![]()
![]()
![]()
K.D. Singh
K.D. Singh, du service forestier de l'Inde, est spécialiste des inventaires forestiers et du 8 traitement des données.
L'auteur analyse en détail trois types de variation spatiale dans les inventaires de forêts tropicales humides. Il en conclut que ce qui ressemble à un déconcertant chaos végétal a, en fait, une structure organisée.
Cet article décrit quelques types de variation fréquemment observés en forêt humide; ces descriptions sont fondées principalement sur des éléments déjà connus, mais aussi sur des documents inédits. On discutera d'abord des macrovariations en montrant leur intérêt pour la stratification. Puis on décrira les types de variation à une échelle intermédiaire, en ayant en vue leur rôle dans le choix du plan d'échantillonnage, à savoir dispositif en grappes ou non, aléatoire ou systématique. Pour terminer, on étudiera les microvariations en indiquant leur importance pour le choix de la dimension et de la forme de l'unité d'échantillonnage.
La forêt tropicale humide est la végétation climatique naturelle de la ceinture équatoriale - s'étendant jusqu'à environ 8 à 10 degrés de part et d'autre de l'équateur. Les facteurs climatiques essentiels pour son développement sont une: température uniformément élevée (environ 24-26°C) et une longue saison humide de 10 à 12 mois environ.
La forêt tropicale humide se trouve principalement dans les régions suivantes:
Amérique du Sud et Amérique centrale - Bassin. de l'Amazone (Brésil), s'étendant à l'ouest jusqu'aux pentes inférieures des Andes, à l'est jusqu'aux Guyanes, au sud jusqu'au Gran Chaco, et au nord le long du côté oriental de l'Amérique centrale.
Afrique - Bassin du Congo (principalement au Zaïre et au Congo), Gabon et Cameroun. La forêt tropicale pénètre vers l'ouest dans le Nigeria, le Ghana, la Côte-d'Ivoire, le Libéria et la Guinée; vers l'est elle s'étend jusqu'à l'Ouganda.
Asie - Principalement dans l'Asie du Sud-Est, y compris la péninsule de Malaisie, les îles de Sumatra, de Bornéo, des Célèbes et de Nouvelle-Guinée. On trouve aussi la forêt tropicale en Inde, à Sri Lanka, en Birmanie, en Thaïlande, en Indochine et aux Philippines.
Bien que la forêt tropicale humide soit présente dans des régions très éloignées les unes des autres, les études d'écologie mettent en évidence des ressemblances frappantes entre les formations. Selon Richards (1971) elles sont composées de synœcies ou communautés d'arbres semblables, aux caractéristiques homogènes et montrant une grande similitude dans leur disposition spatiale. Les espèces composant les synœcies correspondantes dans différentes régions géographiques ont, dans une large mesure, une physionomie comparable et une similitude biologique. Ainsi, La structure fondamentale est la même pour toute la forêt humide.
Ces constatations incitent à rechercher les type, courants de variation, ce qui peut être utile pour établir les dispositifs d'inventaires. Il est évident qu'une telle connaissance peut faciliter l'échange d'informations en matière d'inventaire entre les régions et accroître l'efficacité des plans d'enquête.
Pour planifier un inventaire, il est intéressant de connaître des types de variation à des échelles très différentes. Pour la stratification, on recherchera les macrovariations concernant par exemple les changements dans l'utilisation des terres, les types de forêts, et la classe d'exploitabilité. Pour définir un plan d'échantillonnage, il faut connaître la structure de variation à l'échelle intermédiaire. Pour le choix de la dimension et de la forme de l'unité d'échantillonnage, la microvariation a plus d'importance.
On trouvera ci-après une brève description des structures de variation à chacune des trois échelles.
Les causes de différenciation à l'intérieur de la forêt humide peuvent être divisées en deux grandes catégories: les causes naturelles et les causes culturales.
Pour ce qui est des facteurs naturels, le climat est relativement uniforme dans toutes les régions de forêt humide. Le sol et la topographie apparaissent comme les principaux agents de variation. Parmi les types courants on citera:
- La forêt humide climatique (en général sur sols bien drainés et à des altitudes inférieures à 500-800 mètres).- La mangrove.
- La forêt de marécage.
- La forêt périodiquement inondée.
- La forêt ripuaire.
- La forêt de collines (en général à des altitudes supérieures à 500-800 mètres).
- La forêt de podzols tropicaux.
- La forêt de sables côtiers.
Des facteurs culturaux - influence positive ou négative de l'homme - provoquent d'autres modifications dans la forêt humide naturelle; ainsi, on peut distinguer les grands types ci-après:
- La forêt humide primaire (la moins influencée par l'homme).
- La forêt exploitée.
- La forêt secondaire (reconstituée après des cultures itinérantes).
- L'association forêt-pâturage.
- La forêt aménagée (naturelle et artificielle).
- Les autres classes d'utilisation des terres non boisées.
Il y a des différences marquées entre les types ci-dessus en ce qui concerne les essences, le nombre de pieds à l'hectare, les étages et le volume à l'hectare. On peut également observer que la tendance à l'uniformité est d'autant plus marquée que le milieu est moins favorable.
D'après certaines études, il est possible d'identifier la plupart des macrovariations, à la fois en forêt naturelle et en forêt artificielle, au moyen de la photographie aérienne classique à l'échelle du 1/20000e ou à une échelle inférieure.
Comme la forêt humide primaire occupe une superficie relativement étendue par rapport aux autres types, des recherches ont été faites pour explorer les possibilités d'une stratification plus poussée. Nyyssönen (1961) et Lötsch et al. (1973) en ont analysé les résultats. Il semble assez difficile de faire une distinction plus fine entre les types écologiques à l'aide de photographies aériennes, parce que la forêt humide constitue un couvert végétal d'apparence très uniforme, sans changement ni démarcation sur de vastes étendues.
En revanche, il s'est révélé possible de distinguer de grandes classes de volume ou de surface terrière, à partir de la corrélation entre la densité des cimes des arbres dominants et le volume brut à l'hectare de toutes les essences. L'auteur a entrepris une analyse pilote d'un inventaire effectué au Laos en 1969, d'après les données de 185 placeaux; il en ressort que la valeur du carré du coefficient de corrélation multiple (R2) était de 0,64. Une relation étroite a aussi été constatée dans l'inventaire FAO de la vallée de l'Amazone.
TABLEAU 1. - AMÉLIORATION DE LA PRÉCISION GRÂCE A LA STRATIFICATION1
|
Plan d'échantillonnage |
Estimations de précision pour les régions de l'inventaire (±) |
|||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
|
|
...Pourcentage... |
||||||||
|
Sans stratification |
6,7 |
8,1 |
9,6 |
6,6 |
7,2 |
7,9 |
10,4 |
10,0 |
|
Stratification (après coup) |
4,5 |
6,8 |
7,6 |
4,0 |
3,7 |
4,2 |
5,4 |
5,6 |
1 Estimation de la précision du paramètre du volume industriel net de toutes essences d'un diamètre de 18 inches (45 cm) et plus (seuil de confiance 95 %).
Une classification approximative par densités a été utilisée pour plusieurs inventaires en Asie, Afrique et Amérique latine. L'amélioration de la précision signalée à propos de l'inventaire FAO à Sarawak est indiquée au tableau 1.
1. GRAPPE DE NEUF POINTS INVENTAIRE DE SARAWAK
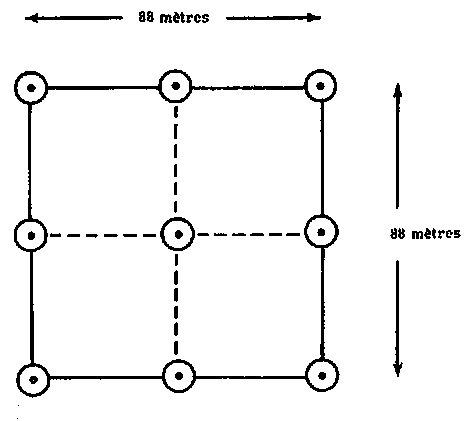
La mésovariation, intermédiaire entre la macrovariation et la microvariation, est un concept relatif qui dépend tant de la superficie à inventorier que de la forme et des dimensions de l'unité d'échantillonnage. Elle est intéressante pour étudier les variations entre les unités d'échantillonnage.
Dans les premières phases de l'inventaire forestier de Sarawak, une analyse de variance a été entreprise pour dégager le coefficient de variation dans les unités d'échantillonnage et entre celles-ci afin de déterminer le nombre optimal de placeaux dans la grappe. On disposait de données pour deux unités, la première et la troisième, couvrant une superficie de 176000 et de 225000 hectares respectivement. L'unité d'échantillonnage consistait en une grappe de 9 points (voir figure 1).
TABLEAU 2. - ANALYSE DE VARIANCE DU VOLUME TOTAL/ACRE1 TOUTES ESSENCES DE 18 INCHES (45 CM) ET PLUS DE DIAMÈTRE
|
Source de la variation |
Degré de liberté |
Somme des carrés |
Moyenne de la somme dés carrés |
Composantes de variance |
|
Entre les unités |
81 |
431448800 |
5326528,3 |
s b2 = 474223,86 |
|
Dans les unités |
656 |
694383300 |
1058511,1 |
s w2 = 1058511,10 |
1 Moyenne: 2031 ft3/acre (142 m3/ha).
TABLEAU 3. - INFLUENCE DE LA TAILLE ET DE LA FORME DU PLACEAU SUR LE COEFFICIENT DE VARIATION DU VOLUME TOTAL TOUTES ESSENCES
|
Type de forêt |
Volume moyen |
Coefficient de variation estimé |
Effectif de l'échantillon |
|||
|
Placeau carré |
Placeau rectangulaire |
|||||
|
0,3 ha |
0,6 ha |
10x300 m |
20x300 m |
|||
|
m3/ha |
...Pourcentage... |
|||||
|
Forêt humide |
393 |
98 |
82 |
75 |
48 |
14 |
|
Forêt décidue |
177 |
63 |
60 |
60 |
58 |
25 |
SOURCE: FAO, Inventaire des ressources forestières, Inde, 1970.
TABLEAU 4. - MOYENNE ET COEFFICIENT DE VARIATION DE LA SURFACE TERRIÈRE PAR HECTARE AU NIVEAU DES GROUPES ET DES ESSENCES POUR LES ARBRES DE 18 INCHES (45 CM) ET PLUS DE DIAMÈTRE
|
Groupes/Essences |
Surface terrière moyenne |
Coefficient de variation |
|
...m2/ha... |
...Pourcentage... |
|
|
Toutes essences |
14,70 |
27 |
|
Diptérocarpacées |
9,89 |
32 |
|
Merantis rouge |
4,10 |
40 |
|
Shorea parvifolia |
2,46 |
111 |
|
Shorea sagittata |
1,52 |
122 |
|
Shorea rubra |
0,95 |
142 |
|
Shorea macroptera |
0,35 |
187 |
|
Merantis jaune |
1,28 |
85 |
|
Shorea collaris |
1,14 |
234 |
|
Kapurs |
1,21 |
141 |
|
Dryobalanops oblongifolia |
2,37 |
202 |
|
Dryobalanops aromatica |
1,59 |
191 |
|
Keruings |
1,16 |
82 |
|
Dipterocarpus pachyphyllus |
0,086 |
270 |
|
Autres que diptérocarpacées |
4,68 |
39 |
|
Essences à faible densité |
2,12 |
50 |
|
Essences à densité moyenne |
1,23 |
67 |
|
Essences à densité élevée |
0,93 |
68 |
SOURCE: FAO, Inventaire forestier en Malaisie (Sarawak), 1973.
Un relevé fut effectué avec un relascope de BAF1 10 pieds carrés par acre (2,3 m2/ha). Il y avait de 50 à 100 échantillons de ce genre uniformément répartis sur les deux unités d'inventaire. Les résultats figurent au tableau 2.
1 BAF (basal area factor) = coefficient de surface terrière.
La valeur de R2, correspondant à s b2/(s b2 + s w2), s'est établie à 0,3, la variation entre échantillons est plus faible que dans un échantillon. L'auteur, en analysant les données de l'inventaire FAO pour la péninsule de Malaisie, a aussi observé une variation élevée à l'intérieur des différentes unités d'échantillonnage.
Pour la forêt humide de la vallée de l'Amazone, Ramos et al. ont trouvé pour R2 la valeur de 0,3 dans un cas et de 0,5 dans un autre. L'unité d'échantillonnage était composée de quatre rectangles très rapprochés, de 250 x 10 mètres, situés à 10 mètres du centre de la grappe et orientés orthogonalement.
Une analyse au niveau des groupes d'essences (31 groupes en tout) pour les forêts de Sarawak (unité 2) a montré une corrélation interne beaucoup plus faible, de l'ordre de 0,2 et moins, pour tous les groupes à l'exception d'un groupe n'appartenant pas aux diptérocarpacées. Pour ce dernier groupe on a trouvé, pour R2, 0,5.
Boon (1962) a étudié la variation de Eusideroxylon zwageri en divisant la surface à inventorier en blocs de dimensions variables: 2 x 1, 4 x 1, 4 x 2 kilomètres. Chaque bloc était ensuite partagé en deux blocs secondaires pour permettre de comparer la variance entre les blocs primaires et à l'intérieur de ceux-ci. L'analyse a montré que la variance entre blocs primaires n'est pas significativement différente de la variance à l'intérieur des blocs. On a constaté qu'il en allait de même pour tous les blocs et quelles que soient les dimensions du placeau.
Giudicelli et al. (1972) ont étudié la corrélation spatiale en ce qui concerne le nombre de pieds pour trois essences importantes et un groupe d'espèces dans une forêt de 1500 hectares au Cameroun en utilisant un placeau de 50 x 50 mètres. On trouvera à la figure 2 le corrélogramme qu'ils ont obtenu. Il montre une variabilité élevée pour de faibles distances pour toutes les essences. En outre, le corrélogramme est concave vers l'origine. En pareils cas, l'échantillonnage systématique doit être plus efficace que l'échantillonnage au hasard, ainsi que l'a constaté Cochran (1963).
Les résultats ci-dessus font apparaître une caractéristique importante de la forêt humide: une variabilité élevée sur de petites surfaces et une relative uniformité sur des superficies étendues. Cela indique que le recours à un type d'échantillonnage par grappes n'entraînera pas une grande perte de précision.
2. CORRÉLOGRAMME POUR 3 ESPÈCES ET 1 GROUPE DE 10 ESPÈCES DE LA FORÊT SEMI-SEMPERVIRENTE AU CAMEROUN
La complexité de la microvariation qu'un inventaire doit échantillonner dépend du degré où on veut pousser la ventilation par essence. Si les paramètres doivent être évalués seulement toutes essences groupées, la structure de variation est relativement simple.
Pour étudier l'influence de la taille et de la forme de l'unité d'échantillonnage sur la précision, un inventaire pilote a été réalisé en Inde. En tout, il y avait 39 unités réparties sur deux types de forêts, couvrant une superficie d'environ 80000 hectares. Les résultats ont montré que dans la forêt humide la forme et la taille influent beaucoup sur le coefficient de variation. En revanche, dans la forêt décidue voisine, la forme et la taille ont eu un faible effet sur la précision, comme le montre le tableau 3.
D'après des expériences menées dans la forêt humide de l'Indonésie, avec des placeaux rectangulaires de 20 mètres de largeur et de longueur variable, le coefficient de variation diminue de façon exponentielle quand les dimensions du placeau augmentent. Sa valeur tend vers 16 pour cent au-delà d'une dimension de 1,5 hectare (voir Nyyssönen, 1961).
Au niveau des essences, la disposition spatiale dans la forêt humide devient très compliquée. Comme l'a dit Richards, celle-ci apparaît d'abord comme un chaos végétal. Des recherches détaillées menées par plusieurs écologistes montrent cependant que la forêt humide, comme n'importe quelle autre communauté, a une structure organisée. Les plantes qui la constituent forment un nombre limité de synœcies avec un arrangement dans l'espace discernable, bien que complexe. Cet arrangement se retrouve pratiquement tel quel avec seulement une faible variation dans l'ensemble de la forêt humide.
Heinsdijk (1961) a étudié la distribution spatiale des essences dans la vallée de l'Amazone. Il a trouvé que la distribution de nombreuses essences se rapprochait courbe de Poisson, mais qu'il y avait souvent des exceptions là où les unités d'échantillonnage comprenaient un grand nombre d'espèces différentes, avec une tendance à la distribution contagieuse.
Utilisant les données d'inventaire en provenance de la vallée de l'Amazone, du Surinam et de Sarawak, l'auteur a entrepris une série de recherches. Pour les 10 à 20 espèces les plus importantes, le nombre moyen de pieds par unité d'échantillonnage (m) et son coefficient de variation (cv) ont été relevés, et la relation entre les deux paramètres a été étudiée.
Pour une distribution de Poisson proprement dite, les points représentant 100/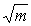 et cv devraient tomber sur une ligne droite inclinée à 45° par rapport à l'axe des X. Cela traduit une caractéristique importante de la distribution de Poisson, à savoir que la moyenne est égale à la variance. Dans tous les cas (voir figure 3), les espèces ont eu tendance à dévier vers le haut par rapport à la relation linéaire attendue, ce qui indique des tendances à l'agglomérat. Il est clair qu'en pareil cas l'expression 100/ n'est pas un indicateur très satisfaisant pour évaluer le coefficient de variation.
et cv devraient tomber sur une ligne droite inclinée à 45° par rapport à l'axe des X. Cela traduit une caractéristique importante de la distribution de Poisson, à savoir que la moyenne est égale à la variance. Dans tous les cas (voir figure 3), les espèces ont eu tendance à dévier vers le haut par rapport à la relation linéaire attendue, ce qui indique des tendances à l'agglomérat. Il est clair qu'en pareil cas l'expression 100/ n'est pas un indicateur très satisfaisant pour évaluer le coefficient de variation.
Les recherches de Boon (1962) sur Eusideroxylon zwageri montrent que cette essence a une tendance marquée à l'agglomérat.
Jack (1961) a étudié l'arrangement spatial des sept essences commerciales les plus importantes de la forêt semicaducifoliée du Ghana en utilisant des unités d'échantillonnage de tailles et de formes diverses. Pour Triplochiton scleroxylon, Khaya ivorensis, et Entandrophragma cylindricum, il a trouvé une différence significative par rapport à la distribution de Poisson et une bonne correspondance avec la distribution binomiale négative. Entandrophragma utile et Mimusops heckelii ont montré des tendances à l'agglomérat, mais cela n'a pu être observé qu'avec des dimensions de placeaux supérieures à 2 hectares, tandis que pour Chlorophora excelsa, la répartition varie, selon les dimensions du placeau, entre une distribution aléatoire et une distribution contagieuse.
Giudicelli et al. (1972), dans un type de forêt semblable au Cameroun, ont observé une distribution contagieuse prononcée pour Triplochiton scleroxylon et Terminalia superba, mais une distribution à peu près aléatoire dans le cas de Entandrophragma cylindricum. Ils ont également trouvé que le modèle de distribution a tendance à devenir régulier si l'on considère un ensemble de 10 essences importantes.
Quand la distribution n'obéit pas à la loi de Poisson, le travail se complique du fait que le modèle, comme l'ont révélé les chiffres observes, est partiellement modifié en fonction des dimensions et de la forme du placeau. Selon Greig-Smith (1954), de nombreux types de modèles non aléatoires semblent aléatoires quand les placeaux d'échantillonnage sont très petits ou très grands mais non aléatoires quand ils sont de taille intermédiaire. D'où la nécessité d'être prudent lorsqu'on tire des conclusions générales sur les caractéristiques de distribution des essences. Cependant, les considérations ci-dessus, ainsi que les études écologiques citées plus haut, montrent clairement que, dans la gamme de dimensions de placeaux utilisées, de nombreuses espèces dominantes de la forêt humide présentent une distribution contagieuse.
Dans un inventaire on s'intéresse moins aux structures; en tant que telles qu'à leurs implications pour l'établissement d'un plan de sondage. Pour les populations distribuées strictement au hasard (distribution de Poisson), le travail de planification est relativement simple. Dans ce cas, la précision de l'estimation est seulement fonction du nombre d'individus dénombrés. L'erreur type d'estimation sera la même, qu'on utilise beaucoup de petits placeaux, ou un petit nombre de grands, ou un seul placeau très vaste (Greig-Smith, 1964). La forme du placeau n'a pas d'influence. De même, la précision de l'inventaire sera pratiquement la même que les placeaux soient répartis au hasard ou systématiquement, qu'ils soient en grappes ou non.
La situation se complique pour les distributions contagieuses. Une conséquence a déjà été mentionnée, à savoir le manque de corrélation nette entre la densité des sujets et le coefficient de variation. En outre, les dimensions aussi bien que la forme de l'unité d'échantillonnage influent sur la précision des estimations. On observe couramment que, pour la même intensité d'échantillonnage, des placeaux petits mais dispersés donnent une précision plus grande qu'un petit nombre de gros placeaux compacts (Dawkins, 1952; Lanly et al., 1969; Greig-Smith, 1964).
3. RELATION ENTRE LE COEFFICIENT DE VARIATION (C.V.) ET 100/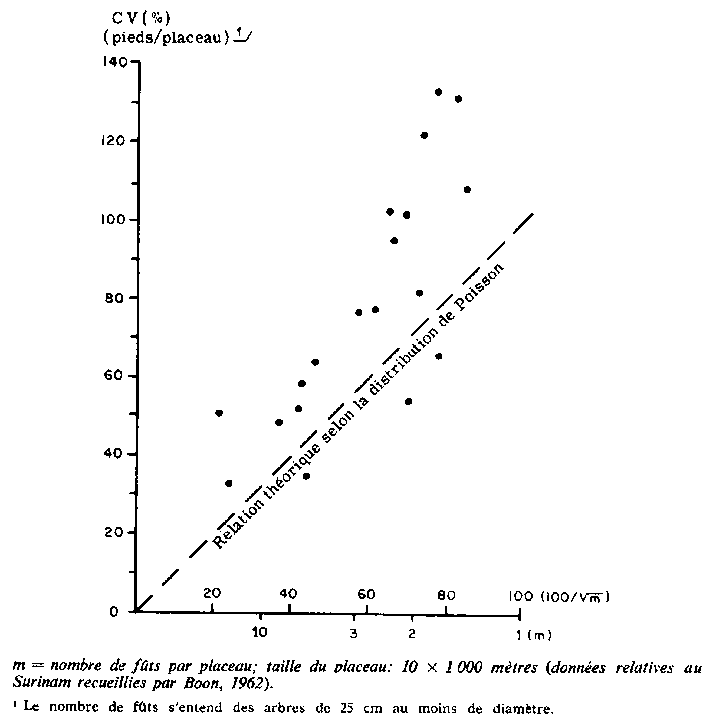
L'effet de l'augmentation des dimensions du placeau n'est pas le même pour toutes les essences. Ces différences peuvent être corrélées avec la disposition spatiale des essences. Triplochiton et Terminalia qui ont toutes deux une distribution nettement contagieuse montrent, relativement parlant, une plus grande perte de précision quand les dimensions du placeau augmentent que Entandrophragma utile, dont la disposition spatiale est proche de la distribution de Poisson (voir figure 2). Eusideroxylon zwageri et Khaya ivorensis, qui toutes deux font preuve d'une tendance à l'agrégat, se comportent d'une façon semblable.
S'agissant d'une distribution contagieuse, les unités d'échantillonnage de forme allongée permettent des estimations plus précises que celles de forme compacte de mêmes dimensions. On peut se représenter ce fait aisément: pour une même surface, il est plus probable que les premières plus que les secondes traversent à la fois des taches denses et des taches clairsemées.
Les recherches de Jack (1961) montrent que les unités longues et étroites, orientées en général perpendiculairement au sens du drainage, donnent des résultats plus précis que les unités relativement compactes. A l'occasion des études écologiques, des expériences semblables ont été décrites par de nombreux chercheurs, par exemple, Greig-Smith (1964) et Aubréville (1971).
Jusqu'ici, l'attention s'est concentrée sur l'étude de la distribution contagieuse, largement répandue dans les forêts humides. Il y a beaucoup d'autres possibilités d'arrangement des sujets dans l'espace. Un exemple en est la distribution régulière, dont il a été question plus haut pour la distribution des groupes de 10 essences. D'après les études décrites ci-dessus il apparaîtrait qu'un inventaire de forêt humide ventilé par essences serait probablement plus coûteux que pour toutes les essences ensemble. Une autre solution serait de s'intéresser au groupe d'essences. Un effet prévisible du groupement des essences est la diminution du coefficient de variation.
La moyenne et le coefficient de variation pour diverses essences et groupements d'essences pour les forêts mélangées de diptérocarpacées de Sarawak figurent au tableau 4. Celui-ci montre clairement qu'on réalise une économie dans l'échantillonnage en travaillant par groupes d'essences. Il y a beaucoup moins de variabilité au niveau du groupe qu'à celui des espèces. Le nombre d'unités d'échantillonnage vides diminue et la courbe de fréquence devient moins asymétrique. L'effet de la dimension et de la forme de l'unité d'échantillonnage est encore marqué, et comme on l'a noté plus haut, il continue à l'être même au niveau de toutes les essences.
AUBRÉVILLE, A., 1971, Regeneration patterns in the closed forests of Ivory Coast. Dans World vegetation types ed. by S.R. Eyre, p. 41. London, Macmillan.
BOON, D.A., 1962, Plot size and variability. Delft, International Training Centre for Aerial Survey. ITC publication, Series B, N°. 17.
COX, F., 1971, Dichtebestimmung und Strukturanalyse von Pflanzenpopulationen mit Hilfe von Abstandsmessungen. Mitt. BundForsch-Anst. Forst-u. Holzw., 87.
DAWKINS, H.C., 1952, Experiments in low percentage enumerations of tropical high forest with special reference to Uganda. Emp. for. rev. 31:131.
DAWKINS, H.C., 1958, The management of natural tropical high forest with special reference to Uganda, Oxford, Imperial Forestry Institute. Paper No. 34.
GIUDICELLI, X., LANLY, J.P., OUAKAM, J.B. & PIETRY, M., 1972, Application de la théorie des processus aléatoires a l'estimation de la précision d'un inventaire forestier par échantillonnage systématique. Annals sci. for., 30:267.
GREIG-SMITH, P., 1964, Quantitative plant ecology. London Butterworths.
HEINSDIJK, D., 1961, Inventaire forestier dans la vallée de l'Amazone. Unasylva, 15:167.
JACK, W.H., 1961, The spatial distribution of tree stems in a tropical high forest. Emp. for. rev. 40:234.
KEMPTHORNE, O., 1957, An introduction to genetic statistics. New York, Wiley.
LANLY, J.P. & VANNIÈRE, B., 1969, Précision d'un inventaire forestier en fonction de certaines de ses caractéristiques. Bois Forêts Trop. 152:35.
LOETSCH, F. & HALLER, K.E., 1973, Forest inventory. Vol. 1. München, BLV Verlagsgesellschaft.
LOETSCH, F., ZÖHRER, F. & HALLER, K.E., 1973, Forest inventory. Vol. 2. München, BLV Verlagsgesellschaft.
MATÉRN, B., 1960, Spatial variation. Meddn St. SkogsforskInst., 49(5).
MATÉRN, B., 1972, Poisson process in the plane and related models for clumping and heterogeneity. Symposium lecture on Analysis of spatial patterns and ecological relations.
MEYER, H.A., 1953, Forest mensuration. State College, Pa.
NYYSSÖNEN, A., 1961, Survey methods of tropical forests, Rome, FAO.
RAMOS, A.A., VIEIRA, A.N., VIVACQUA, C.A., ALENCAR, J.C., BARROS, J.C.M. & PÉLLICO NETTO, S., 1972, Inventario florestal do distrito agroperuario da zona Franca De Manus. Revta flor. 4(1).
RICHARDS, P.W., 1952, The tropical rain forest, p. 10, 249. Cambridge, University Press
RICHARDS, P.W., 1971, The structure of tropical rain forest. Dans World vegetation types, ed. by S.R. Eyre, p. 26. London, Macmillan.
SCHIMPER, A.F.W., 1903, Plant-geography upon a physiological basis. Tr. by W.R. Fischer. Ed. by P. Groom and I.P. Balfour. Oxford, Clarendon Press.
WECK, J. & WIEBECKE, C., 1961, Weltforstwirtschaft und Deutschlands Forst- und Holzwirtschaft. München.
BRÉSIL
Report to the Government of Brazil on the forest inventory in the Amazon valley. 1957, FAO/ETAP Report No. 601.
Report to the Government of Brazil on forest inventories in the Amazon. 1965, FAO/EPTA Report No. 2080.
GUYANE
Inventory of the Ebini-ltaki area. 1970, Guyana. Forest Industries Development Survey Technical Report No. 9. FO-UNDP/SF GUY/9.
INDE
Forest resource survey. 1970, India. Preinvestment Survey of Forest Resources. Technical Report No. 1. FO-UNDP/SF IND/23.
MALAISIE
A description of the data processing system used in the inventory of selected areas of the mixed dipterocarp forest in Sarawak. 1973, Malaysia. Forestry and Forest Industries Development Project. Working Paper No. 22. FO: DP/MAL/72/009.
The timber species of the mixed dipterocarp forest of Sarawak and their distribution. 1973, Malaysia. Forestry and Forest Industries Development Project. Working Paper No. 21. FO: DP/MAL/72/009
![]()
![]()
![]()
{kind=link}
{kind=link}