![]()
![]()
![]()
Bunvong Thaiutsa y Orman Granger
BUNVONG THAIUTSA es profesor de la Facultad de Ciencias Forestales de la Universidad Kasetsart, Bangkok, Tailandia y ORMAN GRANGER es profesor del Departamento de Geografía de la Universidad de California, Berkeley, California.
Desde hace mucho tiempo se conoce la importancia del piso del bosque como parte integral del ecosistema. Uno de los principales procesos que tiene lugar en el piso del bosque es el de la «descomposición», término que se emplea en general para referirse a todos sus cambios biológicos, químicos y físicos, tales como el peso. Wood (1974) define la descomposición de las hojas de Eucalyptus como la pérdida de peso debida a varios factores, entre los que se cuentan la eliminación y/o el consumo de tejidos por invertebrados que se alimentan de hojas, por lixiviación, por degradación bioquímica por microorganismos y por degradación bioquímica a su paso por el tubo digestivo de los invertebrados. La descomposición se produce en gran parte en el piso del bosque, donde los primeros colonizadores de la hojarasca fresca son las bacterias y los hongos.
Los ecólogos que se ocupan de las áreas silvestres han prestado mucha atención a la descomposición de la hojarasca, en relación con el ciclo de los nutrientes y con la productividad del suelo. Es evidente que lo hacen porque la descomposición de la hojarasca influye mucho en la disponibilidad de nutrientes, que es un determinante fundamental del crecimiento de los árboles y de la producción de madera.
Para estudiar la descomposición de la hojarasca se emplean tres técnicas principales: (1) medición de la evolución del dióxido de carbono o de la absorción de oxígeno (Reiners, 1968; Wiant, Jr., 1967a, 1967b; Witkamp, 1966); (2) observación de la desaparición del disco de las hojas y de un constituyente específico, como la celulosa y la lignina (Edwards y Heath, 1963; Heath et al., 1964; Heath et al., 1966; Madge, 1965) y (3) las actividades de la macro y microfauna, en las condiciones particulares del substrato y del ambiente.
El follaje de las coníferas suele descomponerse más lentamente que el de las deciduas (Daubemire y Prusso, 1963; Gosz et al., 1973; Mikola, 1960; Ovington, 1954), porque la hojarasca de las latifoliadas contiene más potasio, más fósforo, menos lignina y casi siempre menos fracción soluble en éter que la hojarasca de las coníferas. También se observan diferencias significativas entre especies (Edwards y Heath, 1963; Heath et al., 1966). Pradhan (1973) señala que la descomposición de la hojarasca de la teca es más rápida que la de la hojarasca de Acacia arabica. Además, las hojas caídas desaparecen mucho más rápidamente que las ramas (John, 1973; Rochow, 1974) y las hojas de sombra, que son más blandas, más rápidamente que las hojas de luz (Williams y Gray, 1974).
Por lo general, el índice de descomposición es más alto en las especies que tienen un máximo de ceniza y de nitrógeno y una mínima relación C/N y de lignina. El índice más bajo de descomposición se observa en especies relativamente pobres en ceniza y nitrógeno, que tienen un máximo de lignina y una máxima relación C/N. Las especies que tienen una cantidad mediana de ceniza, nitrógeno y lignina y una relación C/N media, experimentan una descomposición mediana (Singh, 1969). Broadfoot y Pierre (1939) observaron una correlación muy significativa entre la descomposición de la hojarasca y cinco variables independientes: exceso de bases, materia orgánica soluble en agua, nitrógeno total, ceniza total y calcio total. El coeficiente de correlación múltiple entre el porcentaje de descomposición y tres variables: nitrógeno total, materia orgánica soluble en agua y exceso de bases fue de 0,86. Kucera (1959) también indica una correlación positiva entre la rapidez de la descomposición y un alto contenido de ceniza y de sustancias solubles en agua caliente. Como la composición química de la hojarasca influye en su descomposición, se supone que determina en forma muy significativa la liberación de nutrientes.
Los factores dominantes de la actividad microbiana en las hojas secas son la especie vegetal y el clima. El efecto de la especie vegetal sobre la microflora depende de las propiedades físicas y químicas de las hojas; ambas influyen en la relación C/N. Crossley y Hoglund (1962) señalan una correlación significativa entre el número de microartrópodos en los sacos de hoja rasca y la descomposición de la hojarasca. En invierno hay pocos animales en los discos de las hojas y éstos comen menos tejido de hojas que en el verano, cuando un mayor número de animales de la superficie del suelo come más hojarasca.
El número de artrópodos en la hojarasca depende además de la humedad. En efecto, Madge (1965) llega a la conclusión de que en la época húmeda hay más animales en los discos de las hojas que en la época seca. Al excluir los animales por completo, durante un periodo de nueve meses, no se registra descomposición visible de los discos de las hojas de encina y de haya (Edwards y Heath, 1963). Estos autores afirman que las lombrices eliminan los discos tres veces más rápidamente que los invertebrados pequeños, siendo los más importantes entre estos últimos los tisanuros, los enquitraeidos y las larvas de dípteros. Un insecticida corriente de carbamato (carbofurán), aplicado en las dosis recomendadas, reduce la descomposición del arce rojo a valores de 0,99 y 1,26 g/m²/día. Esto se debe principalmente a que los insecticidas de este tipo son muy tóxicos para las lombrices, según se ha observado (Weary y Merriam, 1978).
UN BOSQUE TROPICAL - el mecanismo de la descomposición de la hojarasca son criterios importantes
El clima modifica notablemente la índole y la rapidez de la descomposición de los restos vegetales en la superficie del suelo, de modo que ejerce una influencia importante en el tipo y la abundancia de la materia orgánica. La humedad y la temperatura se cuentan entre las variables más determinantes (Brinson, 1977; Singh, 1969), porque influyen tanto en el desarrollo de la vegetación como en las actividades de los microorganismos, que son factores muy críticos de la formación del suelo. Kononova (1975), citando varias otras publicaciones, llega a la conclusión de que la intensidad máxima de la descomposición de la materia orgánica se observa en condiciones de temperatura moderada (alrededor de 30°C) y con un contenido de humedad de alrededor del 60-80% de su capacidad máxima de retención de agua. El aumento o disminución de la temperatura y de la humedad simultáneamente, más allá de los niveles óptimos, produce una disminución de la descomposición de la materia orgánica, de modo que no conviene comparar estudios de descomposición de la hojarasca si los regímenes de temperatura y humedad no son iguales.
Resultados de un estudio de cuatro años sobre la descomposición de la hojarasca en un bosque perenne
Wood (1974) presenta regresiones lineales entre: (1) la descomposición microbiana y la lluvia (r = 0,80) (2) la descomposición y el carbono orgánico (r = 0,75) y (3) la descomposición y el pH (r = - 0,60).
El pH obtenido refleja la interacción entre la lluvia y la temperatura, revelando una descomposición lenta tanto con pH bajo (temperatura baja y precipitación abundante) como con pH alto (temperatura alta y precipitación escasa) y descomposición rápida con pH de nivel medio.
Williams y Gray (1974) señalan que hay diferencias en el tiempo que tarda la descomposición a diferentes altitudes, debido a la variación de la temperatura. Shanks y Olson (1961) comparan la descomposición de la hojarasca en rodales naturales a diversas altitudes y llegan a la conclusión de que hay una disminución media de la descomposición de casi 2% por 1°C de disminución de la temperatura media. Este resultado es semejante al obtenido por Mikola (1960), quien observa que la descomposición disminuye con el aumento de la latitud. Es notable la influencia de la temperatura sobre la descomposición de la lignina. A 37°C la lignina se descompone rápidamente y en nueve meses desaparece en un 50-60% (Waksman y Gerretsen, 1931). Meentemeyer (1978) emplea la evapotranspiración anual efectiva como variable predictiva índice de la descomposición.
Reiners (1968) establece la correlación entre los factores ambientales y la tasa de evolución del dióxido de carbono y observa que a 5 y 15 cm de profundidad en el suelo las temperaturas medias tienen una correlación muy grande con la producción de dióxido de carbono. Por cada 10°C de aumento de la temperatura, entre 20 y 40°C, la producción de dióxido de carbono aumenta al doble (Wiant, Jr., 1967a). No se registra la presencia de dióxido de carbono a 10°C, que disminuye notablemente a 50°C o más. La humedad también influye mucho en la evolución del dióxido de carbono. En materia orgánica no incorporada, en la cual la humedad ha disminuido a menos de 20%, o en suelo mineral donde ha disminuido a sólo 5%, no se puede detectar la producción de dióxido de carbono (Wiant, Jr., 1967b).
Pero se informa de algunas conclusiones contrarias sobre los efectos del ambiente climático. Van der Drift (1963) señala que la precipitación es más importante que la temperatura en la descomposición de la hojarasca, a pesar de que los inviernos benignos y las primaveras tibias, sumados a una humedad del suelo relativamente normal, pueden acelerar la descomposición. La humedad que atraviesa los restos orgánicos puede ser importante para la aceleración de la descomposición. El agua de lluvia que se infiltra puede arrastrar los excrementos y los restos de organismos hacia horizontes más profundos, donde otros microbios especializados atacan las moléculas orgánicas restantes. Por tal motivo en los años de sequía la descomposición es más lenta que en los años normales. Por otra parte, Jenny et al. (1949) y Witkamp (1966) subrayan la importancia de la temperatura, y luego la de la precipitación.
Como el clima es el factor principal que controla la descomposición, ésta varía según la época del año. Gosz et al. (1973) y Loomis (1975) observan que la descomposición de los residuos vegetales es rápida en el verano; en cambio Lang (1974) estima que la descomposición de la hojarasca es de 3,75 g/m²/día en los meses de otoño y de 0,80 g/m²/día en los restantes. Boonyawat y Ngampongsai (1974) confirman los resultados de Lang, ya que establecen que la descomposición máxima de la hojarasca en bosques perennes de colina tiene lugar a fines de la época de las lluvias y a comienzos del invierno (0,36 t/ha/mes) y la mínima en verano (0,14 t/ha/mes). Madge (1965) llega a la conclusión de que la desaparición de la hojarasca en Nigeria tiene lugar principalmente en la época de las lluvias, debido a la actividad de los ácaros y de Collembola.
A pesar de que Jenny et al. (1949) estudiaron hace algunos decenios las diferencias de la descomposición en las regiones templadas y tropicales, se sabe de pocas investigaciones en el Asia sudoriental. En la mayoría de los estudios anteriores sobre la producción y la descomposición de la hojarasca en los bosques de Tailandia (Aksornkoae et al., 1972; Boonyawat y Ngampongsai, 1974; Paovongsar, 1976; Thaiutsa et al., 1978) se registra el ambiente climático, pero se descuida la interpretación analítica. Por tal motivo, el presente estudio tiene por objeto elaborar modelos de regresión, para predecir la descomposición en base a la información climática.
Cuadro 2. Ecuaciones de regresión lineal entre el porcentaje de descomposición (D) y la humedad relativa (HR, % ), la precipitación (P, mm), la temperatura media (T, °C), la temperatura máxima (Tmáx, °C), la evapotranspiración efectiva mensual (EEm, mm), la relación de sequedad (RS) y el hidrofactor (Hf) del bosque perenne montano (Chiang Mai, Tailandia)
|
Ecuaciones |
r |
r 2 |
|
D = 2,286 HR - 125,481 |
0,89 |
0,79 |
|
D = 30,096 + 0,149 P |
0,77 |
0,59 |
|
D = 242,712 - 7,860 Tmáx |
-0,52 |
0,27 |
|
D = 26,267 + 14,122 RS |
0,85 |
0,72 |
|
D = 0,891 Hf - 2,103 |
0,75 |
0,56 |
|
D = 1,626 HR - 0,466 EEm + 0,899 P - 53,948 |
0,93 |
0,86 |
|
D = 2,284 HR - 0,910 EEm + 6,222 T + 0,607 P - 192,715 |
0,94 |
0,88 |
La investigación se realizó en la estación de investigación de hoyas hidrográficas Kgma situada en el norte de Tailandia, a 18°45'N de latitud y 98°54'E de longitud. El declive es de menos de 20% y la altitud entre 1250 y 1500 m. El origen del suelo es granito, gneiss y cuarzita; ha sido clasificado como perteneciente al gran grupo de los suelos lateríticos pardorrojizos. Tiene una textura francoarenosa que, sumada a una gran lixiviación, produce un horizonte B profundo. Según Spurr y Barnes (1973) el subtipo de bosque perenne que se observa aquí se denomina «bosque pluvial montano». Las principales especies arbóreas son las de la familia Fagaceae: Castanopsis acuminatissima, Lithocarpus polystachyus, Quercus kerrii y Quercus kingiana. Las copas cubren alrededor del 85%.
En el piso de bosque se colocaron al azar 156 trampas de madera abarcando toda la superficie de la hoya hidrográfica de 0,65 km², para medir el insumo y el producto de la hojarasca. Las dimensiones de las trampas eran de 0,75 × 0,75 m. Se protegieron de la descomposición 12 trampas. Se recogió hojarasca mensualmente desde enero de 1968 hasta diciembre de 1972. Se secó la hojarasca a 70°C durante 24 horas y se anotó el peso, suponiéndose que la pérdida de peso representa la tasa mensual de descomposición. Se registraron la temperatura, la precipitación y la humedad relativa.
Se calcularon las componentes del balance del agua con el programa de computadora del balance del agua de Stone, Jr. (1971). Este cómputo se basa en las tablas de Thornthwaite y Mather (1957) que contienen los valores de la evapotranspiración potencial no reajustada y se basan en los valores de la temperatura media y de I (índice del calor). Estas tablas dan además la insolación mensual media a esta latitud.
De los datos climáticos primarios se derivaron dos variables secundarias. Se estimó el hidrofactor con la ecuación empírica citada por Kononova (1975): Hf =43,2 log P - T, en la cual Hf es el hidrofactor, P es la precipitación mensual en mm y T es la temperatura media en °C. La otra variable, relación de sequedad (RS), se aproximó con la ecuación RS = P/EP, en la cual P es la precipitación mensual en mm y EP es la evapotranspiración potencial expresada en mm.
Cuadro 3. Matriz de la correlación y cuadro resumen de la correlación múltiple gradual
|
Variable |
X1 Descomposición |
X2 Humedad |
X3 EE mensual |
X4 Precipitación |
|
Porcentaje |
mm. |
|||
|
X1 |
1,00 |
0,89 |
0,17 |
0,77 |
|
X2 |
|
1,00 |
0,35 |
0,80 |
|
X3 |
|
|
1,00 |
0,61 |
|
X4 |
|
|
|
1,00 |
|
Grado número |
Variable incluida |
r |
r2 |
r2 aumento |
Significación |
|
1 |
X2 |
0,89 |
0,79 |
0,79 |
0,000 |
|
2 |
X3 |
0,90 |
0,81 |
0,02 |
0,001 |
|
3 |
X4 |
0,93 |
0,86 |
0,05 |
0,001 |
Se hicieron regresiones simples y múltiples entre el porcentaje de descomposición de la hojarasca del bosque y las variables climáticas primarias y secundarias. Se empleó una técnica gradual selectiva en las regresiones múltiples. Con este método se calculó una secuencia de ecuaciones de regresiones múltiples en forma gradual para cada relación. En cada grado, la variable sumada es la que produce la reducción máxima de la suma de los cuadrados de los errores, o sea la que tiene la correlación parcial máxima con la variable dependiente parcializada sobre la variable que ya se habla sumado o con la que, si se hubiese sumado, tuviese el valor F máximo. La secuencia de las interacciones se subdividió con criterios predeterminados basados en los valores F y en un nivel de tolerancia.
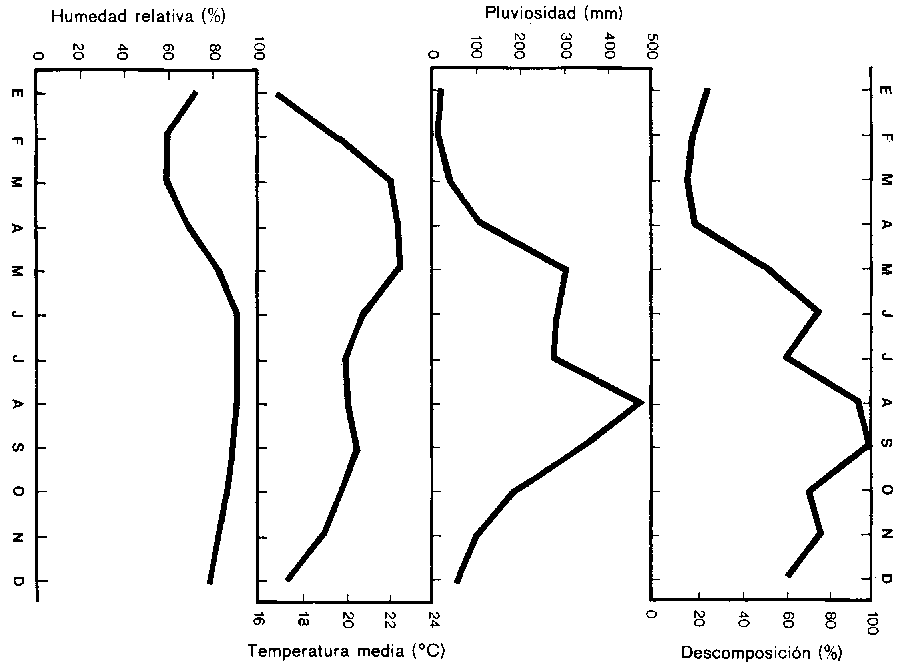
Los datos climáticos originales y derivados y la descomposición mensual de la hojarasca del bosque perenne montano aparecen en el Cuadro 1. Los datos climáticos primarios son: humedad relativa, precipitación, temperatura máxima y temperatura media. Se observó que el porcentaje de descomposición aumenta con la precipitación y con la humedad. Con la temperatura sucede lo contrario. En la figura puede observarse que la descomposición sigue de cerca la intensidad de la precipitación, con un rezago de alrededor de un mes: por ejemplo, la primera cumbre de la descomposición se encuentra en junio y la primera cumbre de la precipitación en mayo. La segunda cumbre de la descomposición y de la precipitación se encuentran en septiembre y en agosto, respectivamente.
Una componente climática importante, derivada de la precipitación y de la temperatura registradas, es la evapotranspiración. «Evapotranspiración efectiva» o cantidad de agua efectivamente evaporada y transpirada es el único término que expresa la energía y la humedad a disposición de los sistemas naturales simultáneamente. Por otra parte, «evapotranspiración potencial» expresa una pérdida teórica de agua si se suministra agua en cantidades ilimitadas. En el trópico, la razón por la cual no se considera la temperatura como elemento climático limitante es el papel que desempeña en la evapotranspiración potencial invariablemente alta. Esta característica de la temperatura hace que la precipitación sea el más crítico y limitante de los elementos climáticos.
Meentemeyer (1974) cita 100 comunicaciones sobre la función de la evapotranspiración en la descomposición de la hojarasca y desarrolla regresiones positivas entre el porcentaje de descomposición y la evapotranspiración efectiva, pero el resultado del presente estudio de la descomposición de la hojarasca en el bosque no indica una relación tan marcada con la evapotranspiración efectiva. El valor r en este estudio es sólo 0,17, en cambio en los anteriores es más de 0,90. Se reconoce por lo general que en la fórmula de Thornthwaite se subestima la evapotranspiración efectiva de los regímenes climáticos tropicales y esto puede haber influido en los resultados. Es posible también que el valor de la capacidad de retención de humedad del suelo empleado en el algoritmo de la computadora (10 pulgadas) sea demasiado bajo para el suelo tropical.
Cuadro 4. Descomposición de la hojarasca del bosque en diferentes lugares durante el primer año.
|
Tipo de bosque o especie arbórea |
Lugar |
Descomposición primer año (%) |
Referencias |
|
Perenne de colina |
Tailandia |
56 |
Investigación de los autores |
|
Plantación de teca |
Tailandia |
95 |
Aksornkoae et al., 1972 |
|
Rodal de teca |
Nigeria |
100 |
Egunjobi, 1974 |
|
Bosque pluvial tropical |
Trinidad |
100 |
Cornforth, 1970 |
|
Bosque húmedo tropical |
Ghana |
100 |
Nye, 1961 |
|
Haya y encina |
Inglaterra |
75 |
Edwards y Heath, 1963 |
|
Vegetación anual |
Nueva Jersey |
58 |
Lang, 1973 |
|
Hierbas y arbustos perennes |
Nueva Jersey |
35 |
Lang, 1973 |
|
Encina, arce rojo, cedro rojo |
Nueva Jersey |
39-48 |
Lang, 1973 |
|
Encina |
Nueva Jersey |
85 |
Lang, 1974 |
|
Arce rojo |
Canadá |
100 |
Weary y Merriam, 1978 |
|
Eucalyptus delegatensis |
Australia |
35-97 |
Wood, 1974 |
Las ecuaciones de regresión múltiple y simple derivadas en este estudio se resumen en el Cuadro 2. Se observa que el porcentaje de descomposición de la hojarasca del bosque perenne montano del norte de Tailandia tiene una correlación significativa con cuatro componentes climáticas: la humedad relativa, la precipitación, la relación de sequedad y el hidrofactor. El coeficiente de correlación de la temperatura máxima (r = - 0,52) es mucho mejor que el de la temperatura media (r = - 0,10). Además, se observa que hay un aumento medio de la hojarasca del 2,29% por un 1% de aumento de la humedad relativa. De manera semejante, la descomposición en el sitio donde se realizó este estudio aumentó en un 0,15% por 1 mm de aumento de la precipitación.
En el Cuadro 3 aparecen los factores climáticos que produjeron el mayor porcentaje de la varianza en la descomposición aislados aplicando la técnica de regresión gradual. La humedad sola representa el 79 %, la evapotranspiración efectiva mensual el 2% y la precipitación el 5% de la varianza total en la descomposición observada. Los valores r2 de ambas regresiones lineales múltiples revelan que los factores multiclimáticos producen una predicción mejor que las variables aisladas. Ni la temperatura media ni la máxima tuvieron una influencia importante en la descomposición de la hojarasca, como habrían permitido esperar los resultados obtenidos por investigaciones anteriores. Por tal motivo es claro que la precipitación juega un papel más importante que la temperatura en la descomposición.
Conviene tener presente que la cantidad de lluvia medida con pluviómetro puede ser menos significativa que el agua del suelo y sobre todo menos significativa que la humedad de la capa superior del suelo y que la humedad de la hojarasca. En las actividades de la fauna, el agua del suelo desempeña una función más directa que la precipitación registrada por el pluviómetro, en parte porque tal función no depende ni de los excedentes de agua ni del escurrimiento de superficie. En el experimento, al tratar de determinar la relación entre el escurrimiento de superficie y la descomposición de la hojarasca, se obtuvo la ecuación:
Y= 30,869 + 2,644X
en la cual Y es el porcentaje de descomposición y X es el escurrimiento expresado en cm (el valor de r es 0,90). Pero esta correlación al parecer es sólo empírica, porque en los meses de escurrimiento máximo (agosto y septiembre) la mayor parte de la desaparición de la hojarasca puede deberse al acarreo más que a la descomposición. Esta es la principal limitación de la técnica de la bandeja para aproximar la pérdida de peso de la hojarasca. Por lo tanto se espera que la humedad del suelo y la humedad de la hojarasca sean más importantes que la precipitación agregada. Estas pueden considerarse como «precipitación útil», o sea como humedad total disponible, después de restar el escurrimiento de superficie inmediato.
Como la descomposición varía con la especie vegetal, el clima y los microorganismos, el tiempo necesario para la mineralización completa es diferente. El Cuadro 4 contiene cifras acerca de la descomposición de los residuos vegetales en varios sitios durante el primer año. La descomposición en los primeros 12 meses (56,23% de la producción de hojarasca) parece ser menor que la obtenido en estudios anteriores en el trópico, pero empleando la ecuación de estado invariable propuesta por Olson (1963):
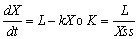
se observa que esta área de la hoya hidrográfica tiene un valor k de 2,28. El parámetro de descomposición k en los bosques tropicales, según Olson, fluctúa entre poco más de 1 y poco más de 4 en un rodal dado. Por lo tanto, el valor k de 2,28 obtenido aquí está dentro de este margen. A esta tasa la mineralización completa tarda 1,8 años. Las diferencias de la duración del ciclo influyen en el desarrollo de los ecosistemas de las regiones silvestres, especialmente en las zonas tropicales donde la hojarasca del bosque influye en los incendios y en la lixiviación, de modo que se requieren métodos apropiados de manejo de la hojarasca.
Al estudiar los efectos de la precipitación, de la temperatura y de algunas componentes modificadas sobre la descomposición de la hojarasca en el bosque perenne montano del norte de Tailandia se observó que el 56,23% de la hojarasca caída anualmente se descompone en 12 meses, con un parámetro de descomposición k de 2,28. La mineralización completa en este ecosistema tarda 1,8 años. El porcentaje de descomposición revela una correlación significativa con la humedad relativa, la precipitación, la relación de sequedad y el hidrofactor. Se observó una relación negativa entre el porcentaje de descomposición y la temperatura máxima.
Una regresión gradual selectiva indica que el 86,1% de la descomposición del primer año depende de la humedad, la precipitación y la evapotranspiración efectiva. Este porcentaje no cambia significativamente añadiendo la temperatura media o máxima como cuarta variable independiente. Además se observó que la precipitación juega un papel mayor que la temperatura en la descomposición de la hojarasca. Hay un aumento medio de la descomposición de la hojarasca de 0,15% por 1 mm de aumento de la precipitación. No conviene dejar de tomar en consideración en investigaciones futuras el agua de la capa superficial del suelo y la humedad de la hojarasca.
Para elaborar los modelos matemáticos de la descomposición de la hojarasca en relación con los factores climáticos quizás se requieran ciertos estudios de laboratorio, pero sólo la investigación en el terreno puede dar un conocimiento práctico del desarrollo de los ecosistemas. Conviene que los experimentos en el terreno abarquen toda la vasta gama de las condiciones climáticas. Es preciso tomar en cuenta el agua de la capa superficial del suelo y la humedad de la hojarasca. Se espera que se realicen pronto estudios detallados sobre la producción y la descomposición de la hojarasca en varios tipos de bosques en toda Tailandia.
AKSORNKOAE, S., KHENMARK, C. y KAEWLAIAD,T. 1972. Study on organic matter in teak plantation. Kasetsart Univ. For Res. Bull., No 23. 36 p. (En tailandés; resumen en inglés.)
BOONYAWAT, S. y NGAMPONGSAI, C. 1974. An analysis of accumulation and decomposition of litter fall in hill evergreen forest, Doi Pui, Chiangmai. Kasetsart Univ. Kogma Watershed Res. Bull., N° 17. 21 p. (En tailandés; resumen en inglés.)
BRINSON, M.M. 1977. Decomposition and nutrient exchange of litter in an alluvial swamp forest. Ecol., 58: 601-609.
BROADFOOT, W.M. y PIERRE, W.H. 1939. Forest soil studies: I. Relation of rate of decomposition of tree leaves to their acid-base balance and other chemical properties Soil Sci., 48: 329-348.
CORNFORTH, I.S. 1970. Leaf-fall in a tropical rain forest. J. Appl. Ecol., 7: 603-608.
CROSSLEY, D.A. y HOGLUND, M.P. 1962. A litter-bag method for the study of microarthropods inhabiting leaf litter. Ecol., 43: 571-574.
DAUBEMIRE,R y PRUSSO, D.C. 1963. Studies of the decomposition rates of tree litter. Ecol., 44: 589-592.
EDWARDS, C.A. y HEATH, G.W. 1963. The role of soil animals in breakdown of leaf material. En Soil organisms. ed. J. Doeksen y J. Van der Drift. Amsterdam, North-Holland Publ. Co., pp. 76-84
EGUNJOBI, J.K. 1974. Litter fall and mineralization in a teak (Tectona grandis) stand. Oikos, 25: 222-226.
GOSZ, J.R., LIKENS, G.E. y BORMANN, F.H. 1973. Nutrient ralease from decomposing leaf and branch litter in the Hubbard Brook Forest, New Hampshire. Ecol. Monogr., 43: 173-191.
HEATH, G.W., ARNOLD, M.K. y EDWARDS, C.A. 1966. Studies in leaf litter breakdown. I. Breakdown rates of leaves of different species. Pedobiol., 6: 1-12.
HEATH, C.W., EDWARDS, C.A. y ARNOLD, M.K. 1964. Some methods for asssessing the activity of soil animals in the breakdown of leaves. Pedobiol., 4: 80-87.
JENNY, H., GESSEL S.P. y BINGHAM, F.T. 1949. Comparative study of decomposition rates of organic matter in temperate and tropical regions. Soil Sci., 68: 419-432.
JOHN, D.H. 1973. Accumulation and decay of litter and net production of forest in tropical West Africa. Oikos, 24: 430-435.
KONONOVA, M.M. 1975. Humus of virgin and cultivated soils. En Soil components Vol. I, ed. J.E. Gieseking. Nueva York, Springer-Verlag. pp. 475-526.
KUCERA, C.L. 1959. Weathering characteristics of deciduous leaf litter. Ecol. 40: 485-487.
LANG, G.E. 1973. Litter accumulation through ecosystem development. Tesis doctoral, Rutgers Univ., New Brunswick, Nueva Jersey. 105 p.
LANG, G.E. 1974. Litter dynamics in a mixed oak forest on the New Jersey Piedmont. Bull. Torrey Bot. Club., 101: 277-286.
LOOMIS, R.M. 1975. Animal changes in forest floor weights under a southeast Missouri oak stand. USFS Res. Note., NC - 184. 3 p.
MADGE, D.S. 1965. Leaf fall and litter disappearance in a tropical forest. Pedobiol., 5: 273-288.
MEENTEMEYER, V. 1974. Climatic water budget approach to forest problems. II. The prediction of regional differences in decomposition rafe of organic debris. Publ. Climatol., 27(1): 35-74.
MEENTEMEYER,V. 1978. Macroclimate and lignin control of litter decomposition rates. Ecol., 59: 465-472.
MIKOLA, P. 1960. Comparative experiment on decomposition rates of forest litter in southern and northern Finland. Oikos, II:161 - 166.
NYE, P.H. 1961. Organic matter and nutrient cycles under moist tropical forest. Plant & Soil, 13: 333-346.
OLSON, J.S. 1963. Energy storage and the balance of producers and decomposers in ecological systems. Ecol., 44:322-331.
OVINGTON, J.D. 1954. Studies of development of woodland conditions under different trees. II. The forest floor. J. Ecol., 42: 71-80.
PAOVONGSAR, S. 1976. Litter fall and mineral nutrient content of litter in dry dipterocarps forest. Tesis de licenciatura Kasetsart Univ. 75 p. (En tailandés; resumen en inglés.)
PRADHAN, I.P. 1973. Preliminary study of interception through leaf litter. Indian For., 99: 440-445.
REINERS, W.A. 1968. Carbon dioxide evolution from the floor of three Minnesota forests. Ecol., 49: 471-483.
ROCHOW, J.J. 1974. Litter fall relations in a Missouri forest. Oikos, 25: 80-85.
SHANKS, R.E. y OLSON, J.S. 1961. First year breakdown of leaf litter in southern Appalachain forests. Sci., 134: 194-195.
SINGH, K.P. 1969. Studies in decomposition of leaf litter of important trees of tropical deciduous forest at Varanasi. Trop. Ecol., 20: 292-311.
SPURR, S.H. y BARNES, B.V. 1973. Forest ecology. Nueva York. Ronald Press, Co. 571 p.
STONE, JR., L.A. 1971. Water balance computer program using consecutive monthly data. En Computer program for the climatic water balance, ed. L.A. Stone, Jr., J.C. Albrecht y G.A. Yoshioka. Publ. Climatol., 24(3): 1-10.
THAIUTSA, B., SUWANNAPINUNT, W. y KAITPRANEET, W. 1978. Production and chemical composition of forest litter in Thailand. Kasetsart Univ. For. Res. Bull., No 52. 32 p.
THORNTHWAITE, C.W. y MATHER, J.R. 1957. Instructions and tables for computing potential evapotranspiration and the water balance. Publ. Climatol., 10(3): 185-311.
VAN DER DRIFT, J. 1963. The disappearance of litter in mull and mor in connection with weather conditions and the activity of the macrofauna. En Soil organisms, ed. J. Doeksen y J. Van der Drift. Amsterdam, North-Holland Publ. Co. pp. 124-133
WAKSMAN, S.A. y GERRETSEN, F.C. 1931. Influence of temperatura and moisture upon the natura and extent of decomposition of plant residues by microorganisms. Ecol., 12: 33-60.
WEARY, G.C. y MERRIAM, H.G. 1978. Litter decomposition in a red maple woodlot under natural conditions and under insecticide treatment. Ecol., 59: 180-184.
WIANT, JR., H.V. 1967a. Influence of temperature on the rate of soil respiration. J. For., 65: 489490.
WIANT, JR., H.V. 1967b. Influence of moisture content on soil respiration. J. For., 63: 902-903.
WILLIAMS, S.T. y GRAY, T.R.G. 1974. Decomposition of litter on soil surface. En Biology of litter decomposition. Vol. II, C.H. Dickinson y G.J.E. Pugh, ed. Nueva York, Academic Press. pp. 611-630.
WITKAMP, M. 1966. Decomposition of litter in relation to environmental factors. Plant & Soil, 15: 295-311.
WOOD, T.G. 1974. Field investigations on the decomposition of leaves of Eucalyptus
delegatensis in relation to environmental factors. Pedobiol., 14:
343-371.
FAO Departamento de Montes
La silvicultura para los pueblos
la contribución forestal a las comunidades rurales

ORGANIZACIÓN DE LAS NACIONES UNIDAS PARA LA AGRICULTURA Y LA ALIMENTACIÓN
publicaciones que usted necesita

Pedidos a:
Sección de Distribución y Ventas
Organización de las Naciones Unidas para la Agricultura y la Alimentación
00100 Roma, Italia
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}