![]()
![]()
![]()
It is important to understand that crises in fishery resources are not limited to particular stocks in the world, but occur in many regions, with a frequency that indicates a major problem to be taken into consideration (Fig. 1). Although catches are not a good indication of abundance, it is generally admitted that big fluctuations reflect mainly variations in abundance. The rapid rise in catches and their sharp fall after several years of intensive exploitation should be noted, particularly over the last two decades. Figure 2 shows that these variations can take the form of regular fluctuations over a long period (nearly two solar cycles, for example). Figure 3 shows that the fluctuations observed for stocks in various parts of the world can be synchronized, which would suggest that climatic factors play an important role in such fluctuations.
In the eastern Atlantic, very important variations have also been observed. The catches and the biomass of sardines have increased in the area to the north of Cap Blanc in an exceptional way (Belvéze and Erzini, 1983), whereas fishing grounds stretched about 850 km to the south (see Fig. 1 and Garcia, 1982). In the same region and especially to the north of Cap Bojador, unexploited stocks of oyster-catchers (Macroranphosus scolopax) underwent considerable fluctuations of biomass (ISPM,1983), providing, in the mid-1970s, a recently created potential of 400 000 t, which practically disappeared without exploitation at the beginning of the 1980s. Stocks of sardinella and mackerel simultaneously dropped sharply along the coasts of the Sahara, while the geographic distribution of sardinella regressed toward the south, and a collapse of sardinella stocks and disappearance of mackerel were observed in the west of the Gulf of Guinea, from the Ivory Coast to Togo (Fig. 4). At the same time, triggerfish, a traditionally scarce species, developed biomasses estimated at several hundreds of thousands of tons in the Gulf of Guinea and in the Sherbro-Cape Verde sector, even seasonally invading the northern coasts of Senegal as far as Mauritania (Caverivière, 1982).
Along Namibia, fluctuations seem linked to the collapse of sardine and anchovy stocks and the gradual increase of certain stocks of horse mackerel (Fig. 1).
In California, South Africa and Peru, it has been shown that variations in abundance are associated with variations in a large number of biological parameters such as growth, age of first sexual maturity, factors of condition, fertility, natural mortality, geographic distribution, etc., indications of profound “mutations” in the population as a result of very heavy stress. These marked changes over the years seem to have been only another aspect of variability, whose importance at geographical level has already been pointed out, since variations in biological parameters are also considerable from one region to another. Also, it is now well established that the gregarious behaviour of these species leads to significant changes in their catchability (which increases when the biomass drops).
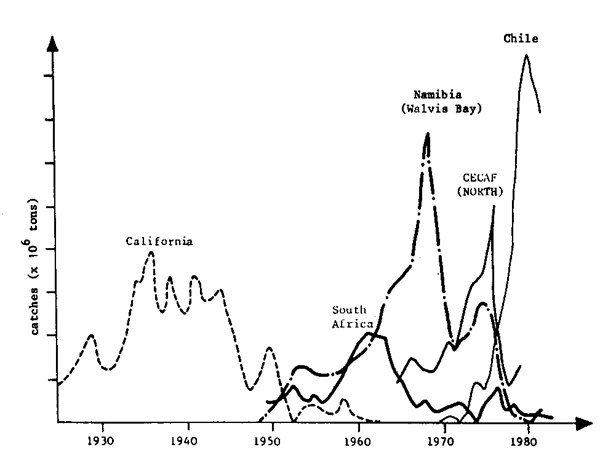
Fig. 1 Variations in sardine landings in different regions of upwelling (farrish, 1983)
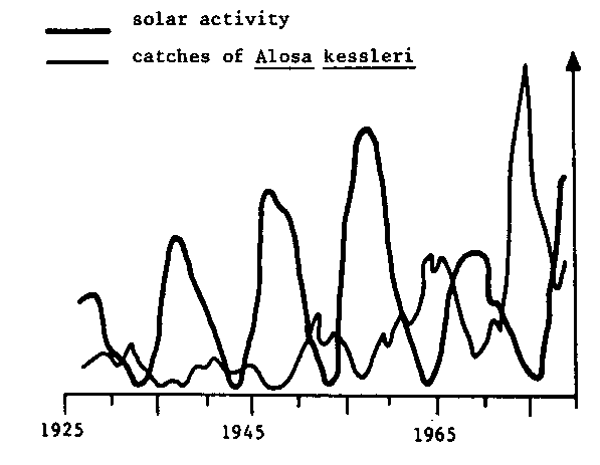
Fig. 2 Variations in catches of Alosa kessler=i in the Black Sea (Ivanov, 1984)
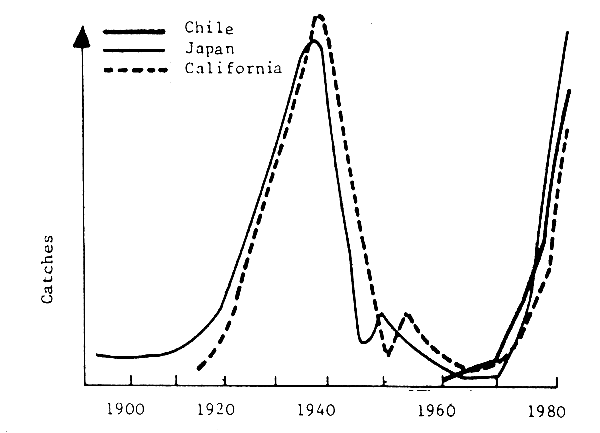
Fig. 3 Simultaneous variations in sardine landings (ref.)
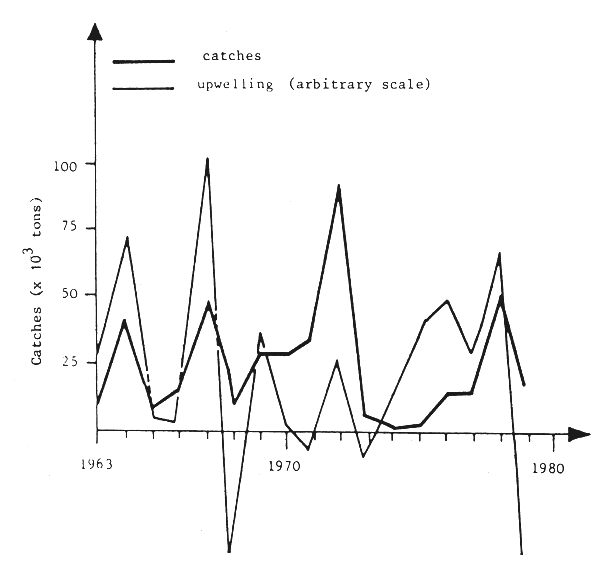
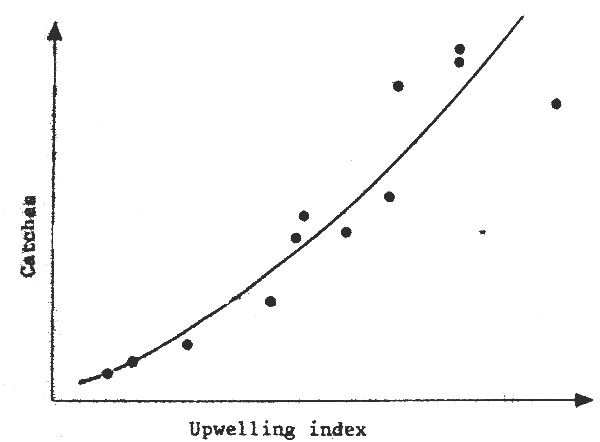
Fig. 4 Upwelling and catches of S. aurita in the Gulf of Guinea (data from CRO, Abidjan, FAO/CECAF, 1980)
It has been suggested many times that the biomass produced by an ecosystem is more constant than that of the specific elements that compose it, or in other words, the species whose stock has collapsed will be replaced by another. The case of the sardine and the anchovy of California has become a classic case used to expound the theory of interspecific competition. However, more recently, MacCall (1983) contested the hypothesis of “replacement” of species in the strict sense (implying phenomena of interspecific competition) in the case of California, and considered it more likely to be a question of important changes in the biomass and the geographic distribution of resources, as a result of environmental action.
There are many examples of apparent replacements (Figs. 5 to 9) in most regions of the world, and the eastern central Atlantic has also been the scene of very significant replacements (Garcia, 1982).
Between Cap Blanc and Cap Barbas, the replacements observed with small pelagic species are due to changes in geographic distribution and in stocks (Belvèze, 1983). It is nonetheless true that these movements, in the case of sardine and triggerfish, are accompanied by considerable increases in the biomass of these species, to the detriment, it seems, of other species (Caveriviére, 1982; Gulland and Garcia, 1984). It is important to note that these replacements, which do not always appear as immediate or automatic, can be advantageous, like the replacement of the Sparidae by the Cephalopodae in the northern CECAF zone, or disadvantageous, like the replacement of a traditionally marketed species by triggerfish throughout the Gulf of Guinea.
Important work has been done in describing variations in resources and attempting to understand their mechanisms, in California (McCall, 1983), in Peru (Jordan, 1983), in Japan (Hayasi, Tanaka, 1983), etc., and in Morocco (Belvéze and Erzini, 1983), Senegal (Fréon, 1983), the Ivory Coast and Ghana (FRU/ORSTOM, 1975; FAO/CECAF, 1980; Binet, 1982; Gulland and Garcia, 1983). The Expert Consultation organized by FAO in April 1983 discussed the subject at length (Sharp and Csirke, 1983).
Although there are still divergences of opinion as regards the detailed mechanisms, it is now widely admitted that these variations are linked essentially to climatic fluctuations whose consequences are aggravated by excessively intensive exploitation.
Climatic variations are probably reflected in variations in the rate of survival of recruits, and therefore, their abundance. Variations, which are reflected in short series of good or bad recruitments, are accompanied by variations in sexual maturity, growth, fertility, natural mortality, etc. The stock-recruitment ratio is also affected, and it may be said that variations in abundance are linked to changes in the biological capacity of the environment vis-à-vis the larva stage.
It is not impossible that interspecific competition, predator-prey relationships, or the side effects of fishing (on another species than the one whose fluctuations are under consideration) may also have an influence, but this has never been satisfactorily demonstrated, whereas the effects of the environment are often obvious.
Information collected from sediments on the abundance of stocks of sardines and anchovy in the past, have shown that biological collapses took place, practically without any exploitation, over the last two centuries (Soutar and Isaacs, 1974).
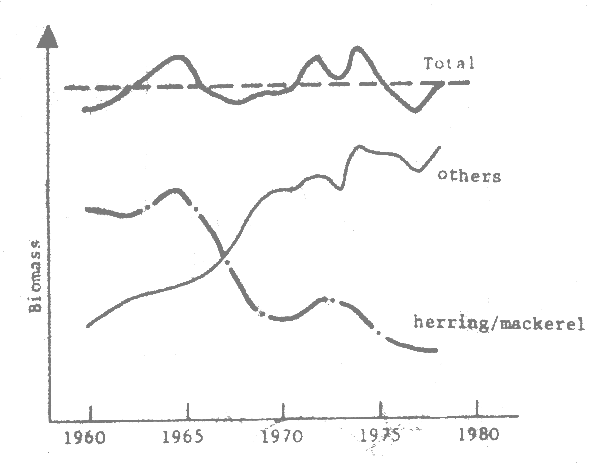
Fig. 5 North Sea
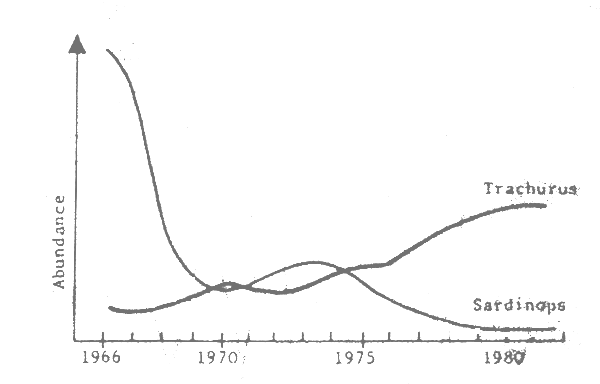
Fig. 6 Namibia
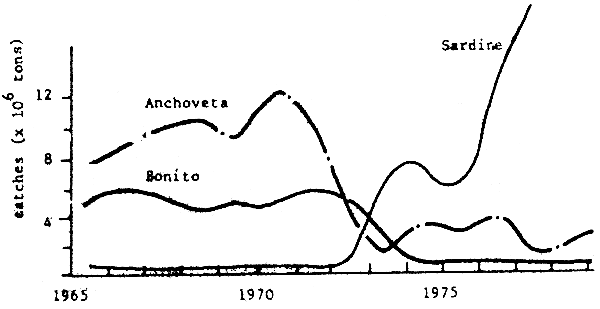
Fig. 7 Peru
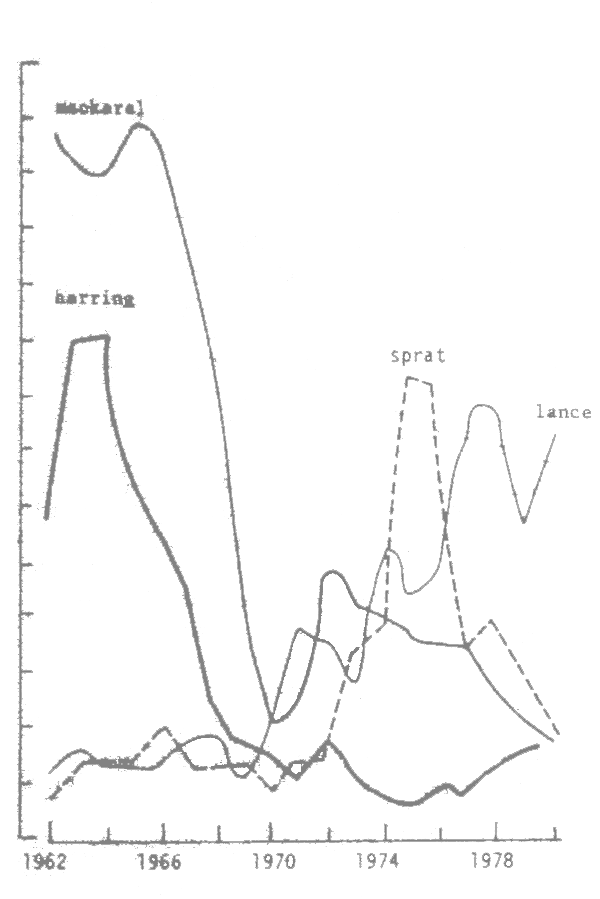
Fig. 8 Variation of the biomass of herring and mackerel spawn and landings of sprat and lance in the North Sea (Jones, 1983)
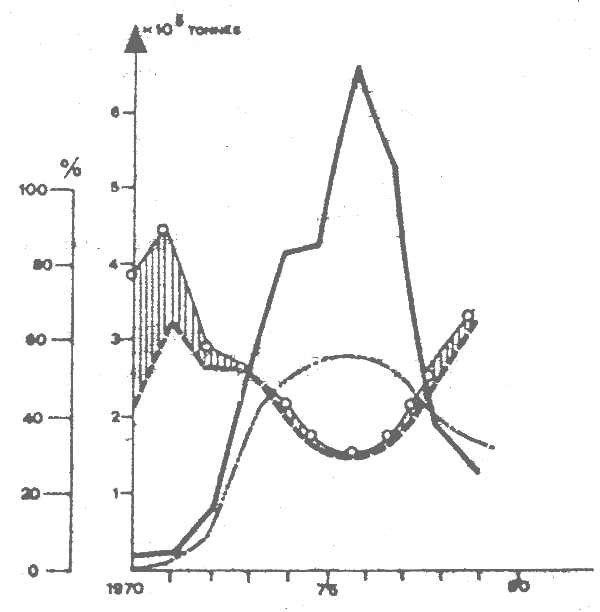
Fig. 9 Variations in sardine catches (—) and proportions ofsardine (—), horse mackerel and mackerel (—) horse macherel, macherels and sardinella (0—0). Source:CECAF statistical Bulletin No. 3, 1981
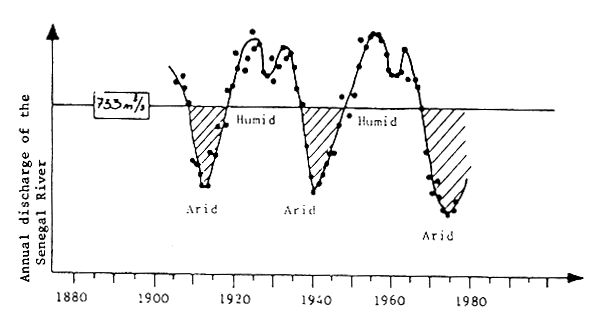
Fig. 10 Long-term climatic variations in West Africa (Faure and Gac, 1981)
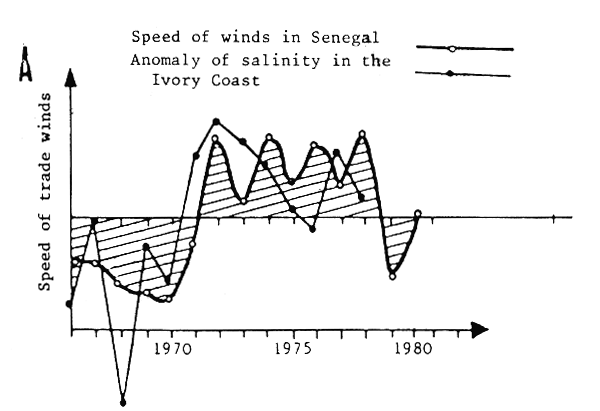
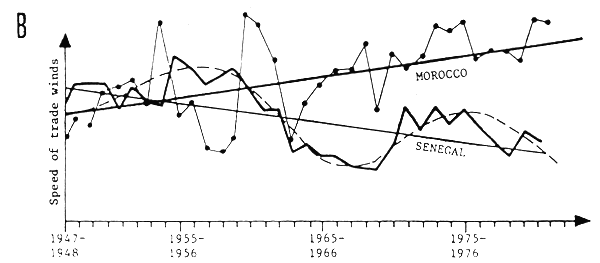
Fig. 11 Hydroclimatic variations in West Africa: A - Anomaly of salinity at Abidjan (FAO/CECAF, 1981) and speed of trade winds in Senegal (Fréon, 1983) B - Index of upwelling in Morocco (zone B), (Belvèze, 1983) and speed of winds in Senegal (Fréon, 1983)
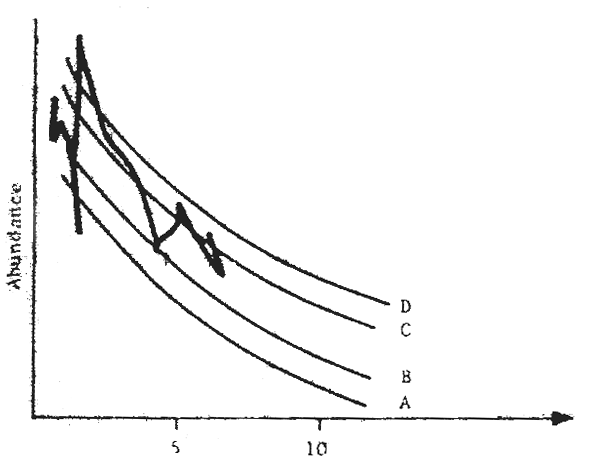
Fig. 12 Production model modified by wind section (and therefore by upwelling for the sardinella in Senegal) (Fréon, 1983)
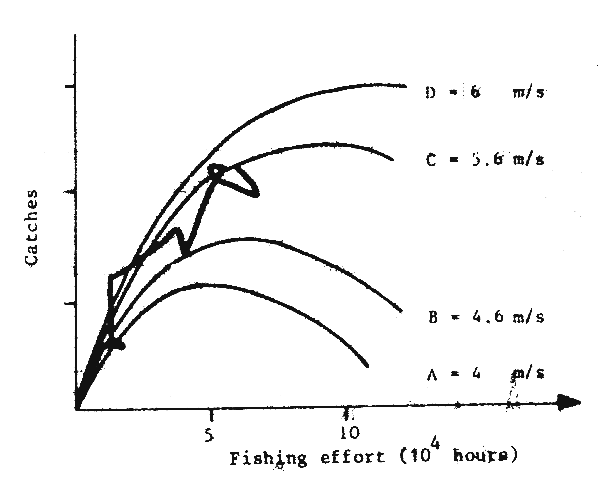
Fig. 13 Catches of sardines, and upwelling in Morocco (Belvè and Erzini, 1983)
It is however, admitted that fluctuations are aggravated by fishing, which also considerably increases the risk of collapse.
Analysis of this variability is interesting, since it indicates the type of variability to be expected at the level of the resource. Figure 10 clearly demonstrates that one of the phenomena marking the climate of West Africa is the existence of a very clear cycle of Sahelian droughts. These droughts are linked to movements of atmospheric fronts, and cause considerable variations in river discharges, and therefore in the salinity and primary productivity of the coastal areas (Binet, 1982).
They are accompanied by major changes in the speed and direction of prevailing winds, particularly winds in upwelling regions (Figs. 4, 11A and 11B), which are also reflected in major variations in primary productivity. Figures 11A and 11 suggest, moreover, that hydroclimatic variations observed along the coast and over the Atlantic are perhaps correlated. The variations appear similar or opposite depending on the case, and taking into account the shortness of the time series. This hypothesis must, however, be considered with extreme caution. Figure 11B indicates that apart from the medium frequency fluctuations, there are also long-term trends toward a weakening of the upwelling (Senegal) or its strengthening (Morocco).
The work of Belvèze and Erzini (1983) in Morocco, and Fréon (1983) in Senegal, indicates that variations in abundance of stocks are linked to the environment (Figs. 12 and 13). Fréon has even proposed a production model for the sardinella of Senegal, including environmental fluctuations.
![]()
![]()
![]()