![]()
![]()
![]()
Role of vitamin A in human metabolic processes
Population at risk and consequences of vitamin A deficiency
Units of expression
Dietary sources
Evidence for making recommendations
Determining mean requirements and setting safe levels of intake
Recommended safe intakes
Toxicity
Future research
References
Vitamin A (retinol) is an essential nutrient needed in small amounts by humans for the normal functioning of the visual system; growth and development; and maintenance of epithelial cellular integrity, immune function, and reproduction. These dietary needs for vitamin A are normally provided for as preformed retinol (mainly as retinyl ester) and pro-vitamin A carotenoids.
Overview of vitamin A metabolism
Preformed vitamin A in animal foods occurs as retinyl esters of fatty acids in association with membrane-bound cellular lipid and fat-containing storage cells. Pro-vitamin A carotenoids in foods of vegetable origin are also associated with cellular lipids but are embedded in complex cellular structures such as the cellulose-containing matrix of chloroplasts or the pigment-containing portion of chromoplasts. Normal digestive processes free vitamin A and carotenoids from embedding food matrices, a more efficient process from animal than from vegetable tissues. Retinyl esters are hydrolysed and the retinol and freed carotenoids are incorporated into lipid-containing, water-miscible micellar solutions. Products of fat digestion (e.g., fatty acids, monoglycerides, cholesterol, and phospholipids) and secretions in bile (e.g., bile salts and hydrolytic enzymes) are essential for the efficient solubilisation of retinol and especially for solubilisation of the very lipophilic carotenoids (e.g., aa- and bb-carotene, bb-cryptoxanthin, and lycopene) in the aqueous intestinal milieu. Micellar solubilisation is a prerequisite to their efficient passage into the lipid-rich membrane of intestinal mucosal cells (i.e., enterocytes) (1-3). Diets critically low in dietary fat (under about 5-10 g daily) (4) or disease conditions that interfere with normal digestion and absorption leading to steatorrhea (e.g., pancreatic and liver diseases and frequent gastroenteritis) can therefore impede the efficient absorption of retinol and carotenoids. Retinol and some carotenoids enter the intestinal mucosal brush border by diffusion in accord with the concentration gradient between the micelle and plasma membrane of enterocytes. Some carotenoids pass into the enterocyte and are solubilized into chylomicrons without further change whereas some of the pro-vitamin A carotenoids are converted to retinol by a cleavage enzyme in the brush border (3). Retinol is trapped intracellularly by re-esterification or binding to specific intracellular binding proteins. Retinyl esters and unconverted carotenoids together with other lipids are incorporated into chylomicrons, excreted into intestinal lymphatic channels, and delivered to the blood through the thoracic duct (2).
Tissues extract most lipids and some carotenoids from circulating chylomicrons, but most retinyl esters are stripped from the chylomicron remnant, hydrolysed, and taken up primarily by parenchymal liver cells. If not immediately needed, retinol is re-esterified and retained in the fat-storing cells of the liver (variously called adipocytes, stellate cells, or Ito cells). The liver parenchymal cells also take in substantial amounts of carotenoids. Whereas most of the body's vitamin A reserve remains in the liver, carotenoids are also deposited elsewhere in fatty tissues throughout the body (1). Usually, turnover of carotenoids in tissues is relatively slow, but in times of low dietary carotenoid intake, stored carotenoids are mobilised. A recent study in one subject using stable isotopes suggests that retinol can be derived not only from conversion of dietary pro-vitamin carotenoids in enterocytes - the major site of bioconversion - but also from hepatic conversion of circulating pro-vitamin carotenoids (5). The quantitative contribution to vitamin A requirements of carotenoid converted to retinoids beyond the enterocyte is unknown.
Following hydrolysis of stored retinyl esters, retinol combines with a plasma-specific transport protein, retinol-binding protein (RBP). This process, including synthesis of the unoccupied RBP (apo-RBP), occurs to the greatest extent within liver cells but it may also occur in some peripheral tissues. The RBP-retinol complex (holo-RBP) is secreted into the blood where it associates with another hepatically synthesised and excreted larger protein, transthyretin. The transthyretin-RBP-retinol complex circulates in the blood, delivering the lipophilic retinol to tissues; its large size prevents its loss through kidney filtration (1). Dietary restriction in energy, proteins, and some micronutrients can limit hepatic synthesis of proteins specific to mobilisation and transport of vitamin A. Altered kidney functions or fever associated with infections (e.g., respiratory infections [6] or diarrhoea [7]) can increase urinary vitamin A loss.
Holo-RBP transiently associates with target-tissue membranes, and specific intracellular binding proteins then extract the retinol. Some of the transiently sequestered retinol is released into the blood unchanged and is recycled (i.e., conserved) (1,8). A limited reserve of intracellular retinyl esters is formed, that subsequently can provide functionally active retinol and its oxidation products (i.e., isomers of retinoic acid) as needed intracellularly. These biologically active forms of vitamin A are associated with specific cellular proteins which bind with retinoids within cells during metabolism and with nuclear receptors that mediate retinoid action on the genome (9). Retinoids modulate the transcription of several hundreds of genes (10-12). In addition to the latter role of retinoic acid, retinol is the form required for functions in the visual (13) and reproductive systems (14) and during embryonic development (15).
Holo-RBP is filtered into the glomerulus but recovered from the kidney tubule and recycled. Normally vitamin A leaves the body in urine only as inactive metabolites which result from tissue utilisation and as potentially recyclable active glucuronide conjugates of retinol in bile secretions (8). No single urinary metabolite has been identified which accurately reflects tissue levels of vitamin A or its rate of utilisation. Hence, at this time urine is not a useful biologic fluid for assessment of vitamin A nutriture.
Biochemical mechanisms for vitamin A functions
Vitamin A functions at two levels in the body. The first is in the visual cycle in the retina of the eye; the second is in all body tissues systemically to maintain growth and the soundness of cells. In the visual system, carrier-bound retinol is transported to ocular tissue and to the retina by intracellular binding and transport proteins. Rhodopsin, the visual pigment critical to dim-light vision, is formed in rod cells after conversion of all-trans retinol to retinaldehyde, isomerization to the 11-cis-form, and binding to opsin. Alteration of rhodopsin through a cascade of photochemical reactions results in ability to see objects in dim light (13). The speed at which rhodopsin is regenerated relates to the availability of retinol. Night blindness is usually an indicator of inadequate available retinol, but it can also be due to a deficit of other nutrients, which are critical to the regeneration of rhodopsin, such as protein and zinc, and to some inherited diseases, such as retinitis pigmentosa.
The growth and differentiation of epithelial cells throughout the body are especially affected by vitamin A deficiency (VAD). Goblet cell numbers are reduced in epithelial tissues. The consequence is that mucous secretions with their antimicrobial components diminish. Cells lining protective tissue surfaces fail to regenerate and differentiate, hence flatten and accumulate keratin. Both factors - the decline in mucous secretions and loss of cellular integrity - diminish resistance to invasion by potentially pathogenic organisms. The immune system is also compromised by direct interference with production of some types of protective secretions and cells (11). Classical symptoms of xerosis (drying or nonwetability) and desquamation of dead surface cells as seen in ocular tissue (i.e., xerophthalmia) are the external evidence of the changes also occurring to various degrees in internal epithelial tissues.
Current understanding of the mechanism of vitamin A action within cells outside the visual cycle is that cellular functions are mediated through specific nuclear receptors. These receptors are activated by binding with specific isomers of retinoic acid (i.e., all-trans and 9-cis retinoic acid). Activated receptors bind to DNA response elements located upstream of specific genes to regulate the level of expression of those genes (12). The synthesis of a large number of proteins vital to maintaining normal physiologic functions is regulated by these retinoid-activated genes. There also may be other mechanisms of action that are as yet undiscovered (10).
Definition of vitamin A deficiency
VAD is not simply defined. WHO defines it as tissue concentrations of vitamin A low enough to have adverse health consequences even if there is no evidence of clinical xerophthalmia (16). In addition to the specific signs and symptoms of xerophthalmia and the risk of irreversible blindness, non-specific symptoms include increased morbidity and mortality, poor reproductive health, increased risk of anaemia, and contributions to slowed growth and development. Because these non-specific adverse consequences may occur from other nutrient deficits as well, it is difficult to attribute non-ocular symptoms specifically to VAD in the absence of biochemical measurements reflective of vitamin A status.
Geographic distribution and magnitude
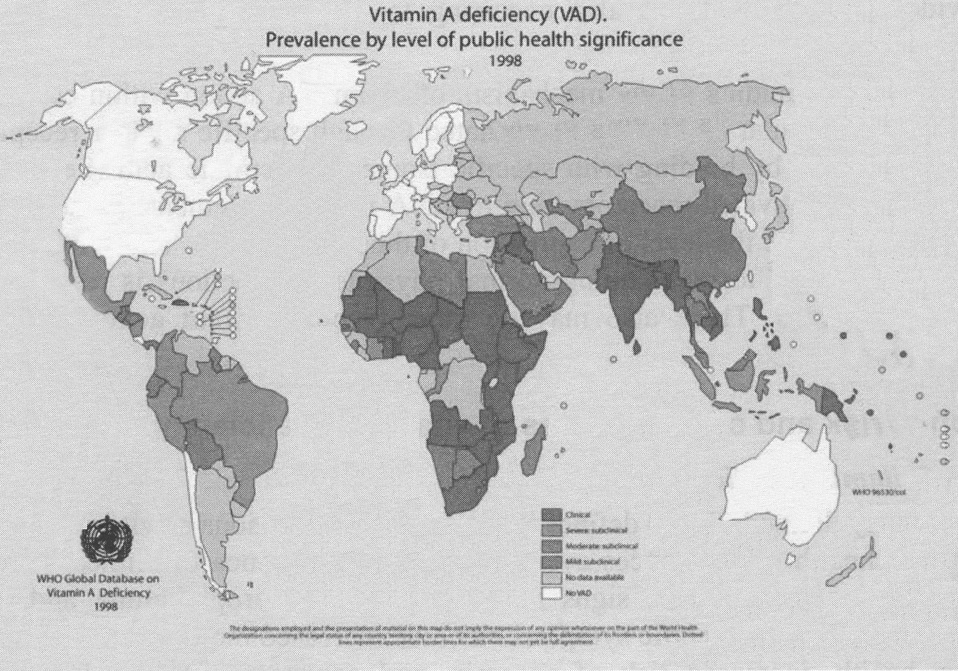
In 1996 WHO mapped the global distribution of VAD (Figure 9) and categorised countries by degree of significance as a public health problem on the basis of both clinical and moderate and severe sub-clinical (prevalence of low blood levels of retinol) indicators of deficiency (16,18). In the early 1990s, WHO estimated that about 3 million children had some form of xerophthalmia annually and, on the basis of blood levels, another 250 million were sub-clinically deficient (18). The magnitude of the sub-clinical estimate is currently being re-evaluated to quantitatively establish a benchmark for measuring prevalence trends. The actual number of sub-clinical deficiencies based on the prevalence of low serum levels of retinol, however, remains uncertain because of the confounding and poorly quantitative role of infections (see later discussion).
Epidemiologic studies repeatedly report clustering of VAD, presumably resulting from concurrent occurrence of several risk factors. This clustering may occur among both neighbourhoods and households (19).
Figure 9. Prevalence of VAD in the world (17)
Age and gender
VAD can occur in individuals of any age. However, it is a disabling and potentially fatal public health problem for children under 6 years of age. VAD-related blindness is most prevalent in children under 3 years of age (20). This period is characterised by high requirements for vitamin A to support early rapid growth, the transition from breast-feeding to dependence on other dietary sources of the vitamin, and increased frequency of respiratory and gastrointestinal infections. The increased mortality risk from concurrent infections extends at least to 6 years of age and is associated with both clinical and sub-clinical VAD (21). There is little information regarding the health consequences of VAD in school-age children. The prevalence of Bitot's spots (i.e., white foamy patches on the conjunctiva) may be highest in this age group but their occurrence may reflect past more than current history of VAD (22). Women of reproductive age are thought also to be vulnerable to VAD during pregnancy and lactation because they often report night blindness (23,24) and because their breast-milk is frequently low in vitamin A (25,26). Not all night blindness in pregnant women, however, responds to vitamin A (24).
There is no consistent, clear indication in humans of a gender differential in vitamin A requirements during childhood. Growth rates and presumably the need for vitamin A from birth to 10 years for boys are consistently higher than those for girls (27). In the context of varied cultural and community settings, however, variations in gender-specific child-feeding and care practices are likely to subsume a small gender differential in requirements to account for reported gender differences and prevalence of xerophthalmia. Pregnant and lactating women require additional vitamin A to support maternal and foetal tissue growth and lactation losses, additional vitamin A which is not needed by other post-adolescent adults (28).
Risk factors
VAD is most common in populations consuming most of their vitamin A needs from pro-vitamin carotenoid sources and where minimal dietary fat is available (29). About 90 percent of ingested preformed vitamin A is absorbed, whereas the absorption efficiency of pro-vitamin A carotenoids varies widely depending on the type of plant source and the fat content of the accompanying meal (30). Where it is possible to increase dietary fat, this will likely improve the absorption of vitamin A activity from the diet.
In areas with endemic VAD, fluctuations in the incidence of VAD throughout the year reflect the balance between intake and need. Periods of general food shortage (and specific shortages in vitamin A-rich foods), peak incidence of common childhood infectious diseases (diarrhoea, respiratory infections, and measles), and periodic seasonal growth spurts affect the balance. Seasonal food availability can influence VAD prevalence by directly influencing access to pro-vitamin A sources; for example, the scarcity of mangoes in hot arid months followed by the glutting of the market with mangoes during harvest seasons (31). Seasonal growth spurts in children frequently follow seasonal postharvest increases in energy and macronutrient intakes. These increases are usually obtained from staple grains (e.g., rice) and tubers (e.g., light-coloured yams) which are not good sources of some micronutrients (e.g., vitamin A to support the growth spurt) (32).
Food habits and taboos often restrict consumption of potentially good food sources of vitamin A (e.g., mangoes and green leafy vegetables). Culture-specific factors for feeding children, adolescents, and pregnant and lactating women are common (29,33-35). Illness- and childbirth-related proscription of the use of specific foods pervade in many traditional cultures (36). Such influences alter short- and long-term food distribution within families. However, some cultural practices can be protective of vitamin A status and they need to be identified and reinforced.
Morbidity and mortality
The consequences of VAD are manifested differently in different tissues. In the eye, the symptoms and signs, together referred to as xerophthalmia, have a long, well-recognised history and have until recently been the basis for estimating the global burden from the disease (20). Although ocular symptoms and signs are the most specific indicators of VAD, they occur only after other tissues have impaired functions that are less specific and less easily assessed.
The prevalence of ocular manifestations (i.e., xerophthalmia or clinical VAD) is now recognised to far underestimate the magnitude of the problem of functionally significant VAD. Many more preschool-age children and perhaps older children and women who are pregnant or lactating have their health compromised when they are sub-clinically deficient. In young children, sub-clinical deficiency, like clinical deficiency, increases the severity of some infections, particularly diarrhoea and measles, and the risk of dying (21,37). The incidence (38) and prevalence (39) of diarrhoea may also increase with sub-clinical VAD. Meta-analyses conducted by three independent groups using data from several randomised trials provide convincing evidence that community-based improvement of the vitamin A status of deficient children 6 months to 6 years of age reduces their risk of dying by 20-30 percent on average (21,40,41). Mortality in children who are blind from keratomalacia or who have corneal disease is reported to be from 50 percent to 90 percent (20,42) and measles mortality associated with VAD is increased up to 50 percent (43). Limited data are available from controlled studies of the possible link between morbidity history and vitamin A status of pregnant and lactating women (44).
There are discrepancies in the linkage between incidence and severity of infectious morbidity of various aetiologies and vitamin A status. The weight of evidence supports an association of VAD with severity of an infection once acquired, except for respiratory diseases, which are nonresponsive (16,37-39,45). The severity of the pneumonia associated with measles, however, is an exception because it decreases with treatment with vitamin A supplements (43,46).
Infectious diseases depress circulating retinol and contribute to vitamin A depletion. Enteric infections may alter absorptive-surface area, compete for absorption-binding sites, and increase urinary loss (7,47,48). Febrile systemic infections also increase urinary loss (6,49) and metabolic utilisation rates and may reduce apparent retinol stores if fever occurs frequently (50). In the presence of latent deficiency, disease occurrence is often associated with precipitating ocular signs (51,52). Measles virus infection is especially devastating to vitamin A metabolism, adversely interfering with both efficiencies of utilisation and conservation (43,52,53). Severe protein-energy malnutrition affects many aspects of vitamin A metabolism, and even when some retinyl ester stores are still present, malnutrition - often coupled with infection - can prevent transport-protein synthesis, that results in immobilisation of existing vitamin A stores (54).
The compromised integrity of the epithelium, together with the possible alteration in hormonal balance at severe levels of deficiency, impairs normal reproductive functions in animals (9,14,15,25,55,56). Controlled human studies are, of course, lacking. In animals and Humans, congenital anomalies can result if the foetus is exposed to severe deficiency or large excesses of vitamin A at critical periods early in gestation (first trimester) when foetal organs are being formed (25,57). Reproductive performance measured by infant outcomes in one community-based clinical intervention trial, however, was not influenced by vitamin A status (44).
The growth of children may be impaired by VAD. Interventions with vitamin A only have not consistently demonstrated improved growth in community studies because VAD seldom occurs in isolation of other nutrient deficiencies that also affect growth and may be more limiting (58).
A lack of vitamin A can affect iron metabolism when deficiencies of both nutrients coexist and particularly in environments that favour frequent infections (59). Maximum haemoglobin response occurs when iron and vitamin A deficiencies are corrected together (60). VAD appears to influence the availability of storage iron for use by haematopoietic tissue (60,61). However, additional research is needed to clarify the mechanisms of the apparent interaction.
In blood, tissues, and human milk, vitamin A levels are conventionally expressed in µg/dL or µmol/l of all-trans retinol. Except for postprandial conditions, most of the circulating vitamin A is retinol whereas in most tissues (such as the liver), secretions (such as human milk), and other animal food sources it exists mainly as retinyl esters, that are usually hydrolysed before analytical detection.
To express the vitamin A activity of carotenoids in diets on a common basis, a joint FAO/WHO Expert Group (62) in 1967 introduced the concept of the retinol equivalent (RE) and established the following relationships among food sources of vitamin A:
|
1 µg retinol |
= 1 RE |
|
1 µg b-carotene |
= 0.167 µg RE |
|
1 µg other pro-vitamin A carotenoids |
= 0.084 µg RE |
Retinol equivalents in a diet are calculated as the sum of the weight of the retinol portion of preformed vitamin A with the weight of b-carotene divided by its conversion factor and with the weight of other carotenoids divided by their conversion factor (69). Most recent food composition tables report b-carotene and sometimes other pro-vitamin A carotenoids as µg/g edible portion. However, older food composition tables frequently report vitamin A as international units (IUs). The following applies to determining comparable values as µg:
|
1 IU retinol |
= 0.3 µg retinol |
|
1 IU b-carotene |
= 0.6 µg b-carotene |
|
1 IU retinol |
= 3 IU b-carotene |
Preformed vitamin A is found almost exclusively in animal products, such as human milk, glandular meats, liver and fish liver oils (especially), egg yolk, and whole milk and dairy products. Preformed vitamin A is also used to fortify processed foods, that may include sugar, cereals, condiments, fats, and oils (70). Pro-vitamin A carotenoids are found in green leafy vegetables (e.g., spinach, amaranth, and young leaves from various sources), yellow vegetables (e.g., pumpkins, squash, and carrots), and yellow and orange noncitrus fruits (e.g., mangoes, apricots, and papaya). Red palm oil produced in several countries worldwide is especially rich in pro-vitamin A (71). Some other indigenous plants also may be unusually rich sources of pro-vitamin A. Such examples are the palm fruit known in Brazil as burití, that is found in areas along the Amazon (as well as elsewhere in Latin America) (72), and the fruit known as gac in Vietnam, that is used to colour rice, particularly on ceremonial occasions (73). Foods containing pro-vitamin A carotenoids tend to be less biologically available but more affordable than animal products. It is mainly for this reason that carotenoids provide most of the vitamin A activity in the diets of economically deprived populations.
Dietary intake and patterns
Vitamin A status cannot be assessed from dietary intake alone, but dietary intake assessment can provide evidence of risk of an inadequate status. Quantitative collection of dietary information is fraught with measurement problems. These problems arise both from obtaining representative quantitative dietary histories from individuals, communities, or both and from interpreting these data while accounting for differences in bio-availability, preparation losses, and variations in food composition data among population groups (70). This is especially difficult in populations consuming most of their dietary vitamin A from pro-vitamin carotenoid sources. Simplified guidelines have been developed recently in an effort to improve the obtaining of reliable dietary intake information from individuals and communities (68,74).
World and regional supply and patterns
In theory the world’s food supply is sufficient to meet global requirements. Great differences exist, however, in the available sources (animal and vegetable) and in per capita consumption of the vitamin among different countries, age categories, and socio-economic groups. VAD as a global public health problem, therefore, is largely due to inequitable food distribution among and within countries and households in relation to need for ample bio-available vitamin A sources (75,76).
Earlier FAO global estimates in 1984 indicated that preformed vitamin A constituted about one-third of total dietary vitamin A activity (69). World availability of vitamin A for human consumption at that time was approximately 220 µg of preformed retinol per capita daily and 560 µg RE from pro-vitamin carotenoids (about 3400 µg carotenoids for a 6:1 conversion factor) per person per day, for a total of about 790 µg RE. These values are based on supply estimates and not consumption estimates. Losses commonly occur during food storage and processing, both industrially and in the home (70).
The estimated available regional supply of vitamin A from a more recent global evaluation (76) shown in Table 16 illustrates the variability in amounts and sources of vitamin A. The variability is further complicated by access to the available supply, that varies with household income, poverty being a yardstick for risk of VAD. VAD is most prevalent in Southeast Asia, Africa, and the Western Pacific, where vegetable sources contribute nearly 80 percent or more of the available supply of retinol equivalents. Furthermore, in Southeast Asia the total available supply is about half of that of most other regions and is particularly low in animal sources. In contrast, the Americas, Europe, and Eastern Mediterranean regions have a supply ranging from 800 to 1000 mmg RE/day, one-third of which comes from animal sources. Recent national data from the USA Continuing Survey of Food Consumption (77) and the National Health and Nutrition Examination Survey (78) included mean dietary intakes of children 0-6 years of age of 864 ± 497 and 921 ± 444 mmg RE daily. In the Dietary and Nutritional Survey of British Adults (79), the median intake of men and women 35-49 years old was 1118 mmg RE and 926 mmg RE, respectively, which corresponded to serum retinol concentrations of 2.3 mmol/l and 1.8 mmol/l, respectively. In another selected survey in the United Kingdom, median intakes for nonpregnant women who did not consume liver or liver products during the survey week were reported to be 686 mmg RE daily (80).
The available world supply figures in Table 16 were recently reassessed based on a bio-availability ratio of 1:30 for retinol to other pro-vitamin A carotenoids (81). This conversion factor was justified on the basis of one published controlled intervention study conducted in Indonesia (82) and a limited number of other studies not yet published in full. Applying the unconfirmed conversion factor to the values in Table 16 would lead to the conclusion that regional and country needs for vitamin A could not be met from predominantly vegetarian diets. This is inconsistent with the preponderance of epidemiologic evidence. Most studies report a positive response when vegetable sources of pro-vitamin A are given under controlled conditions to deficient subjects freed of confounding parasite loads and provided with sufficient dietary fat (83,84). Emerging data are likely to justify a lower biologic activity for pro-vitamin A carotenoids because of the mix of total carotenoids found in food sources in a usual meal (67,68). This Consultation concluded that the 1:6 bioconversion factor originally derived on the basis of balance studies should be retained until there is firm confirmation from ongoing studies that use more precise methodologies.
Table 16
Available supply of vitamin A by WHO region
|
Region |
Total mg
RE/day |
Animal sources mg
RE/day |
Vegetable sources mg
RE/day |
|
Africa |
775 |
122 |
654 (84)a |
|
Americas |
814 |
295 |
519 (64) |
|
Southeast Asia |
431 |
53 |
378 (90) |
|
Europe |
738 |
271 |
467 (63) |
|
Eastern Mediterranean |
936 |
345 |
591 (63) |
|
Western Pacific |
997 |
216 |
781 (78) |
|
Total |
782 |
212 |
565 (72) |
Source: ACC/SCN, 1993 (76).
Indicators of vitamin A deficiency
Ocular signs of VAD are measured by clinical examination and history and are quite specific in preschool-age children. However, these are rare occurrences that require examination of large populations to obtain incidence and prevalence data. Sub-clinical VAD is more prevalent, requiring smaller sample sizes to obtain valid prevalence estimates (16).
A full description of clinical indicators with coloured illustrations for each is found in the WHO Field Guide (20). The most frequently occurring is night blindness, which is the earliest manifestation of xerophthalmia. In mild form it is generally noticeable after stress from a bright light that bleaches the rhodopsin (visual purple) found in the retina. VAD prolongs the time to regenerate rhodopsin, thus delays adaptation time in dark environments. Night-blind young children tend to stumble when going from bright to dimly lighted areas and they, as well as night-blind mothers, tend to remain inactive at dusk and at night (85). No field-applicable objective tool is currently available for measuring night blindness in children under about 3 years of age. It can be measured by history in certain cultures (86). When night blindness is prevalent, many cultures coin a word descriptive of the characteristic symptom that they can reliably recall on questioning, making this a useful tool for assessing the prevalence of VAD (87). Questioning for night blindness is not consistently a reliable assessment measure where a local term is absent. In addition, there is no clearly defined blood retinol level that is directly associated with occurrence of the symptom. Vitamin A - related night blindness, however, responds rapidly, usually within 1-2 days, to administration of vitamin A.
Sub-clinical vitamin A deficiency
Direct measurement of concentrations of vitamin A in the liver where it is stored or in the total body pool relative to known specific vitamin A-related functions (e.g., night blindness) would be the indicator of choice for determining requirements. This cannot be done with the methodology now available for population use. There are several practical biochemical methods for estimating sub-clinical vitamin A status but all have limitations (16,86,88,89). Each method is useful to identify deficient populations, but no one of these indicators is definitive or directly related quantitatively to disease occurrence. The indicators of choice are listed in Table 17. These indicators are less specific to VAD than clinical eye signs and less sensitive for measuring sub-clinical vitamin A status. WHO recommends that where feasible at least two sub-clinical biochemical indicators, or one biochemical and a composite of non-biochemical risk factors, should be measured and that both types of indicators should point to deficiency in order to identify populations at high risk of VAD (16). Cut-off points given in Table 17 represent the consensus gained from practical experience comparing populations with some evidence of VAD with those without VAD. There are no field studies that quantitatively relate the prevalence of adverse health symptoms (e.g., incidence or prevalence of severe diarrheal disease) and relative levels of biologic indicator cut-off values. Furthermore, each of the biochemical indicators listed is subject to confounding factors, which may be unrelated to vitamin A status (e.g., infections).
Table 17
Biochemical indicators of sub-clinical VAD in mothers and in children 6-71 months of age
|
Indicator |
Cut-off to indicate deficiency |
|
Night blindness (24-71 mo) |
³ 1% report a history of night
blindness |
|
Biochemical |
|
|
Breast milk retinol |
|
|
|
£1.05 mmol/l |
|
|
(£ 8 mg/g milk fat) |
|
Serum retinol |
£ 0.70 mmol/l |
|
Relative dose response |
³ 20% |
|
Modified relative dose response |
ratio ³ 0.06 |
Although all biochemical indicators currently available have limitations, the biochemical indicator of choice for population assessment is the distribution of serum levels of vitamin A (serum retinol). Only at very low blood levels (<0.35 mmol/l) is there an association with corneal disease prevalence (90). Blood levels between 0.35 and 0.70 mmol/l are likely to characterise sub-clinical deficiency (91), but sub-clinical deficiency may still be present at levels between 0.70 and 1.05 mmol/l and occasionally above 1.05 mmol/l (92). The prevalence of values below 0.70 mmol/l is a generally accepted population cut-off for preschool-age children to indicate risk of inadequate vitamin A status (16) and above 1.05 mmol/l to indicate an adequate status (93,94). As noted elsewhere, clinical and sub-clinical infections can lower serum levels of vitamin A on average as much as 25 percent independently of vitamin A intake (95,96). Therefore, at levels between about 0.5 and 1.05 mmol/l, the relative dose response or modified relative dose response test on a subsample of the population can be useful for identifying the prevalence of critically depleted body stores when interpreting the left portion of serum retinol distribution curves.
Requirement and safe level of intake for vitamin A in this report do not differ significantly from those of the 1988 FAO/WHO Expert Consultation (69) except for adapting to the age categories defined in this consultation and during pregnancy. The term safe level of intake used in the 1988 report is retained in this report because the levels in Table 18 do not strictly correspond to the definition of a recommended nutrient intake.
The mean requirement for an individual is defined as the minimum daily intake of vitamin A as presented in mmg retinol equivalents (mg RE) to prevent xerophthalmia in the absence of clinical or sub-clinical infection. This intake should account for proportionate bio-availability of preformed vitamin (about 90 percent) and pro-vitamin A carotenoids from a diet that contains sufficient fat (e.g., at least 5-10 g). Bio-availability of carotenoids varies widely by source (e.g., fibrous green leafy vegetables or soft-tissue fruits). The required level of intake is set to prevent clinical signs of deficiency, allow for normal growth, and reduce the risk of vitamin A - related severe morbidity and mortality on a population basis. It does not allow for frequent or prolonged periods of infections or other stresses.
The safe level of intake for an individual is defined as the average continuing intake of vitamin A required to permit adequate growth and other vitamin A-dependent functions and to maintain an acceptable total body reserve of the vitamin. This reserve helps offset periods of low intake or increased need resulting from infections and other stresses. Useful indicators include a plasma retinol concentration above 0.70 mmol/l, that is associated with a relative dose response below 20 percent, or a modified relative dose response below 0.06. For lactating women, breast-milk retinol levels above 1.05 mmol/l (or above 8 mg/g milk fat) are considered to reflect minimal maternal stores because levels above 1.75 mmol/l are common to populations known to be healthy and without evidence of insufficient dietary vitamin A (25,26).
Infants and children
Vitamin A requirements for infants are calculated from the vitamin A provided in human milk. During at least the first 6 months of life, exclusive breast-feeding can provide sufficient vitamin A to maintain health, permit normal growth, and maintain sufficient stores in the liver (97).
Reported retinol concentrations in human milk varies widely from country to country (0.70-2.45 mmol/l). In some developing countries the vitamin A intake of breast-fed infants who grow well and do not show signs of deficiency is from 120 to 170 mg RE/day (26,97). Such intakes are considered adequate to cover infant requirements if the infant’s weight is assumed to be at least at the 10th percentile according to WHO standards (69). However, this intake is unlikely to build adequate body stores because xerophthalmia is common in preschool-age children in the same communities with somewhat lower intakes. Because of the need for vitamin A to support the growth rate of infancy, which can vary considerably, a requirement estimate of 180 mg RE/day seems appropriate.
The safe level for infants up to 6 months of age is based on observations of breast-fed infants in communities in which good nutrition is the norm. Average consumption of human milk by such infants is about 750 ml/day during the first 6 months (97). Assuming an average concentration of vitamin A in human milk of about 1.75 mmol/l, the mean daily intake would be about 375 mg RE, which is therefore the recommended safe level. From 7-12 months, human milk intake averages 650 ml, which would provide 325 µg vitamin A daily. Because breast-fed infants in endemic vitamin A-deficient populations are at increased risk of death from 6 months onward, the requirement and recommended safe intake are increased to 190 mg and 400 mg, respectively.
The requirement (with allowance for variability) and the recommended intake for older children may be estimated from those derived for late infancy (i.e., 20 and 39 mg RE/kg body weight/day) (69). On this basis, including allowances for storage requirements and variability, requirements for preschool children would be in the range of 200-400 mg RE daily. In poor communities where children 1-6 years old are reported to have intakes of about 100-200 mg RE/day, signs of VAD do occur; in southern India these signs were relieved and risk of mortality was reduced when the equivalent of 350-400 mg RE/day was given to children weekly (98). In the United States most preschool-age children maintain serum retinol levels of 0.70 mmol/l or higher while consuming diets providing 300-400 mg RE/day (from the databank for the third National Health and Nutrition Examination Survey).
Adults
Estimates for the requirements and recommended safe intakes for adults are also estimated from those derived for late infancy, i.e. 4.8 and 9.3 mg RE/kg body weight/day (69). Detailed accounts for arriving at the requirement for vitamin A is provided in the FAO/WHO report of 1988 (69) and will not be repeated here because there are no new published studies to indicate a need to revise the assumptions on which the calculations were based. The safe intakes recommended are consistent with the per capita vitamin A content in the food supply of countries that show adequate vitamin A status in all sectors of the population. Additional evidence that the existing safe level of intake is adequate for adults on a population basis is provided by an analysis of dietary data from the 1990 survey of British adults in whom there was no evidence of VAD (79). The median intake for another survey of nonpregnant UK women who did not consume liver or liver products during the survey week was 686 mg/day (80). This value is substantially above the estimated mean requirement for pregnant women and falls quite short of the amount in which teratology risk is reported (99-101). About one-third of the calculated retinol equivalents consumed by the British women came from pro-vitamin A sources (20 percent from carrots).
Pregnancy
During pregnancy additional vitamin A is needed for the growth and maintenance of the foetus for providing a limited reserve in the foetal liver and for maternal tissue growth. There are no reliable figures available for the specific vitamin A requirements for these processes (28).
Newborn infants need around 100 mg retinol daily to meet their needs for growth. During the third trimester the foetus grows rapidly and, although obviously smaller in size than the infant born full term, the foetus presumably has similar needs. Incremental maternal needs associated with pregnancy are assumed to be provided from maternal reserves in populations of adequately nourished healthy mothers. In populations consuming at the basal requirement, an increment of 100 mg/day during the full gestation period should enhance maternal storage during early pregnancy and allow adequate amounts of vitamin A for the rapidly growing foetus in late pregnancy. However, this increment may be minimal for women who normally ingest only the basal requirement level of vitamin A inasmuch as the needs and growth rate of the foetus will not be affected by the mother’s initial vitamin A reserves.
A recent study in Nepal (44), where night blindness is prevalent in pregnant women, provided 7000 µg RE (about 23,300 IU) weekly to pregnant and lactating women (equivalent to 1000 µg RE/day). This level of intake normalised serum levels of vitamin A and was associated with a decrease in prevalence of night blindness and a decrease in maternal mortality. The findings of this study need to be confirmed. In the interim period, however, it seems prudent, recognising that a large portion of the world’s population of pregnant women lives under conditions of deprivation, to increase by 200 µg the recommended safe level to ensure adequacy of intake during pregnancy. Because therapeutic levels of vitamin A are generally higher than preventive levels, the safe intake level recommended during pregnancy is 800 mg RE/day. Women who are or who might become pregnant should carefully limit their total daily vitamin A intake to a maximum of 3000 mg RE (10 000 IU) to minimise risk of foetal toxicity (102).
Lactation
If the amounts of vitamin A recommended for infants are supplied by human milk, mothers should absorb at least as much in their diets to replace maternal losses. Thus, the increments in basal and recommended intakes during lactation are 180 mg RE and 350 mg RE, respectively, for the safe recommended intake per day. After the infant reaches the age of 6 months or when solid foods are introduced, the mother’s need for additional amounts of vitamin A lessens.
Elderly
There is no indication that the vitamin A requirements of healthy elderly individuals differs from those of other adults. It should be remembered, however, that diseases that impede vitamin A absorption, storage, and transport might be more common in the elderly than in other age groups.
Table 18 provides the estimated mean requirements for vitamin A and the recommended safe intakes, taking into account the age and gender differences in mean body weights. For most values the true mean and variance are not known. Values in the table have been rounded. It should be noted that there are no adequate data available to derive mean requirements for any group and, therefore, a recommended nutrient intake cannot be calculated. However, information is available on cures achieved in a few vitamin A-deficient adult men and on the vitamin A status of groups receiving intakes that are low but nevertheless adequate to prevent the appearance of deficiency-related syndromes. The figures for mean dietary requirements are derived from these, with the understanding that the curative dose is higher than the preventive dose. They are at the upper limits of the range so as to cover the mean dietary requirements of 97.5 percent of the population (69).
In calculating the safe intake, a normative storage requirement was calculated as a mean for adults equivalent to 434 mg RE/day, and the recommended safe intake was derived in part by using this value + 2 standard deviations. It is doubtful that this value can be applied to deal with growing children. The safe intake for children was compared with the distribution of intakes and comparable serum vitamin A levels reported for children 0-6 years of age from the United States and with distributions of serum levels of vitamin A of children 9-62 months in Australia (103), where evidence of VAD is rare.
Table 18
Estimated mean requirement and safe level of intake for vitamin A
|
Age group |
Mean requirement µg RE/day |
Recommended safe intake µg RE/day |
|
|
Infants and children |
|
|
|
|
|
0-6 months |
180 |
375 |
|
|
7-12 months |
190 |
400 |
|
|
1-3 years |
200 |
400 |
|
|
4-6 years |
200 |
450 |
|
|
7-years |
250 |
500 |
|
Adolescents, 10-18 years |
330-400 |
600 |
|
|
Adults |
|
|
|
|
|
Females,19-65 years |
270 |
500 |
|
|
Males, 19-65 years |
300 |
600 |
|
65+ |
300 |
600 |
|
|
Pregnant women |
370 |
800 |
|
|
Lactating women |
450 |
850 |
|
Because vitamin A is fat soluble and can be stored, primarily in the liver, routine consumption of large amounts of vitamin A over a period of time can result in toxic symptoms, including liver damage, bone abnormalities and joint pain, alopecia, headaches and vomiting, and skin desquamation. Hypervitaminosis A appears to be due to abnormal transport and distribution of vitamin A and retinoids caused by overloading of the plasma transport mechanisms (104).
The smallest daily supplement associated with liver cirrhosis that has been reported is 7500 µg taken for 6 years (100,101). Very high single doses can also cause transient acute toxic symptoms that may include bulging fontanels in infants; headaches in older children and adults; and vomiting, diarrhoea, loss of appetite, and irritability in all age groups. Rarely does toxicity occur from ingestion of food sources of preformed vitamin A. When this occurs, it usually results from very frequent consumption of liver products. Toxicity from food sources of pro-vitamin A carotenoids is not reported except for the cosmetic yellowing of skin.
Infants, including newborns (105), administered single doses equivalent to 15 000-30 000 mg retinol (50 000-100 000 IU) in oil generally show no adverse symptoms. However, daily prophylactic or therapeutic doses should not exceed 900 mg, that is well above the mean requirement of about 200 mg daily for infants. An excess of bulging fontanels occurred in infants under 6 months of age in one endemically deficient population given two or more doses of 7500 mg or 15 000 mg preformed vitamin A in oil (106,107), but other large-scale controlled clinical trials have not reported excess bulging after three doses of 7500 mg given with diptheria-pertussis-tetanus immunisations at about 6, 10, and 14 weeks of age (108). No effects were detected at 3 years of age that related to transient vitamin A-induced bulging that had occurred before 6 months of age (105,109).
Most children 1-6 years of age tolerate single oral doses of 60 000 mg (200 000 IU) vitamin A in oil at intervals of 4-6 months without adverse symptoms (100). Occasionally diarrhoea or vomiting is reported but is transient with no lasting sequelae. Older children seldom experience toxic symptoms unless they habitually ingest vitamin A in excess of 7500 mg (25 000 IU) for prolonged periods of time (100).
When taken by women at early stages of gestation at daily levels of more than 7500 mg (25 000 IU), foetal anomalies and poor reproductive outcomes are reported (101). One report suggests an increased risk of teratogenicity at intakes as low as 3000 mg (10 000 IU) but this is not confirmed by other studies (102). Women who are pregnant or might become pregnant should avoid taking excessive amounts of vitamin A. A careful review of the latest available information by a WHO Expert Group recommended that daily intakes in excess of 3000 mg (10 000 IU) or weekly intakes in excess of 7500 mg (25 000 IU), should not be taken at any period during gestation (102). High doses of vitamin A (60 000 mg, or 200 000 IU) can be safely given to breast-feeding mothers for up to 2 months postpartum and for 6 weeks for women who are not breast-feeding.
Further research is needed:
1. Blomhoff, R. 1991. Vitamin A metabolism: new perspectives on absorption, transport, and storage. Physiol. Revs., 71: 951-990.
2. Ong, D.E. 1994. Absorption of vitamin A. In: Blomhoff R, ed. Vitamin A in Health and Disease. p. 37-72. New York, Marcel Dekker, Inc.
3. Parker, RS. Absorption, metabolism, and transport of carotenoids. FASEB J., 1996, 10: 542-551.
4. Jayarajan, P., Reddy, V. & Mohanram, M. 1980. Effect of dietary fat on absorption of bb-carotene from green leafy vegetables in children. Indian J. Med. Res., 71: 53-56.
5. Novotny, J.A., Dueker, S.R., Zech, L.A. & Clifford, A.J. 1995. Compartmental analysis of the dynamics of bb-carotene metabolism in an adult volunteer. J. Lip. Res., 36: 1825-1838.
6. Stephensen, C.B. 1994. Vitamin A is excreted in the urine during acute infection. Am. J. Clin. Nutr., 60: 388-392.
7. Alvarez, J.O. 1995. Urinary excretion of retinol in children with acute diarrhea. Am. J. Clin. Nutr., 61: 1273-1276.
8. Green, M.H. & Green, J.B. 1994. Dynamics and control of plasma retinol. In: Blomhoff R, ed. Vitamin A in Health and Disease. P. 119-133. New York, Marcel Dekker, Inc.
9. Ross, C. & Gardner, E.M. 1994. The function of vitamin A in cellular growth and differentiation, and its roles during pregnancy and lactation. In: Allen L, King J., Lönnerdal B, eds. Nutrient Regulation during Pregnancy, Lactation, and Infant Growth. P. 187-200. New York, Plenum Press.
10. Chambon, P. 1996. A decade of molecular biology of retinoic acid receptors. FASEB J., 10: 940-954.
11. Ross, A.C. & Stephensen, C.B. 1996. Vitamin A and retinoids in antiviral responses. FASEB J., 10: 979-985.
12. Pemrick, S.M., Lucas, D.A. & Grippo, J.F. 1994. The retinoid receptors. Leukemia. 3: S1-10.
13. Rando, R.R. 1994. Retinoid isomerization reactions in the visual system. In: Blomhoff R, ed. Vitamin A in Health and Disease. p.503-529. New York, Marcel Dekker, Inc.
14. Eskild, L.W. & Hansson, V. 1994. Vitamin A functions in the reproductive organs. In: Blomhoff R, ed. Vitamin A in Health and Disease. p. 531-559.
15. Morriss-Kay, G.M. & Sokolova, N. 1996. Embryonic development and pattern formation. FASEB J., 10: 961-968.
16. WHO/UNICEF. Indicators of VAD and their use in monitoring intervention programmes. WHO/NUT/96.10. pp. 66. World Health Organization, Geneva.
17. WHO. 1998. Vitamin A Deficiency (VAD) Prevalence by level of public health significance (map) . WHO Global Database on Vitamin A Deficiency.
18. WHO. 1995. Global prevalence of vitamin A deficiency. MDIS Working Paper #2. World Health Organization, Geneva.
19. Katz, J. 1993. Clustering of xerophthalmia within households and villages. Int. J. Epidemiol., 22: 709-715.
20. Sommer, A. 1994. VAD and its consequences: A field guide to their detection and control. 3rd ed. Geneva, World Health Organization, 1994.
21. Beaton, G.H. 1993. Effectiveness of vitamin A supplementation in the control of young child morbidity and mortality in developing countries. ACC/SCN State-of-the-art Series, nutrition policy discussion paper no. 13. United Nations Administrative Committee on Coordination, Subcommittee on Nutrition. Geneva, World Health Organization.
22. Sommer, A., Emran, N. & Tjakrasudjatma, S. 1980. Clinical characteristics of vitamin A responsive and nonresponsive Bitot's spots. Am. J. Ophthalmol., 90: 160-171.
23. Bloem, M.W., Matzger, H. & Huq, N. 1994. Vitamin A deficiency among women in the reproductive years: an ignored problem. Proceedings of the 16th IVACG Meeting, 24-28 October, 1994, Chiang Rai, Thailand.
24. Christian, P. 1998. Night blindness of pregnancy in rural Nepal-nutritional and health risks. Int. J. Epidemiol., 27: 231-237.
25. Wallingford, J.C. & Underwood, B.A. 1986. Vitamin A deficiency in pregnancy, lactation, and the nursing child. In: Baurenfeind JC, ed. Vitamin A deficiency and its control. p.101-152. New York, Academic Press.
26. Newman, V. 1994. Vitamin A and breast-feeding: a comparison of data from developed and developing countries. Food and Nutrition Bulletin, Series: 161-176.
27. WHO. 1995. Physical status: The use and interpretation of anthropometry. Report of a WHO Expert Committee. WHO Technical Report. Series 854. Geneva, World Health Organization.
28. National Academy Sciences, Food and Nutrition Board, Institute of Medicine. 1990. Nutrition during pregnancy. Part II. Nutrient supplements. p. 336-341. Washington, DC, National Academy Press.
29. Mele, L. 1991. Nutritional and household risk factors for xerophthalmia in Aceh, Indonesia: a case-control study. Am. J. Clin. Nutr., 53: 460-1465.
30. Erdman, J. Jr. 1988. The physiologic chemistry of carotenes in man. Clin. Nutr., 7: 101-106.
31. Marsh, R.R., Talukder, A., Baker, S.K. & Bloem, M.W. 1995. Improving food security through home gardening: A case study from Bangladesh. In: Technology for rural homes: research and extension experiences . UK, AERDD, University of Reading.
32. Sinha, D.P. & Bang, F.B. 1973. Seasonal variation in signs of vitamin A deficiency in rural West Bengal children. Lancet, ii: 228-231.
33. Johns, T., Booth, S.L. & Kuhnlein, H.V. 1992. Factors influencing vitamin A intake and programmes to improve vitamin A status. Food and Nutrition Bulletin, 14: 20-33.
34. Tarwotjo, I. 1982. Dietary practices and xerophthalmia among Indonesian children. Am. J. Clin. Nutr., 35: 574-581.
35. Zeitlan, M.F. 1992. Mothers' and children's intakes of vitamin A in rural Bangladesh. Am. J. Clin. Nutr., 56: 136-147.
36. Mahadevan, I. 1961. Belief systems in food of the Telugu-speaking people of the Telengana region. Indian J. Soc. Work,, 21: 387-396.
37. Ghana VAST study team. 1993. Vitamin A supplementation in northern Ghana: effects on clinic attendance, hospital admissions, and child mortality. Lancet, 342: 7-12.
38. Barreto, M.L. 1994. Effect of vitamin A supplementation on diarrhoea and acute lower-respiratory-tract infections in young children in Brazil. Lancet, 344: 228-231.
39. Bhandari, N., Bhan, M.K. & Sazawal, S. 1994. Impact of massive dose of vitamin A given to preschool children with acute-diarrhoea on subsequent respiratory and diarrhoeal morbidity. BMJ, 309: 1404-7.
40. Fawzi, W.W. 1993. Vitamin A supplementation and child mortality. A meta-analysis. JAMA, 269: 898-903.
41. Glasziou, P.P. & Mackerras, D.E.M. 1993. Vitamin A supplementation in infectious diseases: a meta-analysis. BMJ, 306: 366-70.
42. Menon, K. & Vijayaraghavan, K. 1979. Sequelae of severe xerophthalmia: a follow-up study. Am. J. Clin. Nutr., 33: 218-20.
43. Hussey, G.D. & Klein, M. 1990. A randomised controlled trial of vitamin A in children with severe measles. N. Engl. J. Med., 323: 160-164.
44. West, K.P. 1997. Impact of weekly supplementation of women with vitamin A or beta-carotene on foetal, infant and maternal mortality in Nepal. In: Report of the XVIII International Vitamin A Consultative Group Meeting. p. 86. 22-26 September. Cairo, Egypt.
45. WHO. 1995. The Vitamin A and Pneumonia Working Group. Potential interventions for the prevention of childhood pneumonia in developing countries: a meta-analysis of data from field trials to assess the impact of vitamin A supplementation on pneumonia morbidity and mortality. Bulletin of the World Health Organization. 73: 609-619.
46. Coutsoudis, A., Broughton, M. & Coovadia, H.M. 1991. Vitamin A supplementation reduces measles morbidity in young African children: a randomised, placebo-controlled, double blind trial. Am. J. Clin. Nutr., 54: 890-895.
47. Solomons, N.W. & Keusch, G.T. 1981. Nutritional implications of parasitic infections. Nutr. Revs., 39: 149-161.
48. Feachem, R.G. 1987. Vitamin A deficiency and diarrhoea: a review of interrelationships and their implications for the control of xerophthalmia and diarrhoea. Tropical Disease Bulletin, 84: R1-R16.
49. Thurnham, D.I. & Singkamani, R. 1991. The acute phase response and vitamin A status in malaria. Transactions of the Royal Society of Tropical Medicine and Hygiene, 85: 194-199.
50. Campos, FACS., Flores, H. & Underwood, B.A. 1987. Effect of an infection on vitamin A status of children as measured by the relative dose response (RDR). Am. J. Clin. Nutr., 46: 91-94.
51. Curtale, F., Pokhrel, R.P., Tilden, R.L. & Higashi, G. 1995. Intestinal helminths and xerophthalmia in Nepal. J. Tro. Pediatr., 41: 334-337.
52. Sommer, A. & West, K.P. Jr. 1996. Infectious morbidity. In: Vitamin A Deficiency, Health, Survival, and Vision. p.19-98. New York, Oxford University Press.
53. Foster, A. & Yorston, D. 1992. Corneal ulceration in Tanzanian children: relationship between measles and vitamin A deficiency. Transactions of the Royal Society of Tropical Medicine and Hygiene, 86: 54-455.
54. Arroyave, G. 1961. Serum and liver vitamin A and lipids in children with severe protein malnutrition. Am. J. Clin. Nutr., 1961, 9:180-185.
55. Bates, C.J. 1983. Vitamin A in pregnancy and lactation. Proc. Nutr. Soc., 42: 65-79.
56. Takahashi, Y. 1975. Vitamin A deficiency and foetal growth and development in the rat. J. Nutr., 105: 1299-1310.
57. Public Affairs Committee of the Teratology Society. 1987. Teratology society position paper: Recommendations for vitamin A use during pregnancy. Teratology, 35: 269-275.
58. Underwood, B.A. 1994. The role of vitamin A in child growth, development and survival. In: Allen L et al., eds. Regulation during pregnancy, lactation, and infant growth. p. 195-202. New York, Plenum,
59. IVACG. 1998. IVACG statement on vitamin A and iron interactions. Washington, DC, International Vitamin A Consultative Group.
60. Suharno, D. 1993. Supplementation with vitamin A and iron for nutritional anaemia in pregnant women in West Java, Indonesia. Lancet, 342: 1325-1328.
61. Sijtsma, K.W. 1993. Iron status in rats fed on diets containing marginal amounts of vitamin A. Br. J. Nutr., 70: 777-785.
62. FAO/WHO. 1967. Requirements of vitamin A, thiamine, riboflavine and niacin. Report of a Joint FAO/WHO Expert Group. WHO technical report series. 362, World Health Organization, Geneva.
63. Adams, W.R. & Green, M.H. 1994. Prediction of liver vitamin A in rats by an oral isotope dilution technique. J. Nutr., 124: 1265-1270.
64. Furr, H.C. 1989. Vitamin A concentrations in liver determined by isotope dilution assay with tetradeuterated vitamin A and by biopsy in generally healthy adult Humans. Am. J. Clin. Nutr., 49: 713-716.
65. Haskell, M.J. 1998. Plasma kinetics of an oral dose of [2H4]retinyl acetate in Human subjects with estimated low or high total body stores of vitamin A. Am. J. Clin. Nutr., 68: 90-95.
66. Van den Berg, H. & van Vliet, T. 1998. Effect of simultaneous, single oral doses of bb-carotene with lutein or lycopene on the bb-carotene and retinyl ester responses in the triacylglycerol-rich lipoprotein fraction of men. Am. J. Clin. Nutr., 68: 82-89.
67. Castenmiller, J.J. & West, C.E. 1998. Bio-availability and bioconversion of carotenoids. Ann. Rev. Nutr., 18: 19-38.
68. Parker, R.S. et al. 1999. Bio-availability of carotenoids in Human subjects. Proc. Nutr. Soc., 58 :1-8.
69. FAO/WHO. 1988. Requirements of vitamin A, iron, folate and vitamin B12. Report of a Joint FAO/WHO Expert Consultation. Rome, Food and Agriculture Organization.
70. Rodriguez-Amaya, DB. 1997. Carotenoids and food preparation: the retention of pro-vitamin A carotenoids in prepared, processed, and stored foods. Arlington, VA, John Snow, Inc./OMNI Project.
71. Booth, S.L., Johns, T. & Kuhnlein, H.V. 1992. Natural food sources of vitamin A and pro-vitamin A. UNU Food and Nutrition Bulletin, 14: 6-19.
72. Burití Palm. 1975. In: Report, Ad Hoc Panel of the Advisory Committee on Technology Innovations, Board on Science and Technology for International Development, Commission on International Relations. Underexploited tropical plants with promising economic value. P. 133-137. Washington, DC, National Academy Sciences.
73. Vuong, L.T. 1997. An indigenous fruit of North Vietnam with an exceptionally high ß-carotene content. p. 2. Sight and Life Newsletter.
74. IVACG. 1989. Report of the International Vitamin A Consultative Group. Guidelines for the development of a simplified dietary assessment to identify groups at risk for inadequate intake of vitamin A. Washington, DC, International Life Sciences Institute-Nutrition Foundation.
75. Périssé, J. & Polacchi, W. 1980. Geographical distribution and recent changes in world supply of vitamin A. Food and Nutrition, 6: 21-27.
76. ACC/SCN. Second report on the world nutrition situation. Vol.1. Global and regional results, October 1992. Vol. 2, March 1993.
77. US Department of Agriculture, Agricultural Research Service. 1998. Food and Nutrient Intakes by Individuals in the United States, by Sex and Age, 1994-96, pp. 197. Nationwide Food Surveys Report No. 96-2.
78. Centers for Disease Control and Prevention. 1998. National Health and Nutrition Examination Survey III, 1988-1994. CD-ROM Series 11, No. 2A, April 1998. Hyatsville, MD.
79. Gregory, J., Foster, K., Tyler, H. & Wiseman, M. 1990. The Dietary and Nutritionl Survey of British Adults. London, HMSO.
80. Tyler, H.A., Day, M.J.L. & Rose, H.J. 1991. Vitamin A and pregnancy [letter]. Lancet, 337:48-49.
81. Bloem, M.W., de Pee, S. & Darnton-Hill, I. 1997. Vitamin A deficiency in India, Bangladesh and Nepal. In: Gillespie S, ed. Malnutrition in South Asia. A regional profile. p.125-144. UNICEF Regional Office for South Asia.
82. de Pee, S. 1995. Lack of improvement in vitamin A status with increased consumption of dark-green leafy vegetables. Lancet, 346: 75-81.
83. Yin, S. 1998. Green and yellow vegetables rich in pro-vitamin A carotenoids can sustain vitamin A status in children. FASEB J., 12: A351.
84. Jalal, F. 1998. Serum retinol concentrations in children are affected by food sources of bb-carotene, fat intake, and anthelmintic drug treatment. Am. J. Clin. Nutr., 68: 623-9.
85. Christian, P. 1998. Working after the sun goes down. Exploring how night blindness impairs women’s work activities in rural Nepal. Eur. J. Clin. Nutr., 52: 519-524.
86. Underwood, B.A. & Olson, J.A., eds. 1993. A brief guide to current methods of assessing vitamin A status. A report of the International Vitamin A Consultative Group (IVACG). Washington, DC, Nutrition Foundation.
87. Sommer, A. History of nightblindness: a simple tool for xerophthalmia screening. Am. J. Clin. Nutr.,1980, 33:887-891.
88. Underwood, B.A. 1990. Biochemical and histological methodologies for assessing vitamin A status in Human populations. In: Packer L, ed. Methods in Enzymology: Retinoids, Part B. pp. 242-250.New York, Academic Press.
89. Olson, J.A. 1992. Measurement of vitamin A status. Voeding, 53: 163-167.
90. Sommer, A. & Muhilal. 1982. Nutritional factors in corneal xerophthalmia and keratomalacia. Arch. Ophthalmol., 100: 399-403.
91. Wachtmeister, L. 1988. Attempts to define the minimal serum level of vitamin A required for normal visual function in a patient with severe fat malabsorption. Acta Ophthalmol., 66: 341-348.
92. Flores, H. 1984. Assessment of marginal vitamin A deficiency in Brazilian children using the relative dose response procedure. Am. J. Clin. Nutr., 40: 1281-1289.
93. Flores, H. 1991. Serum vitamin A distribution curve for children aged 2-6 y known to have adequate vitamin A status: a reference population. Am. J. Clin. Nutr., 54: 707-711.
94. Pilch, S.M., ed. 1987. Analysis of vitamin A data from the health and nutrition examination surveys. J. Nutr., 117: 636-640.
95. Christian, P., Schulze, K., Stoltzfus, R.J., West, K.P. Jr. 1998. Hyporetinolemia, illness symptoms, and acute phase protein response in pregnant women with and without night blindness. Am. J. Clin. Nutr., 67: 1237-1243.
96. Filteau, S.M. 1993. Influence of morbidity on serum retinol of children in a community-based study in northern Ghana. Am. J. Clin. Nutr., 58: 192-197.
97. WHO/UNICEF/ORSTOM/UC Davis. 1998. Complementary feeding of young children in developing countries: a review of current scientific knowledge. Pp.228. WHO/NUT/98.1. Geneva, World Health Organization.
98. Rahmathullah, L. Reduced mortality among children in Southern India receiving a small weekly dose of vitamin A. N. Engl. J. Med., 323: 929-935.
99. Miller, R.K. 1998. Periconceptional vitamin A use: How much is teratogenic? Reproductive Toxicology, 12: 75-88.
100. Hathcock, J.N. Evaluation of vitamin A toxicity. Am. J. Clin. Nutr., 52: 183-202.
101. Hathcock, J.N. 1997. Vitamins and minerals: efficacy and safety. Am. J. Clin. Nutr., 66: 427-437.
102. WHO. 1998. Safe vitamin A dosage during pregnancy and lactation. pp.34. WHO/NUT/98.4. Geneva, World Health Organization.
103. Karr, M. 1997. Age-specific reference intervals for plasma vitamin A, E and beta-carotene and for serum zinc, retinol-binding protein and prealbumin for Sydney children aged 9-62 months. Int. J. Vit. Nutr. Res., 67: 432-436.
104. Smith, F.R. & Goodman, D.S. 1976.Vitamin A transport in Human vitamin A toxicity. N. Engl. J. Med., 294: 805-808.
105. Humphrey, J.H. 1998. Neonatal vitamin A supplementation: effect on development and growth at 3 y of age. Am. J. Clin. Nutr., 68: 109-117.
106. Baqui, A.H. 1995. Bulging fontanelle after supplementation with 25,000 IU vitamin A in infancy using immunisation contacts. Acta Paediatrica, 84: 863-866.
107. de Francisco, A. 1993. Acute toxicity of vitamin A given with vaccines in infancy. Lancet, 342: 526-527.
108. WHO/CHD Immunisation-Linked Vitamin A Supplementation Study Group. 1998. Randomised trial to assess benefits and safety of vitamin A supplementation linked to immunisation in early infancy. Lancet, 352: 1257-1263.
109. Van Dillen, J., de Francisco, A., & Ovenrweg-Plandsoen, W.C.G. 1996. Long-term effect of vitamin A with vaccines. Lancet, 347:1705.
![]()
![]()
![]()
{kind=link}