![]()
![]()
![]()
Para animales tales como los peneidos, con una corta duración de vida, donde la captura anual depende casi enteramente de una clase anual reclutada durante ese año, los análisis de las variaciones interanuales de abundancia y el establecimiento de los modelos predictivos son esenciales para la explotación racional de los recursos. De hecho, las consecuencias de tales variaciones para la eficacia de las medidas potenciales de ordenación son importantes y serán tratadas en los siguientes Capítulos (11 y 12).
En primer lugar recordemos brevemente las diferentes causas posibles de variaciones en la abundancia anual, y después revisaremos las correlaciones varias establecidas con propósitos predictivos y sus mecanismos subyacentes en peneidos. En este tipo de población, las variaciones más importantes están obviamente ligadas a variaciones en reclutamiento anual lo que, a causa del número muy pequeño de generaciones que contribuyen a la captura, se traducen directamente en variaciones prácticamente proporcionales en la captura total, si el modelo de explotación permanece estable. Las variaciones en la capturabilidad son ciertamente responsables en parte de las variaciones temporales de las capturas pero, a priori, juegan un papel despreciable en las variaciones interanuales.
Existen también otras causas de variación, asociadas con las dinámicas propias de la problación. Aunque estas han sido hasta ahora muy escasas o en absoluto citadas en la literatura, nos pareció que requieren su inclusión para que esta revisión resulte completa.
9.1.1 Naturaleza de las Variaciones
La explotación industrial de los camarones peneidos es un hecho reciente en la mayoría de las regiones tropicales del mundo. Por lo tanto, las capturas en los últimos 20 años han variado enormemente como una función del esfuerzo de pesca, de las mejoras hechas a la potencia pesquera de las flotas y del desarrollo progresivo de la estrategia de explotación de especies individuales (a la iniciación de la pesca), a pesquerías multiespecíficas (cuando la pesquería está plenamente desarrollada), etc. Si a esto se añade el hecho de que raramente se obtienen buenas series de estadísticas pesqueras suficientemente completas y detalladas, que permitan análisis refinados, se comprende que los datos que permiten estudios a largo plazo de variabilidad de capturas a realizar, son mínimos.
Así, una de las primeras cuestiones es: ¿varía la abundancia de una población de camarones independientemente del esfuerzo? La respuesta es ambigua. De acuerdo con ciertos autores hay variaciones importantes, de acuerdo con otros la abundancia es estable. La estabilidad de la producción artesanal en ciertas zonas de Côte d'Ivoire a lo largo de un número de años es un argumento a favor del reclutamiento estable. Por otro lado, hay variaciones importantes en la pesquería artesanal de Senegal (Le Reste, 1979). Además Boerema (1974), señaló que las variaciones de abundancia en el mar son mínimas en Panamá pero grandes en Medio Oriente.
Las opiniones divergentes pueden relacionarse parcialmente a la amplitud de las series temporales disponibles, y obviamente las variaciones interanuales a corto plazo o “ruido” deben ser diferenciadas de las variaciones a largo plazo (“la señal interanual”). Cuando se dispone de series cortas de datos, la “señal” está a menudo enmascarada por el proceso de desarrollo de una pesquería que no está estabilizada y por lo tanto no es aparente. Se puede confundir con variaciones relacionadas al esfuerzo de pesca, por lo que sesga completamente la relación cpue/esfuerzo y la evaluación resultante.
Cuando por otra parte los datos disponibles lo permiten (Figuras 63–66) la “señal interanual” se distingue claramente. Es tentador buscar la causa de este fenómeno en variaciones climáticas a largo plazo, como han hecho muchos autores, según veremos más tarde.
Siendo este el caso, los espectros de variabilidad climática no son idénticos en la zona ecuatorial, tropical ni en las templadas. Por lo tanto, no sería sorprendente que la abundancia de las poblaciones de camarones en esas regiones esten sujetas a variaciones diferentes. Por ejemplo, se puede ver en la Figura 63 que el “ruido” a cada lado de las “señales” es mucho mayor en Texas (30 %, entre 1930 y 1945) que en Louisiana (15 % aproximadamente durante el mismo período).
El ejemplo utilizado es solamente una indicación, sin prejuicio por nuestra parte en cuanto a las verdaderas causas de esta variabilidad a largo y corto plazo. Nuestra intención es solamente llamar la atención respecto a los diferentes tipos de variaciones posibles y sus combinaciones, ya que pueden ser diferentes las consecuencias para la ordenación y el desarrollo de una estrategia de explotación óptima. Sería útil identificar las diferentes poblaciones mundiales por su espectro de variabilidad (amplitud y frecuencia de la señal interanual, amplitud de “ruido”) pero esto está fuera del alcance de esta revisión. Sin embargo, anotemos también que todos los estudios de este tipo deberían procurar establecer hasta qué punto la estabilidad “aparente” de las capturas en una pesquería multiespecífica refleja la habilidad de una flota para mantener su captura total ajustando su estrategia pesquera, antes que la estabilidad verdadera de las poblaciones subyacentes.
9.1.2 Las Variaciones del Reclutamiento
El ciclo vital de la mayoría de los peneidos incluye una importante fase litoral cuyo resultado óptimo se asocia con varios fenómenos.
9.1.2.1 Las variaciones de la reproducción
Hay pocos estudios a largo plazo que hacen posible examinar el orígen de esta variación. Aquí haremos referencia esencialmente al trabajo de Lhomme (1979).
La Figura 62 muestra la variación, desde 1973 a 1977, del porcentaje de hembras maduras en las capturas en función del tamaño. La información en esta curva está demasiado integrada como para permitir un análisis preciso, pero la figura muestra que el fenómeno de reproducción varía y que, para un tamaño dado, la proporción de individuos maduros observados depende del año. Aunque no es posible calcular la significación estadística de las diferencias observadas, está claro que 1975 fue un año anormal para las dos poblaciones de San Luis y Roxo-Bissagos y el de 1977 fue significativamente anormal con respecto a este último.
No conocemos hasta qué punto estas anomalías de la puesta afectaron al reclutamiento.
9.1.2.2 Supervivencia y emigración larvaria
Después de la puesta, las larvas resultantes deben llegar a zonas más someras donde encuentran la protección necesaria para su supervivencia y crecimiento. Obviamente la oportunidad de penetrar en los estuarios está íntimamente ligada a las condiciones hidrológicas (corrientes costeras, corrientes marinas, balance del total de agua del estuario, etc.).
Las fuertes lluvias y las riadas resultantes parecen ser, de acuerdo con los autores respectivos, un factor favorable ya que estos aumentan la extensión de la zona con un fuerte gradiente de salinidad a la cual las larvas pueden dirigirse por sí mismas, o desfavorable por simplemente obstruir la entrada de las larvas al estuario.
9.1.2.3 Supervivencia en los estuarios
Es evidente que está en el nivel larvario del mar y ciertamente con los estuarios, donde se efectúa lo esencial de la regulación del número en los penaeidos costeros. Muchos autores han sugerido que las lluvias fuertes y las riadas tienden a favorecer la supervivencia y el crecimiento, aumentando por una parte la superficie de las zonas de alevinaje disponibles (pero para las especies con requerimientos de sal esto puede tener el efecto opuesto), y de aquí reducir la competición intraespecífica, y por la otra aumentando la productividad del medio ambiente aportando nutrientes de los alrededores.
9.1.3 Las Variaciones Naturales de la Talla de Primera Captura
Estas variaciones están esencialmente ligadas a variaciones interanuales de la talla media en la migración de camarones jóvenes de regreso al mar.
La biomasa capturable justo antes o durante esta migración depende del número de poslarvas reclutadas, de su supervivencia hasta la migración, de su crecimiento y en consecuencia de las temperaturas predominantes y de la duración de la estancia en el estuario (Ford y St. Amant, 1971; Le Reste, 1979). El último autor mostró que el tamaño larvario en la migración variaba considerablemente en función de la salinidad del ambiente, con consecuentes variaciones en la biomasa capturable (Figura 66). No se debe olvidar que aquí se usaron redes fijas para pescar y que de hecho cualquier retraso en la migración resulta en un aumento natural de la edad a la primera captura en el estuario. El hecho de que haya un aumento resultante en la biomasa indicaría una mortalidad natural menor que aquella calculada por muchos autores (ver Capítulo 7).
Se resaltará más tarde (ver Capítulo 10 Sección 1.2) el hecho de que estos sucesos tienen consecuencias en el mar. La migración tardía de los camarones, de un mayor tamaño, tiene una curva de reclutamiento que se desplaza hacia tamaños mayores, originando aquí de nuevo variaciones interanuales de la talla de primera captura en el mar. Aunque los datos disponibles no permiten la medida de los efectos de esta variación en la abundancia anual, es probable que no sean despreciables.
9.1.4 Conclusiones
Hay muchas causas de variaciones interanuales en la abundancia de las poblaciones peneidos. Las relaciones de causa y efecto son difíciles de demostrar, incluso si a veces son una cuestión de pleno sentido común. Es posible que ciertos factores tales como las riadas, por ejemplo, puedan tener un efecto favorable aquí y desfavorable allí, dependiendo de su intensidad (efecto de umbral) o la forma del estuario (río o laguna, boca ancha o estrecha).
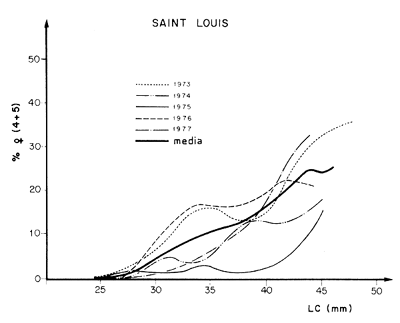
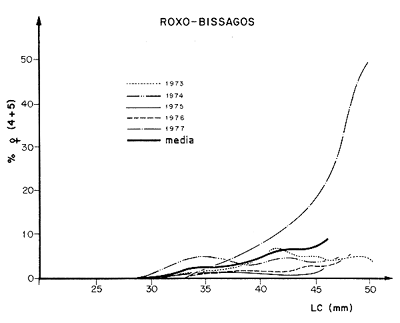
Figura 62 Variaciones del % de hembras maduras con la talla para Penaeus notialis en Senegal (según Lhomme, 1979b)
Desde que estas variaciones se han descubierto en muchas regiones, se han intentado predicciones, a partir de modelos de regresión simple, considerando la población marina o la del estuario como una “caja negra” que reacciona a un factor dado (lluvia, riadas, temperatura anual, etc.) para producir una cierta biomasa y así una cierta captura anual.
Estos se establecen por correlación, a partir de datos históricos, con la intención de llegar a las predicciones de captura. El principio es demostrar una relación cuantitativa suficientemente precisa entre la abundancia y un parámetro biológico o climatológico con objeto de predecir la variación interanual de captura. Para ser útil, el modelo debe permitir una predicción precisa con suficiente anticipación como para que la información sea utilizable.
Para los camarones, la captura anual depende casi enteramente del reclutamiento durante el año y solamente se pueden hacer predicciones a corto plazo. A largo plazo solamente se puede prever la tendencia general si es posible asociarla con un factor climatológico periódico (actividad solar, por ejemplo).
Se pueden distinguir dos tipos de modelos para los camarones según se establezcan en relación con un factor climatológico (lluvia, riadas, luz solar, actividad solar) o con un factor biológico (generalmente, la abundancia de una fase previa del ciclo vital).
9.2.1 Predicciones a Partir de Datos Climatológicos
9.2.1.1 Relaciones con las riadas y las lluvias
Debido a la importancia de las zonas costeras en el ciclo vital del camarón, y las principales funciones que parecen jugar los gradientes litoral-oceánico en los procesos migratorios durante el desarrollo, la influencia de los aportes continentales fue la primera en ser estudiada. Los resultados son contradictorios ya que las relaciones demostradas son bien positivas o negativas, de acuerdo con los casos que fueron examinados.
En los Estados Unidos de América, Hilderbrand y Gunter (1953) y Gunter y Hilderbrand (1954) demostraron, ya desde 1953, que en Texas había para P. setiferus buena relación entre la captura total y la lluvia de los dos años previos (r = 0,66) o durante el año en curso y los dos previos (r = 0,70). La relación es significativa al nivel del 1 % y las capturas altas se relacionan con lluvias más fuertes.
Los autores sugirieron una relación causa-efecto con la desalinización de las aguas costeras, correspondiendo posiblemente el tiempo de desfase observado al retraso entre las lluvias y su efecto sobre la salinidad del agua y, en consecuencia, sobre la producción de las poblaciones de camarón.
Varios años más tarde, Gunter y Edwards (1969) confirmaron estos resultados utilizando series más largas de tiempo (1927 a 1964) de las mismas especies en la misma región. Los análisis de los datos disponibles en Louisiana para las mismas especies no demostraron sin embargo una relación entre las capturas anuales y las riadas del Mississipi. La conclusión lógica fue que la salobridad estuvo allí siempre por encima del mínimo.
Este mismo tipo de análisis se aplicó por los autores a P. aztecus en Texas y Louisiana y no mostró una relación significativa, sugiriendo que incluso con más datos habría una correlación negativa; este resultado se consideró lógico para una especie que está supuestamente adaptada a un área con pequeños caudales de agua dulce y alta salinidad.
Más recientemente, este problema ha sido estudiado de nuevo por Barrett y Gillespie (1973, 1975) quienes demostraron que de hecho existe una correlación negativa en Louisiana entre el caudal del río en primavera y las capturas de P. aztecus; y también entre el de verano y las capturas de P. setiferus1.
Y = 3365 D-0,5064 (1,0 - 0,994746E)
donde E es el esfuerzo pesquero y D el aporte anual de agua dulce. En este modelo no fue considerada la posibilidad sugerida de que la relación entre aporte de agua dulce y producción puede no ser monótona
Según estos autores, los caudales excesivos en esta región donde el aporte de agua dulce es ya importante, limitarían las zonas de alevinaje disponibles (con la salinidad apropiada) limitando la producción. Esta explicación se confirmaría por el hecho de que la producción de P. aztecus en los estuarios de Louisiana es mayor cuando la salinidad es más alta en primavera.
Los aportes de agua dulce parecen así favorables para la producción en Texas y desfavorables en Louisiana para la misma especie, indicando por lo tanto que la relación positiva entre la baja salinidad y la producción de P. setiferus puede no ser una función lineal pero podría presentar un umbral tras el cual se invierte.
Más recientemente, Lhomme y García (1984) demostraron también en Senegal que las anomalías de la lluvia y la abundancia de la población estaban positivamente correlacionadas en la zona pesquera de San Luis (con baja entrada de agua dulce) y correlacionadas negativamente en Roxo-Bissagos donde el aporte de agua dulce es mucho mayor.
El trabajo de Gunter y Edwards (1969) sobre P. setiferus también depara comentario adicional. De hecho, la comparación de las variaciones de capturas anuales en Louisiana y Texas muestran una concordancia bastante buena entre las variaciones a largo plazo (Figura 63). Como resultado, mientras los párrafos previos parecen indicar diferencias en las reacciones a los aportes de agua dulce entre estas dos poblaciones, es también cierto que existen variaciones sincrónicas a largo plazo que podrían implicar la acción de un factor común, probablemente de origen climático.
Las variaciones sincrónicas también han sido demostradas por Berry y Baxter (1969) para las capturas anuales de P. aztecus en el noroccidente del golfo de México (Figura 64) pero este sincronismo podría derivar de la existencia de migraciones importantes dentro de toda el área (Barrett y Gillespie, 1975, página 3).
Habría sido interesante conocer si existía también algún sincronismo entre las variaciones de dos especies diferentes pero la observación positiva sobre esto por Barrett y Gillespie (1973) se contradice por los mismos autores en 1975.
En Australia, Thompson (1956) ha demostrado desde hace tiempo una correlación positiva entre las variaciones de lluvia y las capturas anuales de los juveniles (1924 a 1941) sin hacer alguna sugerencia en cuanto a las causas que subyacen. El observó en particular una buena correlación entre la captura y la lluvia de los dos años previos (r = 0,76), o las lluvias caídas de 6 a 12 meses antes de la temporada de pesca. También demostró, sobre 6 años solamente (1948–1953) variaciones similares entre la lluvia de un año y las capturas de adultos en el mar en la misma región al año siguiente.
Esta observación preliminar fue confirmada por Ruello (1973), con series más largas de datos (Figura 65). El observó las mejores correlaciones con la lluvia del año anterior (r = 0,64) así como aquellas del mismo año añadidas al año anterior (r = 0,69). Los aportes del río también demostraron tener una buena correlación positiva.
La observación de Thompson (1956) de una correlación positiva entre la lluvia y la producción juvenil en los estuarios está en contradicción con las observaciones de Castello y Ollinto Moller (1978) en Brazil, Le Reste (1979) en Senegal (Figura 65) y Barrett y Gillespie (1975) en Louisiana que habían demostrado una correlación negativa entre la lluvia y la producción estuarina.
De nuevo, las observaciones disponibles en la literatura indican una correlación negativa o positiva dependiendo del área o de la especie.
Se han adelantado varias hipótesis para explicar los fenómenos observados: el efecto directo en la fecundidad, la facilidad de la migración larvaria por un aumento en los grados de salinidad o el efecto detrimente de caudales excesivos durante el reclutamiento larvario, efectos sobre el habitat disponible en los estuarios y por lo tanto sobre la competición, crecimiento y supervivencia, etc., algunos de estos factores posiblemente actúan conjuntamente.
El aparente sincronismo de las variaciones a largo plazo en una gran parte del golfo de México sugieren el efecto de factores climatológicos operando a gran escala por un mecanismo difícil de analizar a causa de la complejidad del ecosistema y del ciclo vital concerniente.
La mayoría de las hipótesis adelantadas asumen que los factores climatológicos afectan el número de reclutas. Como se ha dicho ya anteriormente, algunos factores tales como los desbordamientos de los ríos, pueden ser o no beneficiosos y se puede asumir que quizás la relación no es lineal, aumentando la producción hasta un umbral dado, dependiente de las condiciones locales, y estando limitado más allá de él.
Un mecanismo bastante diferente, que implica a la salinidad induciendo a variaciones naturales de las tallas de primera captura, se puede deducir de los resultados obtenidos por Le Reste (1979). Este autor estudió las variaciones de producción de una gran pesquería artesanal en Casamance (Senegal), que es parte del año un estuario hipersalino. el demostró que el aumento en el tamaño medio anual de los camarones capturados, registrado desde el principio de la sequía del Sahel, estaba asociado con el aumento de la salinidad en el estuario. El también demostró que en los años secos, cuando el tamaño en la migración (y por lo tanto en la primera captura) es grande, las capturas son cuantiosas y viceversa. La salinidad en el estuario de Casamance, que se nutre principalmente del agua subterránea, depende de las lluvias caídas años anteriores y Le Reste demostró una buena correlación negativa entre las capturas de un año y la lluvia de los años anteriores. Esto se interpreta por la influencia de la salinidad en la duración de la estancia en el estuario, una explicación avanzada y sostenida por García (1977) para la Côte d'Ivoire y Nigeria. Una estancia más extendida en el estuario aumentaría, a través de la interacción de crecimiento y mortalidad, la edad a la primera captura y la biomasa capturable. El uso de la correlación como modelo predictivo ha permitido buena concordancia entre las capturas observadas y las calculadas, que serán obtenidas (Figura 66). La calidad y credibilidad de la predicción deberá probarse en los próximos años.
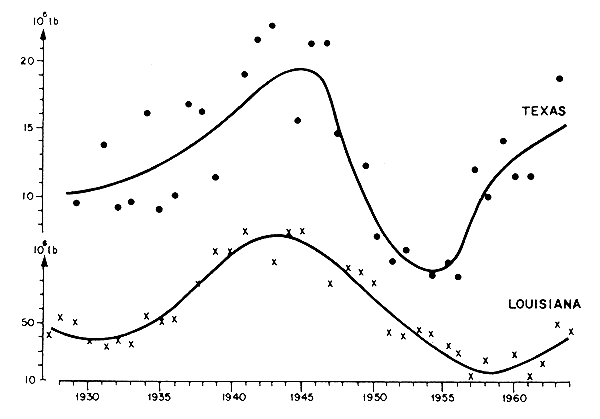
Figura 63 Variaciones a largo plazo de los desembarcos de camarón Penaeus setiferus en Texas (puntos) y en Louisiana (cruces) (según los datos de Gunter y Edwards, 1969)
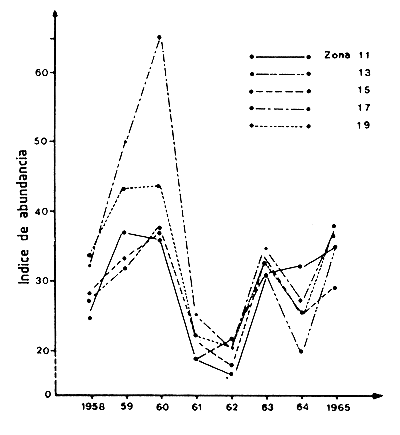
Figura 64 Variaciones interanuales de la abundancia de Penaeus aztecus en diversas zonas estadísticas del norte y noroeste del golfo de México, de Alabama a México (según Berry y Baxter, 1969)
Por último, debe señalarse que ciertos autores han intentado relacionar, mes a mes, la lluvia y las capturas con un cierto desfase de tiempo. La correlación es generalmente excelente ya que relaciona dos factores temporales, pero no puede utilizarse para la predicción de las capturas anuales.
9.2.1.2 Relación con la temperatura
Williams (1969) demostró, en los Estados Unidos, una relación entre la temperatura característica del año y la abundancia de camarones expresada como captura por unidad de esfuerzo. La introducción de un desfase de tiempo no mejoró notablemente la relación. Cuando él agrupó los datos de Georgia, las Carolinas y Texas, las capturas fueron mayores durante los años más templados. Sin embargo, el autor anotó que, aunque estas relaciones eran todavía aparentes, eran peores si se consideraban las diferentes regiones separadamente.
De hecho, esta relación no parece ser útil para predecir, con buena precisión, las variaciones interanuales en cada región separadamente y es solamente significativa a nivel de toda la costa sudeste de los Estados Unidos; es solamente de esta forma que el rango de los valores térmicos anuales es suficientemente amplio como para permitir que ocurra un tipo de relación morfo-edáfica, demostrando de hecho que todo el potencial de camarón de una región relaciona con su perfil térmico anual (Figura 67). Esto se comprende fácilmente cuando se reconoce que el crecimiento esencial y los mecanismos reproductivos de estos animales tropicales son dependientes de la temperatura. El autor ha señalado además que los datos a lo largo de la línea de regresión están de hecho dispuestos en un orden latitudinal. Estas observaciones se deben tomar en unión con las de Turner (1977) que serán en la siguiente sección.
Baxter (1969) estableció en Texas una relación de tipo diferente demostrando que había una conexión entre la fecha de mayor migración anual juvenil hacia el mar y la temperatura del aire en abril del mismo año (Figura 68). Esta predicción podría ser útil para determinar con precisión el período en el que debería hacerse el muestreo así como para determinar un índice de abundancia para los juveniles en migración. En la misma línea, un trabajo llevado a cabo en Louisiana (Barrett y Gillespie, 1973, 1975) indica que la captura en mayo de P. aztecus en el estuario depende de la temperatura en abril.
9.2.1.3 Relación con la latitud
Turner (1977) demostró que había, a escala mundial, una conexión entre la producción (Y) a la latitud (X en grados) de la forma
Y = 159e-0,070(x)
9.2.1.4 Relación con la actividad solar
Aunque no existe este tipo de estudio sobre camarones peneidos, es interesante señalar que Driver (1979) demostró buena correlación entre la producción de Crangon crangon (un camarón carideo) y el ritmo de actividad solar. Un estudio previo demostró que se podrían hacer predicciones a uno o dos años vistas basadas en mediciones de la pluviosidad y las temperaturas del aire. Al tener las variaciones climáticas interanuales demostradas conexiones con la actividad solar, el autor intentó y obtuvo buenas correlaciones (r = 0,86) con una periodicidad de 11 años.
9.2.1.5 Relación con la superficie de las zonas intermareales
Turner (1977) también demostró que las capturas de las lagunas de una región estaban asociadas con la superficie de las zonas intermareales. No obstante, él aceptó que esta relación podría ser simplemente una consecuencia de la relación entre la producción del estuario y el tamaño del “interfase” mar/estuario. De la misma manera, Barret y Gillespie (1973, 1975) demostraron una relación entre la variación interanual del área del estuario adecuada para la vida del camarón (salinidad por encima de 10 ppm) y la captura.
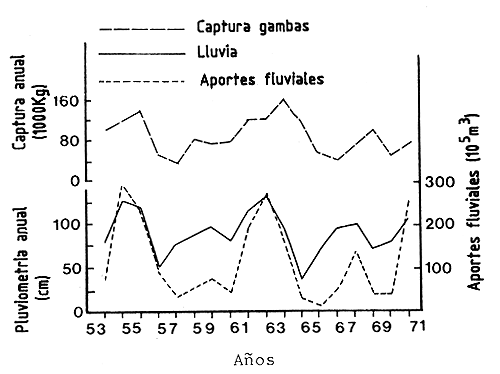
Figura 65 Variaciones interanuales de las lluvias, los aportes fluviales y las capturas de Metapenaeus mackeyi (según Ruello, 1973)
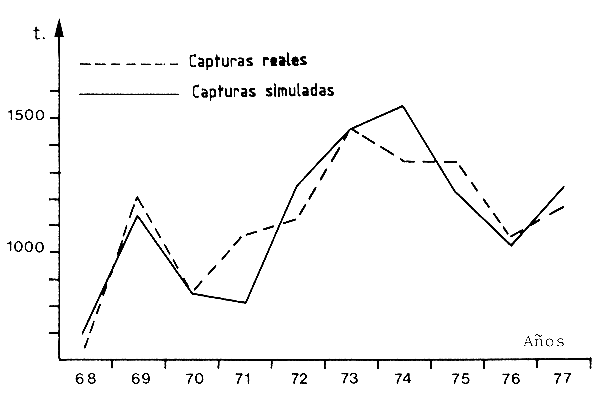
Figura 66 Evolución de las capturas reales (----) y simuladas (—) en la pesquería de juveniles de Casamance (Senegal) (según Le Reste, 1979)
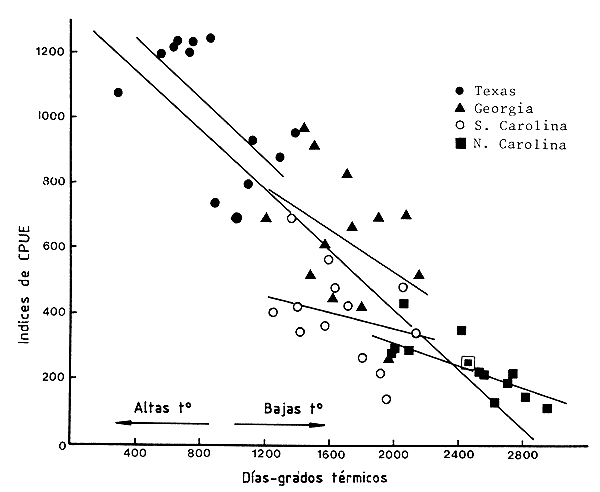
Figura 67 Relación entre el balance térmico anual y los índices de abundancia (según Williams, 1969)
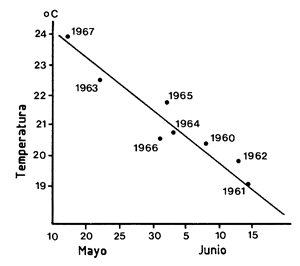
Figura 68 Relación entre la temperatura en abril y la fecha de máxima abundancia de juveniles en la bahía de Galveston (según Baxter, 1969)
9.2.1.6 Conclusiones
Existen suficientes indicaciones para demostrar que la producción anual de camarón varia de un año a otro en relación con factores climatológicos a larga escala. Se puede asumir que subyace una relación causa-efecto, lo que sin embargo será muy difícil de probar debido a que los mecanismos son ciertamente complejos e incluyen la modificación de varios parámetros (fecundidad, reclutamiento poslarvario, edad en migración, crecimiento y supervivencia, capturabilidad) por las condiciones climatológicas.
Los parámetros climatológicos que se han medido (lluvia, caudales de ríos, perfil térmico anual, o actividad solar) pueden ser en el fondo sólo indicadores de variaciones climatológicas que afectan otros muchos parámetros (horas de sol y, en consecuencia, producción primaria y fotoperiodicidad, salinidad, cronología o amplitud relativa de las estaciones, desplazamientos de frentes, posición o fuerza de corrientes y corrientes submarinas) que son también capaces de influenciar la producción biológica de las poblaciones de camarones.
Finalmente, no es necesario entender sin embargo todos los mecanismos a fin de usar una relación empírica como herramienta predictiva si la precisión del diagnóstico se estima ser suficiente. Es obvio que las relaciones anteriores se establecieron con este propósito. No obstante, se debe enfatizar que, de acuerdo con Marr (1972), este tipo de relación es a menudo sólo precisa en condiciones extremas y que, cuando son intermedias, la variación más fuerte está a menudo ligada a factores bióticos (competición, etc.). Se deduce, por lo tanto, que un parámetro dado pueda ser solamente informativo en zonas donde sea un factor limitante y que de aquí sólo se puedan predecir los años anormales con suficiente precisión.
Ya que la relación no es necesariamente lineal, las previsiones no deben estar basadas en extrapolaciones fuera del rango de las observaciones.
Los ciclos vitales del camarón atraviesan varias etapas, poslarvaria, juvenil y adulta. Dos de estas corresponden a una migración donde se concentran los camarones y el muestreo es más simple. Muchos estudios han intentado demostrar desde los sesenta la relación entre la abundancia en una fase temprana y en otra posterior, fase explotada, con el fin de obtener un índice de pre-reclutamiento y un simple modelo predictivo.
9.3.1 La Relación entre Abundancia de Adultos y la de su Prole Larvaria y Poslarvaria
Según Neal (1975), no ha sido posible demostrar ninguna conexión. El sugiere que esto indica que la variabilidad en la fase larvaria y poslarvaria está relacionada esencialmente con el medio ambiente. Los autores no están al tanto de observaciones directas sobre este tema. No obstante, es cierto que la reproducción está sujeta a fuertes variaciones de un año al siguiente (Sección 9.1.2.1.)
9.3.2 La Relación entre Abundancia Larvaria en el Mar y Abundancia de Adultos de la Misma Generación
Siempre de acuerdo con Neal (1975), no se ha observado alguna relación. Debe notarse que el obstáculo principal para estos estudios reside en obtener un índice suficientemente preciso de abundancia larvaria en el mar a un costo razonable. Los problemas proceden de una parte de las características de la distribución del plancton (ver Capítulo 2) y, por otra parte, del rápido desarrollo (con alta mortalidad larvaria) a través de las muchas etapas larvarias sucesivas que son una característica del desarrollo peneidos. De hecho, la abundancia en un momento dado de una cierta fase larvaria (protozoea por ejemplo) depende de la intensidad de la puesta y también del tiempo transcurrido entre la puesta y el muestreo, a causa de la “migración” permanente hacia y de la fase considerada y a la mortalidad larvaria.
9.3.3 La Relación entre la Abundancia Poslarvaria y la Subsecuente de Juveniles o Adultos
Cuando se dispone de datos suficientes sobre la variación de la abundancia de dos estados sucesivos dentro de un año, es posible encontrar una relación entre los máximos respectivos de abundancia (Figura 69).
Entonces, cuando hay largas series de abundancias en varias etapas de desarrollo, de muchos años, se pueden calcular los índices respectivos de abundancia y su correlación, de ser significativa, podría ser utilizada como una herramienta predictiva (Figura 70).
Baxter (1963) en la conclusión de un estudio de 3 años sobre P. aztecus en Texas, demostró buena correlación entre variaciones estacionales de abundancia de poslarvas, juveniles y adultos en las lagunas. El sugirió que la predicción puede ser posible pero que solamente un estudio detallado puede confirmarlo (Figura 69). Posteriormente, Berry y Baxter (1969), después de siete años de estudios en la misma región, señalaron la ausencia de una relación utilizable entre la abundancia poslarvaria y la captura para carnada o comercial. Ellos sugirieron que el muestreo mediante la red Renfro (ver Sección 2.3.2.4) debería ser revisado. Estos resultados están corroborados por los de St. Amant, Broom y Ford (1965) quienes demostraron que en Louisiana no existía una buena relación entre la abundancia poslarvaria y la juvenil. Ello indicaría que la producción depende principalmente de la supervivencia larvaria en los estuarios y que había una regulación considerable de los números de la población en ese estado. Subrahmanyam (1966) y Subrahmanyam y Rao (1970) señalaron que, durante dos años sucesivos, la abundancia poslarvaria y las capturas variaron en la misma dirección. Sin embargo, estos estudios se hicieron sobre períodos demasiado cortos como para evaluar el valor cuantitativo de la relación. Lo mismo se aplica al trabajo de dos años de Christmas, Gunter y Musgrave (1966), cuyos resultados fueron sin embargo confirmados por Christmas, Langley y van Devender (1976) quienes obtuvieron una predicción (válida en aproximadamente un 10 %) sobre un período de 3 años.
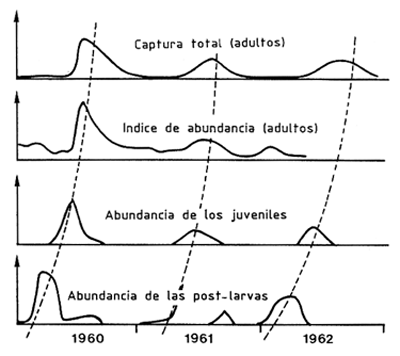
Figura 69 Relaciones entre las variaciones estacionales de los índices de abundancia de las post-larvas, juveniles y adultos (simplificado de Baxter, 1963)
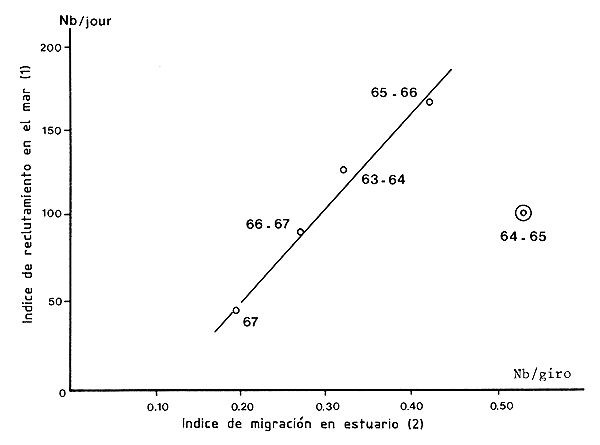
Figura 70 Relación entre la migración y el reclutamiento (modificado de Yokel, 1969)
1. Captura anual media/día de subadultos (68/libra y mes)
2. Indice de abundancia anual de subadultos en migración (número/giro de flujómetro)
Aún más recientemente, y mediante un estudio de 2 años, Roessler y Rehrer (1975), demostraron la buena correlación entre las variaciones climáticas en la abundancia poslarvaria en los Everglades y el índice de reclutamiento de los subadultos en el mar. La alta correlación observada (r = 0,78) es normal ya que se establece entre dos factores estacionales.
De hecho, aparte del trabajo de Berry y Baxter, no existen, a nuestro entender, ejemplos bien fundamentados que se extiendan sobre un período suficientemente largo, y la cuestión de la utilidad práctica de los índices de abundancia poslarvaria para predecir la abundancia juvenil o adulta queda sin una respuesta definitiva. De hecho, la respuesta podría variar según el ecosistema considerado.
9.3.4 La Relación entre la Abundancia de Juveniles y de la Población de Adultos en el Mar
Yokel, Iversen e Idyll (1969) y Yokel (1969) demostraron con P. duorarum de Florida que si se tomaban en cuenta las variaciones estacionales de tamaño en la migración (implicando diferentes desfases de tiempo), existía buena correlación entre los índices de abundancia númerica de los juveniles migratorios y los de los jóvenes reclutas en las zonas de pesca (Figura 70). Los rendimientos pesqueros de los juveniles podrían ser por lo tanto utilizados como índices de reclutamiento. Berry y Baxter (1969) han demostrado, sobre P. aztecus de Texas, que había una buena correlación entre las capturas de juveniles en la pesquería de camarón para carnada (desde el 25 de abril al 31 de agosto) y las capturas de adultos en el mar (entre julio y septiembre). Caillouet y Baxter (1973) dieron ecuaciones de las relaciones obtenidas para Texas y Louisiana. En Texas existen excelentes correlaciones (r = 0,85) para P. aztecus si la captura anual en el mar se relaciona con el índice de abundancia juvenil en la bahía de Galveston1 y otras mediocres (r = 0,28) si esta captura se relaciona con la captura anual de los estuarios2. Con el mismo tipo de datos de P. setiferus, la correlación es regular (r = 0,58). En Louisiana, la correlación entre la captura anual en el estuario y la del mar es excelente para P. aztecus (r = 0,73) y P. setiferus (r = 0,86), lo que ha sido confirmado por Barrett y Gillespie (1975).
Las correlaciones entre las capturas anuales en estuario y en el mar indican que los años buenos en estuarios son también años buenos en el mar (y viceversa) pero no pueden ser utilizadas como predicciones porque las dos variantes se basan sobre el mismo período de tiempo.
9.3.5 Conclusiones
Cuando se intenta relacionar las capturas de camarones en el mar con la abundancia de una de las fases precedentes del ciclo vital, parece que sólo los subadultos migratorios pueden dar un índice de predicción útil a corto plazo (aproximadamente con 3 meses de anticipación). Esto necesita ser confirmado para poslarvas, con series de datos más largos.
Las conclusiones optimistas de Berry y Baxter (1969, p. 797) sobre el posible uso de la abundancia poslarvaria con propósitos de predicción, si puede mejorarse el muestreo, no han sido ni confirmadas ni desmentidas ya que no han sido llevados a cabo otros trabajos de similar envergadura. Debido al relativo alto costo de la obtención de tales datos, en comparación con los obtenidos de las pesquerías sobre los juveniles, no está claro que tal programa de trabajo pueda implantarse y sea continuado rutinariamente en el futuro.
![]()
![]()
![]()