![]()
![]()
![]()
6.1 Effect of nutrition on heifer pubertal development
6.2 Effect of nutrition on the postpartum cow
6.3 The relationship between body condition and cow reproduction
6.4 Hormonal changes associated with undernutrition
6.5 Summary
6.6 References
Many studies show the influence of nutrition on cattle fertility. Differences in nutrition probably account for most variation in reproductive performance between herds and among animals within herds (Wiltbank et al, 1964; McDowell, 1972; Holness et al, 1978). Level of feeding (Wiltbank et al, 1962) and bodyweight (Ward, 1968; McClure, 1970; Lamond, 1970) affect cow fertility.
The effects of underfeeding are greatest on pre-pubertal animals and lactating cows. Weight loss postpartum, due to underfeeding or high lactation demands, extends the postpartum anoestrous period (Entwistle, 1983). Underfeeding also reduces milk yield, which reduces the growth of the calf. This reduces calf weaning weight and delays puberty, which reduces the potential lifetime productivity of the female calf.
The effects of poor nutrition differ depending on whether the main deficiency is in energy, protein, vitamins, minerals or trace elements. Under traditional management, usually more than one component is deficient (Roberts, 1971).
The effects of nutrition on cattle reproduction are covered extensively here because most cattle in the tropics are poorly fed and improving their feeding can immediately increase their reproductive performance. For more detail, consult Cunha et al (1967), Lamond (1970), McClure (1970), Preston and Willis (1974) and Topps (1976,1977).
The nutritional status of animals is difficult to measure, and this complicates interpretation of nutrition x reproduction interactions (Haresign, 1984). An animal's nutritional status is usually assessed on changes in its liveweight and body condition. However, these are long-term changes while many of the events of reproduction, e.g. ovulation, fertilization and placentation, take only a short time.
Poor nutrition delays puberty, reduces conception rate and increases pregnancy losses in heifers (Short and Bellows, 1971; Milagres et al, 1979; Fleck et al, 1980; Lemenager et al, 1980). Wiltbank et al (1966) referred to a critical age-to-weight ratio which must be reached before heifers attain puberty.
Drought delayed the onset of puberty in heifers of 10 breed groups in Australia and stopped ovarian activity in half of those that had already reached puberty (Post and Reich, 1980). Bartha (1971) found that feeding concentrates to Azaouak zebu cows advanced puberty and first conception by 4 to 18 months. Penzhorn (1975) found that puberty was delayed by 7 months in Africander heifers on a restricted diet. All heifers attained puberty at about the same bodyweight (279-295 kg) but at different ages. Conception rates after a 3-month breeding period were 80, 93, 87 and 40% for heifers on high, medium, low and restricted levels of nutrition, respectively. These data also indicate that overfeeding can reduce reproductive performance. Reid et al (1964) also found that heifers reared on a very high level of nutrition had more breeding problems subsequently than those fed moderately.
Supplementation need not be continuous. In Ethiopia, Azage Tegegne and his co-workers (ILCA, unpublished data) supplemented Boran heifers with 1.5 kg of concentrate (1/3 oilseed cake, 2/3 wheat bran) for 90 days during the dry season preceding puberty. Supplemented heifers reached puberty earlier (596.4 vs 633.5 days), were heavier at puberty (230.7 vs 202.4 kg) and had larger ovaries than unsupplemented heifers. The proportion of supplemented heifers bred in the subsequent breeding season (61.9%) was nearly twice that of unsupplemented heifers (33.3%). A similar trend was observed among Boran x Friesian heifers. These results agree with those of Olivares et al (1981) in Brahman heifers (Table 27).
Table 27. Effect of prepartum and postpartum supplementation on the reproductive performance of Brahman heifers
|
|
I |
II |
III |
IV |
|
Prepartum cow weight loss (g/day) |
770 |
737 |
1362 |
1366 |
|
Postpartum weight loss to 120 days (g/day) |
41 |
81 |
163 |
162 |
|
% cows cycling |
67 |
58 |
25 |
8 |
|
% cows conceiving |
50 |
25 |
8 |
8 |
|
Calf weight at birth (kg) |
27 |
29 |
28 |
29 |
|
Calf weight at 120 days (kg) |
104 |
102 |
119 |
108 |
I = Fed 2 kg concentrates for 22-55 days prepartum and 90 days postpartum
II = Fed concentrates prepartum only
III = Fed concentrates postpartum only
IV = Received no concentrates (control)
Source: Olivares et al (1981).
Cohen et al (1980) found a direct relationship between bodyweight and incidence of oestrus in 603 heifers aged 17.2 to 19.2 months and weighing between 130 and 376 kg. The relationship between the percentage of heifers showing oestrus (Y. expressed as probit units) for a given weight (X) was:
Y = - 40.69 + 19.34 log10 X
The authors estimated that 5% of the heifers would show oestrus when weighing 187 kg or less, 50% would show oestrus when weighing 231 kg or less, and 95% would show oestrus when or before they weighed 280 kg.
The reproductive performance of the postpartum cow is related to nutritional status (Dunn et al, 1969; van Niekerk, 1982). Cows fed a high energy diet after calving conceive sooner than those with a lower energy intake (Wiltbank et al, 1962, 1964; Dunn et al, 1969; Hill et al, 1970). Although protein is generally regarded as less important than energy for reproduction, low protein intake can also cause infertility. However, it may be´ difficult to differentiate the effects of low protein intake from concurrent low energy intake, because protein deficiency usually leads to decreased appetite.
Cattle in the tropics are usually dependent on natural pastures and crop byproducts for feed. The crude protein content of the feed is often below 7.5%, which reduces rumen efficiency and reduces the true digestibility of the feed. As a result, lactating cows are unable to meet their nutritional requirements and lose weight and condition during lactation. This prolongs the lactation anoestrous period, and cows tend to calve in alternate years (Ward, 1968). The percentage change in the cow's bodyweight during the first 2 weeks after calving is inversely related to the number of days to first ovulation (Stevenson and Britt, 1980; Butler et al, 1981).
High levels of feeding before calving reduced the postpartum anoestrous period in taurine cows (Bellows and Short, 1978). In addition, more cows exhibited oestrus before the breeding season and subsequent pregnancy rates were increased. King (1968) estimated that a 1% change in body weight resulted in a 1% change in first service conception rate. Similar results have been found in zebu cattle.
Feeding a high plane of nutrition to five anoestrous cows of each of four breeds for 45 days resulted in 65% resuming cycling and 55% ovulating, whereas the 20 cows kept on a low plane diet neither cycled nor ovulated (Dindorkar et al, 1982).
In Zambia, feeding zebu cows a sub-maintenance diet resulted in 55% of the animals stopping cycling within a year, whereas those on a maintenance diet continued to cycle normally (Rakha and Igboeli, 1971). The cows on the sub-maintenance diet also had a higher incidence of silent heats than the maintenance-fed cows. Two out of three oestrous cycles may be silent in underfed animals (Hale, 1974).
The growth and development of the foetus, parturition, lactation and involution of the uterus, all use energy. The energy used by these processes must be supplied to the cow if she is to rebreed soon after calving. Generally, the farmer will not be able to meet the cow's whole energy needs, and some will be met from body reserves or fat. Thus a cow in good condition is better able to meet the energy requirements of parturition, lactation and involution of the uterus, and will therefore rebreed sooner, than a cow in poor condition. Cows should be fed well for 22-55 days before parturition and, if possible, for 90 days after parturition (Olivares et al, 1981) (Table 27).
Improved pastures can also be used to improve cow nutrition. In Colombia, cows grazing improved pasture and supplemented with legume fodder had a conception rate of up to 64.4%, compared with 6.3% for cows grazing only unimproved pasture (Kleinhesterkamp et al, 1981). In Florida, beef cows grazing mixed clover/grass pastures at 0.55 ha per cow had higher conception rates than cows grazing pure grass pastures at 0.83 ha per cow (Warnick, 1976) (84 vs 48% in lactating cows): they also had higher wearing rates (84 vs 64%), a shorter postpartum anoestrous period (72 vs 90 days) and required fewer services to conceive (1.34 vs 1.40). Most lactating cows grazing the grass pasture did not cycle during the 90-day breeding season.
Hale (1975) found that underfed dry zebu cows stopped cycling when their weight fell to 320 kg from 390 kg. However, when the cows regained weight, they did not start cycling again until they were significantly heavier than the weight at which they stopped cycling.
Mukasa-Mugerwa et al (1989) estimated that traditionally raised zebu cattle in the Ethiopian highlands needed 8 months after they stopped lactating to attain a bodyweight and condition that allowed them to reconceive successfully. The average calving interval was 26 months despite a lactation length of only 8 months. Fertility of grazing cows is therefore closely related to the liveweight change during the calving-to-service interval. The animal is likely to become sexually active only after it has regained much of its pre-calving weight.
Patil and Deshpande (1981) found that Gir cows that gained weight in the first three months after parturition showed heat during that period, while those that lost weight remained anoestrous. Cows that lost weight had lower blood glucose and serum protein concentrations than cows that gained weight, and the authors suggested that measurements of these could be used to indicate cows that might not show oestrus soon after parturition.
McClure (1968) found that cows with a blood glucose concentration of less than 30 mg glucose per 100 ml blood tended to return to service. Cows must, therefore, be on an adequate or rising plane of nutrition and gaining mass during the mating season if conception is to be successful (van Niekerk, 1982).
In Zimbabwe, Richardson et al (1975) found that a cow's ability to reconceive was a function of its final change in bodyweight at mating time, but was not related to its rate of bodyweight change from calving to midway through the mating season (Table 28). Similar results were reported by Ward (1968), Trail et al (1971), Steenkamp et al (1975), Meaker (1975) and Buck et al (1976).
Table 28. Change in bodyweight as percentage of initial weight of cows and their subsequent conception rates
|
Bodyweight 1 change (% ) |
Number of cows |
Calving rate (%) |
Bodyweight 2 change (%) |
Number of cows |
Calving rate (%) |
|
-24 |
11 |
55 |
|
|
|
|
-20 |
13 |
46 |
-20 |
4 |
25 |
|
-16 |
28 |
82 |
-16 |
6 |
67 |
|
-12 |
32 |
82 |
-12 |
11 |
64 |
|
-8 |
32 |
91 |
-8 |
29 |
69 |
|
-4 |
23 |
87 |
4 |
36 |
78 |
|
0 to +8 |
31 |
84 |
0 to +20 |
87 |
90 |
1 Bodyweight change between peak weight in early pregnancy and parturition.
2 Bodyweight change between peak weight in early pregnancy and the following mating season.
Source: Richardson et al (1975).
In Botswana, Buck et al (1976) found average conception rates of 50% for cows weighing less than 300 kg at the beginning of the breeding season, 85% for cows weighing 430 kg, 67% for cows that lost weight over the breeding period and 76% for cows that gained 20 kg weight over this period.
Ward (1968) suggested that every cow has an optimum bodyweight for conception, the so-called "target" or "critical" bodyweight. Animals weighing less than this are less able to reproduce. Wiltbank et al (1964) added that breeding cows must be improving in "condition" during the mating period. This is emphasised in the work of Wiltbank (1977) (Table 29) and Haresign (1984).
Table 29. Effect of cow body condition at calving on the cumulative percentage return to oestrus
|
Body condition |
Cumulative percentage return to oestrus |
|||||
|
Days after calving |
||||||
|
40 |
50 |
60 |
70 |
80 |
90 |
|
|
Thin |
19 |
34 |
46 |
55 |
62 |
66 |
|
Moderate |
21 |
45 |
61 |
79 |
88 |
92 |
|
Good |
31 |
42 |
91 |
96 |
98 |
100 |
Source: Wiltbank (1977).
Condition scoring was started in Australia for assessing sheep fatness: it was introduced into the United Kingdom for the same purpose, and has since been extended to cattle. Condition scoring is a subjective visual assessment of animals, but with practice a high level of repeatability, both between measurements and between scorers, can be obtained (Nicholson and Butterworth, 1986). A condition score is based on the amount of fat and muscle tissue covering the skeletal frame and is indicative of the animal's nutritional status (van Niekerk, 1982).
The relationship between condition score and body fatness has been established from data on cows slaughtered at different body condition scores (Wright and Russell, 1984). Although the condition score gives a good indication of fatness, breeds differ in the way they deposit fat reserves. This is especially true of cows with more than 15% body fat. Dairy cattle generally deposit more fat internally than do beef cattle. Condition scoring tends to assess subcutaneous fat reserves, and therefore at a given condition score value dairy cows tend to have more fat reserves than beef cows.
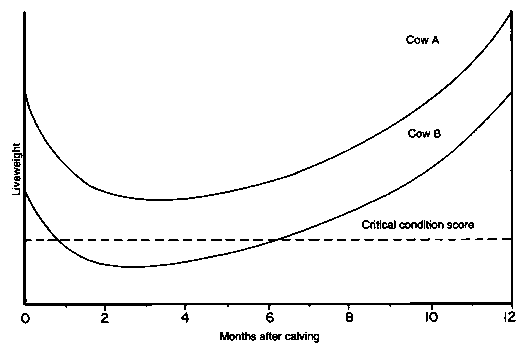
Changes in bodyweight or condition score of cows indicate likely levels of subsequent reproductive performance. The fertility of cows in poor condition is low. Cows below the "critical condition score» are least likely to reproduce (Figure 13).
Steenkamp et al (1975) compared the conception rates of cows of similar weight that differed in condition score and found that condition at mating was more important than weight. This agrees with the findings of van Niekerk (1982), who observed a calving rate of 78% for cows in optimum condition compared with just 8% for animals in the poorest condition (Table 30). The feed costs of maintaining the animals in this better condition are more than covered by increased reproductive performance.
Thus animals should be fed well to promote good reproductive performance. It is more efficient to feed animals to maintain good body condition than to allow them to lose weight in the hope that it can be regained before the mating season. Wright (1985) estimated that the loss of one unit in condition score would supply 3200 MJ of metabolisable energy: restoring the animal's condition score would require about 6500 MJ of dietary metabolisable energy. This agrees with van Niekerk (1982) who concluded that the feed cost of maintaining a cow at a condition score of 3.0 was half that required to raise a cow's condition from 1.5 to 3.0.
The benefits of feeding animals well in terms of better reproductive performance are often easily appreciated by peasant farmers. However, smallholders usually have only small supplies of supplementary feed and will need advice on which animals to feed it to, how much to feed and when.
Figure 13. Pattern of liveweight change during the breeding cycle of the cow and its possible relationship to nutritionally induced infertility in the postpartum period
Remarks:
Negative energy balance itself is not likely to have much effect on the length of the postpartum period as long as the cow is in good condition at calving (Cow A) and its condition remains above the critical condition score, which is defined as that score below which cows still in negative energy balance (Cow B) are likely to suffer from extended anoestrous periods and silent heats.Source: Haresign (1984).
Table 30. Effect of condition score at mating on calving rate of cows
|
Condition score at mating |
n |
Number of calves born per 100 cows mated |
|
1.5 |
12 |
8 |
|
2.0 |
72 |
43 |
|
2.5 |
124 |
64 |
|
3.0 |
211 |
78 |
|
3.5 |
31 |
77 |
Source: van Niekerk (1982).
Few studies have been made on the relationship between bodyweight, condition and hormone synthesis or secretion in zebu cattle, and their results are inconsistent. However, in general the results suggest that poor feeding postpartum reduces luteal function and responsiveness of the ovaries to luteinising hormone (Gombe and Hansel, 1973; Martinez et al, 1984; Rutter and Randel, 1984; Whisnant et al, 1985).
Inadequate nutrition delays puberty and sexual maturity in heifers and resumption of ovarian activity and oestrus in postpartum cows. If a cow is underfed when pregnant it will be in poor condition at calving, and will be slow to resume cycling and reconceive. Ideally, the cow's body condition should improve gradually through pregnancy, but excessive fatness should be avoided.
Cows will probably lose weight after calving, but weight loss should be minimised through good feeding to allow them to start cycling again as soon as possible.
More studies are needed to determine the physiological basis of the nutrition/reproduction interaction in zebu cattle.
Bartha R. 1971. Studien zu Fragen der Zebu-Rinderzucht in den Tropen. Weltforum Verlag, Munich, Federal Republic of Germany. 172 pp.
Bellows R A and Short R E. 1978. Effect of precalving feed level on birthweight, calving difficulty and subsequent fertility. Journal of Animal science 46: 1522-1528.
Buck N G. Light D, Rutherford A, Miller M, Prachett T W. Capper B and Trail J C M. 1976. Environmental factors affecting beef cow reproductive performance in Botswana. Animal Production 23: 357-363.
Butler W R. Everett R W and Coppock C E. 1981. The relationships between energy balance, milk production and ovulation in postpartum Holstein cows. Journal of Animal Science 53: 742.
Cohen R D H. Garden D L and Langlands J P. 1980. A note on the relationship between liveweight and the incidence of oestrus in Hereford heifers. Animal Production 31: 221-222.
Cunha T J, Warnick A C and Koger M. 1967. Factors affecting calf crop. University of Florida Press, Gainesville, Florida, USA. 376 pp.
Dindorkar C V, Muzaffar S J and Kaikini A S. 1982. Note on the effect of feeding extra energy diets to anoestrous cattle. Indian Journal of Animal Sciences 52: 1228-1229.
Dunn T G. Ingalls J E, Zimmerman D R and Wiltbank J N. 1969. Reproductive performance of 2-year old Hereford and Angus heifers as influenced by pre- and post-calving energy intake. Journal of Animal Science 29: 7l9-726.
Entwistle K W. 1983. Factors influencing reproduction in beef cattle in Australia. AMRC Reviews No. 43. Australian Meat Research Committee, Sydney, NSW, Australia. 30 pp.
Fleck A T. Schalles R R and Kiracofe G H. 1980. Effect of growth rate through 30 months on reproduction performance of beef heifers. Journal of Animal Science 51: 86.
Gombe S and Hansel W. 1973. Plasma LH and progesterone levels in heifers on restricted energy intakes. Journal of Animal Science 37: 728-733.
Hale D H. 1974. Studies on the level of LH and prolactin in serum of cycling and anoestrous cows. South African Journal of Animal Science 4: 51.
Hale D H. 1975. Nutrition, hormones and fertility. Rhodesia Agricultural Journal 72: 69.
Haresign W. 1984. Underfeeding and reproduction: Physiological mechanisms. In: C Chemineau, D Gauthier and J Thimonier (eds), Reproduction des ruminants en zone tropicale. INRA Publication No. 20. INRA (Institut national de la recherche agronomique), Paris, France. pp. 339-365.
Hill J R. Lamond D R. Henricks D M, Dickey J F and Niswender G D. 1970. The effects of undernutrition on ovarian function and fertility in beef heifers. Biology of Reproduction 2: 78-84.
Holness D H. Hopley J D H and Hale D H. 1978. The effect of plane of nutrition, live weight, temporary weaning and breed on the occurrence of oestrus in beef cows during the postpartum period. Animal Production 26: 47-54.
King J O L. 1968. The relationship between conception rate and changes in body weight, yield and S.N.F. content of milk in dairy cows. Veterinary Record 83: 492.
Kleinhesterkamp I, Botero R. Muller F. Amezquita M C and Cabrales R D. 1981. Conception rates in lactating cows on the eastern plains of Colombia. Memoria, Asociacion Latinoamericana de Produccion Animal 16: 121 (Animal Breeding Abstracts 52: 6492).
Lamond D R. 1970. Nutrient status in relation to reproduction. Journal of Animal Science 30: 322.
Lemenager R P. Smith W H. Martin T G. Singleton W L and Hodges J R. 1980. Effects of winter and summer energy levels on heifer growth and reproductive performance. Journal of Animal Science 51: 83 7.
Martinez N D, Lopez S R and Combellas J. 1984. Effect of postpartum nutrition on blood level of progesterone in dairy cows. In: C Chemineau, D Gauthier and J Thimonier (eds), Reproduction des ruminants en zone tropicale. INRA Publication No. 20. INRA (Institut national de la recherche agronomique), Paris, France. pp. 367-377.
McClure T J. 1968. Hypoglycaemia, an apparent cause of infertility of lactating cows. British Veterinary Journal 124: 126-130.
McClure T J. 1970. An experimental study of the causes of a nutritional and a lactational stress infertility of pasture-fed cows, associated with loss of body weight at about the time of mating. Research in Veterinary Science 11: 247254.
McDowell R E. 1972. Improvement of livestock production in warm climates. W H Freeman, San Francisco, California, USA. 711 pp.
Meaker H J. 1975. Relationship between body mass and conception in beef cattle. South African Journal of Animal Science 5: 45.
Milagres J C, Dillard E U and Robinson O W. 1979. Influence of age and early growth on reproductive performance of yearling Hereford heifers. Journal of Animal Science 48: 1089.
Mukasa-Mugerwa E, Ephraim Bekele and Tadesse Tessema. 1989. Type and productivity of indigenous cattle in central Ethiopia. Tropical Animal Health and Production (in press).
Nicholson M J and Butterworth M H. 1986. A guide to condition scoring of zebu cattle. International Livestock Centre for Africa, Addis Ababa, Ethiopia. 29 pp.
van Niekerk A. 1982. The effect of body condition as influenced by winter nutrition, on the reproductive performance of the beef cow. South African Journal of Animal Science 12: 383-387.
Olivares R. Chicco C, Redrigvez C F. Hernandez R and Verde O. 1981. Prepartum and postpartum supplements for Brahman heifers. Memoria, Asociacion Latinoamericana de Produccion Animal 16: 48 (Animal Breeding Abstracts 52: 6521).
Patil J S and Deshpande B R. 1981. Study of bodyweight changes during antepartum, parturition and postpartum periods in Gir cows with special reference to exhibition of postpartum actions. Indian Veterinary Journal 58: 376379.
Penzhorn E J. 1975. Wintering levels and reproduction in Afrikander heifers. Agroanimalia 7: 49-59 (Animal Breeding Abstracts 45: 3777).
Post T B and Reich M M. 1980. Puberty in tropical breeds of heifers as monitored by plasma progesterone. Proceedings of the Australian Society of Animal Production 13: 61-62.
Preston T R and Willis M B. 1974. Intensive beef production. Pergamon Press, New York, USA. 566 pp.
Rakha A M and Igboeli G. 1971. Effects of nutrition, season and age on the estrous cycle of indigenous Central African cattle. Journal of Animal Science 32: 943-945.
Reid J T. Loosli J K, Trimberger G W. Turk K L, Asdell S A and Smith S E. 1964. Causes and prevention of reproductive failures in dairy cattle. IV. Effect of plane of nutrition during early life on growth, reproduction, health and longevity of Holstein cows. 1. Birth to fifth calving. Cornell University Agricultural Experiment Station Bulletin 987. Ithaca, New York, USA.
Richardson F D, Oliver J and Clarke G P Y. 1975. Analyses of some factors which affect the productivity of beef cows and of their calves in marginal rainfall areas of Rhodesia. I. Factors affecting calving rate. Animal Production 21: 41-49.
Roberts S J. 1971. Veterinary obstetrics and genital diseases. 2nd edition. Published by the author. Distributed by Edwards Brothers, Ann Arbor, Michigan, USA. 776 pp.
Rutter L M and Randel R D. 1984. Postpartum nutrient intake and body condition: Effect on pituitary function and onset of estrus in beef cattle. Journal of Animal Science 58: 265-274.
Short R E and Bellows R A. 1971. Relationships among weight gains, age at puberty and reproductive performance in heifers. Journal of Animal Science 32: 127-131.
Steenkamp J D G. van der Horst C and Andrew M J A. 1975. Reconception in grade and pedigree Afrikander cows of different size: Postpartum factors affecting reconception. South African Journal of Animal Science 5: 103-110.
Stevenson J S and Britt J H. 1980. Models for the prediction of days to first ovulation based on changes in endocrine and non-endocrine traits during the first two weeks postpartum in Holstein cows. Journal of Animal Science 50: 103.
Topps H J. 1976. Effect of energy and protein deprivation on the performance of beef cattle. In: A J Smith (ed), Beef cattle production in developing countries. Proceedings of the Conference held in Edinburgh, 1-6 September 1974. University of Edinburgh, Centre for Tropical Veterinary Medicine, Edinburgh, UK. pp. 204-215.
Topps H J. 1977. The relationship between reproduction and undernutrition in beef cattle. World Review of Animal Production 10: 43-49.
Trail J C M, Sacker G O and Fisher I L. 1971. Crossbreeding of beef cattle in Western Uganda. I. Performance of Ankole, Boran and zebu cows. Animal Production 13: 127-141.
Ward H K. 1968. Supplementation of beef cows grazing on veld. Rhodesian Journal of Agricultural Research 6: 93-101.
Warnick A C. 1976. Management factors affecting reproductive efficiency of beef cattle in developing countries. In: A J Smith (ed), Beef cattle production in developing countries. Proceedings of the Conference held in Edinburgh, 1-6 September 1974. University of Edinburgh, Centre for Tropical Veterinary Medicine, Edinburgh, UK. pp. 132-137.
Whisnant C S. Kiser T E, Thompson F N and Hall J B. 1985. Effect of nutrition on the LH response to calf removal and GnRH. Theriogenology 24: 565-573.
Wiltbank J N. 1977. Effect of level of nutrition on growth and reproduction of beef females. Georgia Nutrition Conference, 16-18 February 1977. pp. 138-146.
Wiltbank J N. Rowden W W, Ingalls J E, Gregory K E and Koch R M. 1962. Effect of energy level on reproductive phenomena of mature Hereford cows. Journal of Animal Science 21: 219-225.
Wiltbank J N. Rowden W W. Ingalls J E and Zimmerman D R. 1964. Influence of postpartum energy levels on reproductive performance of Hereford cows restricted in energy intake prior to calving. Journal of Animal Science 23: 1049-1053.
Wiltbank J N. Gregory K E, Swiger L A, Ingalls J E, Rothlisberger J A and Koch R M. 1966. Effect of heterosis on age and weight at puberty in beef heifers. Journal of Animal Science 25: 744-751.
Wright I A. 1985. The relationship between body condition, nutrition, and performance of beef cows. In: F H Baker (ed), Emerging technology and management for ruminants. Winrock International, Arkansas, USA. pp. 110-118.
Wright I A and Russell A J F. 1984. Partition of fat, body composition and body condition score in mature cows. Animal Production 38: 23.
![]()
![]()
![]()