![]()
![]()
![]()
3.1 Ecologie et répartition des adultes
3.2 Comportement et rendement
3.3 Reproduction et migration larvaire
3.4 Migration des juvéniles
3.5 Recrutement et maturité sexuelle
3.6 Croissance
3.7 Mortalité
Elle a été étudiée de façon approfondie par Garcia (1976) en Côte d’Ivoire. Comme le schéma du cycle vital qui a été observé paraît très général, un résumé de ces travaux et principaux résultats (Garcia 1978, sous presse) sera repris ici.
Tableau 1
SUPERFICIE DES FONDS A CREVETTE DANS LE SECTEUR COTE D’IVOIRE/ANGOLA
|
PAYS |
STOCKS |
SUPERFICIE EN MILLES |
|
COTE D’IVOIRE
|
Tabou - Sassandra |
250 |
|
Grand Lahou |
80 |
|
|
Bassam |
60 |
|
|
GHANA
|
Axim - Takoradi |
120 |
|
Ada-Keta |
80 |
|
|
TOGO |
|
90 (max.) |
|
BENIN |
|
130 |
|
NIGERIA |
Lagos |
13001 |
|
CAMEROUN |
|
190 |
|
GUINEE EQUATORIALE |
|
? |
|
GABON |
Cap Esterias |
200 |
|
CONGO |
|
Pas de contentration exploitable |
|
TOTAL |
|
2 500 ou 3 700 |
1 Burukovski et Bulenkov (1969).
2 Raitt et Niven (1969).
L’espèce se rencontre du cap Hatteras, sur la côte nord-américaine, au cap Frio, sur la côte est-africaine (Perez Farfante, 1969). De chaque côté de l’Atlantique, Penaeus duorarum occupe une aire géographique dans laquelle les eaux de surface atteignent au moins 24°C dans l’année et descendent rarement au-dessous de 18°C. Ce préférendum pourrait être lié à des exigences métaboliques concernant notamment les mécanismes de reproduction.
A l’intérieur de ces limites thermiques la distribution des adultes est conditionnée par des salinités supérieures à 35‰. Dans les zones hydrologiquement convenables, la répartition est liée à la présence de lagunes ou d’estuaires et à la nature du sédiment dont la teneur en particules fines (£ 50m) doit dépasser 25 pour cent. L’une des caractéristiques importantes des fonds à crevette est leur grande richesse en matière organique. La répartition bathymétrique de l’espèce est liée à la structure du sédiment: les concentrations intéressantes se rencontrent là où la teneur en particules fines dépasse 50 pour cent. La limite inférieure de la distribution, vers 60-65 m, est vraisemblablement liée aux conditions hydrologiques.
Dans les régions où existent des oscillations de la thermocline, la distribution bathymétrique varie saisonnièrement au rythme de ces oscillations. Les déplacements tendent à maintenir l’espèce à la partie supérieure de la couche de discontinuité, entre 23-25°C d’une part, et entre 35-35,5‰ d’autre part. En saison d’upwelling, ces eaux disparaissent du plateau continental et les crevettes sont alors menées, dans certaines zones, à supporter des conditions moins favorables (16-18°C).
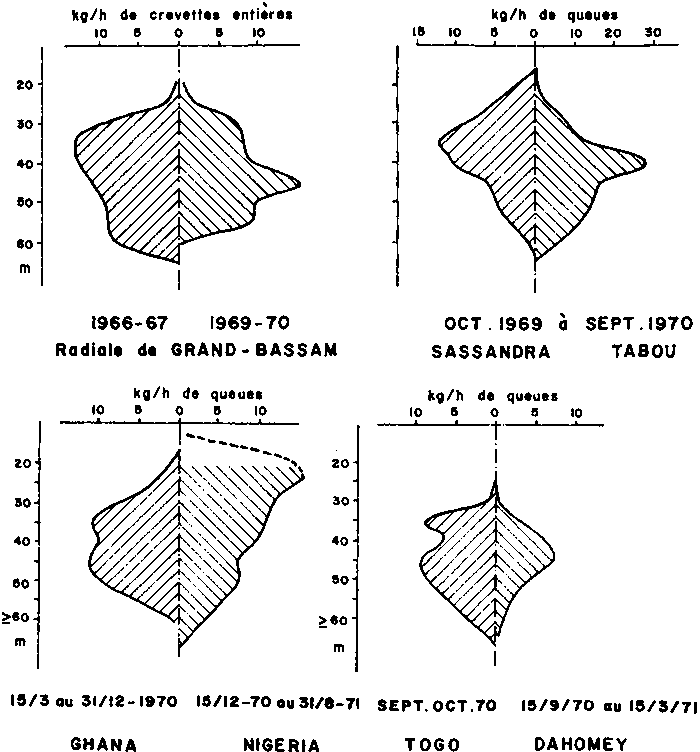
Les distributions bathymétriqnes de tailles et de sexes sont très caractéristiques (Figure 1) et différentes de celles que l’on observe dans le golfe du Mexique. Les individus de tailles petite et moyenne occupent les immersions centrales (30-45m) de l’aire de répartition. Ce sont les plus abondants et leur sex-ratio est proche de la normale. La taille moyenne augmente et l’abondance diminue vers le large d’une part (où les mâles dominent) et vers la côte d’autre part (où les femelles l’emportent). La distribution bathymétrique des sexes change saisonnièrement, vraisemblablement en relation avec la reproduction. Devant le delta du Niger, en revanche, l’abondance est plus élevée aux plus faibles profondeurs exploitées (vers 20-25 m).
Sur la côte d’Afrique, les rendements les plus élevés s’obtiennent, soit le jour, soit la nuit, soit indifféremment. L’obtention de rendements maximaux de jour est extrêmement fréquente en Côte d’Ivoire, au Nigéria et au Sénégal (Garcia, 1977; Raitt et Niven, 1966). Au Gabon, les meilleurs rendements sont obtenus de nuit (Fontana et Bâ, 1972).
Ce comportement apparent dépend de la zone, de la période et de la profondeur considérées. Il est admis que l’activité est essentiellement nocturne et que le comportement apparent n’est que la manifestation de modifications nycthémérales de la vulnérabilité des crevettes. Ces modifications liées à la turbidité des eaux résulteraient de migrations verticales mettant périodiquement les crevettes hors de portée des chaluts (hypothèse avancée par Le Guen et Crosnier, 1968, au Congo). Fontana et Bâ (1972) notent que le chalutage contre le vent permet des rendements de 25 pour cent plus élevés que lorsque le chalutage s’effectue vent arrière.
En Côte d’Ivoire, la proportion de femelles mûres dans les captures augmente dès octobre quand les eaux se réchauffent après la saison froide. Elle passe par des valeurs élevées d’octobre à décembre et décroît ensuite de façon irrégulière jusqu’en mai-juin où apparaît parfois un maximum secondaire.
L’abondance des postlarves en surface, à l’entrée des lagunes suit des rythmes nycthéméraux, lunaires et saisonniers. Les larves ne sont présentes que la nuit. La courbe d’abondance nocturne est unimodale ou bimodale suivant la phase lunaire, en liaison avec le cycle des marées. L’abondance globale dépend de la phase lunaire; elle est maximale en nouvelle lune, minimale en pleine lune et intermédiaire aux quartiers. Ces variations reflètent, soit un rythme lunaire de ponte, soit plus vraisemblablement un rythme de marée avec interaction de la luminosité lunaire, entraînant une baisse d’abondance des larves en surface lors de la pleine lune.
La taille des postlarves à stade de développement égal, varie saisonnièrement avec la richesse planctonique et la température (relation inverse).
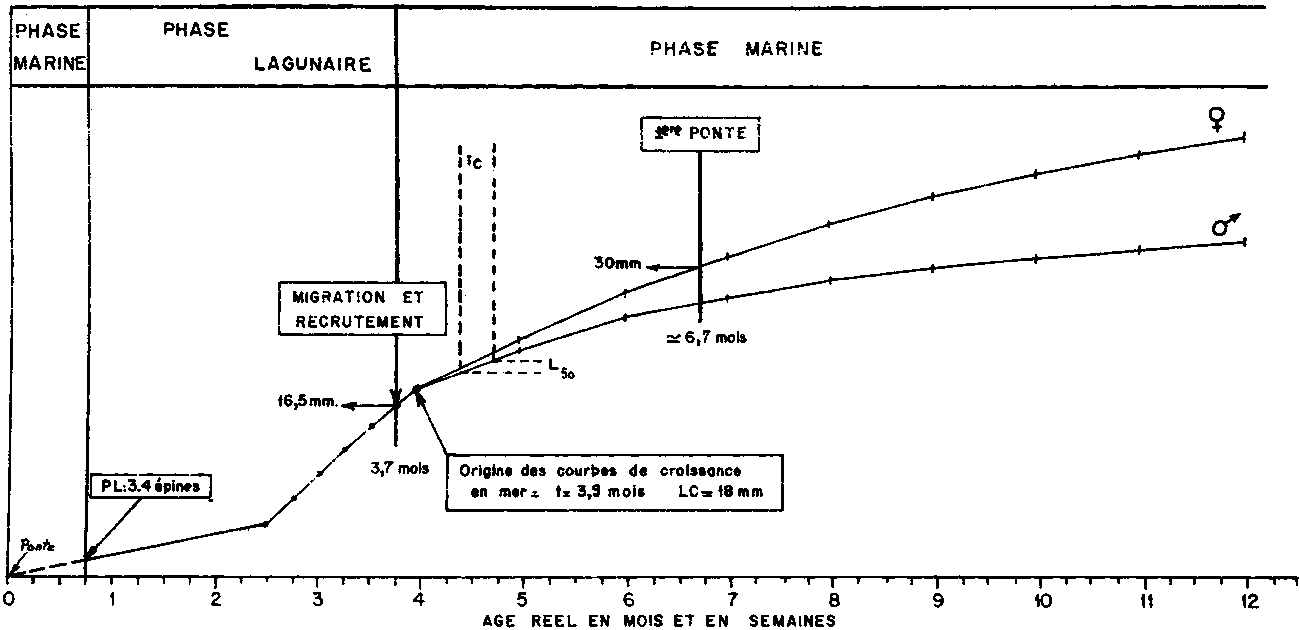
Elle a lieu à l’âge de 3-4 mois après un séjour en lagune de 2,5 à 3 mois. Il existe un rythme nycthéméral et tidal: la migration a lieu la nuit à marée descendante. Les variations saisonnières d’amplitude annuelle sont liées de façon évidente au. cycle de la reproduction et aux facteurs externes qui la gouvernent. Des pics de migration se produisent au moment des crues quand la salinité est faible et les courants rapides. Les captures les plus importantes s’obtiennent de novembre à mai en lagune Ebrié (Côte d’Ivoire), de janvier à mai en lagune Tagba (Côte d’Ivoire) et de février à mai dans la lagune de Lagos (Nigéria) (Obakin, 1969 et 1970).
Les limites de distribution des juvéniles en lagune sont celles de l’influence marine (courants de marée, onde saline). La taille de migration varie saisonnièrement. Elle dépend des régions. Elle est de 16-17 mm (longueur céphalothoracique) en Côte d’Ivoire. Elle est en moyenne légèrement plus faible au Nigéria (13 - 15 mm; Obakin, 1969 et 1970). La taille de migration varie également, d’un secteur à l’autre d’une lagune, et d’une lagune à l’autre, en liaison avec la salinité ambiante. Elle varie aussi d’une année à l’autre dans une même lagune en fonction de la pluviosité annuelle, donc également de la salinité ambiante. Cette action de la salinité sur la taille de migration se ferait par l’intermédiaire d’un raccoursissement de la durée du séjour en lagune quand la salinité devient faible.
En Côte d’Ivoire le recrutement en mer, estimé à partir du pourcentage d’individus de petite taille (LC inférieure à 22 mm) dans les captures réalisées en mer suit très bien la courbe moyenne annuelle de migration hors des lagunes. Il est maximum de février à avril. C’est également le cas au Nigeria (Bayagbona, Sagha et Afinowi, 1971) et au Gabon (Fontana et Bâ, 1972). La comparaison des cycles annuels moyens de ponte, de migration et de recrutement a permis de reconstituer en Côte d’Ivoire la chronologie du cycle vital (Figure 2). Si les postlarves qui entrent en lagune au stade 3-4 épines rostrales sont bien âgées de 3 semaines, la grande migration s’effectue vers 3.4 mois à une taille de 16 17 mm (LC). Les premiers modes (18 mm, LC) dans les distributions de fréquence de longueur des captures en mer apparaissent vers 4 mois. Le délai entre la migration et le recrutement dans la population exploitée est très court. La taille de première maturité (30 mm, LC en Côte d’Ivoire) est atteinte ver 6-7 mois. La plus petite femelle mûre trouvée au Nigeria par De Vries et Lefevere (1966) mesurait 28 mm (LC). Les crevettes vivent au moins deux ans.
Elle a été étudiée en Côte d’Ivoire par diverses méthodes, dont celle des marquages. Elles ont toutes fourni des résultats cohérents. La croissance est rapide en saison chaude et fortement perturbée en saison froide, ainsi apparemment qu’en période de ponte. Aucune différence significative n’a été observée dans la croissance moyenne entre la côte africaine et la côte américaine.
La courbe de croissance moyenne annuelle peut être représentée par les paramètres suivants:
|
|
|
Marquage |
Méthode de Petersen |
|
Mâles |
Kmois |
0,30 |
0,26 ± 0,50 |
|
L¥ |
32,3 mm |
34,4 mm ± 2,0 |
|
|
Femelles |
Kmois |
0,18 |
0,16 ± 0,05 |
|
LC¥ |
51,7 mm |
51,7 mm ± 3,4 |
En Côte d’Ivoire les marquages ont permis d’estimer la mortalité naturelle à 0,25/mois; l’analyse des cohortes confirme l’ordre de grandeur (M = 0,21/mois) de cette estimation. Des variations saisonnières de capturabilité ont été mises en évidence.
![]()
![]()
![]()
{kind=link}
{kind=link}