![]()
![]()
![]()
Una de las condiciones esenciales para la evaluación u ordenación de los recursos es la obtención de una buena estimación del esfuerzo pesquero. Para evitar los sesgos es necesario satisfacer la relación clásica F = qf. Ello supone que cualquier variación de q, tanto periódica como a largo plazo, implica, para un nivel dado de esfuerzo nominal, una variación de la mortalidad por pesca correspondiente. Además, la captura por unidad de esfuerzo C/f se usa frecuentemente como un índice de la abundancia en la medida que admitamos la relación, igualmente clásica, C/f = qD, donde D es la densidad (abundancia por unidad de superficie). El resultado es que toda variación de q se traducirá por una variación “aparente” de la densidad.
Por estas razones, ha parecido útil a los autores examinar diferentes tipos conocidos de q en los camarones y sus consecuencias, incluso a costa de algunas repeticiones. Para facilitar la exposición distinguiremos los siguientes tipos de variaciones: nictimerales, mareales, lunares, estacionales, espaciales, específicas de la edad e interanuales. Las diferencias entre algunas de ellas son quizás arbitrarias; está claro que para estos animales de vida muy corta que permanecen alrededor de un año en la pesquería, hay un solapamiento entre las variaciones estacionales, espaciales y específicas de la edad, desde el momento que están íntimamente ligadas al desarrollo de las cohortes en el tiempo y en el espacio.
Han sido mencionadas de forma resumida en el Capítulo 2. Estas variaciones han sido puestas en evidencia hace muchos años y por muchos autores, ya que su manifestación evidente es la aparición de variaciones nictimerales del rendimiento. Los camarones son, en general, animales de actividad nocturna que permanecen enterrados en el sedimento durante el día. Los rendimientos, reflejando el ritmo de actividad, son mejores por la noche, tanto en el mar como en los estuarios y lagunas. No obstante este no es siempre el caso en el mar y, cuando los rendimientos son más elevados durante el día, podemos pensar que por causas asociadas con la luz (y la transparencia del agua), los camarones descansan sobre el fondo durante el día y nadan libremente durante la noche, particularmente fuera del alcance de las artes de arrastre. El ritmo de actividad “aparente” se invierte con respecto al ritmo real.
Estas variaciones temporales se “borran” usando datos medios de captura por unidad de esfuerzo que permite calcular un índice de abundancia diaria. García, Petit y Fonteneau (1973), han estudiado este fenómeno con gran detalle en el golfo de Guinea, sobre diversos fondos de pesca de Nigeria y Senegal, y en varias estaciones. Ellos demostraron que, en esta región donde las aguas se caracterizan por ser portadoras de altas concentraciones de partículas sólidas, los rendimientos eran más frecuentemente altos durante el día pero que periódicamente sucedían cambios del ritmo de actividad “aparente”, asociados con cambios estacionales de las condiciones hidrológicas (temperatura en el límite septentrional de la distribución de la especie y turbidez). Más adelante veremos que estas modificaciones estacionales introducen variaciones estacionales de q.
Las variaciones ligadas al ritmo de las mareas son claramente evidentes para la pesca artesanal de juveniles con redes fijas donde las capturas se producen durante el reflujo. La capturabilidad puede variar con la velocidad de la corriente; Yokel (1969) demostró que si la velocidad era inferior a 0,31 m/segundo, los camarones podrían nadar fuera de la red y en contra la corriente. Las variaciones estacionales de las corrientes de mareas y de las inundaciones de los ríos pueden también ocasionar variaciones estacionales en la capturabilidad en este tipo de pesca.
Prácticamente todos los pescadores hablan de estas variaciones ligadas a las fases de la luna pero, en el mar, estas son difíciles de demostrar de forma incuestionable. No obstante, su existencia ha sido claramente demostrada por ciertos autores (Capítulo 2). Por el contrario, estas son muy evidentes en las lagunas o en los estuarios en la pesca de juveniles en emigración donde se observan máximos principales y secundarios en luna nueva y llena respectivamente.
Estas se conocen desde hace mucho tiempo para prácticamente todas las poblaciones explotadas. Sin embargo, su estudio ha sido frecuentemente descuidado porque, en la dinámica de poblaciones clásica, el uso de índices medios anuales de abundancia permite obviar esta dificultad.
1 El término capturabilidad se utiliza aquí en su sentido más amplio
Por el contrario, el estudio de especies de vida corta como los camarones requiere el uso de un intervalo de tiempo inferior al período de oscilación de los fenómenos biológicos (un año), lo que pone en relieve la importancia de estudiar el ritmo estacional cuyo efecto no puede ser por más tiempo enmascarado por el efecto medio en un intervalo básico anual.
Las fuentes de variaciones estacionales de la capturabilidad son numerosas. Un primer inventario ha sido preparado por García (1977b) que aquí repetiremos y completaremos. Las principales fuentes de variaciones estacionales son:
8.4.1 Las Condiciones Hidrológicas y el Comportamiento
8.4.1.1 La turbidez
Hemos visto en el primer párrafo de este Capítulo que las variaciones nictimerales del rendimiento pueden dar una imagen correcta o invertida del ritmo de actividad de los camarones. García, Petit y Fonteneau (1973) han demostrado la existencia de variaciones estacionales netas del ritmo de actividad “aparente” en diversos fondos de pesca del golfo de Guinea. Según la turbidez sea elevada o baja, este ritmo presenta un máximo de día o de noche. Estos autores han indicado que el paso del comportamiento “aparente” diurno (rendimientos más elevados por el día) a un comportamiento “aparente” nocturno (rendimientos más elevados por la noche) se traduce en una baja de la densidad aparente ligada a una baja de la capturabilidad. Ellos igualmente han indicado la existencia de estaciones con ritmo estable y estaciones con ritmo inestable. Es verosímil que esta inestabilidad tenga influencia sobre la eficiencia del esfuerzo de pesca.
En un trabajo mas reciente sobre la misma región, García (1978) demostró por medio del análisis de cohortes la existencia de variaciones estacionales de la capturabilidad que son sincrónicas con variaciones de la turbidez (Figura 59).
Marcille (1978), en Madagascar, ha demostrado igualmente que durante la estación seca la potencia de pesca relativa de ciertos barcos se aumenta por lluvias repentinas que aumentan la turbidez de las aguas costeras. Estos cambios de capturabilidad, ligados al comportamiento de los camarones y en particular a cambio de la extensión vertical de los cardúmenes, pueden afectar de manera diferente las capturas de artes de arrastre de diversa abertura vertical, perturbando las relaciones de estandarización (ver Sección 4.1.1.2).
8.4.1.2 La temperatura
Su acción sobre la capturabilidad ha sido percibida de forma contradictoria. García, Petit y Fonteneau (1973) han sugerido que en Senegal, las bajas temperaturas invernales (14–15°C por término medio) mejoran la capturabilidad global diaria y suprimen los ritmos de actividad. Los camarones entumecidos serán entonces igualmente capturados por el día que por la noche.
Por el contrario, Penn (1976) en Australia atribuyó el aumento de la capturabilidad observada a un aumento de la temperatura.
8.4.1.3 Las corrientes
Podemos recordar aquí el caso particular de los camarones en migración. Cuando los juveniles salen de las lagunas o estuarios, y la pesca se efectúa con la ayuda de artes pasivos, la captura obtenida por hora de pesca, o por día, dependerá por una parte de la abundancia real de camarones en estuarios (magnitud ligada al ciclo estacional de la reproducción), pero igualmente al volumen de agua filtrada por la red en la unidad de tiempo de pesca. Por lo tanto la captura dependerá de la velocidad de la corriente y la capturabilidad será más elevada en período de aguas vivas que en los de aguas muertas. También será mayor en el período de inundaciones que durante el estío.
Para un análisis más detallado de los problemas de evaluación del esfuerzo de pesca en este tipo de pesquerías nos referiremos a la Sección 4.1.2.2.
8.4.2 Las Variaciones Ligadas al Reclutamiento y a la Selectividad
Son conocidas las variaciones de capturabilidad relacionadas con la selectividad de las artes de arrastre, en función de la edad de una cohorte. Las curvas de evolución son en general de tipo sigmoideo, simétricas o no. En los casos de animales de larga vida donde la clase anual nuevamente reclutada no representa más que una pequeña parte del stock reclutado, este fenómeno no tiene una notable incidencia estacional sobre la capturabilidad global de la población. En el caso de animales de vida corta como los camarones, donde la duración de la vida en la pesquería es frecuentemente inferior a un año, la estructura de edades de la población está sujeta a unas variaciones estacionales importantes ligadas al mecanismo del reclutamiento (García, 1977). Una ilustración de este fenómeno y de su amplitud se presenta en la Figura 77. En estas poblaciones, la combinación de los fenómenos de selectividad y reclutamiento puede inducir a variaciones estacionales de la capturabilidad global de la población y, en particular, un aumento de q con la edad media de la población hasta que se complete el proceso de selección de la generación anual considerada. Dado que la participación de una cohorte en la pesquería es ligeramente superior a un año, esta evolución de q con la edad ligada a la selectividad se traduce, de hecho, en una variación estacional.
8.4.3 Las Variaciones de Abundancia Numérica
Por las mismas razones anteriores, la abundancia numérica de la población disponible está sujeta a unas variaciones estacionales cuya amplitud depende de las variaciones estacionales del reclutamiento. Si consideramos que todos los reclutamientos elementales dentro de un año constituyen una generación teórica es evidente que la proporción de esta generación disponible para la pesca varía a lo largo del año. Ello implica que la proporción de la generación anual total capturada por una unidad de esfuerzo variará con el tiempo, pasando por un máximo justo después del reclutamiento para luego disminuir hasta el siguiente reclutamiento. Estas variaciones son asimilables a variaciones de q y un esfuerzo dado producirá una mortalidad por pesca global distinta, según el período considerado del año.
8.4.4 Los Cambios de Indices de Concentración
Cuando el recurso está constituido por muchas especies diferentes y solamente se conoce el esfuerzo global, la variación estacional de la distribución relativa del esfuerzo total sobre las distintas especies produce variaciones estacionales de la capturabilidad específica. Veremos en las Secciones 8.5 y 8.6 que los cambios de la concentración del esfuerzo sobre las diferentes cohortes o grupos de edad de una especie dada, provocan unas variaciones de q en el espacio y según las especies.
8.4.5 Conclusiones
Las fuentes de variaciones estacionales de la capturabilidad, tomada aquí en su sentido más amplio, son numerosas. A pesar del hecho de que la mayor parte de ellas no han sido cuantificadas, su tendencia puede ser apreciada cualitativamente. Las variaciones ligadas a la dinámica de la población (abundancia numérica y edad media) tienden a contraponerse. Aquellas que están ligadas a las condiciones hidrológicas pueden combinarse u oponerse según los casos, siendo a priori difícil de definir analíticamente la resultante estacional de todas estas variaciones. Estas pueden, no obstante, ser calculadas por medio de marcados sucesivos o por análisis de cohortes (ver Capítulo 6).
Las consecuencias de este tipo de variaciones son las siguientes:
las variaciones de F con la edad serán distintas para cada cohorte elemental que contribuye a una generación anual;
la mortalidad total aplicada a la población por un esfuerzo anual dado dependerá de la distribución estacional de este último. Esto también es válido para la captura total y el rendimiento por recluta;
el resultado de una medida de ordenación, tal como una veda estacional, dependerá esencialmente de su posición dentro del año.
Estas variaciones estacionales deben ser tenidas en cuenta, tanto como sea posible, cuando se utilicen técnicas de simulación para determinar el efecto probable sobre la pesca y las medidas de ordenación.
En el caso de la pesca al arrastre, estas pueden estar ligadas a dificultades en el arrastre y dependen de la configuración del fondo, de su naturaleza, de la presencia de vegetación, de fuertes corrientes, etc. Este es un problema generalmente poco estudiado porque es fácil de evitar por el uso del esfuerzo medio, si el esquema del esfuerzo relativo sobre la especie considerada no cambia con el tiempo (índices de concentración constantes).
No obstante, cuando la distribución es heterogénea y la pesca está bien dirigida, como es el caso de los camarones gracias al arte de ensayo, el esfuerzo se concentra generalmente sobre las biomasas más grandes (Figura 60). La consecuencia es que si en cada punto la capturabilidad es la misma (desde que la proporción de la población presente en ese punto y capturada por unidad de esfuerzo es la misma), esta es globalmente superior en las zonas de gran abundancia (donde la proporción del conjunto de la población que es capturada por unidad de esfuerzo es más elevada). El resultado es que todo cambio en la distribución del esfuerzo sobre la biomasa implica variaciones en la relación entre F y f. Para resolver este problema, Beverton y Holt (1957, página 148) desarrollaron el concepto de intensidad de pesca global efectiva (overall effective fishing intensity).
La variabilidad espacial de q es aún más evidente en las pesquerías artesanales debido, por una parte, a la morfología de los estuarios y, por la otra, a la competencia entre artes de pesca. En efecto, la cantidad capturada por las redes fijas corrientemente utilizadas en la India y en Africa depende del tiempo de calado pero sobre todo del volumen filtrado. Es decir, del flujo de agua por el punto de pesca. Desde que éste as más elevado en zonas estrechas observamos frecuentemente un incremento de los rendimientos en los pasos estrechos (canales, pasos, puentes, etc.). Más aún esto complica singularmente la eventual reglamentación de la pesca ya que la consecuencia de la reducción de un número de redes variará según la zona considerada y la mortalidad total dependerá de la localización de los artes que sean retirados y de los que permanezcan.
Estas son de origen diverso y ya hemos señalado en los apartados 8.4.2 y 8.4.3 las variaciones relativas al fenómeno de la selectividad y del reclutamiento así como las variaciones ligadas a la abundancia numérica de la población y a su variación a lo largo del año. Además, es necesario destacar que la consecuencia de una concentración del esfuerzo sobre las zonas de mayor abundancia es la existencia de variaciones de q ligadas a la edad. En efecto, hemos visto en el apartado anterior (8.5) que la pesca se concentra sobre los estratos batimétricos y geográficos donde la densidad es más elevada (Figura 60). Al existir una estratificación paralela de tallas y edades, la consecuencia es una concentración del esfuerzo sobre las principales cohortes durante el período en que su biomasa es más importante (Figura 61).
La comprensión de estas variaciones es importante bajo la óptica de la utilización de los datos de esfuerzo anual para un modelo global de evaluación de recursos (tipo Schaefer o Fox, por ejemplo).
8.7.1 Cambios en la Distribución Estacional del Esfuerzo
Debido a la existencia de variaciones estacionales de q, todo cambió en la distribución relativa del esfuerzo en el transcurso de un año tendrá como consecuencia una modificación de la relación entre el esfuerzo nominal y la mortalidad por pesca que genera.
8.7.2 Cambios en las Especies Objetivo
Cuando se explotan muchas especies conjuntamente, la experiencia muestra que, a pesar del hecho de que son más o menos simultáneamente explotadas, la pesca se dirige en general hacia los lugares donde se encuentran las especies más rentables y se dirige más hacia especies secundarias cuando la abundancia de las primeras disminuye. Un fenómeno similar ha sido detectado en Senegal, donde se explota una sola especie, pero donde la transferencia del esfuerzo se efectúa del camarón hacia las especies de peces acompañantes que viven en los mismos fondos o en otros próximos (Lhomme, comunicación personal).
Sin una buena medida de la distribución real del esfuerzo por especies, su evolución para una especie dada será en general mal evaluada y la baja de los rendimientos de las especies objetivo será exagerada. Un ejemplo de un intento de solución de este problema en Australia ha sido presentado por Hall y Penn (1979).
8.7.3 Modificaciones en la Relación entre el Tiempo de Búsqueda y el Tiempo de Pesca
Hancock (1979) indicó que este concepto clásico bien conocido en pelágicos es válido para los camarones de comportamiento gregario muy marcado como Penaeus merguiensis. Si la disminución de la población a lo largo de los años se acompaña por una disminución del número de cardúmenes, con un aumento del tiempo de búsqueda en detrimento del de pesca, este último ya no es suficiente para medir correctamente la tendencia de la mortalidad por pesca y se hace necesario evaluar el tiempo utilizado en la búsqueda de cardúmenes.
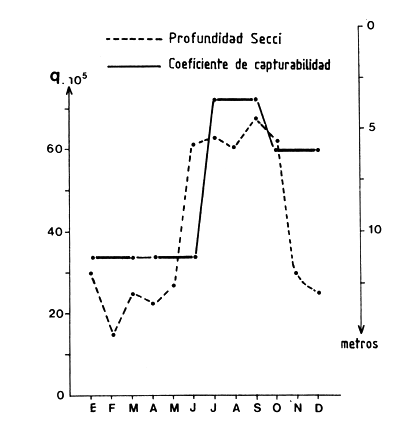
Figura 59 Variaciones estacionales del coeficiente de capturabilidad y de la turbidez, en Penaeus notialis (según García, 1979)
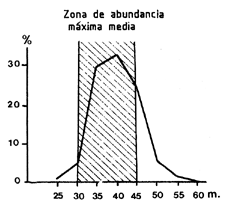 | 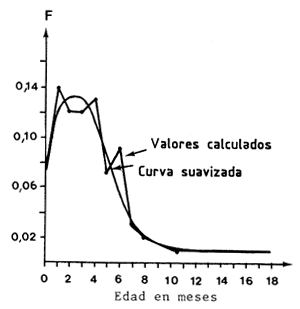 |
Figura 60 Repartición batimétrica del esfuerzo anual y de la abundancia en Côte d'Ivoire (según García, 1977) | Figura 61 Variaciones de F con la edad en Panaeus notiales (según García, 1977) |
![]()
![]()
![]()