27-41. (H)¹
SHARPLES, A. & GUNNERY, H. 1933. Callus formation in Hibiscus Rosa-sinensis L. and Hevea brasiliensis Mull. Arg. Ann. Bot., 47: 827-840. (A) 1
STIGTER, H. C. M. DE. 1956. Studies on the nature of the incompatibility in a cucurbitaceous graft. Mededel. Landbouwhogeschool, Wageningen, 56 (8). (H) 1
THIEL, K. 1954. Untersuchungen zur Frage der Unverträglichkeit bei Birnendelsorten auf Quitte A. (Cydonia EMA). Gartenbauwiss., 1: 127-159. (H)1
1 (A) = anatomía; (H) = injerto heteroplástico.
Observations on the bisexual RUGGERI, CECILIA behavior of a population of (Italia) Populus tremula L.
Borradores de capítulos
Section
|
1. Algunos principio sobre genética y citilogía arbórea |
GUSTAFSSON, ÅKE (Suecia) |
|
2a Genética de poblaciones como base de selección |
STERN, KLAUS (Alemania, Fed. Rep.) |
|
2b Hibridación entre especies y razas |
WRIGHT. JONATHAN W. (Estadis Unidos) |
|
3 Investigación de procedencias |
CALLAHAM, R. Z. (Estadis Unidos) |
|
4 Genética y mejora de las especies exóticas forestales |
MORANDINI, RICCARDO (Italia) |
|
5 La fisiología del árbol en relación con la genética y sus aplicaciones |
WAREING, P. F. (Reino Unido) |
|
6a Mejora genética de la resistencia a las enfermedades en las especies forestales |
BJÖRKMAN, E. (Suecia) |
|
6b Mejora genética de la resistencia a los ataques de insectos en las especies forestales |
SOEGAARD, BENT (Dinamarca) |
|
7 Mejora genética de las propiedades de la madera de especies forestales |
ZOBEL, BRUCE J. (Estados Unidos) |
|
8 Producción y certificación de semilas |
MATTHEWS, JOHN D. (Reino Unido) |
|
9 La genética en la práctica forestal de Suecia |
PLYM FORSHELL, W. (Suecia) |
Contribuciones
SECCIÓN 1. GENÉTICA ARBÓREA Y CITOLOGÍA
|
Evaluation of spontaneous, chemical and radiation-induced mutations in Pinaceae |
MERGEN, F (Estados Unidos) |
|
Cytogenetical evolution in conifers |
KHOSHOO, T. N. (India) |
|
Some characteristics of autotetraploids of tree species induced by colchicine treatment |
KYUN, SIN-KYU & KIM, CHUNG-SUK (Corea) |
|
Etude d'un peuplier tetraploide obtenu artificiellement et de sa descendance |
SEKAVIN, MICHELE (Italia) |
|
Genetics of flavonoids in Prunus wood |
HASEGAWA, MASAO & SHIRATO, TERUO (Japón) |
|
Sensitivity of pine seed to neutron, gamma and X-ray irradiation |
YIM, KYONG BIN - (Corea) |
|
Inheritance of needles discoloration of sugi (Cryptomeria japonica D. Don) |
FUKUHARA, NARAKATSU (Japón) |
SECCIÓN 2a. GENÉTICA DE POBLACIONES, HEREDABILIDAD, APITUD COMBINATORIA Y PRUEBAS DE DESCENDENCIA
|
Arrangement and design of field experiments in progeny testing |
JOHNSSON, HELGE (Suecia) |
|
Mass selection and heritability studies in forest tree breeding |
TODA, RYOOKITI (Japón) |
|
Estimates of heritability published in forest tree breeding research |
HATTEMER, H. H. (Alemania, Fed. Rep.) |
|
Estimation of genetic parameters in forest trees without raising progeny |
SAKAI, KAN-ICHI & HATEKEYAMA, SUEKICHI (Japón) |
|
One parent progeny testing with black wattle (Acacia mearnsii de Wild) |
MOFFETT, A. A. & NIXON, KATHLEEN M. (Sudáfrica) |
|
Population genetics - breeding methods |
WETTSTEIN, W. W. (Austria) |
|
The height growth in seedlings of Picea abies (L.) Karst; a possibility to determine the material portion |
HOLZER, KURT (Austria) |
|
Some causes of variation in the initial development of Scots pine (Pinus sylvestris L.) |
HADDERS, GUSTAF (Suecia) |
|
Results from an 11-year-old progeny test with Picea abies (L.) Karst in southeastern Noruega |
RUDEN, TOLLEF (Noruega) |
|
Early tests |
SCHMIDT, W.(Alemania, Fed. Rep.) |
|
Prognose prématurée dans l'amélioration de la forme du tronc du peuplier noir |
ZUFA LAJOS (Yugeoslavia) |
|
Application de la méthode graphique a l'etude de la variabilité des populations |
DUMITRIU-TATARANU (Rumania) |
SECCIÓN 2b. HIBRIDACIONES ENTRE ESPECIES Y RAZAS
|
Experiments on forest tree hybrids in Bavaria from 1936 to 1962 |
ROHMEDER, E. (Alemania, Fed. Rep.) |
|
Performance of some aspen seed sources and hybrids in eastern Massachusetts |
PAULEY, SCOTT S. (Estados Unidos) |
|
On the fertilization in the crossings of Pinus rigida with P. taeda, P. radiata and P. elliottii |
KYUN, SIN KYU & YIM, KYONG BIN (Corea) |
|
The progress of seedling growth of poplar hybrids in relation to their selection |
BIALOBOK, S. (Polonia) |
|
Interspecific hybridization of several pine species from the subgenus Diploxylon koehne |
VIDAKOVIC, MIRKO (Yugeoslavia) |
|
Intraspecific hybridization and heterosis within Picea abies |
NILSSON, B. O. (Suecia) |
|
On the growth of artificial hybrids of sugi Cryptomeria japonica |
NOHARA, YUTA (Japón) |
|
Cruzamientos dentro de la sección Leuce y un híbrido notable entre Leuce y Aigeiros |
CATALÁN, G. (España) |
|
Considerations sur l'amelioration du genre Populus et spécialement sur la section Aigeiras |
MUHLE LARSEN, C. (Bélgica) |
|
Investigations of crossability of some Mediterranean pines and the American red pine |
MOULOPOULUS, C. & BASSIOTIS, C. (Grecia) |
|
Recherches biométriques sur les fruits et graines dans les travaux de sélection et d'amélioration du frêne |
BENEA, V., LEANDRU, L. & LAZARESCU, C. (Rumania) |
SECCIÓN 3. INVESTIGACIONES DE LA PROCEDENCIA
|
Practical results and current problems in provenance research in Suecia |
LANGLET, OLOF (Suecia) |
|
Provenance in tree improvement with particular references to Eucalyptus |
PRYOR, LINDSAY D. (Australia) |
|
Growth of Noruega spruce (Picea abies [L.] Karst.) provenances in eastern north America |
HOLST, MARK (Canada) |
|
The design of field tests |
WRIGHT, JONATHAN W. (Estados Unidos) |
|
Standardization of methods in provenance research and testing |
VYSKOT, MIROSLAV (Checoeslovaquia) |
|
Report on the state and preliminary evaluation of Czechoslovak provenance trial plots of Noruega spruce in the international series from the year 1938 |
VINS, B. (Checoeslovaquia) |
|
Some results from an anatomic investigation of Noruega spruce provenances in four international spruce tests of 1938 in Suecia and Noruega |
DIETRICHSON, JON (Noruega) |
|
Douglas-fir archives and their use in tree improvement |
IRGENS-MOLLER, H (Estados Unidos) |
|
Selection for drought resistance in sugar maple |
KRIEBEL, HOWARD B. (Estados Unidos) |
|
Les paroelles de recherche concernant les provenances du melèze fondées par Rubner sur le territoire de la Tchécoslovaquie |
SINDELAR, JIRI (Checoeslovaquia) |
|
Recherches anatomiques sur les feuilles comme complément a la connaissance morphologique des unites systématiques inférieures dans les populations de pin noir |
NICOTA, BOZIDAR (Yugeoslavia) |
|
Report on researches undertaken in Polonia on Scots pine (Pinus sylvestris) |
TYSZIEWICZ, St. (Polonia) |
|
Pinus cembra L. as a pioneer at timberline in the European Alps |
KURT, HOLZER (Austria) |
|
The geographical variation, the intraspecific taxonomy and the breeding of Scots pine (Pinus sylvestris) |
PRAVDIN, L. F. (U.R.S.S.) |
|
Les possibilitiés d'utiliser les mesures statistiques portent sur des cones et graines d'Epicea des Carpathes orientales du nord (Roumanie) en vue de la recon naissance des populations d'élite |
DUMITRIU-TATARANU, I. & BENEA, V.(Rumania) |
|
Monophage analysis of elm popuIations |
RICHENS, R. H. (Reino Unido) |
SECCIÓN 4. ESPECIES EXÓTICAS
|
Planning a genetical survey of Pinus radiata populations |
BANNISTER, M. H. (Nueva Zelandia) |
|
The use of exotic trees in increasing production with particular reference to northwestern Europe |
EDWARDS, M. V. (Reino Unido) |
|
Initiating a breeding program with Pinus patula in Southern Rhodesia |
HODGSON, L. M. (Rhodesia del Sur) |
|
Génétique des peupliers |
SEKAWIN, MICHELE (Italia) |
|
Experiences with the Douglas fir in Europe |
SCHOBER, R. (Alemania, Fed. Rep.) |
|
Experiences and present situation of foreign tree species and provenances in Suecia |
KIELLANDER, C. L. (Suecia) |
|
Ensayo de introducción de pings subtropicales en el nordeste Argentino |
BARRETT., W. H. H. (Argentina) |
|
La résistance des Eucalyptus au calcaire actif dans le sol. Compte rendu d'un test précoce |
LACAZE, M. (Francia) |
|
Contribution du Maroc pour l'amelioration génétique des Eucalyptus introduits dans le bassin méditerranéen |
CLAUDOT, JEAN (Marruecos) |
|
Sélection de certaines populations valeureuses de robinier faux Acacia et possibilitiés de Ieur multiplication par vole végetative |
ENESCO, Val., BIRLANESCU, E. & COSTEA, A. (Rumania) |
|
The geographical distribution of Populus Nigra L. var. Pubescens Parlat, and of Populus Thevestina Dode, and their hybrids |
PAPAIOANNOU, J. (Grecia) |
SECCIÓN 5. GENÉTICA Y FISIOLOGÍA ARBÓREA
|
Physiological implications in tree improvement |
KOZLOWSKI, T. T (Estados Unidos) |
|
Some physiological actions of Gibberellins on forest boos |
SATO, KEJI (Japón) |
|
Physiological investigations on Picea abies (L.) Karst in view of genetics |
HOLZER, KURT (Austria) |
|
Tree physiology-genetics studios at Oregon State University |
FERREL, WILLIAM K. (Estados Unidos) |
|
Amélioration des roboisements d'Eucalyptus par multiplication végétative |
FRANCLET, A.(Túnez) |
|
Grafting as an aid in the brooding of teak (Tectona grandis L. f.) and semal (Salmalia malabarica Schott et endl.) |
KEDHARNATH, S. & VENKATESH, C. S. (India) |
|
Plastic greenhouses in tree breeding practice |
LESKINEN, USKO (Finlandia) |
|
Enraizamiento de Eucalyptus camnaldulensis |
MARCAVILLACA, MANUEL & MONTALDI, EDGARDO, R. (Argentina) |
|
Comparison of nutritional properties in forest tree species |
INGESTAD, TORSTEN (Suecia) |
|
The possibility of using cuttings for the establishment of commercial plantations of Monterey pine |
FIELDING, JOHN M. (Australia) |
|
Etudes physiologiques et biochimiques dans les travaux d'améIioration du pin |
BENEA, V., LEANDRU, L. & NITU, C. (Rumania) |
|
Influence de la topophysie sur l'affinement apparent des clones de peuplier a écorce grise et rugueuse |
MAY, S. A. (Italia) |
SECCIÓN 6a. MEJORA DE LA RESISTENCIA A LAS ENFERMEDADES
|
Problems and progress in improvement of rust resistance of North American trees |
BINGHAM, R. T. (Estados Unidos) |
|
Improvement of disease resistance in Populus |
SCHREINER, ERNST J. (Estados Unidos) |
|
The possibility of establishing clinical indices in boos to determine their level of vigor and vulnerability to diseases caused by facultative parasites |
BIER, JOHN E. (Canadá) |
|
Resistance of pines to southern fusiform rust |
HENRY, B. W. & JEWELL, F. F. (Estados Unidos) |
|
Breeding sandal for resistance to the spike disease |
VENKATESH, C. S. & KEDHARNATH, S. (India) |
|
Brooding for resistance to disease of fast-growing trees and need for a more complex selection |
KRSTIC, M. M. (Yugeoslavia) |
SECCIÓN 6b. MEJORA DE LA RESISTENCIA A LOS ATAQUES DE INSECTOS
|
Integrated selection for white pine weevil resistance and its components |
GERHOLD, H. D. & STROH, R. C. (Estados Unidos) |
|
Studios on resistance of pings to insects |
SMITH, RICHARD H. & EATON, CHARLES B. (Estados Unidos) |
|
Breeding resistance in pines to Rhyacionia moths |
HOIST, MARK (Canadá) |
|
The brooding of white pine for resistance to weevil |
HEIMBURGER, C. (Canadá) |
SECCIÓN 7. MEJORAS DE LAS PROPIEDADES DE LA MADERA
|
Evaluating the environment for studios of the inheritance of wood properties |
LARSON, PHILIP R. (Estados Unidos) |
|
Inheritance of wood properties and their relation to growth rate in Pinus taeda |
BUIJTENEN, J. P. VAN (Estados Unidos) |
|
The influence of environment of the wood density of Radiata pine grown in New Zealand |
MADDERN HARRIS, J. (Nueva Zelandia) |
|
Variation in fiber length in Populus tremuloides clones |
VALENTINE, F. A. (Estados Unidos) |
|
Timber improvement by tree selection and breeding |
PHILLIPS, E. W. J. (Reino Unido) |
|
Present status of forest tree breeding in India |
KEDHARNATH, S. (India) |
|
Observations on timber quality and out-turn of Scots pine |
OPENSHAW, K. (Reino Unido) |
|
Wood density in progenies after hybridization within Picea abies |
NILSSON, Bo (Suecia) |
SECCIÓN 8. PRODUCCIÓN Y CERTIFICACIÓN DE SEMILLA
|
Seed stands and seed orchards in the breeding of conifers |
ANDERSSON, E. (Suecia) |
|
Problems of flowering and seed production |
RISTO, SARVAS (Finlandia) |
|
Forest tree seed certification in the Estados Unidos and some proposals for uniformity |
RUDOLF, P. O. (Estados Unidos) |
|
Tree seed certification and forest genetics |
ELIASON, E. J. (Estados Unidos) |
|
Consequences of the development of varieties in forestry |
LANGNER, W. (Alemania, Fed. Rep.) |
|
Basic principles of origin certification |
BARNER, H. (Dinamarca) |
|
Relation entre la hauteur des arbres et la fructification du pin maritime (Pinus pinaster Sol.) |
ILLY, GEORGES (Francia) |
|
Some results on the determination of seed sources by the ignitibility of forest tree seed |
DEICHMANN, V. VON (Brasil) |
|
Critères phénotypiques de choix des plantes plus de pin pigeon |
CAPPELLI, M. (Italia) |
|
Contribution de la morphologic du pollen a la génétique forestière. Disparition d'une espèce Cedrus libani Loud.) dans certaines regions |
AYTUG, BURHAN (Turquía) |
|
Identification des peuplements d'élite pour les réservations de semences par cartographie seminologique |
ENESCO, Val., BENEAN, V. and CATANESCU, D. (Rumania) |
SECCIÓN 9. GENÉTICA DE LA PRÁCTICA FORESTAL EN SUECIA
Los siguientes fueron presentados por los miembros suecos de la Consulta durante los tres viajes de estudio y visitas sobre el terreno a Bogesund.
|
The Noruega spruce provenance experiments at Dönjelt and Hjuleberg |
LANGLET, O. |
|
Genetic variation in progeny tests of Scots pine (Pinus sylvestris) |
EHRENBERG, C. |
|
Graft production, grafting and the production of cuttings |
PERSSON, S. |
|
On the anatomy of graft unions in Scots pine and Noruega spruce |
DORMLING, I. |
|
On grafting methods |
DORMLING, I. |
|
The Department of Forest Genetics, Royal College of Forestry, Stockholm. Research Notes No. 1, 1963 |
GUSTAFSSON, A. |
|
The Swedish Forest Tree Breeding Association |
ARNBORG, T.& JOHNSSON, H. |
|
Aspen breeding end research. The Swedish Match Company, Mykinge Experiment Station |
HÜPPEL, A. & JOHNSSON, H. |
|
Experimental provenance plantation with Scots pine at Alvkarleby, Suecia |
EICHE, V. |
Miembros de las mesas
|
Presidente: |
LUNDH, ERIK |
|
Vice-Presidentes: |
AMAREL GURGEL, J. DE |
|
TODA, RYOOKITI |
|
|
LANGNER, W. |
|
|
WETTSTEIN, W. |
|
|
Relator principal: |
MATTHEWS, J. D. |
|
Secretario General: |
MÉTRO, A. |
|
Secretario General Asistente: |
FUGALLI, O. |
|
Secretario Técnico: |
SIRÉN, G. |
N. A. OSARA, Director, Dirección de Montes y Productos Forestales, en representación del Director General de la FAO.
COMITÉ ORGANIZADOR
|
Miembro honorario: |
SYLVÉN, N. |
|
Presidente: |
PLYM FORSHELL, W. |
|
Vice-Presidente: |
GUSTAFSSON, ÅKE |
|
Secretario General: |
JOHNSSON, H. |
|
Secretario: |
EHRENBERG, C. |
|
Miembros: |
ANDERSSON, E. |
|
ANDRÉN, TH. |
|
|
BJÖRKMAN E. |
|
|
EKMAN, S - G. |
|
|
HAGBERG, E. |
|
|
HÄGGSTROM, B. |
|
|
LANGLET, O. |
|
|
STEFANSSON, E. |
|
|
WIJLSTRÖM, S. |
SECCIÓN 1. GENÉTICA ARBÓREA Y CITOLOGÍA
|
Presidente: |
SAX, K. |
|
Relator principal: |
GUSTAFSSON, A. |
|
Co-Relatores: |
MERGEN, F. |
|
KHOSHOO, T. N. |
|
|
HYUN, S. K. |
|
|
Secretario: |
MERGEN, F. |
SECCIÓN 2. GENÉTICA DE POBLACIONES, HEREDABILIDAD, APTITUD COMBINATORIA Y PRUEBAS DE DESCENDENCIA
|
Presidente: |
PHILIPPIS, A. DE |
|
Relator principal: |
STERN, K. |
|
Co-Relatores: |
JOHNSSON, H. |
|
TODA, R. |
|
|
Secretario: |
STRAND, L. |
SECCIÓN 2b. HIBRIDACIONES ENTRE ESPECIES Y BAZAS
|
Presidente: |
HEIMBURGER, C. C. |
|
Relator principal: |
WRIGHT, J. W. |
|
Co-Relatores: |
MUHLE LARSEN, C. |
|
ROHMEDER, E. |
|
|
Secretario: |
JONES, E. W. |
SECCIÓN 3. INVESTIGACIONES DE EA PROCEDENCIA
|
Presidente: |
BOUVAREL, P. |
|
Relator principal: |
CALLAHAM, R. Z. |
|
Co-Relatores: |
LANGLET, O. |
|
PRYOR, L. D. |
|
|
Secretario: |
LACAZE, J. |
SECCIÓN 4. ESPECIES EXÓTICAS
|
Presidente: |
FIELDING, J. M. |
|
Relator principal: |
MORANDINI, R. |
|
Co-Relatores: |
BANNISTER, M. H. |
|
SEKAWIN, M. |
|
|
EDWARDS, M. V. |
|
|
SCHOBER, R. |
|
|
Secretario: |
FAULKNER, R. |
SECCIÓN 5. GENÉTICA Y FISIOLOGÍA ARBÓREA
|
Presidente: |
FOWELLS, H. A. |
|
Relator principal: |
WAREING, P. F. |
|
Co-Relatores: |
HOLZAR, K. |
|
SATO, K. |
|
|
KOZLOWSKI, T. T. |
|
|
Secretario: |
INGESTAD, T. |
SECCIÓN 6a. MEJORA DE BA RESISTENCIA A LAS ENFERMEDADES
|
Presidente: |
SCHREINER, E. J. |
|
Relator principal: |
BJÖRKMAN, E. |
|
Co-Relator: |
BINGHAM, R. T. |
|
Secretario: |
BINGHAM, R. T. |
SECCIÓN 6b. MEJORA DE EA RESISTENCIA A LOS ATAQUES DE INSECTOS
|
Presidente: |
SCHREINER, E. J. |
|
Relator principal: |
SØEGAARD, B. |
|
Co-Relator: |
GERHOLD, H. |
|
Secretario: |
GERHOLD, H. |
SECCIÓN 7. MEJORA DE LAS PROPIEDADES DE MADERA
|
Presidente: |
PHILLIPS, E. W. |
|
Relator principal: |
ZOBEL:, B. J. |
|
Co-Relatores: |
MADDERN HARRIS, J. |
|
VAN BUIJTENEN, J. |
|
|
Secretario: |
LARSON, P. |
SECCIÓN 8. PRODUCCIÓN Y CERTIFICACIÓN DE SEMILLA
|
Presidente: |
KRIEBEL, H. |
|
Relator principal: |
MATTHEWS, J. |
|
Co-Relatores: |
ANDERSSON, E. |
|
SARVAS, R. |
|
|
Secretario: |
BARNER, H. |
SECCIÓN 9. GENÉTICA DE EA PRÁCTICA FORESTAL EN SUECIA
|
Presidente: |
SYRACH LARSEN, C. |
|
Relator principal: |
PLYM FORSHELL, W. |
|
Secretario: |
AMBORG, T. |
Participantes
|
Amaral Gurgel, José de |
Brasil |
|
Andersson, Enar |
Suecia |
|
Andrén, Thorsten |
Suecia |
|
Arnborg, Tore |
Suecia |
|
Aytug, Burhan |
Turquía |
|
Badra, Mahmoud |
Túnez |
|
Barner, Helmuth |
Dinamarca |
|
Barner, Burton V. |
Estados Unidos |
|
Bassiotis, Constantine |
Grecia |
|
Bastos de Menezes, Osvaldo |
Brasil |
|
Bay, Bendt |
Nueva Zelandia |
|
Benea, Vasile |
Rumania |
|
Bergman, Fritz H. F. |
Suecia |
|
Betancourt Barroso, M. R. |
Cuba |
|
Bialobok, Stefan |
Polonia |
|
Bier, John E. |
Canadá |
|
Bingham, Richard T. |
Estados Unidos |
|
Biraghi, Antonio |
Italia |
|
Björkman, Erik |
Suecia |
|
Boonkird, Sa-ard |
Tailandia |
|
Bouvarel, Pierre |
Francia |
|
Bunn, Ernest H. |
Nueva Zelandia |
|
Burdon, Rowland D. |
Nueva Zelandia |
|
Buijtenen, Johannes P. van |
Estados Unidos |
|
Callaham, R. Z. |
Estados Unidos |
|
Castellani, Ettore |
Italia |
|
Catalán, Gabriel |
España |
|
Chalmers, William S. |
Trinidad y Tabago |
|
Clear, Thomas |
Irlanda |
|
Cooling, Edwin N. G. |
Rhodesia |
|
Deasy, Jeremiah J. |
Irlanda |
|
Dietrichson, Jon |
Noruega |
|
Dimpfimeier, Rudolf |
Alemania, Rep. Fed. |
|
Donaubauer, Edwin |
Austria |
|
Edlund, Erik |
- Suecia |
|
Edwards, M. V. |
Reino Unido |
|
Ehrenberg, Carin |
Suecia |
|
Ekman, Sven G. |
Suecia |
|
Eliason, Everett |
Estados Unidos |
|
Esbo, Harald |
Suecia |
|
Fassi, Bruno |
Italia |
|
Faulkner, Roy |
Reino Unido |
|
Ferrell, William K. |
Estados Unidos |
|
Fielding, John M. |
Australia |
|
Fowells, Harry A. |
Estados Unidos |
|
Franclet, André |
Túnez |
|
Fröhlich, Hans I. |
Alemania, Rep. Fed. |
|
Furukoshi, Takanobu |
Japón |
|
Geary, Thomas F. |
Rhodesia |
|
Gerhold, Henry D. |
Estados Unidos |
|
Giertych, Maciej M. |
Polonia |
|
Gonser, Hans |
Alemania, Rep. Fed. |
|
Gustafsson, Åke |
Suecia |
|
Göhrn, Viggo A. |
Dinamarca |
|
Hadders, Gustaf |
Suecia |
|
Hafsia, Hamda |
Túnez |
|
Hagberg, Erik |
Suecia |
|
Hagman, Max |
Finlandia |
|
Hedlund, Hans |
Suecia |
|
Heimburger, Carl C. |
Canadá |
|
Hermelin, S. N. Joakim S. |
Suecia |
|
Heybroek, Hans M. |
Países Bajos |
|
Holst, Mark J. |
Canadá |
|
Holzer, Kurt |
Austria |
|
Häggström, Börje |
Suecia |
|
Härdén, Sven |
Suecia |
|
Höjer, Erik |
Suecia |
|
Ingestad, Torsten |
Suecia |
|
Irgens-Moller, Helge |
Estados Unidos |
|
Jamblinne de Meux, Alain de |
Bélgica |
|
Jaynes, Richard A. |
Estados Unidos |
|
Johansson, Folke L. H. |
Suecia |
|
Johnsson, Helge |
Suecia |
|
Jones, Eustace W. |
Reino Unido |
|
Joranson, Philip N. |
Estados Unidos |
|
Jovancevic, Milorad |
Yugoeslavia |
|
Juma, Salah |
Jordania |
|
Jurriaanse, Aart |
Sudáfrica |
|
Keet, John N. |
Reino Unido |
|
Keiding, Henrik |
Dinamarca |
|
Kiellander, Carl L. |
Suecia |
|
Kittani, Hassan |
Irak |
|
Kleinschmit, Richard |
Alemania, Rep. Fed. |
|
Koster, Robbert |
Países Bajos |
|
Kriebel, Howard B. |
Estados Unidos |
|
Krstic, Mihailo |
Yugoeslavia |
|
Krutzsch, Peter |
Suecia |
|
Kunjara Na Ayudhya, V. |
Tailandia |
|
Kärki, Lauri E. P. |
Finlandia |
|
Lacaze, Jean F. |
Francia |
|
Langlet, Olof |
Suecia |
|
Langner, Wolfgang |
Alemania, Rep. Fed. |
|
Larson, Philip R. |
Estados Unidos |
|
Leskinen, U. |
Finlandia |
|
Ljunger, Åke |
Suecia |
|
Lundh, Erik |
Suecia |
|
Lyytinen, Oiva |
Finlandia |
|
Løfting, Ebbe C. L. |
Dinamarca |
|
Marcet, Enrique |
Suiza |
|
Matéern, Bertil |
Suecia |
|
Malos, Gonzáles E. |
Cuba |
|
Matthews, J. D. |
Reino Unido |
|
May, Silvio |
Italia |
|
McLean, Colín |
Reino Unido |
|
Mergen, François |
Estados Unidos |
|
Morandini, Riccardo |
Italia |
|
Moulopoulos, Christos |
Grecia |
|
Muhle Larsen, Carl |
Bélgica |
|
Nanson, Alphonse W. J. |
Bélgica |
|
Nicovescu, Horia |
Rumania |
|
Nilsson, Bo |
Suecia |
|
O'Carroll, Niall |
Irlanda |
|
O'Driscoll, John |
Irlanda |
|
Openshaw, Keith |
Reino Unido |
|
Palosuo, Veikko J. |
Finlandia |
|
Papaioanuou, John |
Grecia |
|
Philippis, Alessandro de |
Italia |
|
Phillips, Edgar William J. |
Reino Unido |
|
Plym Forshell, Wilhelm |
Suecia |
|
Pryor, Lindsay Dixon |
Australia |
|
Puente E., José M. de la |
México |
|
Richens, Richard Hook |
Reino Unido |
|
Robak, Håkon |
Noruega |
|
Rohmeder Ernst |
Alemania, Rep. Fed. |
|
Ruden, Tollef |
Noruega |
|
Ruggeri, Cecilia |
Italia |
|
Samapuddhi, Krit |
Tailandia |
|
Sarvas, Risto Olavi |
Finlandia |
|
Sax, Karl |
Estados Unidos |
|
Scattolin, Mario |
Italia |
|
Schmidt, Helmut |
Alemania, Rep. Fed. |
|
Schmitt, Richard |
Alemania, Rep. Fed. |
|
Schreiner, Ernst T. |
Estados Unidos |
|
Sekawin, Michele |
Italia |
|
Simolinna, Jaakko Einar |
Finlandia |
|
Sriburi, Taweesakdi |
Tailandia |
|
Steenackers, Victor Maria |
Bélgica |
|
Stefansson, Eric |
Suecia |
|
Stern, Klaus |
Alemania, Rep. Fed. |
|
Streyffert, Torsten |
Suecia |
|
Swabey, Christopher |
Reino Unido |
|
Syrach Larson, Carl |
Dinamarca |
|
Søegaard, Bent |
Dinamarca |
|
Toda, Ryookiti |
Japón |
|
Valentine, Fredrick, A. |
Estados Unidos |
|
Vecchi, Emma de |
Italia |
|
Vidakovic, Mirko |
Yugoeslavia |
|
Visuthakul, Ninnat |
Tailandia |
|
Wareing, Philip F. |
Reino Unido |
|
Wettstein, Wolfgang |
Austria |
|
Wijkström, Stig |
Suecia |
|
Wright, Jonathan W. |
Estados Unidos |
|
Yli-Vakkuri, Paavo Olavi |
Finlandia |
|
Zobel, Bruce |
Estados Unidos |
|
Zufa, Lajos |
Yugoeslavia |
|
Strand, Lars |
Biom. Soc. |
|
Dyson, William G. |
EAAFRO |
|
Callaham, Robert Z. |
IUBS |
|
Matthews, John D. |
IUFRO |
|
Sindelar, Jirí |
IUFRO |
|
Vyskot, Miroslav |
IUFRO |
|
Esbo, Harald |
OECD, ISTA |
|
Osara, Nils A. |
Director, Dirección de Montes y Productos Forestales, FAO |
|
Metro, André |
Jefe, Subdirección de Producción Forestal, FAO |
|
Fugalli, Oscar |
Subdirección de Producción Forestal, FAO |
|
Sirén, Gustaf |
Subdirección de Producción Forestal, FAO |
|
Deichmann, Vollrat von |
Oficial de Montes de la FAO, Estola Nacional de Florestas Minas Gerais, Brásil |
|
Silow, R. A. |
Subdirección de Energía Atómica, FAO |
![]()
![]()
![]()
{kind=link}