![]()
![]()
![]()
El marcado es un procedimiento utilizado desde hace mucho tiempo para el estudio de las migraciones, el crecimiento y la mortalidad de animales marinos. Rounsefell en 1963 y, mas recientemente, Jakobsson (1970) y Neal (1970) han hecho una revisión general de las técnicas de marcado utilizadas en peces e invertebrados. En el caso particular de los camarones nos encontramos con las dificultades principales siguientes:
su pequeño tamaño y consiguiente fragilidad;
el proceso de muda. Toda perturbación durante este proceso puede producir irregularidades en el crecimiento o aumento de mortalidad. Existen dos clases de métodos de marcado, aquellos que permiten distinguir individualmente los ejemplares, y los que sólo permiten el marcado de un grupo de individuos que luego son seguidos como una cohorte artificial.
Una operación de marcado debe ser cuidadosamente planificada desde el principio para asegurar las mayores posibilidades de supervivencia de los ejemplares marcados. Por ejemplo, para un marcado en los estuarios, conviene seleccionar lugares donde los parámetros hidrológicos (temperatura, salinidad, oxígeno disuelto) son favorables para la especie bajo estudio, evitando, por ejemplo, salinidades muy bajas o temperaturas muy altas.
Las instalaciones de marcado (circulación de agua, volumen de tanques disponible, aireación) deben ser diseñadas para asegurar las mejores condiciones.
Esta operación se debe realizar cuidando al máximo no dañar a los camarones. Es preferible evitar, cuando sea posible, el uso de artes de arrastre, particularmente en estuarios donde son mejores las artes de estacada o las nasas. Si el arrastre es inevitable, deberá llevarse a cabo con lances cortos (10–15 minutos, por ejemplo). La duración del arrastre se puede reducir en función de la abundancia de la especie objetivo, las especies acompañantes y la cantidad de cuerpos “extraños” en el fondo (piedras, esponjas) que aplastarán a los camarones durante la subida del arte. Costello (1964) sugirió que la presión en el copo podría ser reducida ajustando anillos rígidos, cosidos en el interior, para mantener la bolsa abierta.
Los problemas de mantenimiento de camarones vivos son más agudos si el marcado se lleva a cabo en tierra. En el mar es suficiente una buena circulación de agua. Pueden surgir problemas si la salinidad del agua bombada por el barco desde la superficie es muy diferente a la del fondo donde viven los camarones. La temperatura también debe ser observada, siendo a veces preferible suspender el marcado durante las horas más calurosas del día.
En barcos pequeños, y en los estuarios, una solución sencilla consiste en atar jaulas flotantes al barco anclado, asegurando de este modo la circulación con las corrientes naturales.
Si el marcado se lleva a cabo en la orilla, la instalación será más compleja y deberá incluir una bomba y sistema de aireación. Este último equipo puede ser eliminado inyectando agua con dispositivos especiales con un efecto venturi, que asegurarán la aireación o, aún más sencillo, dejando que el agua agite la superficie con suficiente presión.
La mesa de marcado debe tener dos cubetas de poca profundidad con agua de mar, donde se coloca el camarón durante la operación de marcado (Figura 40) y, si es posible, un canal que transporte el camarón al tanque de recuperación. Puede ser necesario bajar la temperatura de los tanques de almacenamiento y Emiliani (1971) describió el sistema utilizado en el laboratorio de Galveston (USA).
El uso de anestésicos puede recomendarse para reducir los sobresaltos del camarón durante el marcado. Zein Eldin (citado por Neal, 1970) señaló que se requerían altas concentraciones de anestésicos y subrayó lo difícil que era no sobrepasar el estrecho límite de dosificación no letal con el que se consigue una sedación rápida.
A menudo es preferible marcar los camarones inmediatamente después de la captura, cuando todavía están conmocionados y relativamente inmóviles. La experiencia muestra que su recuperación es muy rápida y que se vuelven extremadamente activos después de 20 ó 30 minutos. Más tarde el marcado se vuelve más difícil para el operador y más violento para el animal.
El objetivo de la operación es marcar un grupo de ejemplares de unas características dadas para seguirlos a lo largo del tiempo. Es preferible marcar grupos homogéneos, especialmente cuando los fenómenos a observar pueden variar con la edad.
6.3.1 Separación en Grupos
Los jóvenes camarones en emigración están en general separados naturalmente, en el sentido de que el rango de tamaños y de edades en la captura es pequeño. Este no es el caso en el mar, por lo que la distribución se hace necesaria. Allen y Costello (1962) desarrollaron un método simple usando separadores transparentes. El paso de los camarones a marcar a través de dos separadores malla diferente provoca la selección de un grupo de tallas homogéneo. Los camarones de talla superior quedan retenidos en el primer separador, y los de talla pequeña escapan a través del fondo del segundo.
6.3.2 La Utilización de Colorantes Vitales
Menzell y Racek han utilizado colorantes vitales desde 1955. Arnold (1965) revisó su uso en peces. Diversos métodos pueden usarse:
6.3.2.1 La inmersión total
En una solución colorante que impregna las membranas y el tubo digestivo. Según Dawson (1957), colorante sólo persiste por unos días. Este método fue discutido por Racek (1956). La persistencia del azul Trypan es de dos semanas, pero con un 50% de mortalidad inmediata. El sulfato azul Nilo da, según este autor, mejores resultados. La marca desaparece con la muda (Meyer-Waarden y Tiews, 1965).
6.3.2.2 La ingestión
De pastillas alimenticias coloreadas también coloreará el tubo digestivo. Dawson, desde 1957, ha experimentado con ambas técnicas, aunque no son de uso común debido a la rápida desaparición del colorante. A veces la persistencia es buena, más de 200 días, aunque la sobreingestión del colorante produce un envenenamiento fatal. No obstante, según el autor, este inconveniente se puede evitar.
6.3.2.3 La inyección intramuscular
Esta ha sido utilizada en camarones desde 1955 por Menzel (1955), y por Dawson (1957), Costello (1959), Costello y Allen (1965), Klima (1965, 1974), Berry (1967), Kutkuhn (1962) y McCoy (1968) y parece dar los mejores resultados.
Costello (1964), ha descrito un procedimiento detallado de esta técnica, desde la captura hasta la puesta en libertad, y aquí recopilaremos los detalles esenciales de este trabajo. El material estándar consiste en jeringuillas de tuberculina de 0,5 ó 1 cc y con agujas hipodérmicas muy finas. Puede usarse una jeringa automática (Figura 41). La solución colorante se inyecta lateralmente a través de la membrana articular, entre el 5o y 6o segmento abdominal (o entre el 3o y el 4o, Neal 1970). Para los camarones de tallas inferiores a 60 mm (longitud total), se puede inyectar a través del primer segmento abdominal. Es necesario evitar el perforar el tubo digestivo e inyectar burbujas de aire. La velocidad de este marcado es de 200/240 camarones por hora y puede llegar a ser 300/hora.
Los camarones deben ser contados y medidos cuando están siendo marcados. Luego, los que estén en malas condiciones se substraen del total. El colorante inyectado en el sistema vascular a través de la arteria dorsal-abdominal se fija en las branquias (Figura 42a) donde persiste por varias semanas1. El inconveniente de esta técnica es la rápida desaparición del color que puede producir una reducción artificial en la tasa de recaptura durante el período de recaptura.
6.3.3 Los Pigmentos Fluorescentes
6.3.3.1 La pulverización en masa
Los camarones pueden ser pulverizados a alta presión (14 kg/cm) con un polvo fluorescente (Benton y Lightner, 1972). Las partículas de este polvo penetran en las membranas flexibles del caparazón y su fluorescencia puede ser observada bajo la luz ultravioleta. Este método presenta diversos inconvenientes.
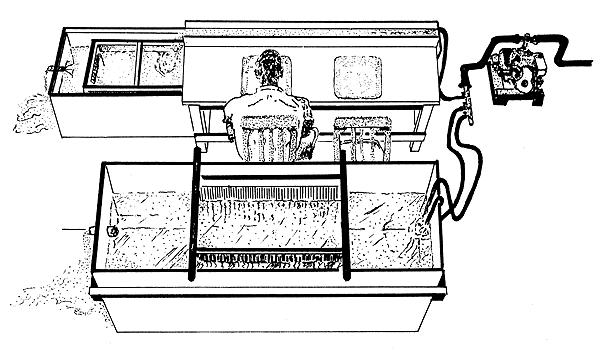
Figura 40 Instalación de marcado en tierra (según Costello, 1964)
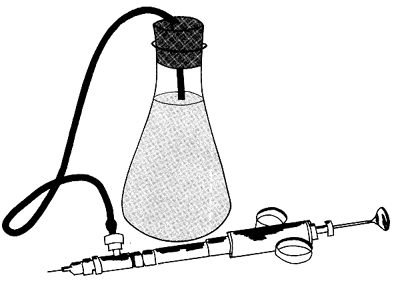
Figura 41 Sistema de inyección automática (según Neal, 1969)
El más importante es que la marca tiene poca persistencia y desaparece con la primera muda, salvo circunstancias excepcionales en que ha sido retenida durante 130 días. Loesch et al (1976) utilizando este método demostraron que las partículas pueden pasar a través del caparazón y persistir después de varias mudas.
La necesidad de usar luz ultravioleta para su detección, lo que limita las experiencias de marcado y recaptura a ser llevadas a cabo por operadores científicos.
La pulverización en masa en el mar puede posiblemente contravenir las normas en materia de colorantes comestibles ya que los camarones no detectados pueden ser ingeridos accidentalmente.
El marcado fluorescente de este tipo tiene la ventaja de que, para estudios a corto plazo, es posible que una sola persona efectúe el marcado de varios cientos de individuos por hora (Neal, 1970).
6.3.3.2 La inyección
Puede efectuarse utilizando un producto fluorescente disuelto en aceite de parafina (Klima, 1965). El colorante tiene una fuerte persistencia (276 días) pero requiere, para su detección, la supervisión personal de científicos y el uso de luz ultravioleta. Smith (1970) estudió el uso de estos colorantes para el marcado de peces jóvenes. En esta experiencia, la marca estuvo visible de 8–11 meses. Clark y Caillouet (1973) utilizaron esta técnica con juveniles de Penaeus setiferus y Edwards (1977) con Penaeus vannamei.
Neal (1970) y Penn (1981) destacaron que las partículas del aceite de parafina avanzan lentamente hacia las branquias donde se pueden acumular especialmente si se inyecta demasiado producto.
6.3.4 Las Tintas de Impresión
Probadas por Klima (1965), para un marcado secundario, no fueron satisfactorias porque no permanecían en el punto de inyección, sino que se difundían hacia las branquias, donde ocultaban la marca primaria.
6.3.5 Las Mutilaciones
El recorte parcial de un urópodo (“clipping”) es un método útil para experimentos a corto plazo. Ha sido utilizado por Barr (1971) en carídeos.
Tiene la ventaja de permitir la identificación de cada ejemplar por un número. Este marcado puede aplicarse a todos los ejemplares que puedan soportar una marca y su extensión en el tiempo y espacio no causa dificultades ya que las características del ejemplar marcado pueden ser fácilmente idéntificadas gracias al número específico.
6.4.1 Marcas Externas
Son visibles externamente y pueden fijarse al camarón por diversos procedimientos.
6.4.1.1 La marca de Atkins (Figura 42b)
Consiste en un hilo de nylon que atraviesa al camarón de lado a lado, atado con un bucle que lleva un disco de plástico coloreado y numerado (Allen y Costello, 1962; Penn, 1975; Ruello, 1970, 1975, 1977). Una marca similar, que simplemente se ata alrededor del caparazón, fue utilizada por Tiews (1964) para Crangon crangon. La marca de Atkins fue colocada por Costello y Ruello a través del primer segmento abdominal, y por Penn en la unión del primer y segundo segmento para no obstaculizar la muda.
6.4.1.2 La cinta de vinilo
Marullo et al. (1976) usaron una cinta de plástico flexible, cosida a través del animal. Esta cinta tiene la ventaja de no dañar los tejidos y de no enredar al camarón. No obstante, esta se puede decolorar y volverse menos visible. La operación de marcado es más rápida que con la marca anterior, pero la toxicidad del plástico de color no ha sido aún comprobada. Tratadas con aureomicina para limitar las infecciones, estas marcas han sido utilizadas recientemente en los Estados Unidos con éxito (Klima, 1980 y Klima y Parrack, 1980) y en Kuwait (Mohamed et al., 1979). Hasta la fecha, es la marca externa más efectiva para el marcado de juveniles ya que pueden marcarse camarones de 50 a 100 mm y obtener unas excelentes tasas de recaptura.
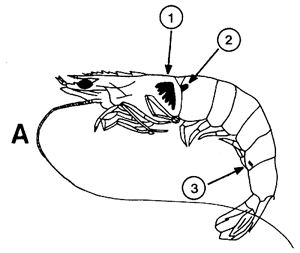
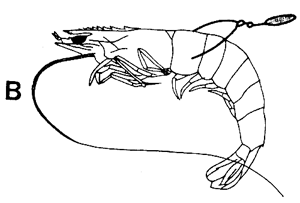
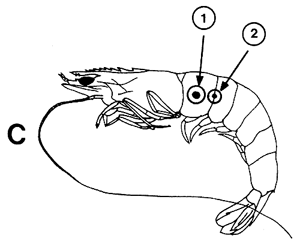
Figura 42 Diferentes tipos de marcas internas y externas
A. Inyección coloreada (1) marca interna de vinilo (2) e inyección fluorescente (3)
B. Marca de Atkins
C. Discos de Petersen: método original (1) y modificación propuesta por Neal, 1970 (2)
6.4.1.3 Los discos de Petersen y sus variantes (Figura 42C)
Esta vieja técnica ha sido ampliamente utilizada durante muchos años y ha evolucionado considerablemente. La marca tenía originalmente la forma de dos discos de 7,9 o 9,5 mm de diámetro, colocados a ambos lados del animal y unidos por un hilo de acero a través del cuerpo del camarón en la mitad del primer segmento abdominal (Iversen e Idyll, 1960; Potter, 1975). Este método fue mejorado por Neal (1970) que redujo el tamaño del disco y utilizó una aguja de acero inoxidable para evitar la corrosión. El enganche de la marca al camarón se simplificó utilizando una pinza de circuitos impresos. Las agujas se sumergieron en una solución antibiótica (2% clorotetraciclina) antes de su utilización. Este tipo de marcas, a las que Kurata (1962) se opone por su inevitable interferencia con la muda, han sido ampliamente utilizadas en los Estados Unidos (Lindner y Anderson, 1956; Iversen y Jones, 1961; Neal, 1969), en Côte d'Ivoire (García, 1973, 1975), en Senegal (Lhomme, 1977), en Madagascar (Le Reste y Marcille, 1976) y en Australia (Lucas, Young y Brundritt, 1972) dando, aparentemente, buenos resultados. Welker et al. (1975) compararon este tipo de marcas con las inyecciones colorantes y las marcas internas; no encontrando diferencias significativas en el crecimiento o en la tasa de recaptura.
El último paso en la evolución de marcas de este tipo es la “toggle tag” (marca articulada) utilizada por Penn (1975) en Australia. La marca numerada se fija al cuerpo entre el primer y segundo segmento abdominal mediante un hilo de PVC (35 mm de longitud y 0,5 mm de diámetro), terminado en dos barras en forma de T, que sobresalen por un lado y otro del animal. La marca es aplicada con una pistola especial (Floy tag). La flexibilidad del hilo de PVC reduce la ulceración de tejidos. La “toggle tag” (marca articulada) y la marca de Atkins dan resultados similares.
6.4.1.4 Las marcas de dardo
Fijadas al músculo de la gamba, estas marcas (“dart tag” y “anchor tag”) han demostrado ser desastrosas para los camarones. Dell (1968) ha descrito la marca y la pistola que se usa para su colocación. Estas han sido utilizadas, por ejemplo, por Bearden y McKenzie (1972) y tienen la desventaja de ser asimétricas (la marca aparece sólo en un lado) y desequilibran al camarón durante la natación. El movimiento excesivo de la marca causa una ulceración permanente que es, asimismo, una constante fuente de mortalidad adicional (Neal, 1969; Lucas, Young y Brundritt, 1972).
6.4.2 Las Marcas Internas
Una marca de este tipo ha sido descrita por Neal (1970), y consiste en una pequeña etiqueta de plástico numerada, de 5 × 2 × 0,25 mm, que se introduce en el músculo abdominal a través de una membrana articular, por lo que es visible y normalmente no se pierde con la muda. Para su detección debe estar acompañada de una marca primaria (inyección colorante). La aplicación de una pomada antibiótica reduce las infecciones (Figura 42B). Penn (1981) indica que esta marca puede ser rechazada en la muda después de infecciones locales y erosiones de los tejidos circundantes.
Las marcas metálicas de acero inoxidable, codificadas con bandas de colores, han sido utilizadas por Jefferts, Bergman y Fiscus (1963) y West y Chew (1968) en Pandalus platyceros. Estas no parecen provocar mortalidad adicional y el crecimiento es normal. Existe un aparato que permite su rápida colocación. Después de la inserción, la marca se magnetiza para facilitar su detección y recuperación. Esta técnica fue utilizada satisfactoriamente para peneidos por Welker (citado por Neal, 1970). No obstante, su detección durante operaciones pesqueras industriales tiene algunos problemas técnicos.
Consiste en colocar simultáneamente dos marcas sobre el animal. En teoría se utiliza para detectar y estimar las pérdidas accidentales de marcas. En camarones, han sido empleadas para ampliar las posibilidades ofrecidas por las inyecciones de colores. Estas, que son claramente visibles desde el exterior, constituyen la marca primaria para distinguir, por ejemplo, diferentes períodos de marcado. La marca secundaria permite identificar, después de la recaptura, y para un examen más detallado, los diferentes subgrupos (diferentes grupos de tamaño, diferentes lugares de puesta en libertad, etc.). La marca secundaria podría ser, en este caso, una inyección de gelatina fluorescente en la cola (Klima, 1965) (Figura 42A). La aplicación e inyecciones en dos lugares diferentes aumentan las combinaciones posibles. Este autor no observó mortalidad adicional alguna. Este método también ha sido aplicado por McCoy y Brown (1967) sobre juveniles. El marcado doble se aplica en una o dos operaciones. En el primer caso, los dos colorantes (vital y fluorescente) son inyectados en una mezcla. El primero 'alcanza las branquias en 24 horas y el segundo después de 48 horas. La mezcla de los dos colorantes puede detectarse hasta 45 días más tarde por medio de luz ultravioleta.
Si está numerada, la marca secundaria puede utilizarse para la identificación del individuo (ver Sección 5.4.2). El doble marcado de este tipo ofrece la ventaja del marcado individual (identificación visual de ejemplares) sin interferir con la muda; por otra parte, tiene el inconveniente de que las inyecciones colorantes se borran con el tiempo. No obstante, este método ha sido utilizado por Berry (1967).
Esta es una de las fases delicadas de la operación de marcado. Es, a lo largo de esta operación, cuando la predación puede causar una mortalidad adicional importante. El principio esencial que debe ser mantenido es: los camarones no deben ser jamás devueltos al mar individualmente. Deberán soltarse, siempre que sea posible, cerca del fondo. Además, sólo se liberaran los que estén en buen estado. Esta comprobación se hace por inspección en una cubeta con agua y mucha aireación. Los camarones, que al principio están conmocionados, se recuperan inmediatamente y la experiencia demuestra que, en general, es suficiente una espera de una hora, ya que los camarones que sobreviven a ese período, luego sobreviven varios días en viveros (García, 1977, 1977b; Lhomme, 1978; Le Reste y Marcille, 1976). Berry (1967) ha conservado camarones marcados durante 24 horas. En el marcado de juveniles, Klima (1965) prolongó este período a 18 días.
Un cierto tiempo de recuperación es deseable pero no debe ser muy largo ya que, contrario al efecto deseado, los camarones marcados pueden finalmente debilitarse (Beverton y Bedford, 1963).
La liberación, propiamente dicha, se efectúa con la ayuda de una caja especial que se abre por el fondo (Costello, 1964); Le Reste y Marcille, 1976) mediante un mensajero enviado desde la superficie o con la ayuda de un recipiente barato de plástico (Figura 43), que se abre simplemente tirando de una cuerda (García, 1973), o más sencillo todavía, colocando en el mar unos recipientes desmontables de plástico, lastrados y sueltos, que se abren en el fondo gracias a un simple pero ingenioso proceso de disolución de un bloque de sal (Emiliani, 1971) (Figura 44). Este sistema se utiliza actualmente y de forma rutinaria en el golfo de México. En lagunas de poca profundidad, se pueden enviar los camarones al fondo a través de un tubo que les permite, una vez introducidos en la superficie, alcanzar el fondo directamente (Emiliani, op. cit.).
En ciertos casos (en zonas limitadas o pequeñas bahías) se efectúa por el equipo de investigación mismo. Más a menudo, los camarones son capturados por los pescadores, por lo que debe tomarse un cuidado considerable en lo que se refiere a la planificación de la operación, antes y después del marcado, para obtener resultados satisfactorios.
6.7.1 La Información Preliminar
Los pescadores deben estar debidamente informados a través de contactos personales con los científicos, con los patrones, tripulaciones, armadores, clasificadores en las fábricas, etc., y por medio de la difusión de anuncios atractivos de color, indicando claramente la clase de marca utilizada, su posición, la dirección postal a la que se debe devolver y la recompensa ofrecida. La organización periódica de una lotería puede animar a la devolución de las marcas, si los premios son buenos.
6.7.2 La Recogida de Información de Recaptura
El camarón debe ser recobrado entero y con toda la información asociada a la recuperación (fecha y lugar de recaptura). No obstante, no es aconsejable disminuir la recompensa cuando el camarón sea devuelto con la información incompleta o bien se devuelva sólo la marca, ya que se corre el riesgo de recibir información falsa. Un contacto muy estrecho con los pescadores asegurará la recogida de camarones recapturados y la información en las mejores condiciones. También puede ser necesario distribuir recipientes con formol para asegurar la buena conservación de los camarones. Cuando los barcos salen por mucho tiempo, es aconsejable que los patrones anoten sobre la marcha en sus cuadernos de bitácora el número de las marcas recobradas con toda la información adicional, ya que el camarón crece rápidamente y se hace necesaria la mayor precisión informativa posible sobre la fecha de recaptura. La recolección de información suplementaria de la pesquería y, en particular, del esfuerzo pesquero debe ser cuidadosamente planificada. Es deseable un esfuerzo suplementario durante unas semanas después del marcado para obtener una detallada información sobre la distribución geográfica del esfuerzo pesquero.
6.7.3 Los Sesgos
Las fuentes de sesgos que afectan a los resultados del marcado han sido revisadas en las obras básicas de Beverton y Holt (1957), Ricker (1975) y, más recientemente, Gulland (1969). Debe recurrirse a estos manuales para un estudio a fondo de estos problemas. En esta sección, y en la siguiente, se resumirán los factores esenciales relacionados con camarones. Z, F y M representan los coeficientes instantáneos de mortalidad total por pesca y natural respectivamente. Para una mejor comprensión del texto que sigue, será de mucha utilidad referirse al Capítulo 7 que trata de las mortalidades.
6.7.3.1 Sesgos de Ricker tipo A
Estos afectan a los valores de F pero no a los de Z.
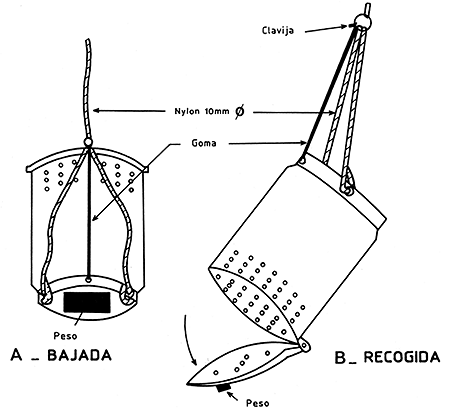
Figura 43 Sistema simple de suelta a nivel del fondo en poca profundidad. La caja desciende libremente, la tracción de la cuerda a nivel del fondo permite la apertura (según García, 1973)
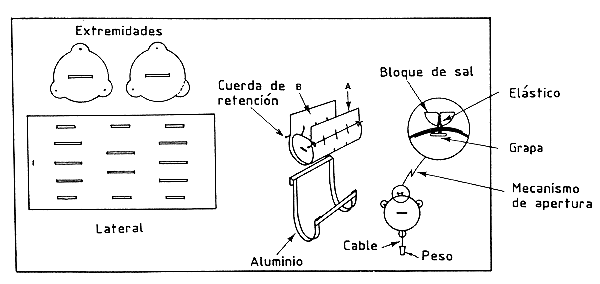
Figura 44 Caja de suelta autónoma con apertura automática. La caja desciende libremente, la dislución de un bloque de sal la abre al cabo de 10 minutos (según Emiliani, 1971)
(a) Mortalidad inicial debido al marcado
Esta ha sido estudiada por Penn (1975). Puede ser reducida guardando los camarones durante una o dos horas en un tanque con corriente de agua debidamente aireada, para eliminar los camarones más débiles. La devolución al mar debe hacerse con mucho cuidado y al nivel del fondo para evitar la predación producida por los peces que siguen al barco atraídos por las operaciones de pesca. García (1975) demostró que la tasa de recaptura depende de la hora de puesta en libertad y que la diferencia entre las tasas de recaptura de los camarones marcados durante el día o la noche es más importante cuando el agua está clara. La mortalidad inmediata puede también reducirse evitando marcar camarones que sean muy pequeños. Lucas, Young y Brundritt (1972) han demostrado que los discos de Petersen provocan una mortalidad adicional en camarones de menos de 30 mm (LC).
(b) Información incompleta de recapturas con tasa constante
Para reducir al mínimo este factor es importante asegurar una buena campaña de información antes del marcado y una buena organización para la recogida de marcas y el pago de las recompensas. Según Penn (1981), una importante pérdida de animales marcados ocurre particularmente con ejemplares jóvenes debido a que una proporción significativa de la captura de pequeñas tallas se procesa mediante máquinas de pelado automático.
La tasa de marcas recapturadas, pero no detectadas por las fábricas, puede calcularse utilizando la técnica de “siembra”. Esta consiste en introducir directamente, entre la captura de los barcos, un determinado número de camarones marcados lo que permite observar el porcentaje comunicado posteriormente. Paulik (1961), presentó un buen tratamiento estadístico de este método. Este ha sido utilizado por Berry (1967), y Costello y Allen (1968) para Penaeus duorarum. La cantidad de marcas no comunicadas depende obviamente del tipo de marca. A priori, esta es menor cuando la marca es más visible. En este sentido, los discos de Petersen no pueden prácticamente pasar desapercibidos durante el descabezado, mientras que las inyecciones colorantes son mucho más discretas. En este último caso, el empalidecimiento de la marca coloreada produce, además, un aumento progresivo en el porcentaje de marcas no detectadas, lo que afecta al valor calculado de Z. La curva de recaptura ya no es, en este caso, rectilínea.
Siguiendo la misma línea de razonamiento debe señalarse que el “interés” del pescador por el marcado, y por ello la atención prestada al descubrimiento de marcas, disminuye rápidamente cuando decrece la abundancia de camarones marcados. Por esta razón, puede ser importante considerar sólo las recapturas obtenidas en las primeras semanas después del marcado (Costello y Allen, 1978).
6.7.3.2 Sesgos de Ricker Tipo B
Cuando aparece este tipo de sesgos la disminución de las recapturas es más importante que la disminución real de la población. Estos afectan al valor de Z pero no al de F.
(a) Pérdida de marcas a tasa constante
Los discos de Petersen son poco propensos a causar estos sesgos, pero la decoloración progresiva de los colorantes inyectados puede contribuir a este fenómeno si a lo largo del tiempo empalidecen, a una velocidad constante, de tal forma que sean difícilmente detectados. La curva de recaptura puede incluso dejar de ser lineal cuando esta tasa no es constante. Ricker (1975, p. 114), propuso un método que permite superar esta dificultad. Se debe recordar que, según Penn (1981), las marcas internas de PVC también pueden ser rechazadas con la muda.
(b) Mortalidad debida al marcado constante
Este es un tipo de sesgo muy clásico. Normalmente se traduce en una sobreestimación de la mortalidad natural calculada por el marcado. Según Lucas, Young y Brundritt (1972), los discos de Petersen no producen este error con camarones de más de 30 mm LC.
(c) Tasa constante de emigración del área de pesca
Este error es potencialmente muy importante cuando el marcado tiene lugar en una laguna o estuario justo antes de la emigración. La tasa de migración no puede distinguirse de la mortalidad natural y en el proceso de estimación aparece como una mortalidad adicional.
Estos errores son difíciles de detectar y más aún de cuantificar y corregir. Por esta razón, el marcado generalmente permite obtener un valor para Z que puede descomponerse en F + X, donde X representa una mortalidad “combinada” que integra pérdidas de marcas, migración, mortalidad debida al marcado y mortalidad natural (M). Esta mortalidad a veces se toma como M a menos de que se indique lo contrario, y puede, de hecho, ser considerada como el límite superior de los posibles valores de M. (A menudo se usa el término “aparente” tasa de mortalidad natural.) Además, la combinación del valor de F determinado mediante el, marcado con el valor de Z hallado, por ejemplo mediante el análisis de distribuciones de edades, permite determinar un mejor valor de M.
6.7.3.3 Sesgos de Ricker tipo C
Estos afectan a Z y F. En este grupo se incluyen los sesgos que producen recapturas inútiles en el período inicial y aquellos que resultan de una diferencia en vulnerabilidad entre camarones marcados y no marcados causada por:
una alteración en el comportamiento de los camarones marcados, causada por la marca, que modifica la capturabilidad al principio del período de recaptura;
una distribución diferencial de camarones marcados y no marcados asociada con una distribución no aleatoria del esfuerzo pesquero en los primeros intervalos de tiempo. Durante los siguientes intervalos, el esfuerzo debe estar distribuido al azar o bien los animales marcados deberán tener la misma distribución que los no marcados para permitir el uso del número de recapturados como índice de abundancia de la población no marcada. Este aspecto se expone ampliamente en la Sección 7.2.3.
El primer tipo de error puede detectarse fácilmente cuando se dispone de buenos datos, ya que los primeros puntos en la curva de recaptura están claramente situados por encima o por debajo del alineamiento de los otros puntos que le siguen. El error es de poca importancia cuando se recuperan muchas marcas después del o de los intervalos sesgados, pero puede ser muy serio en el caso contrario. Gulland (1969) y Ricker (1975) han propuesto métodos para superar estos sesgos. El principio es la utilización de sólo puntos no sesgados (ver Sección 7.5).
El problema de la distribución diferencial de ejemplares marcados y no marcados es extremadamente importante y, para evitar esta dificultad, el marcado debe ser cuidadosamente planificado. Lo más sencillo es cuadricular toda el área de distribución (no sólo la zona o lugar de pesca) y marcar todos los ejemplares capturados por unidad de esfuerzo en cada estación o, si son muy numerosos, una proporción constante de ellos. Estos problemas se tratarán en el capítulo dedicado a la mortalidad.
No obstante, cuando el área es muy grande, puede que sea imposible de llevarse a cabo y Gulland (1969) propone un enfoque para poblaciones no-mezcladas que consiste en calcular F para los camarones marcados o deducir “q” a partir del conocimiento de la intensidad pesquera en la zona limitada donde se encuentran los camarones marcados. Si este valor puede tomarse también como válido para los ejemplares no marcados, la evaluación de la intensidad pesquera aplicada al conjunto de la población, permite el cálculo de un valor válido de F.
6.8.1 Sobre el Crecimiento
Es evidente que las marcas externas son susceptibles de impedir considerablemente el mecanismo de la muda aun cuando la técnica actual de inserción (en la membrana intersegmentaria) es preferible a la original (a través del primer segmento abdominal). Kurata (1962) consideró, debido a estas restricciones, que el marcado externo era totalmente incompatible con la fisiología de los camarones.
Según Penn (1975), las marcas externas causan una ralentización del crecimiento durante los primeros dos meses y frecuentemente no hay crecimiento hasta la primera muda, en condiciones experimentales. Por el contrario Neal (1970) apreció en acuarios que los camarones “inyectados” tienen un crecimiento superior al de aquellos que llevan una rígida marca externa.
A pesar de todas estas restricciones, y a espera de un método mejor, el marcado se ha utilizado frecuentemente. En muchas regiones (golfo de México, Kuwait, Australia, Africa occidental, Madagascar) se han obtenido excelentes resultados demostrando que el crecimiento de los camarones marcados, bajo condiciones naturales, es compatible con el obtenido por otros métodos. El marcado también ha proporcionado una herramienta valiosa para la demostración de variaciones estacionales del crecimiento asociadas con factores ambientales o con la reproducción (García, 1975a, 1977; Penn, 1975; Lhomme, 1978; Klima, 1980).
6.8.2 Sobre la Fisiología
Un estudio comparativo efectuado por Zein-Eldin y Klima (1965) no demostró diferencia entre el consumo de oxígeno de ejemplares marcados por inyección y los no marcados. Penn (1975) y García (1975) observaron, mediante el marcado, un retraso en el crecimiento durante el período de puesta.
El marcado externo causa una herida que se oscurece rápidamente. En la época de puesta este oscurecimiento desaparece, la herida se cicatriza (Penn, 1975), y una cutícula recién formada preserva la aguja de la marca externa, aislándola completamente del músculo (Ruello, 1970; Fontaine y Lightner, 1973; Penn, 1975). Penn notó que la cicatrización se retrasa con el uso de marcas rígidas del tipo “anchor tag”, una inyección que impide que la herida se cierre.
6.8.3 Sobre el Comportamiento
Según Costello y Allen (1961) las marcas externas modifican las posibilidades natatorias y de enterramiento de los camarones, haciéndolos más vulnerables a los predadores. Es exacto que, cuando la marca no está bien ajustada, se percibe un desequilibrio durante la natación. Penn (1975) confirmó que el enterramiento se dificulta por la marca rígida, pero Lucas, Young y Brundritt (1972) no han observado impedimentos para el enterramiento o la natación.
Según Penn, toda la actividad general se reduce durante los dos primeros meses que siguen al marcado (movimientos, mudas).
6.8.4. Sobre la Supervivencia
Muchos autores han observado que el porcentaje recapturado es menor en camarones pequeños (Figura 45) (Lindner y Anderson, 1956; Iversen y Jones, 1961; Lucas, Young y Brundritt, 1972; García, 1973, 1975; Le Reste y Marcille, 1974; Lhomme (1981). No obstante, la disminución observada es difícil de interpretar debido a que son difíciles de separar los medios de selectividad (capturabilidad reducida de pequeñas tallas) de aquellos producidos por el marcado (incremento de mortalidad).
Teniendo en cuenta la evidente conmoción que produce el marcado, es de esperar que el marcado sea responsable de alguna mortalidad. Conviene distinguir la mortalidad inicial, que ocurre en unas pocas horas después del marcado, de la mortalidad de marcado que se viene a sumar a las mortalidades pesquera y natural durante todo el período de observación.
6.8.4.1 La mortalidad inicial debida a la marca
Esa mortalidad inicial puede ser importante y se hace evidente por el número de camarones marcados que mueren durante el tiempo de retención antes de su puesta en libertad. Su magnitud depende en gran parte de las condiciones de captura y de la conservación a bordo. El efecto de esta mortalidad en el experimento puede reducirse guardando a bordo los animales marcados por algún tiempo de manera que los ejemplares más débiles puedan ser eliminados antes de la puesta en libertad. Según Penn (1975) los camarones que sobreviven en acuario, con una marca externa las primeras 72 horas, también sobrevivirán hasta la próxima muda. García (1977) notó que de un 93 a 99 % de los camarones marcados con discos de Petersen, y considerados sanos, sobreviven perfectamente más allá de la primera hora y no muestran, a continuación, una mortalidad superior a los ejemplares de control durante 12 a 24 horas.
No se puede asegurar que toda la mortalidad inicial ocurra antes de la puesta en libertad y es muy importante identificarla por este motivo después, ya que la reducción en el número inicial de ejemplares marcados causa un sesgo en la evaluación de F.
Lucas, Young y Brundritt (1972) han demostrado que los discos de Petersen, en contraste con el marcado de anclaje “anchor tag” (que da un 50 % de pérdidas), no producen una mortalidad inicial. Penn (1975) calculó para las “toggle tags” una mortalidad inicial de un 20 %. Lucas, Young y Brundritt, no han notado ningún efecto letal debido a las inyecciones colorantes, pero se debe recordar que los colorantes “vitales” pueden ser tóxicos en dosis elevadas (Costello, 1964) y se requiere una meticulosa utilización de esta técnica para limitar sus inconvenientes (análisis de las dosis, filtración en las soluciones, eliminación de burbujas de aire en la inyección, etc.).
La predación en el momento de la puesta en libertad puede ser también una fuente importante de mortalidad inicial. La mayoría de los autores coinciden en que es preferible la puesta en libertad de masa en el fondo. García (1975) observó, para diferentes grupos marcados, que la tasa de recaptura depende de la hora de la puesta en libertad (Figura 46) y que las diferencias son menores cuando las aguas están más turbias. El sugirió que este fenómeno podría estar ligado a la predación de los camarones por los peces atraídos por el barco en arrastre y los descartes, que seguirían la jaula en su descenso hacia el fondo.
6.8.4.2 Mortalidad adicional
Esta mortalidad esta implícitamente asumida en todas las operaciones de marcado por las que se obtiene una estimación de F y X, la mortalidad natural, “aparente” o mortalidad residual, incluyendo la mortalidad natural, la emigración, la pérdida de marcas y mortalidad adicional debido al marcado.
Lucas, Young y Brundritt (1972) han mostrado, en acuarios, que la mortalidad de marcado adicional ocurría en pequeños camarones marcados con discos y “anchor tags”, pero no en los adultos. Esto fue confirmado por Penn (1975) con marcas “articuladas” y marcas de Atkins. Este autor también destacó que el color de la marca no tenía influencia en la supervivencia, lo que confirma el trabajo de Costello y Allen (1961) que afirmaron que el color de la marca no tenía efecto en la predación. Según Penn (1975), se produce una importante mortalidad durante la primera muda después del marcado.
6.8.4.3 Conclusiones
A pesar de que la mortalidad debida a las operaciones de marcado y a la presencia de una marca es el problema principal de esta técnica de investigación, se hace muy difícil demostrar claramente tal mortalidad y, más aún, cuantificarla. Es obvio que tal mortalidad depende de las condiciones que prevalecen durante la operación de marcado (equipo, diferencias entre las condiciones a bordo y en el fondo, métodos de captura y marcado, procedimientos de puesta en libertad, etc.). También es cierto que los resultados de las observaciones experimentales, sobre supervivencia, muda, etc., deben ser aceptadas con precaución. De hecho, los resultados son a menudo contradictorios y, por ejemplo, muchos autores han apreciado una tasa de supervivencia superior en los camarones mayor que en los de control (Costello y Allen, 1961; Lucas, Young y Brundritt, 1972; Klima, 1965). Los dos primeros autores sugirieron incluso que los colorantes inyectados tienen un efecto profiláctico. Penn (1975) pensó que esta paradoja surgió porque la mortalidad en los acuarios ocurre principalmente durante la muda por conmoción o canibalismo y como las actividades generales de los camarones marcados se reducen, esto tiene una tasa de muda inferior a la de los de control y, por lo tanto, una mortalidad inferior. También se han observado mortalidades muy altas en lotes de control (Lucas, Young y Brundritt, 1972; Penn, 1975), lo que indica que, en muchos experimentos, los animales están bajo condiciones de tensión. Parece probable que los animales menos conmocionados sobreviven mejor, cuando las condiciones bajo las que se les mantiene se vuelven limitantes.
Por lo tanto, uno puede preguntarse en que medida pueden ser extrapolados los resultados obtenidos en el acuario, a experimentos reales de marcado. Penn (1975) ha perfilado sus limitaciones y Lucas, Young y Brundritt (1972) puntualizaron que los resultados experimentales en acuarios “dependen de las condiciones ambientales” y que su uso posterior debe realizarse con prudencia.
Por ello sería muy útil intentar durante las operaciones de marcado identificar los posibles sesgos por medio de métodos indirectos: comparación de resultados obtenidos con lotes diferentes, con los dos sexos, en diversas estaciones y con diversos tipos de marcas.
Una prueba útil sería comparar las distribuciones de frecuencia de tallas de los camarones marcados, recapturados, y explotados por la flota pesquera con el fin de asegurar que las poblaciones marcadas y explotadas sean idénticas (Figura 47), y que los resultados obtenidos para la población marcada puedan ser extrapolados al conjunto de la población explotada.
Para el análisis de los resultados, es probable que las recapturas obtenidas a “largo plazo” (después de los 30 primeros días que siguen al marcado) darán resultados menos sesgados, tanto en lo que concierne a las mortalidades como al crecimiento.
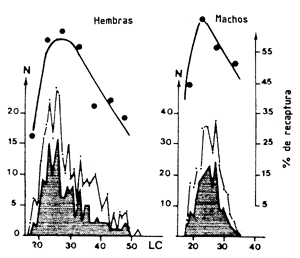 | 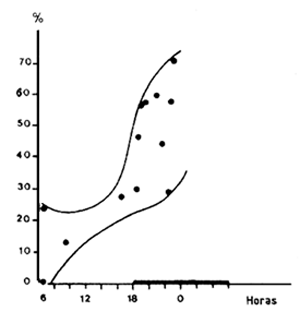 |
Figura 45 Tasa de recaptura en función de la talla en el primer mes que sigue al marcado (según García, 1975). En claro, camarones marcados; en oscuro, camarones recapturados | Figura 46 Variaciones de la tasa de recaptura según la hora de liberación (según García, 1975) |
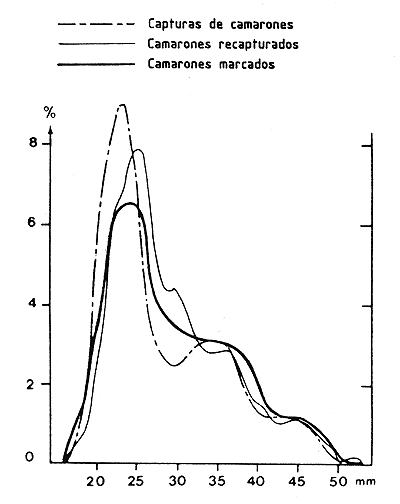
Figura 47 Comparación de las distribuciones de frecuencias de tallas de camarones marcados, recapturados y capturados por los camaroneros durante el mismo período (según García, 1977)
![]()
![]()
![]()