![]()
![]()
![]()
A. YenikoyeUniversité de Niamey
B.P. 237
Niamey
Niger
Resume
Summary
Introduction
Résultats et discussions
Conclusions
Références
Les modifications du niveau et du rythme de sécrétion des gonadotropines PRL, FSH et LH associées aux variations saisonnières de l'activité reproductive ont été étudiées chez la brebis Peul en milieu sahélien au Niger. Il existe un rythme circannuel de la prolactine chez la brebis soumise aux variations normales de photopériode et de température. Les concentrations maximales de prolactine (> 200 ng/ml) sont observées en avril et mai (durée du jour plus longue et température plus élevée). Les concentrations minimales (< 100 ng/ml) sont observées en décembre et en janvier (durée du jour plus courte et température plus basse). La durée du jour est le principal facteur de l'environnement modulant la sécrétion de prolactine. L'évolution circ-annuelle du niveau du LH montre une réduction de la concentration en fev/mars, juin/juil, sep/oct et en décembre due à une baisse de fréquence des pulses de LH (1 pulse/1,6 h contre 1 pulse/4,0 h) sans changement de sensibilité du complexe hypothalamo-hypophysaire aux stéroïdes ovariens. Il n'y a pas de variation saisonnière de l'activité autonome de l'hypophyse à libérer FSH et au rectro-contrôle de cette hormone par les stéroïdes ovariens. L'hypothèse que l'activité reproductive de la brebis Peul est contrôlé par le rythme endogène de libération de LH est soutenue.
Seasonal changes in the levels and patterns of the gonadotropins PRL, FSH and LH were studied in Peul sheep in the Sahel zone in Niger. Highest concentrations (> 200 ng/ml) of prolactin were observed in Apr/May (long days and high temperatures) and lowest (< 100 ng/ml) in Dec/Jan (short days and low temperatures). Photoperiod is the principal environmental factor influencing prolactin secretion. Circannual changes in LH level show a reduction of the concentration in Feb/Mar, Jun/Jul, Sep/Oct and in December, caused by a reduced frequency of LH secretion (1 peak/1.6 hr against 1 peak/4.0 hr) and not by a change in the sensitivity of the hypothalamo-hypophysary complex to ovarian steroids. There are no significant seasonal variations in autonomous pituitary activity for releasing FSH or in feedback by ovarian steroids. The results suggest that seasonal variations in sexual activity in Peul sheep are controlled by the frequency of LH peaks.
La fécondité de la brebis Peul en milieu sahélien est limitée par un anoestrus saisonnier de décembre à avril et des dioestrus prolongés de février à mai et de juillet à octobre (Yenikoye et al, 1982; Yenikoye, 1984). L'étude des variations saisonnières des niveaux de gonadotropines FSH, LH et prolactine (Cognie et al, 1981; Oussaid, 1983; McNatty et al, 1984) a permis de préciser les degrés de dépendance existant entre l'ovaire et l'hypophyse au cours de l'anoestrus chez la brebis. En début d'anoestrus, il est seulement observé une diminution de la fréquence des pulses de LH et le milieu de l'anoestrus est marqué par une chute des niveaux de FSH et LH (McNatty et al, 1981; Oussaid, 1983; McNatty et al, 1984). On parle aussi d'anoestrus "léger" dans le premier cas et d'anoestrus "profond" dans le second. L'allongement de la phase lutéale ou dioestrus prolongé chez la brebis est associé à une diminution de la fréquence de libération des prostaglandines d'origine utérine (Sheldrick et al, 1980; Zarco et al, 1984) mais le rythme de sécrétion des prostaglandines utérines est contrôlé par les oestrogènes ovariennes (Hixon et al, 1975) et l'oxytocine hypophysaire (Zarco et al, 1984). Il est alors possible que le dioestrus prolongé chez la brebis soit dû à une déficience en gonadotropines (Restall et al, 1978).
Les mécanismes endocriniens mis en jeu en milieu tropical et les relations entre ces mécanismes et la reproduction sont encore mal connus. Cet article reporte les modifications des sécrétions de gonadotropines associées aux variations saisonnières de l'activité ovarienne chez la brebis Peul.
Variations circannuelles de la prolactine
La concentration moyenne mensuelle de la prolactine suit une variation saisonnière parallèle à la durée du jour et à la température (Yenikoye et Ravault, 1981). Les concentrations maximales sont observées en avr/mai quand la durée du jour est la plus longue et la température moyenne est la plus élevée (figure 1). Les concentrations minimales coïncident avec les jours les plus courts et les températures les plus basses en déc/jan. Cependant, l'étude de la régression de la concentration de prolactine en fonction de la durée du jour et de la température montre que seule la photopériode est significative (P < 0,025). Il semble que chez la brebis Peul bicolore, comme chez d'autres races ovines étudiées dans les régions tempérées et froides (Thimonier et al, 1978; Munro et al, 1980), la durée du jour est le principal facteur de l'environnement qui module le rythme de sécrétion de la prolactine.
Malgré la faible amplitude de variation (max = 1,5 h) de la photopériode dans les régions subtropicales par rapport aux zones tempérées (max = 8,0 h), le modèle de sécrétion de la prolactine reste similaire. Ceci suggère, d'une part que le climat sahélien influence peu le rythme de sécrétion de la prolactine chez la brebis, et d'autre part qu'une faible variation de la photopériode est capable de stimuler sa sécrétion.
Figure 1. Variations saisonnières de température, de photopériode et de prolactine chez la brebis Peul au Niger.

Variations circannuelles de LH
Contrairement à la prolactine, LH n'est pas sécrétée de façon continue mais sous forme pulsatile (Knobil, 1987). Chez la brebis Peul l'amplitude et la durée des pulses de LH ne changent pas au cours de l'année et seule la fréquence varie (tableau 1). Les 2 périodes maximale et minimale correspondent à la saison sexuelle et à l'anoestrus saisonnier. Le rythme de sécrétion de LH est dépendant de l'activité intrinsèque et de la sensibilité du complexe hypothalamo-hypophysaire aux stéroïdes ovariens (Karsch et al, 1980).
Activité intrinsèque du complexe hypothalamo-hypophysaire. Chez la brebis Peul castrée, car l'activité est mise en évidence chez l'animal ovariectomisé (Goodman et Karsch, 1981), le niveau quotidien de LH chute à 4 périodes de l'année (figure 2). Cette diminution est due à une baisse de fréquence des pulses de LH sans modification d'amplitude, ni de durée (tableau 1).
Tableau 1. Variations saisonnières de la pulsatilité de LH chez les brebis normale et castrée de race Peul au Niger.
|
Traitement et mois |
Pulse |
|||
|
Amplitude (ng/ml) |
Durée (min) |
Fréquence (h/pulse) |
||
|
Brebis normale |
||||
|
|
Septembre (saison sexuelle) |
3,2 ± 0,1 |
64,0 ± 5,1 |
1,6 |
|
|
Mars (anoestrus) |
3,2 ±0,1 |
65,0 ± 15,0 |
4,0 |
|
Brebis castrée |
||||
|
|
Septembre (saison sexuelle) |
5,5 ± 0,3 |
62,5 ±4,8 |
1,3 |
|
|
Mars (anoestrus) |
5,6 ± 0,2 |
57,2 ±4,8 |
2,0 |
Les études effectuées chez la brebis normale ou castrée et chez la vache (Miller et Alliston, 1974; Schillo et al, 1978; Hooley et al, 1979) ont montré que le niveau de sécrétion de LH baisse lorsque la température ambiante augmente ou lorsque la température rectale s'accroît de 1°C. Il a été observé chez la brebis Peul que l'amplitude circadienne de la température rectale en période d'anoestrus augmente de 0,85°C par rapport à la saison sexuelle. Il existe probablement des relations entre les centres de l'homeostasie thermique et ceux qui contrôlent la sécrétion de LH (Riggs et Malven, 1974; Bligh, 1986).
Sensibilité du complexe hypothalamo-hypophysaire aux stéroïdes ovariens. L'élévation relative du niveau de LH après ovariectomie, l'intervalle entre le moment de la castration et le pic de LH suivant l'ovariectomie (Land et al, 1976; Karsch et al, 1980), la diminution du niveau de LH après injection d'oestrogène à des animaux castrés et l'intervalle entre le moment de l'injection et le début d'une décroissance significative de LH (Goodman et Karsch, 1980) sont les critères d'évaluation de la sensibilité du complexe hypothalamo-hypophysaire aux stéroïdes ovariens.
Les paramètres ne subissent pas de variation saisonnière significative chez la brebis Peul (tableau 2). Ceci suggère que la sensibilité du complexe hypothalamo-hypophysaire aux stéroïdes ovariens ne change pas suivant les périodes favorables et défavorables de l'activité sexuelle. Ainsi donc, contrairement aux brebis de races européennes (Land et al, 1976; Goodman et Karsch, 1980), on n'observe pas d'augmentation du feedback négatif aux oestrogènes chez la brebis Peul en période d'anoestrus saisonnier.
Figure 2. Evolution circannuelle du niveau de LH chez deux brebis ovariectomisées au Niger.
Tableau 2. Effets de castration et d'injection d'oestrogène sur les paramètres de sécrétion de LH et de FSH.
|
Paramètre |
Période |
||
|
Sep-Nov |
Fev-Mar |
||
|
LH |
|
|
|
|
|
Niveau avant castration (ng/ml) |
0,99 ± 0,15 |
0,93 ± 0,2 |
|
|
Niveau après castration (ng/ml) |
1,64 ± 0,4 |
1,75 ± 0,3 |
|
|
Taux d'accroissement |
1,21 ± 0,4 |
1,07 ± 0,2 |
|
|
Intervalle, castration-pic LH (h) |
12,85 ± 3,1 |
11,3 ± 5,0 |
|
|
Niveau avant injection (ng/ml) |
4,1 ± 0,3 |
4,2 ± 0,1 |
|
|
Niveau après injection (ng/ml) |
2,3 ± 0,2 |
2,2 ± 0,1 |
|
|
Taux de diminution |
-0,45 ± 0,03 |
-0,49 ± 0,03 |
|
|
Intervalle, injection-chute (h) |
-3,9 ± 0,6 |
4,3 ± 0,6 |
|
FSH |
|
|
|
|
|
Niveau avant castration (ng/ml) |
4,7 ± 0,7 |
3,6 ± 0,1 |
|
|
Niveau après castration (ng/ml) |
8,2 ± 1,1 |
9,9 ± 0,7 |
|
|
Taux d'accroissement |
1,7 ± 0,4 |
1,8 ± 0,2 |
|
|
Intervalle, castration-pic LH (h) |
17,4 ± 7,4 |
11,1 ± 5,5 |
|
|
Niveau avant injection (ng/ml) |
47,7 ± 10,9 |
34,0 ± 4,5 |
|
|
Niveau après injection (ng/ml) |
20,9 ± 0,03 |
-0,59 ± 0,03 |
|
|
Taux de diminution |
-0,56 ± 0,03 |
-0,59 ± 0,03 |
|
|
Intervalle, injection-chute (h) |
-4,04 ± 0,2 |
4,93 ± 0,1 |
Variations interannuelles de FSH
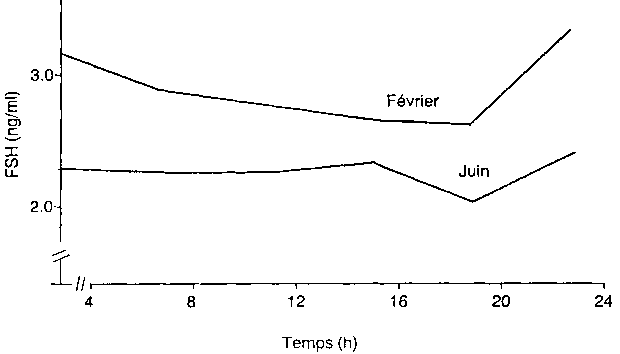
Les concentrations ciradiennes de FSH sont les plus basses à 20 h et les plus élevées à 24 h (Yenikoye, 1986). La concentration de FSH est à tout moment supérieure en période d'anoestrus à ce qu'elle est en saison sexuelle (figure 3). Cette élévation du niveau de FSH est due probablement à une modification de l'activité intrinsèque et/ou de la sensibilité du complexe hypothalamo-hypophysaire aux stéroïdes ovariens (Findlay et Clarke, 1987).
Activité intrinsèque du complexe hypothalamo-hypophysaire. Il n'y a pas de variation saisonnière significative (P > 0,05) du niveau de FSH au cours de l'année chez la brebis Peul castrée (figure 4). La libération de FSH est en partie sous le contrôle du GnRH hypothalamique mais certaines cellules hypophysaires sont capables de libérer FSH même après déconnection de l'hypothalamus (Findlay et Clarke, 1987). On peut penser que les systèmes qui contrôlent la libération hypophysaire de FSH ne subissent pas de variation saisonnière dans leur fonctionnement chez la brebis Peul.
Sensibilité aux oestrogènes. En utilisant les mêmes critères d'évaluation de la sensibilité du complexe hypothalamo-hypophysaire aux stéroïdes que dans le cas de LH, on n'observe pas de variation saisonnière du retrocontrôle des stéroïdes ovariens sur la libération de FSH (tableau 2). La libération de FSH par l'hypophyse est également influencée par l'inhibine d'origine folliculaire (Goodman et al, 1981); or, il n'y a pas de variation saisonnière de la capacité de l'inhibine à réduire le niveau de FSH chez la brebis (Findlay et al, 1985). Il est probable que l'élévation du niveau de FSH en période d'anoestrus chez la brebis Peul est due à une faible sécrétion d'oestrogènes (et d'inhibine) d'origine folliculaire.
Figure 4. Evolution circannuelle du niveau de FSH chez deux brebis ovariectomisées au Niger.
Il existe chez la brebis Peul en zone sahélienne un rythme saisonnier des sécrétions de prolactine, LH et FSH. L'évolution circannuelle de la prolactine suit les variations de la photopériode, et celle de LH est contemporaire de l'augmentation de l'amplitude thermique journalière. La libération de ces hormones hypophysaires intervenant dans la reproduction semble dépendre directement des facteurs climatiques. Le rythme saisonnier de libération de FSH serait influencé par un rythme saisonnier de sécrétion d'oestrogène (et d'inhibine) d'origine folliculaire. La production d'oestrogène est elle-même sous le contrôle de LH.
Bligh J. 1986. La défence contre le chaud et le froid. La Recherche 177: 600-611.
Cognie Y, Gayerie F. Oldham C M, Poulin N et Mauleon P. 1981. Frequent lambing: Underlying physiology. Proceedings of the 32nd Annual Meeting of the European Association for Animal Production.
Findlay J K et Clarke I J. 1987. Regulation of the secretion of FSH in domestic ruminants. Journal of Reproduction and Fertility, Supplement 34: 27-37.
Findlay J K. Gill T W et Doughton B W. 1985. Influence of season and sex on the inhibitory effect of ovine follicular fluid and on plasma gonadotropins in gonadectomized sheep. Journal of Reproduction and Fertility 73: 329-325.
Goodman R L et Karsch F J. 1980. Control of seasonal breeding in the ewe: Importance of changes in response to sex-steroid feedback. Progress in Reproductive Biology 5: 134-154.
Goodman R L et Karsch F J. 1981. The hypothalamic pulse generator: A key determinant of reproductive cycles in sheep. In: B K Follet and D E Follett (eds.), Biological clocks in seasonal reproductive cycles. Wright, Bristol, UK.
Goodman R L, Pickover S M et Karsch F J. 1981. Ovarian feedback control of follicle stimulating hormone in the ewe: Evidence for selective suppression. Endocrinology 108: 772-777.
Hixon J E, Gengenbach D R et Hansel W. 1975. Failure of PGF2 to cause luteal regression in ewes after destruction of ovarian follicles by X-irradiation. Biology of Reproduction 13: 126-135.
Hooley R D, Findley J K et Stephenson R G A. 1979. Effect of heat stress on plasma concentrations of prolactin and luteinizing hormone in ewes. Australian Journal of Biological Science 32: 231-235.
Karsch F J. Robert L, Goodman R L and Legan J S. 1980. Feedback basis of seasonal breeding: Test of an hypothesis. Journal of Reproduction and Fertility 58: 521-535.
Knobil E. 1987. A hypothalamic pulse generator governs mammalian reproduction. News in the Physiological Sciences 2: 42-43.
Land R B. Wheeler A G et Carr W R. 1976. Seasonal variation in the oestrogen-induced LH discharge of ovariectomized Finnish Landrace and Scottish Blackface ewes. Annals of Biology, Animal Biochemistry and Biophysics 16: 521-528.
McNatty K P. Gibb M, Dobson C et Thurley D C. 1981. Evidence that changes in LH secretion regulate the growth of the preovulatory follicle in the ewe. Journal of Endocrinology 90: 375-389.
McNatty K P. Hudson N L, Henderson K M, Lun S. Heath D A, Gibb M, Ball K, McDiarmid J M et Thurley D C. 1984. Changes in gonadotrophin secretion and ovarian follicular activity in seasonal breeding sheep throughout the year. Journal of Reproduction and Fertility 70: 309-321.
Miller H L et Alliston C W. 1974. Influence of programmed circadian temperature changes upon levels of luteinizing hormone in the bovine. Biology of Reproduction 11: 187-190.
Munro C J. McNatty K P et Renshaw L. 1980. Circannual rhythms of prolactin secretion in ewes and the effect of pinealectomy. Journal of Endocrinology 84: 83-89.
Oussaid B. 1983. Etudes de l'activité ovarienne et de sa stimulation pendant l'anoestrus saisonnier chez la brebis Ile-de-France. Thèse Doctorat 3ème Cycle. Université de Paris VI, Paris, France.
Restall B J. Kearings R D, Herdegen J et Carberry P. 1978. The induction of reproductive activity in lactating ewes. Australian Journal of Agricultural Research 29: 181-187.
Riggs B L et Malven P V. 1974. Effects of intraventricular infusion of serotonim, norepinephrine and dopamine on spontaneous LH release in castrate male sheep. Biology of Reproduction 11: 587-592.
Schillo K K, Alliston C W et Malven P V. 1978. Plasma concentrations of luteinizing hormone and prolactin in the ovariectomized ewe during induced hyperthermia. Biology of Reproduction 19: 306-313.
Sheldrick E L, Mitchell M D et Flint A P F. 1980. Delayed luteal regression in ewes immunized against oxytocin. Journal of Reproduction and Fertility 59: 37-42.
Thimonier J. Ravault J P et Ortavant R. 1978. Plasma prolactin variations and cyclic ovarian activity in ewes submitted to different light regimens. Annals of Biology, Animal Biochemistry and Biophysics 18: 1229-1235.
Yenikoye A. 1984. Variations annuelles du comportement d'oestrus, du taux et des possibilités d'ovulation chez la brebis Peulh du Niger. Reproduction, Nutrition, Développement 24: 11-19.
Yenikoye A. 1986. Etudes de l'endocrinologie sexuelle et de la croissance folliculaire chez la brebis nigérienne de race Peulh: Influence de la saison de reproduction. Thèse Docteur-es-Sciences. Université de Tours, Tours, France.
Yenikoye A et Ravault J P. 1981. Rythme circannuel de sécrétion de la prolactine chez la brebis de race Peulh. Comptes Rendues de l'Académie des Sciences, Paris, Série III 293: 523-525.
Yenikoye A, Pelletier J. André D et Mariana J C. 1982. Anomalies in ovarian function of Peulh ewes. Theriogenology 17: 355-364.
Zarco L, Stabenfeldt G H. Kindahl H. Quirke J F et Granstrom E. 1984. Persistence of luteal activity in the non-pregnant ewe. Animal Reproduction Science 7: 245-267.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}