![]()
![]()
![]()
4.3.1 Wooded habitat
4.3.2 Open habitat
4.3.3 Aquatic and wetland habitats
4.3.4 Protected areas
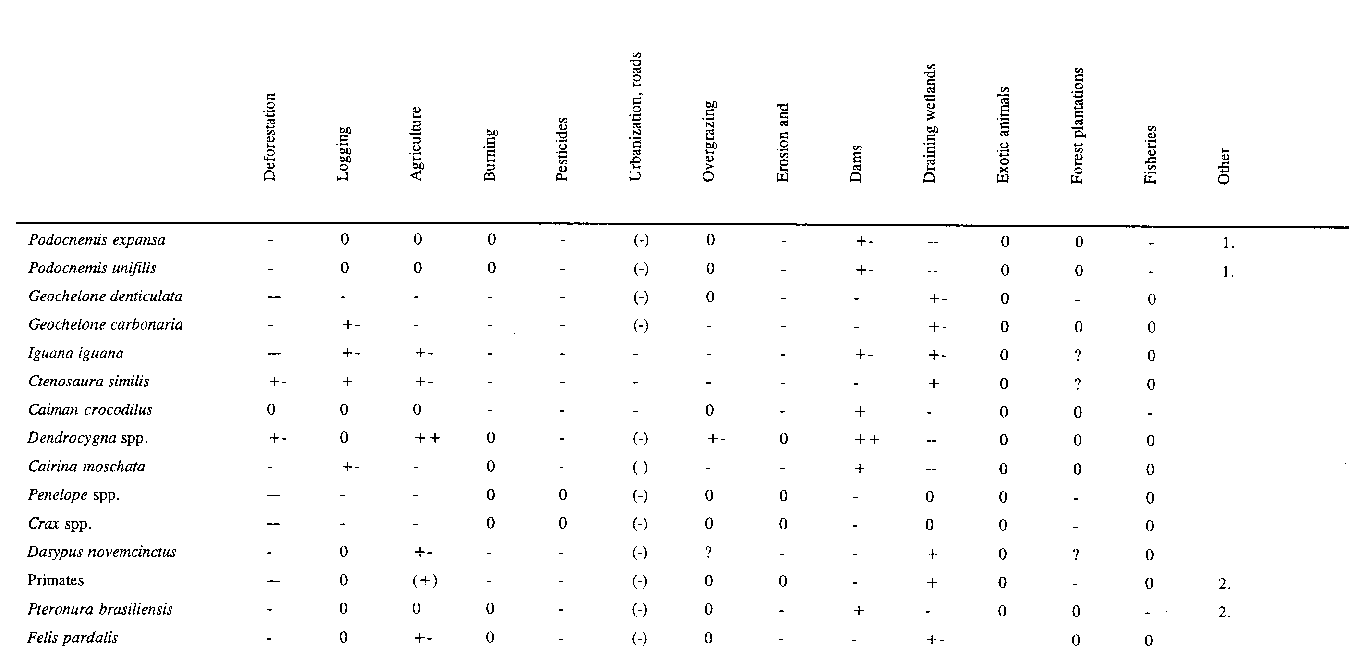
Wildlife populations belong to diverse ecosystems, and their presence and abundance are heavily dependent on habitat size, structure, quality and continuity. At the same time, a number of human activities are acting to modify Latin America's primary ecosystems. In response to the demands of economic growth and burgeoning populations, the pace of environmental modification has been stepped up and wildlife has been heavily affected (Table 29). Some 81 percent of the experts interviewed opined that environmental modifications had a greater impact on wildlife than hunting did.
Anthropogenic environmental modifications cover a vast array of frequently interdependent or synergistic activities. Table 33 summarizes the opinions of the experts surveyed and offers a tentative ranking of environmental modifications in terms of Latin American wildlife. The order of importance was similar for different countries and for the region as a whole (except for Peru), as shown by the statistically significant Kendall coefficients of agreement (W).
Deforestation emerges overwhelmingly as the crucial environmental modifier for wildlife in Latin America, as is true most everywhere. Agriculture-related activities in the broad sense come second, with burning and pesticide contamination in third and fourth place. The fifth environmental modifier is roads, human settlement and urbanization, but here the differences between countries are considerable. Overgrazing and erosion are sixth and seventh in importance (relatively more so in the Andean countries). Hydraulic structures including dams, irrigation systems and the draining of wetlands are eighth. The final factor is introduced plant or animal exotics, which are fairly important in the Southern Cone countries. Under the heading "others" come oil and mining activities, fisheries and the scarcity of wild foods.
Table 33. The main environmental modifications affecting Latin American wildlife, in diminishing order of importance, according to survey findings
|
Modification |
Mexico |
Central America |
Colombia |
Venezuela |
Peru |
Brazil |
Other South American countries |
Argentina |
Chile |
Latin America |
Frequency (times cited) |
|
Deforestation |
3 |
1 |
1 |
1 |
1 |
1 |
2 |
1 |
1 |
1 |
1.00 |
|
Agriculture |
2 |
3 |
3 |
2 |
4.5 |
2 |
1 |
2 |
5 |
2 |
0.98 |
|
Burning |
1 |
2 |
2 |
3 |
2 |
4 |
3 |
5 |
2 |
3 |
1.00 |
|
Pesticides |
6 |
4 |
5 |
6 |
4.5 |
6 |
6 |
3 |
6 |
4 |
0.98 |
|
Urbanization, roads |
4 |
5 |
10 |
4 |
8 |
3 |
4 |
7 |
9 |
5 |
0.83 |
|
Overgrazing |
8 |
7 |
6 |
5 |
3 |
7 |
9.5 |
4 |
7 |
6 |
0.85 |
|
Erosion |
7 |
8 |
4 |
7 |
6 |
8 |
9.5 |
9 |
4 |
7 |
0.80 |
|
Hydraulic structures |
5 |
6 |
7 |
8 |
7 |
5 |
5 |
8 |
8 |
8 |
0.88 |
|
Exotic species |
10 |
10 |
9 |
9 |
10 |
10 |
8 |
6 |
3 |
9 |
0.63 |
|
Other1 |
9 |
9 |
8 |
10 |
9 |
9 |
7 |
10 |
10 |
10 |
0.65 |
|
Number of surveys |
5 |
5 |
4 |
3 |
3 |
4 |
3 |
6 |
5 |
40 |
|
|
Coefficient of agreement W |
0.420 |
0,772 |
0.763 |
0.778 |
0.452 |
0.756 |
0.647 |
0.463 |
0.606 |
0.450 |
|
|
Chi-square and statistical significance |
18.9* |
34.7*** |
27.5** |
21.0* |
12.2 |
27.2** |
29.1*** |
25.0** |
27.3** |
162.0*** |
|
1 Oil and mining activities, fisheries, scarcity of natural food
4.3.1.1 Logging
4.3.1.2 Shifting cultivation
4.3.1.3 Deforestation for agricultural and livestock production
4.3.1.4 Extent of deforestation
4.3.1.5 Impact on wildlife
Most of the key Latin American wildlife groups and species are forest-dwellers, with various types of forest and woodland constituting the natural vegetation in roughly 65 percent of the total area, making environmental modifications of wooded ecosystems singularly important in Latin America. The structure of woodlands and forests can be altered and even virtually eliminated through anthropogenic intervention.
The main modifiers of forest environments are logging and shifting cultivation. An estimated 355 million m3 (516) of timber was cut in Latin America in 1980. The bulk (79 percent) of this volume is used for fuel. Most wood for domestic uses is removed from the dry open forests around villages and towns: this is not necessarily destructive unless demand is much higher than supply. Logging for charcoal for commercial and industrial purposes, however, may seriously reduce forest cover (211, 213, 256, 336, 423, 536). The demand for wood and charcoal, the principal sources of fuel (221, 516), is expected to parallel the increase in rural population.
Industrial logging in tropical forests concentrates on mature forests in areas subject to flooding or near roads. They generally contain a great diversity of tree species, only a few of which are of substantial commercial value. This means that logging is usually highly selective, directly affecting only a small portion of the trees - usually 3 to 10 percent. However, access roads and the felling and hauling of trunks does damage or destroy up to half of the remaining trees, and so the global impact of industrial logging is felt by all tree species (307, 308, 422, 628). The trend now is to take more species and more trees per hectare, thus more profoundly modifying the ecosystem. The growing demand for fine woods for sawmills and raw material for pulp, plywood and paperboard, etc. to meet both domestic and export demand will also mean a dizzying rise in the amount of land affected (422, 516). On the other hand, the lack of roads, the rugged terrain, the legal restrictions and the great distances involved, all of which drive up costs, have to some extent slowed the progress of logging. Forest access roads also foster the invasion of wooded areas by hunters and settlers (256, 307, 447, 615).
Another powerful modifier of tropical forests is shifting cultivation, the main subsistence resource for millions of people in forest areas (30, 256, 332, 422, 516, 615): it may even spell the disappearance of such forests. Under this system, a stretch of wooded vegetation is slashed, burned and farmed on a no-till basis for one, two or even three years. The plot is then abandoned and the cycle repeated elsewhere. Secondary or fallow vegetation springs up in the abandoned areas, which can be cut over after a few years and once again cultivated. As traditionally practised in areas of low population density, this stable land-use system has coexisted for millennia with the tropical forests (307, 615, 628). It generates a mosaic of cultivated patches and successional forests at various stages of recovery in the midst of the forest. The planting cycle is repeated on earlier sites at long intervals and thus has less effect on primary forest, which is very laborious to clear.
The most aggressive form of shifting cultivation occurs in forest areas invaded by landless peasants. This has been called "shifting cultivation driven by necessity" (615), and it usually occurs at higher population densities than do the traditional slash-and-burn systems. There is less land per person and people are obliged to repeat the cycle more frequently on the same sites, leaving the land less time to lie fallow and recover. The campesino cuts more than he can cultivate to signify that the land is occupied. Land available for settlement is often the least suitable land for cultivation e.g. hillsides. Dance (147) established that 81.8 percent of the land occupied by settlers in the Peruvian Amazon was not suited for farming: indeed 74.8 percent of the deforested area, some 5 million ha by 1980, was subsequently abandoned. Shifting cultivation not only eliminates the indigenous vegetation, it also promotes erosion and vastly impoverishes the soil, forcing the settlers to move on to invade new lands, repeating the cycle (106, 252, 375, 422, 615). Lanly (322) estimates that shifting cultivation is responsible for 35 percent of the deforestation in tropical America, mainly in Mexico, Central America and the Andes. Significantly, shifting cultivation is almost always accompanied by subsistence hunting (livestock production is minimal in shifting cultivation systems) (256, 615).
The conversion of tropical forest to permanent crop and grassland is currently the main cause of deforestation in Latin America (322, 422, 516), and the one most frequently cited as the major threat to wildlife (17, 30, 66, 106, 129, 252, 266, 301, 381, 383, 385, 404, 452, 500, 535, 597). Involved are a wide range of government programmes and private initiatives, and there is also a link with shifting cultivation. The spread of both is conditioned and facilitated by the development of road networks (200, 201, 252, 256, 375, 422, 513, 585). The patches deforested by settlers are often the first step in the development of permanent grasslands (307, 332, 422, 615). This however, is a quite different use of the land than in the typical slash-and-burn subsistence minifundio. It involves vast and contiguous stretches of permanently occupied land, mechanized deforestation, private property and capital investment. The principal objectives are commercial monocropping for sugar-cane, or beef production for domestic and export markets (this is particularly true in Central America) (87, 200, 201, 256, 307, 332, 423, 516).
Clearly, a bigger population requires more farmland. And yet, the conversion of tropical moist forests for agriculture usually occurs on land ill-suited for the purpose. The outcome is low soil fertility, erosion, nutrient leaching, soil compaction and pest and disease invasions. The land-carrying capacity and cost-effectiveness of the resultant grasslands drops off dramatically after about ten years (87, 106, 423), with various kinds of secondary vegetation colonizing the degraded soils. Forest removal may be irreversible when it involves heavy soil degradation. In some areas, however, there seem to be more wooded areas than in the past: this is true of parts of Panama and Venezuela (50, 600). In addition to natural regeneration, reforestation programmes contribute to the recovery of degraded land and produce wood. These forest plantations cover some 60 000 km2 in Latin America, primarily monocropped exotics such as Eucalyptus and Pinus (516).
Deforestation has gradually reduced the extent of tropical America's forests. The remaining area of moist tropical forest is thought to be about two-thirds the original amount (87, 426). The reduction has been greater in the Caribbean islands, Central America and the more populated parts of South America. The Brazilian Atlantic forest in eastern Brazil has been almost totally deforested: only a few remnant forests now cover 1-8 percent of the original 750 000 km2 (125, 213, 536). The best documented case in Central America is Costa Rica, which originally was 96 percent forest (579). By 1977, the remaining land under closed forest was 31 percent of the original, and only 17 percent of the primary forest remained in 1983 (513, 579). Deforestation has made greater inroads into deciduous forests in the plains and foothills, and proportionately less in moist and very moist and montane forests (106, 307, 332, 422, 513, 600). The great forest stands of Amazonia are less affected. In the Brazilian Amazon, for example, 4 percent of the wooded area had been deforested by 1980, but the figures were much higher in heavily settled areas (201, 585).
The published figures on current rates of deforestation (area deforested annually vs. initial existing forest area) vary wildly, fostering controversy among the experts (200, 201, 332, 354, 421, 516). Salcedo (516) estimates that 43 000 km2 of closed forest are deforested every year in Latin America, plus 10 000 km2 of open woodlands, which would be an overall rate of 0.41 percent for broad-leaved closed neotropical forests. Over half of the deforestation takes place in secondary or previously logged forests, so the loss of primary forest is less than the annual rate of deforestation.
The highest rates of deforestation, some 1-35 percent each year, are in Central America, Mexico, Colombia, Ecuador and Paraguay. The amount of closed forest deforested annually is greatest in Brazil at 14 800 km2. Proportionately, however, the rate is a low 0.33-0.38 percent, given the vast size of the Amazon forest. The countries least affected by deforestation are Guyana, Suriname and French Guiana (87, 332, 354, 422, 516). The annual pace of deforestation is quickening over time in most countries of the area (200, 201, 332, 422, 516), but tapering off in Venezuela, probably because most of the more accessible forests have already been cut (371).
Wildlife is affected by the modification of the forest because the habitat shrinks and becomes fragmented, altering its structure and quality. Even the few studies we have on the relation between neotropical fauna and the habitat clearly show that deforestation reduces habitat size and quality for forest species, paving the way for wildlife species that prefer secondary vegetation and open areas, such as Odocoileus virginianus, Sylvilagus floridanus and some species of doves. For the great majority of the key species (Table 27) covered in this work, however, the change is not a favourable one. Strict forest-dwellers include Geochelone denticulata, the Cracid family of birds, all primates, most felids, the tapirs, peccaries, Mazama deer, pacas and agoutis. Even aquatic species such as the turtles Podocnemis expansa and P. unifilis which feed in flooded gallery forests are being affected by deforestation. Moreover, Island Biogeography Theory teaches us that the probability of extinction for populations in isolated patches of habitat is proportionate to the size of the patch (355, 630). In other words, a series of isolated wooded patches can sustain fewer species than a block of forest land equivalent to the sum of the area of these patches. Habitat loss is thus actually greater for forest species than the amount actually deforested would seem to indicate. In heavily deforested areas such as the Brazilian Atlantic Forest, certain species of birds and primates are now in danger of extinction (122, 125, 213, 536, 571).
The impact of habitat fragmentation on wildlife depends on the ecological diversity of the patch, its size, the distances and links between patches, and the biological attributes of the wildlife species they contain. Specialist species and larger animals require more area to maintain their populations (307, 500). Neotropical forest game animals are precisely the largest-bodied game species in their respective ecosystems, and so they are greatly affected by forest fragmentation. Some forest floor species, e.g. certain birds, Tayassu tajacu and the felids, may move from one patch to another (109, 181, 353, 521), but the tree-dwelling species, in particular, are confined to the patch in which they live (266, 307, 500). The synergistic effect of habitat fragmentation and hunting further aggravates the effect of isolation in that forest animals are made more vulnerable to hunting as they move through open areas from one patch to the next. Vaughan (597) found the species most affected by habitat fragmentation in Costa Rica to be Trichechus manatus, Myrmecophaga tridactyla, Harpia harpyja, Tayassu pecari, Crax rubra, Panthera onca, Tapirus bairdii and Ateles geoffroyi. The species most affected by forest fragmentation in Venezuela are the tinamous, the cracids and the primates, Priodontes maximum, Dasypus kappleri, Speothos venaticus, Mazama spp., Tapirus terrestris and Tayassu pecari, according to Mondolfi (404). A knowledge of the survivorship of forest species in patches of remnant forest is of vital importance for their management, but single-focus studies on this topic are still quite rare (307, 500, 597).
The fallow patches at different successional stages created by shifting cultivation in primary forest increase ecological diversity in the habitat and are frequented by many wildlife species: Didelphis marsupialis, Dasypus novemcinctus, Tayassu tajacu, Odocoileus virginianus, Mazama spp., Agouti paca, Dasyprocta spp., certain monkeys, Columba, Leptotila, Iguana, Ctenosaura, etc. (98, 257, 307, 336, 337, 404, 509). The apparently beneficial effect of shifting cultivation on wildlife turns negative, however, where such altered habitats predominate and where there is a pattern of short-cycle fallow and erosion-degraded land (30, 356, 615).
Selective logging of the biggest trees may affect the key frugivores and alter the vertical profile of the forest, allowing more sunlight to penetrate and to reach the understorey. This alters the microclimate and stimulates the development of patches of secondary vegetation (307, 308, 628). Pioneer research in the Brazilian Amazon summarized by Johns (307) shows that bird fauna diversity tends to be reduced by logging. The big specialized fruit-eating monkeys like Ateles and Lagothrix disappear from heavily logged areas, but the less specialized monkeys such as Cebus, Saguinos and Saimiri or the leaf-eating monkeys (Alouatta) may actually be more abundant in forests where logging is present. Undergrowth ungulates such as Tapirus terrestris, Tayassu tajacu and Mazama americana appear relatively unaffected by selective logging and may even tend to concentrate in such areas (242, 307). Tayassu pecari, on the other hand, seems to be confined to intact forests (19, 315, 372, 404, 579). Much study remains to be done on the connection between forestry practices and wildlife. For now, however, moderate logging does not seem to have an unduly alarming effect on wildlife.
Forest plantations help to conserve soils and protect degraded watersheds, contribute to forest production and may attenuate the deforestation of primary forest (256, 307, 422, 516). Monocropping exotics, however, is of very limited value for native wildlife in terms of habitat (30, 307, 404, 536). Pine plantations are frequented by some of the less specialized mammals such as the armadillos Dasypus novemcinctus and D. sabanicola, the deer Odocoileus virginianus and Mazama americana, and the agoutis (Dasyprocta sp.) (307, 567).
4.3.2.1 Agriculture
4.3.2.2 Burning vegetation
4.3.2.3 Livestock and grazing
4.3.2.4 Exotic animals
Open habitat includes the following natural ecosystems: tropical savannah, pampa grasslands, the puna and paramo and desert formations, as well as various types of agro-ecosystems and secondary herbaceous communities resulting from the alteration of the primary ecosystems. All in all these open habitats cover roughly one-third of Latin America, and are advancing apace with deforestation.
Tropical agriculture exhibits a wide range of crops and a variable physiognomy from woods to brushland to grasslands, but all have one thing in common: only one or very few species grown for food or industrial purposes predominate. Establishing the crop means removing the native plant cover, tilling the land, applying fertilizer, and planting. Crop maintenance then requires chemical or mechanical control of plant or animal pests on cultivated areas, and frequently irrigation. Harvesting reduces the amount of natural shelter and exports a substantial portion of the primary production and mineral nutrients. Agro-ecosystems therefore involve highly simplified structures and, for short-cycle crops such as cereals and horticultural crops, much sharper seasonal contrasts than do natural habitats.
Among other things, the impact of agriculture on wildlife will depend on the type of crop, the composition and environmental demands of the local wildlife, and the spatial pattern of the cultivated and non-cultivated areas. The relationship between wildlife and agriculture is a very complex one, but in general habitat change operates to the detriment of most forms of native wildlife. Some opportunistic species do, however, benefit from the cover, the ecotones, and, particularly, the food which crops provide.
Data on the impact of habitat modification on neo-tropical wildlife due to agriculture is quite fragmentary. Most of the data concerns species that find their optimum habitat under certain specific cropping systems, where they can become very abundant. The most striking examples are: the concentration of ducks, particularly Dendrocygna spp., in rice fields (22, 59, 154, 204, 310); the link between sorghum growing and the abundance of Zenaida auriculata and other doves (84, 177, 178, 197, 418, Hernandez Camacho, survey, 1982); and the expansion of the coyote (Canis latrans) as a result of deforestation in Central America (19, 336, 597). The deer Odocoileus virginianus is one of the few big game species that can co-exist with agriculture and even benefit from the advance of the agricultural frontier (81, 149, 238, 336). On the central llanos of Venezuela this deer prefers the growing phase of sorghum to its natural habitat, and makes much use of this cereal during the transition between the rainy and dry season, when most wild fodder is of poor quality (149).
Forest-dwelling mammals are losing their habitat as agriculture gains ground. Many species, such as peccaries, the single-antler deers (Mazama spp.), pacas, agoutis and some monkeys (particularly Cebus) are in the habit of raiding cultivated patches amid the forest (66, 178, 294, 336, 402). They can do considerable damage and even become farm pests. Some species are more vulnerable to hunting from blinds in cultivated areas than they are in their own wild habitat. Additionally, crops can attract and concentrate animals from miles around in a funnel effect. Populations from much larger areas than the actual cropland can thus be culled. In the long run, the above species are gradually disappearing from farm areas for lack of habitat.
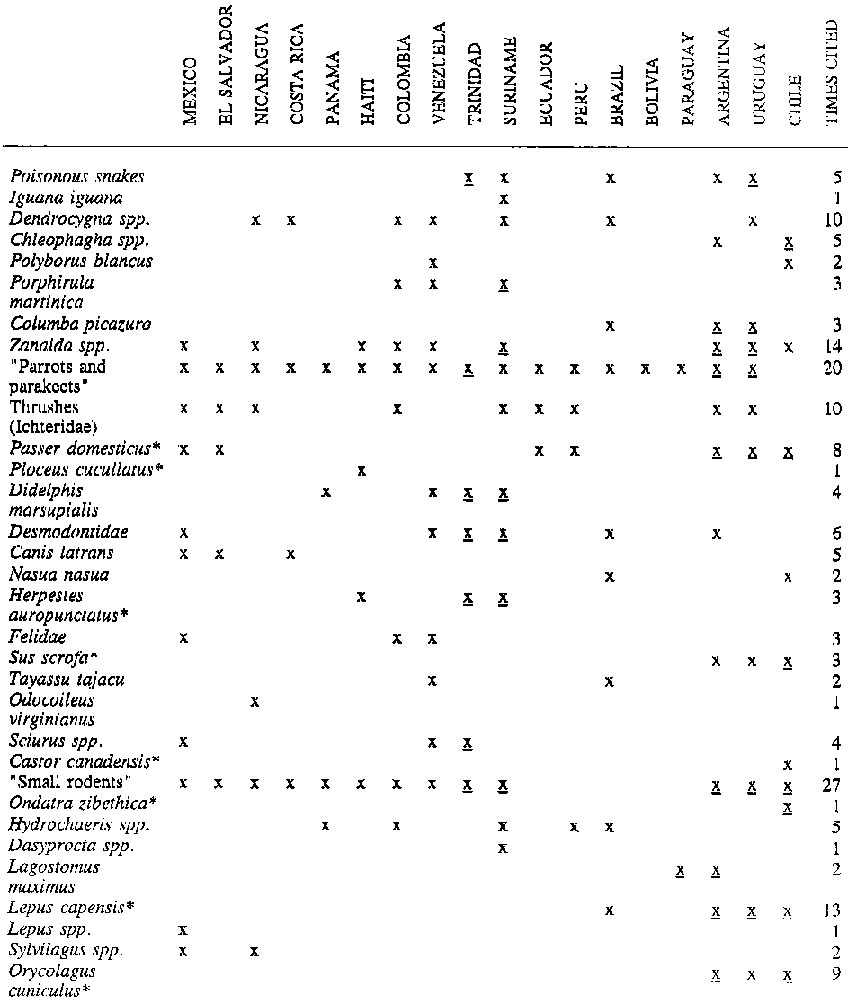
The main vertebrate pests are open habitat species (Table 35). Included in this group are the whistling ducks (Dendrocygna spp.) and the purple gallinule (Porphyrula martinica) in rice fields; geese (Chloephaga spp.) in southern South America; doves and species of parrots, parakeets and cockatoos (Forpus passerinus, Aratinga canicularis, A. pertinax, Pionus senilus, Myiopsitta monachus) and other grain and fruit-eating birds such as Molothrus, Cassidix, Leistes, Dolichonyx, Ploceus, Sporophila. Sicalis, etc. The most important agricultural pests are mice, rats and other small rodents, either native (Cricetidae, Geomydae, Caviidae, Ctenomydae, etc.) or introduced (Rattus, Mus) (164, 177, 178). The exotic leporidae Lepus capensis and Oryctolagus cuniculus and the viscacha Lagostomus maximus are agricultural pests in southern South America, causing particularly heavy damage in degraded grasslands (64, 85). The capybara may also behave as a pest on farmlands (17, 21, 409, 440).
Modern agriculture uses a wide variety of biocides to combat crop pests, weeds and diseases. These toxic agro-chemicals are widely used with few controls over much of Latin America and are believed to have a severe impact on wildlife (Table 33). What data we do have on accidental or intentional poisoning of birds and fishes by biocides is sporadic (30, 129, 269, 505, 536, 592). Wildlife contamination by biocides remains an unknown factor requiring urgent attention, particularly in the main intensive cultivation areas.
Fire is a major ecological factor in forest as in open ecosystems. Forest fires are usually accidental, destructive to the ecosystem, and to be avoided at all costs (106, 242, 252, 404). The frequent and usually intentional burning of vegetation in open tropical habitats is, however, one of the most characteristic and controversial features of these ecosystems (2, 282, 385, 518, 536, 596).
Burning scrub and grasslands during the dry season is an ingrained tradition in tropical America. The fires stimulate the regrowth of fodder grasses, eliminating undesirable dead, woody and grass biomass, killing snakes and other animal pests and accelerating the nutrient cycle. Vegetation is occasionally burned to facilitate hunting as well (296, 460, 507, Ojasti, pers. obs.). Certainly, fire does temporarily reduce the supply of fodder and natural shelter, and in the long run affects floral composition, eliminating some species, whereas the more fire-resistant gain ground. Eliminating plant cover on hillsides can favour erosion and nutrient leaching. In the wake of fire, nests, young, invertebrates and small vertebrates may be destroyed and/or exposed to predators, but most wildlife manage to flee fire or to find shelter in their burrows, it is believed (2, 106, 282, 382, 596).
The burning of vegetation is often cited as a negative factor for wildlife (17, 252, 242, 396, 507, 536), and is prohibited and/or controlled by law in many countries. Objectively speaking, however, not much is really known about the impact of fire on savannah wildlife (445). The periodic burning done in many grassland areas, which eliminates woody species, particularly their seedlings, and favours perennial grasses, is necessary if grasslands are to be maintained for wild or domestic herbivores (2, 85). Burning, therefore, which can either degrade or upgrade wildlife habitat, should be used with great caution. What matters most, in the final analysis, is the type of vegetation involved, which wildlife species have priority, the frequency and timing of burning, and how it is done (2, 282, 385, 440).
The domestic herbivores that came in with the colonization of Latin America assumed the ecological niches of the dominant grazers in open habitats everywhere, or nearly so. Grazing and other livestock activities, which involve vast areas, are singularly important for this reason. Cattle production is at the top of the list, with 265 million head in Latin America. Sheep production, with a population of some 110 million, is more common in the Southern Cone and Andean countries. There are also about 30 million equine species. Swine production is very important economically, but tends to be concentrated in intensive systems and has less impact on grasslands. Generally speaking, Latin American livestock production systems are based on extensive use of natural grassland within the latifundio land tenure system, of which the components are huge properties, low investment, and low production per unit of area. The land tenure system combines with export economics to create a situation where some farm land is used for ranching, and thus underutilized from the social standpoint (87, 227, 422, 423).
Livestock production activities affect indigenous wildlife in that domestic herbivores eat the grass and trample the ground: then there are watering points, fencing, roads and other installations, grasslands management, the sanitary implications and the control or culling of large predators and other wildlife considered as pests. The order of importance and intensity of these factors varies from place to place, and we have only a very general understanding of their impact on indigenous wildlife.
Moderate grazing by domesticated animals does seem to be perfectly compatible with the environmental requirements of native wildlife in the same habitat. Grazing also reduces the mature plant biomass, stimulating regrowth and improving fodder quality for selective grazers such as the capybara (440, 448). The watering points established for livestock also improve the habitat for wild species (80, 149, 242, 272, 446). It is quite likely that the culling of the big cats has improved the survival rate of their native prey. A landowner may also regulate hunter access to his property.
Overgrazing is a very widespread feature of Latin American livestock production systems and one of the gravest threats to wildlife habitat (Table 33), potentially forcing wildlife and domestic species to compete for fodder. Where the carrying capacity of the land is exceeded, the high-value fodder plants are gradually replaced by weeds or stretches of bare ground, promoting erosion and desertification in ranched areas (80, 85, 184, 282, 336, 536). The irreversible loss of soil through erosion due to overgrazing, deforestation and poor farming practices is spreading at an alarming pace in Latin America, particularly in Mexico (46, 336, 615), and in the Andean and Southern Cone countries (85, 147, 164, 184, 300, 256, 615). A few opportunistic species such as hares (Lepus spp.), vizcachas (Lagostomus maximus) and jackrabbits (Pediolagus salinicola) (63, 64, 85, 336) may thrive in severely overgrazed areas. The extent to which other herbivores are affected is not well documented, however, although we do know that overgrazing by sheep has caused a decline in the carrying capacity of vicuña and guanaco habitats (85, 93, 282, 322, 488). Barbed wire fences may hinder the movements of some large mammals such as guanacos and pronghorns, and there are reports of these animals being trapped in such fences (93, 336, 488).
Wildlife are also subject to some of the same diseases as domestic animals. Foot-and-mouth disease, for example, also affects deer, particularly the pampas deer (Ozotoceros bezoarticus) and the marsh deer (Blastocercus dichotomus) (93, 122). Trypanosomiasis, also known as crippling or hip disease, affects both equines and capybaras (191, 432, 440, 522, 524). Very little is known, however, about the role of this and other diseases as a factor in Latin American wildlife mortality.
Most bird and mammal exotics introduced in Latin America live in altered, open or peri-urban habitats, with the greatest numbers found in southern South America. The list of introduced mammals ranges from big game species such as the European boar (Sus scrofa) to the deer species Cervus elaphrus, Dama dama and Axis axis which were introduced in specific hunting reserves in the early part of this century. Many specimens escaped from these reserves, however, and wild populations are now scattered over various areas of Argentina, Chile, Peru and Uruguay (156, 164, 176, 198, 303, 471). These exotic deer are a valuable game resource, particularly when properly managed in game reserves, but they may also alter the vegetation and compete with wild species (176, 198). The European boar, pure or crossbred, is generally considered a pest species (303, Table 35).
Rabbits and hares (Oryctolagus cuniculus, Lepus capensis) are the exotics with the greatest impact in Latin America. They were introduced in 1980 in Argentina and Chile. The European hare is abundant in Argentina, Chile and Uruguay, reaching Paraguay and southern Brazil. The rabbit is distributed throughout Chile, mostly, but populations are also found in Argentina (156). Both species are voracious herbivores, prolific, reach high population density and thrive in open modified environments. They are frequently responsible for serious plant degradation and damage to agriculture and forest plantations. At the same time they are subject to massive commercial hunting, and this does to some extent offset the environmental harm they do (63, 362).
Introduced rodents in South America include the beaver (Castor canadensis) and the muskrat (Ondatra zibethica) in southernmost South America. Both are now very limited in distribution, but they do have a great impact on wetlands and surrounding areas where present (199). Numerically, most of the rodents unintentionally introduced by people in inhabited areas belong to the muridae species Mus musculus, Rattus rattus and R. norvegicus. They do not easily infiltrate primary systems, however, and this limits their impact on wild habitats. They same is true of introduced birds such as Passer domesticus and Columba livia.
The most frequently cited mammal in tropical America is probably the mongoose (Herpestes auropunctatus), which was introduced on various Caribbean islands and in Trinidad, Guyana and Surinam during the last century to control snakes and rodents in crops (156, 294). This small carnivore from India feeds on small vertebrates and can eliminate indigenous species, particularly in island habitats. On the continent, however, the mongoose confines itself to modified environments and does not penetrate the primary forest (294). In the neo-tropical countries where it is found it is considered a pest (Table 35).
Unsupervised domestic animals, many of which have gone wild, can also be considered exotics. From early colonial times to the present herds of cattle, for example, living in the wild in habitats inaccessible to horses, have been hunted or captured just like the indigenous species (93). Packs of wild dogs freely roaming wildlife habitats can be highly destructive. They cause high mortality among the llanos capybaras (312, 440), and are reportedly the principal predators of vicuña in Peru (282), pudu in Chile (276) and the unique fauna of the Galapagos Islands in Ecuador (58). Semi-domestic or wild pigs are locally abundant in a number of countries and environments. Given their voracious and omnivorous feeding habits, these animals can destroy vast tracts of wetland, rooting among the vegetation, contaminating watering holes, destroying eggs and young, and competing with many of the native species (93). Domestic animals that have reverted to the wild are known to be very influential, but there is very little objective data on their ecological impact in Latin America.
Inland water habitats, permanent or temporary flood plains, brackish coastal and estuarine areas and other wetlands, including cultivated ones, cover approximately 10 percent of Latin America (150). Many key neotropical wildlife species (Table 27) are either closely linked with or restricted to these habitats: river turtles, crocodilian species, ducks, manatees, Pteronura, capybara and coypu. Most of the rest require water every day and so there must be some body of water in their habitat.
Man-made environmental modifications of aquatic habitats include various hydraulic structures such as dams and irrigation systems intended to increase the area under water, or others intended to reduce it by drainage. Soil erosion inevitably leads to sedimentation in streams and rivers, whereas environmental pollution is channeled into and builds up in aquatic habitats. Aquatic ecosystems are very much exposed to various kinds of modification, and their research and management should accordingly be given high priority (425, 505).
The main causes of shrinking wetland and brackish freshwater habitats are drainage for agriculture, and destruction and/or damming within the mountain watersheds that once abundantly watered the lower basins (150, 184, 336). These processes are diminishing habitat size and quality for ducks and other semi-aquatic wildlife, particularly in Mexico, Central America and Colombia (24, 67, 336, 618) - although such habitat loss is partly offset by the development of irrigated cropping systems (70, 164, 184, 238, 336). Wild species that are resources in their original habitat may come to behave as pests in farm areas.
The construction of long earthen dykes for roads or water retention is altering the hydrogeography of the lowland llanos of Venezuela and Colombia, the Pantanal de Matogrosso in Brazil, and other flood plain areas. The waters behind these dykes increase the extent and duration of flooding, favouring semi-aquatic and aquatic wildlife (31, 32, 236, 242, 440, 492, 532). In addition, the series of excavations or pits dug during the construction of these dykes or roads come to form permanent bodies of water that are used by ichthyofauna, crocodilian species, turtles, capybaras and waterfowl during the dry season. However, the swift build-up or draw-off of dammed waters through sluices accentuates seasonal contrasts, and may be harmful to both aquatic and terrestrial wildlife (442).
Large dams, built primarily for electric power, are a relatively recent modification on the big South American rivers. The new lakes upriver of the dams cover the terrestrial ecosystems of the dammed valleys, and losses of terrestrial habitat can be quite vast on the local scale. The Tucurui dam on the Tocantis River in Brazil, for example, flooded 2 160 km2. A total of some 35 000 km2, an area nearly as big as the island of Trinidad, will be covered by the dams now planned for the Brazilian Amazon (242).
The best-documented aspect of wildlife and dams is the rescue operations mounted as the dams fill (39, 307, 614). The longer-term effects of damming on indigenous wildlife are not so well- established. It is probable, however, that habitat quality for terrestrial fauna living along the shores of reservoirs or dispersed in islands compares unfavourably with that of the riverine forests and alluvial soils covered by the waters of the dam. The aquatic fauna in these reservoirs are also mainly river species adapted to the rivers' seasonal changes and many species are dependent on allothogenic food. Most aquatic mammals, birds and reptiles are likely to be shore-bound, and not adapted to make use of the vast stretches of water that reservoirs offer. We therefore do not know to what extent aquatic wildlife can compensate for the loss of habitat suffered by terrestrial species.
In addition to drastic alterations of aquatic environments, people may interfere directly with their associated wildlife. Concentrations of urban, tourist and recreational activities along coastal strips can drive off waterfowl, complicating the conservation of endangered species endemic in such areas, such as manatee and caiman Crocodylus acutus. Collisions with speedboats are reportedly a major cause of manatee mortality in Florida (262, 436). Aquatic mammals and reptiles are also killed in the course of fishing operations, or lose habitat as mangroves are cut back and shrimping expanded in coastal lagoons (159, 403, 407, 465).
4.3.4.1 Status and extent of protected areas
4.3.4.2 Protected areas and wildlife
In response to the progressive modification and decline of natural ecosystems, Latin American countries are setting aside parts of their national territories as national parks or other protected areas. Much progress has been made in the establishment of protected areas in the last 20 years, and this is currently a basic thrust of environmental and land management policy in many countries. Statistics on specially administered areas, compiled from various sources (Table 36), can give us a tentative summary of the current situation in Latin America.
Over 200 national parks and similar units cover 1.77 percent of Latin America's territory. The total area under special administrative regimes covers about 4.5 percent of the territory, an amount roughly the size of Venezuela. There are great differences from one country to the next, however. Chile, Costa Rica, Venezuela, Panama, Ecuador and Colombia have far more land under national parks than half the countries in Table 36, where protected land comprises not even 1% of the total. The total amount of protected areas is greater in Venezuela in both relative and absolute terms. The proportion is also high in Chile, the Dominican Republic, Costa Rica, Peru, Panama and Cuba, whereas it is under 1 percent in Mexico, some countries of Central America, Brazil and Uruguay.
The number and size of national parks and other protected areas is only one indicator of their development: the degree of management and protection actually offered in protected areas is as or more important (though harder to quantify). Often enough, faced with the imminent occupation of wild areas of particular interest, an area of land is declared a protected area, even though there are no resources to provide even the bare minimum of administrative and wildlife protection services. The upshot is the so-called "paper parks" that appear only on maps and in statistics, but not in reality. Well-established protected areas are also facing various kinds of external pressure that interfere seriously with their declared purpose: conservation (106, 123, 222, 242, 256, 307, 480). Another factor to consider is the management pattern and whether or not it conforms to the specific situation in each case. There is a great range of legally protected areas. Some involve absolute protection while others allow certain uses, including rational resource exploitation. The park services of the national parks and other protected areas in various countries are now reorganizing and redefining their conservation units and management categories with the collaboration of the FAO Regional Technical Cooperation Network on National Parks, other Protected Areas and Wildlife and the IUCN Commission on National Parks and Protected Areas; the goal being a sounder and more functional protection system. The biosphere reserves concept, sponsored by the "Man and the Biosphere" programme (MAB) of UNESCO, has also given a new international dimension to the management of protected areas.
Table 36. National parks and other protected or specially administered areas in Latin America: number, estimated area and percentage of national territory. Current validity varies according to source of information.
|
Country |
No. |
National Parks km2 |
% |
No. |
All areas km2 |
% |
Source |
|
México |
19 |
2779 |
0.14 |
33 |
5 503 |
0.28 |
222 |
|
Cuba |
1 |
258 |
0.23 |
28 |
9 120 |
8.0 |
IUCN 19861 |
|
Dominican Republic |
8 |
? |
? |
13 |
5 445 |
11.3 |
IUCN 1986 |
|
Guatemala |
4 |
958 |
0.88 |
7 |
987 |
0.91 |
IUCN 1986 |
|
Honduras |
2 |
75 |
0.07 |
9 |
575 |
0.51 |
222 |
|
El Salvador |
3 |
29 |
0.14 |
4 |
40 |
0.19 |
222 |
|
Nicaragua |
2 |
173 |
0.21 |
2 |
173 |
0.21 |
222 |
|
Costa Rica |
13 |
5 2505 |
10.3 |
29 |
5 671 |
11.1 |
Boza s/f |
|
Panama |
5 |
6 555 |
8.51 |
8 |
6 609 |
8.58 |
222 |
|
Colombia |
25 |
38310 |
3.36 |
34 |
39587 |
3.43 |
222 |
|
Venezuela |
32 |
86435 |
9.48 |
119 |
355 960 |
39.0 |
INPARQUES 1988 |
|
Suriname |
13 |
84 |
0.06 |
15 |
8040 |
5.62 |
Baal, 1988 |
|
Ecuador |
5 |
15 302 |
5.80 |
11 |
19902 |
7.54 |
222 |
|
Brazil |
25 |
35750 |
0.42 |
37 |
55 750 |
0.65 |
Quintao 19834 |
|
Peru |
6 |
22591 |
1.76 |
34? |
112794 |
8.785 |
480 |
|
Bolivia |
10 |
19 105 |
1.74 |
18 |
43 507 |
3.69 |
222 |
|
Paraguay |
7 |
11 174 |
2.75 |
11 |
11 185 |
2.75 |
222 |
|
Argentina |
19 |
25 6962 |
0.93 |
122 |
103 647 |
3.73 |
IUCN 1986 |
|
Uruguay |
4 |
194 |
0.11 |
8 |
305 |
0.17 |
222 |
|
Chile |
34 |
93 000 |
12.3 |
74 |
134 200 |
17.7 |
CONAF s/f |
|
Total |
216 |
363718 |
1.77 |
616 |
919 000 |
4.47 |
|
1 Proceedings of the 27th working session of IUCN's Commission on National Parks and Protected Areas; Bariloche, 1986.2 Includes reserves and refugees.
3 Natural park.
4 Brazil Florestal N° 54: 13-28; 1983.
5 42% according to Ponce del Prado (480).
Indigenous wildlife can be a very compelling component of and unique scenic attraction in some protected areas of Latin America. The Pampa Galera and Paracas National Reserves in Peru, the road that runs through the Pantanal de Mato Grosso in Brazil and the Cuare Wildlife Refuge in Venezuela are just a few examples. Even in these cases, only a very small fraction of the protected area was set up explicitly to protect wildlife. The main purpose in most protected areas was to safeguard representative specimens of whole and intact wild ecosystems (strictly protected areas), or else to institute rational resource use as part of a sustained multi-use system. However, there is virtual consensus that protected areas do play a very basic role in wildlife management (46, 71, 92, 123, 142, 153, 222, 245, 269, 301, 336, 385, 404, 597, survey findings). Several countries have also legislated the establishment and management of protected wildlife areas offering incentives to encourage private initiative to emulate these initiatives (25, 26, 76, 117, 128, 135, 171, 249, 387, 467, 468, 584, 601).
Strictly protected areas are the best way of ensuring the survival of critically endangered species, particularly when the main problem is habitat modification. Other than preserving critically endangered species, protected areas should also favour other local wildlife and even produce surplus populations that can spread out to the surrounding areas. Considering the low reproductive capacity and sedentary habits of many forest-dwelling species, the process may be less impressive in forests. The best-documented success of the recovery of an endangered population in a protected area is that of the vicuña on the Andean altiplano (72, 96, 191, 486, 506). Unfortunately, this case is an exception, perhaps owing its success to exceptionally efficient technical backing. Generally speaking, habitat conservation in protected areas seems to be a much more feasible option than the eradication of hunting. And while there is no fully reliable documentation on the subject, it seems quite likely that poaching does take place in most protected areas in Latin America, which of course partially detracts from their major function: wildlife protection. There is a need to combat these irregularities by more efficient wildlife protection and by building awareness among the local people.
Additionally, protected areas which do allow controlled resource use are often larger than those enjoining strict protection. It is also true that rational utilization of such areas for the benefit of the local people can spark popular support for conservation programmes. Simultaneous conservation and utilization, such as IUCN's management category VIII (multi-use management), offers great potential for the sustained production and utilization of wildlife. This principle is explicit in certain legally established zones such as reserves or experimental hunting grounds, but it also applies in many other cases, e.g. conservation efforts where forest management or watershed conservation is the top priority. Unhappily, the general rule seems to be that while hunting is not authorized, these areas lack effective control measures to prevent it. This alone is an incentive to poachers, and minimizes the value of wildlife as a resource. Much remains to be done in the field of applied research, planning, extension and wildlife protection to achieve sustained use of the vast potential of wildlife in protected productive areas. In Venezuela, for example, these occupy more than one-quarter of the national territory. Clearly, a sustained contribution of protected areas to the regional economy would be the best justification both for their existence and in terms of their long-term survival.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}