![]()
![]()
![]()
7.1 Sélectivité du chalut
7.2 Recrutement
7.3 Limites de taille
7.4 Sélectivité des filets maillants
7.5 Exercices
Les poissons de tailles ou d'âges divers ne subissent pas tous la même mortalité par pêche: les petits peuvent s'échapper à travers les mailles d'un filet, ou ne pas se trouver dans la principale zone exploitée. Il est utile de distinguer l'élément provenant uniquement du comportement (au sens le plus large) des animaux mêmes - recrutement - et celui qui est dû à la sélectivité de l'engin de pêche.
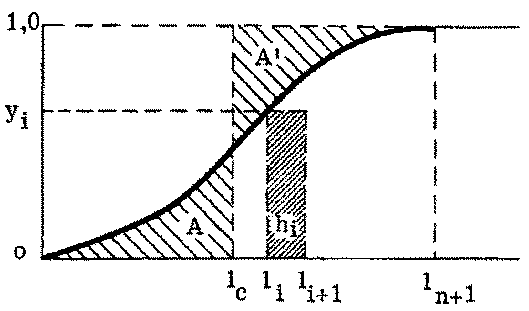
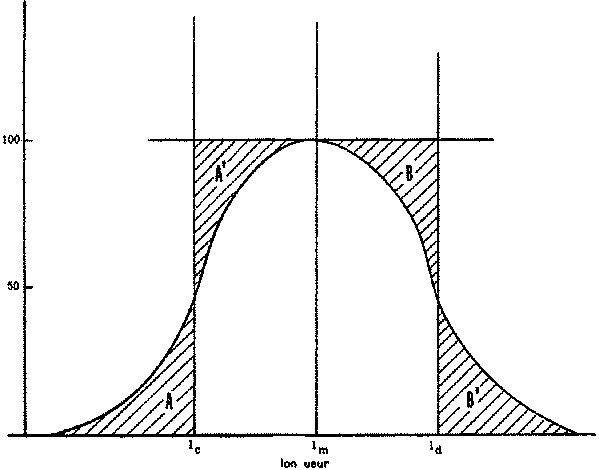
La forme la plus simple de sélection s'effectue dans les engins à poche (chaluts, sennes, etc.). Pour cette catégorie de matériel de pêche, on suppose communément que la composition par taille est la même pour les poissons entrant dans le filet et pour ceux qui se trouvent à proximité immédiate de l'engin. La sélectivité de celui-ci est donc déterminée par la fuite, à travers les mailles, des poissons qui y étaient entrés. Pour beaucoup d'espèces, divers éléments permettent d'établir que la fuite a lieu surtout à travers la poche terminale. Pour pouvoir mesurer directement la sélectivité, il faudra donc être en mesure d'estimer le nombre de poissons de chaque taille pénétrant dans le filet, soit en doublant extérieurement le cul-de-chalut ou une autre partie de l'engin d'une nappe à mailles plus fines, soit en examinant la composition par taille des prises de filets à mailles beaucoup plus petites utilisés en même temps et au même endroit. Quelle que soit la méthode employée, les résultats peuvent être exprimés en proportions de poissons de chaque longueur entrant dans le filet et retenus dans la poche terminale. Ces proportions, portées en fonction de la longueur des poissons, donnent la courbe de sélectivité du filet pour l'espèce considérée. Un exemple typique est fourni par la figure 7.1.
La courbe de sélectivité peut couvrir une gamme de longueurs de 10 centimètres ou davantage, ce qui signifie que les jeunes poissons commençant à croître dans l'intervalle de sélection ne subissent au début qu'une faible mortalité par pêche. Lorsqu'ils deviennent plus grands, leurs chances d'échapper au filet (après y être entrés) deviennent moindres, jusqu'au moment où ils sont trop gros pour pouvoir fuir: c'est seulement alors qu'ils sont exposés au taux de mortalité entier, quel qu'il soit. Il est possible d'introduire dans les évaluations du rendement ce changement progressif du coefficient de mortalité par pêche dans l'intervalle de sélection, mais il suffit généralement de représenter le processus par une seule longueur moyenne de sélection, lc, et de supposer que tous les poissons de longueur inférieure à lc s'enfuient, tandis que tous ceux qui dépassent lc sont retenus par l'engin, subissant donc la pleine mortalité par pêche.
FIGURE 7.1. - Courbe simple de sélectivité. Détermination de la longueur moyenne de sélection.
La quantité importante est donc:
tc = âge moyen de sélection
= âge moyen d'entrée dans la prise, ou âge moyen à la première capture.
Dans la figure 7.1, la longueur moyenne de sélection lc est la longueur pour laquelle les deux surfaces hachurées A et A' sont égales. Si la courbe de sélectivité est symétrique ou presque, ce sera la longueur au point médian de la courbe, c'est-à-dire la longueur correspondant au pourcentage de captures de 50 pour cent, l'autre moitié des poissons entrés dans le filet s'échappant. Si elle n'est pas symétrique, lc peut être calculé en égalant les aires entre la courbe de sélectivité et l'axe des y et entre la ligne l = lc et l'axe des y. Supposons que l'on classe les longueurs et prenons la ième catégorie de taille hi, limitée par les longueurs li, et li +1: l'ordonnée correspondant à la ième catégorie est yi où i = 0, 1, 2,... n. Dans ce cas, l'aire à gauche de la courbe de sélectivité est égale à l'aire du rectangle de hauteur 1 et de base ln+1 moins l'aire située au-dessous de la courbe de sélectivité. La surface de celle-ci est approximativement égale à la somme des surfaces des rectangles pour chacune des catégories de longueur, soit S hi × y1. L'aire totale située à gauche de la courbe de sélectivité est donc ln+i - S hi,yi, qui, comme la hauteur totale de la courbe est 1, sera égal à lc. Si hi, est constant et égal à 1 cm, on aura: lc =ln+1 - S yi..
Pour les chaluts, la longueur moyenne de sélection lc est généralement proportionnelle au maillage m:
lc = b.m
où b est le coefficient de sélectivité.
Cette formule permet d'estimer le coefficient moyen de sélectivité pour toute dimension de maille à partir des résultats d'expériences menées avec un petit nombre de maillages différents. De plus, le coefficient de sélectivité varie peu pour les poissons d'à peu près la même forme. Une certaine variation du coefficient est introduite par l'état du poisson, ainsi que par le type de maille et le matériau employé pour sa construction (les matériaux légers et souples - par exemple le nylon - donnent un coefficient de sélectivité plus élevé que les matériaux épais - comme le chanvre).
En sus de la longueur moyenne de sélection (point des 50 pour cent), les courbes de sélectivité diffèrent aussi par leur pente - qui indique si la sélection s'exerce sur une gamme de tailles étroite ou étendue. Elle est généralement mesurée par l'éventail de sélection - différence entre les longueurs auxquelles 25 pour cent et 75 pour cent des poissons sont retenus par l'engin.
Le recrutement est le processus par lequel les jeunes poissons entrent dans l'aire exploitée et peuvent être capturés par les engins de pêche. Il peut s'agir d'un mouvement effectif, comme dans le cas de la plie de la mer du Nord, qui, à un âge relativement avancé (de 3 à 4 ans), quitte les zones côtières, peu profondes, de développement des jeunes, pour gagner les principaux terrains de pêche. Le recrutement peut aussi ne faire intervenir qu'une modification d'habitudes: par exemple, chez l'églefin de la mer du Nord, on trouve dans les mêmes secteurs des individus de tous âges - les très jeunes sont pélagiques, et entrent dans la phase exploitée lorsqu'ils adoptent un habitat démersal, à l'automne de leur première année d'existence. Du point de vue mathématique, la quantité importante est tr (âge moyen au recrutement).
Le recrutement est, de par sa nature, bien moins facile à exprimer quantitativement que la sélection due au maillage. Comme on s'intéresse surtout à l'effet combiné du recrutement et de la sélection - c'est-à-dire au mode d'«entrée dans les prises» - l'allure du recrutement est très importante quand celui-ci se situe au-dessus de la gamme de sélection de l'engin ou la chevauche, mais non quand il est achevé avant que commence la sélection opérée par l'engin. Par conséquent, si tous les poissons ont été recrutés à une taille tombant au-dessous de l'éventail de sélection d'un maillage probable donné, on n'a pas à tenir compte du schéma précis du recrutement, qui peut être pris arbitrairement comme survenant à n'importe quel âge ou longueur commodes, au-dessous de la gamme de sélection (voir figure 7.1). Lorsque ce facteur est important, on peut déterminer l'allure générale de la courbe de recrutement par une bonne connaissance de la biologie de l'espèce, et peut-être l'estimer avec une précision plus grande au moyen d'enquêtes faites par des navires de recherche sur les jeunes poissons, tant sur les principaux fonds de pêche qu'en dehors de ceux-ci. A défaut, on obtiendra une estimation en comparant la composition par taille des captures effectives des navires de pêche commerciale avec la sélectivité connue des engins employés. Le recrutement intéresse normalement toute une gamme de tailles, et peut être représenté par le même type de courbe que la courbe de sélectivité de la figure 7.1.
Si sélection et recrutement portent sur la même gamme de tailles (comme dans la figure 7.2), la sélection effective, c'est-à-dire la proportion Pl de la pleine mortalité par pêche à laquelle sont exposés les poissons du stock d'une taille donnée, sera fournie par l'équation:
Pl =rl × Sl, (7.1)
où rl = proportion recrutée, c'est-à-dire ordonnée de la courbe de recrutement
et Sl = proportion des poissons entrant dans le filet qui sont retenus, c'est-à-dire ordonnée de la courbe de sélectivité.
FIGURE 7.2. - Combinaison d'une courbe de sélectivité et du recrutement. Recrutement sur une série de longueurs.

Ainsi, la courbe de sélectivité résultante, qui exprime l'entrée effective des poissons dans les captures, est le produit des courbes de recrutement et de sélectivité de l'engin, et à partir d'elle la longueur moyenne de sélection résultante peut être obtenue comme plus haut, en égalant les deux surfaces A et A'.
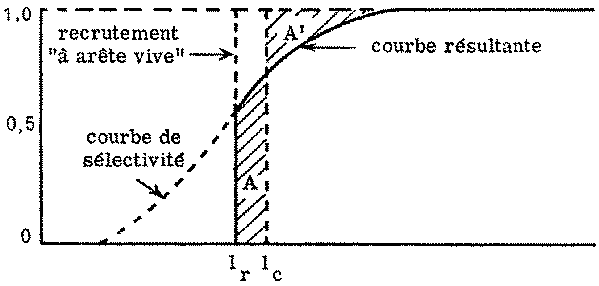
Parfois - par exemple lorsque les recrues pénètrent dans une zone de pêche - le recrutement, exprimé en longueur, peut être tout à fait prononcé, approchant dans les cas extrêmes, à la longueur seuil lr, ce qu'on appelle sélection «à arête vive» (voir figure 7.3). Les poissons de longueur inférieure à lr, ne subissent aucune mortalité par pêche, puisqu'ils ne sont pas capturés par l'engin, mais lorsqu'ils atteignent lr, ils sont brusquement exposés à une forte mortalité par pêche (comme l'indique la figure 7.3: plus de 50 pour cent de la pleine mortalité par pêche). Dans ce cas, la courbe de sélection résultante part de la longueur/, et s'élève verticalement jusqu'à l'intersection avec la courbe de sélectivité de l'engin; ensuite, elle coïncide avec celle-ci. Ici aussi, cependant, la longueur moyenne de sélection lc peut être calculée à partir de cette courbe résultante: lc est telle que les deux aires A et A' de la figure 7.3 soient égales.
FIGURE 7.3. - Recrutement «à arête vive».
Si une taille légale minimale, soit ls, tombe à l'intérieur ou au-dessus de l'éventail de sélection de l'engin employé, l'entrée des poissons dans les apports peut être représentée par une courbe partant de ls et s'élevant verticalement jusqu'à atteindre la courbe de sélectivité réelle (voir figure 7.4). La longueur moyenne de sélection lc est calculée de manière analogue: elle est telle qu'elle égalise les aires A et A' de la figure 7.4.
Les poissons n'atteignant pas la taille légale sont pris aussi, et, puisqu'ils ne peuvent pas être débarqués, doivent être rejetés à la mer. L'effet de cette capture et de ce rejet sur la production du stock dépend du taux de survie des poissons remis à l'eau. Si tous survivent, les choses se passent comme si la courbe de recrutement du point de vue des apports était la courbe de sélectivité réelle.
FIGURE 7.4. - Effet de la taille limite (ls) sur la courbe de sélectivité effective.

Si, par contre, certains des poissons trop petits rejetés, ou la totalité d'entre eux, meurent, chaque classe d'âge est sujette à une certaine mortalité par pêche avant d'atteindre la longueur ls, comme l'indique la partie inférieure de la courbe de sélectivité de la figure 7.4. Le coefficient résultant de mortalité par pêche, qui tient compte de la capture et du rejet des poissons trop petits, est obtenu en calculant la moyenne de la partie de la courbe de sélectivité qui se trouve au-dessous de la taille limite. Cette longueur moyenne, baptisée lp, est telle qu'elle égalise les aires B et B' de la figure 7.4. En d'autres termes, on suppose que les poissons ne sont pas capturés du tout jusqu'au moment où ils atteignent la longueur lp, où ils sont alors soumis à la fraction constante a du plein taux de mortalité par pêche, jusqu'à ce qu'ils parviennent à la taille ls (a est l'ordonnée à la longueur ls, - voir figure 7.4). Si la croissance de lp à ls occupe une période ts - tp (voir ci-dessous) et si la fraction q de poissons rejetés meurt, alors la classe d'âge est réduite du facteur
 , (7.2)
, (7.2)
du fait de la capture et du rejet des poissons n'ayant pas encore atteint la taille marchande. Dans cette expression, F est le coefficient total de mortalité par pêche auquel les poissons sont exposés quand leur taille a dépassé la zone de sélection de l'engin.
La sélectivité des filets maillants est telle que la proportion de poissons retenus est maximale pour une certaine taille, et diminue pour tout poisson de taille supérieure ou inférieure à cette taille optimale. S'il est possible d'établir directement la répartition par taille du poisson de la zone exploitée en employant des engins pratiquement non sélectifs ou dont on connaît la sélectivité - par exemple un chalut - on peut déterminer immédiatement la sélectivité du filet maillant. Si l'on pose GCl et TCl pour les nombres de poissons d'une taille l capturés par unité de temps de pêche respectivement par le filet maillant et par l'engin supposé non sélectif, la mortalité par pêche exercée par le filet maillant sur les poissons de longueur l sera proportionnelle à:
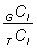
Lorsqu'il n'est pas facile d'avoir recours à un engin de pêche non sélectif, la meilleure manière d'estimer les caractéristiques de sélectivité consiste à employer un jeu de filets maillants de maillages différents. Pour toute taille de poisson, la répartition des prises obtenues avec différentes dimensions de maille sera analogue à la répartition des captures des poissons de diverses tailles par un maillage donné, avec un maximum pour une maille optimale, la prise tombant de part et d'autre de celui-ci. A partir des données des captures effectuées par divers filets (corrigées le cas échéant pour tenir compte des différences dans le temps total de pêche des divers maillages), il est possible d'obtenir ces distributions pour chaque taille de poisson. Ces éléments peuvent être combinés en exprimant les deux variables par des termes normalisés: la dimension de maille par le rapport du périmètre de la maille à la grosseur du corps du poisson d'une longueur donnée, et la prise par le rapport entre le nombre de poissons capturés avec le maillage utilisé et le nombre capturé avec le maillage le plus efficace (voir McCombie et Fry, 1960; Gulland et Harding, 1961.
Si l'on suppose que chaque maille possède la même puissance de pêche optimale, autrement dit détermine la même mortalité par pêche sur la taille de poissons pour laquelle elle est la plus efficace, alors les données combinées peuvent fournir immédiatement, non seulement l'efficacité relative de divers filets sur une taille donnée de poissons, mais aussi, pour un filet d'un maillage donné, l'efficacité relative sur différentes tailles de poissons, autrement dit sa sélectivité.
La sélectivité peut également être estimée en comparant les prises de seulement deux maillages, sous réserve que l'on ait fait une hypothèse touchant l'allure de la courbe de sélectivité. Celle-ci est souvent proche de la courbe normale, et si on la suppose normale on peut écrire:
Cl = prise de poissons de longueur l par unité de temps de pêche
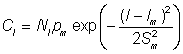
où Nl = nombre de poissons de longueur l susceptibles d'être capturés par l'engin.
Pm = puissance de pêche du filet de maille m
lm = longueur des poissons pour laquelle la maille m est la plus efficace
Sm = écart type de la courbe de sélectivité de la maille.
Si l'on utilise sur la même population deux mailles, A et B, il est évident que l'on a:
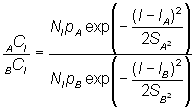
ou, en prenant les logarithmes népériens:

et, si l'on suppose en outre que l'écart type de la courbe de sélectivité est constant (autrement dit que SA = SB = S), l'équation peut être simplifiée:
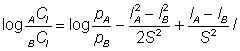
L'expression de droite est une fonction linéaire de l: elle peut s'exprimer sous la forme type a + bl:
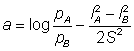
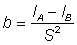
Ainsi, si le logarithme du rapport des prises par unité de temps de pêche de filets de différentes tailles est exprimé en fonction de la longueur, on obtient une droite, dont on peut estimer:

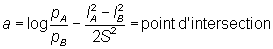
A partir de ces éléments, il n'est pas possible d'estimer séparément lA, lB, et S2, ni par conséquent de déterminer les courbes de sélectivité sans hypothèses supplémentaires. Toutefois, il est raisonnable de supposer (comme on l'a fait précédemment pour une gamme de maillages) que les puissances de pêche des deux filets sont égales, et en outre que la longueur optimale est proportionnelle à la dimension de maille (équivalant communément dans la pratique à un poisson dont la grosseur du corps - la «corpulence» - est légèrement supérieure à environ 1,1 fois le périmètre de la maille). Autrement dit:
i.e. lA = KMA
lB =KMB
On a alors: 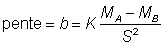

Expression à partir de laquelle on obtient:
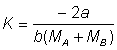
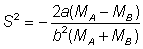
d'où peuvent être déterminées les courbes de sélectivité.
D'autres hypothèses peuvent être faites touchant l'allure de la courbe de sélectivité. On peut notamment supposer, comme on l'a fait plus haut, que la courbe est normale, mais que l'écart type est proportionnel au maillage. Ce modèle cadre fréquemment mieux avec les données observées que celui qui suppose un écart type constant (voir Regier et Robson, 1966, pour une étude complète de la sélectivité des filets maillants).
Lorsqu'on a déterminé la courbe de sélectivité du filet maillant, l'étude ultérieure des populations est facilitée s'il est possible de représenter la courbe par un trait simplifié, analogue à l'approximation «à arête vive» employée pour la sélectivité du chalut. Il faut deux longueurs de sélection: lc, longueur à laquelle le poisson entre dans la zone de sélection, et ld, longueur à laquelle il en sort. Autrement dit, la pêche à l'aide d'un filet maillant présentant la courbe de sélectivité de la figure 7.5 exerce le même effet qu'une mortalité constante subie entre les longueurs lc, et ld, pour lesquelles les paires de zones hachurées A et A' et B et B' sont égales.
FIGURE 7.5. - Conversion, aux fins de l'étude des populations, de la courbe de sélectivité d'un filet maillant en une courbe simplifiée par l'emploi d'approximation faisant intervenir la sélection «à arête vive».
1. a) Des culs-de-chalut de 44 et de 112 mm de maille, utilisés alternativement, ont donné les prises de plies (temps de pêche égaux) indiquées au tableau 1.
TABLEAU 1. - PLIE
|
Longueur |
Maille de 44 mm |
Maille de 112 mm |
|
cm |
||
|
10 |
0 |
0 |
|
11 |
1 |
0 |
|
12 |
2 |
0 |
|
13 |
6 |
0 |
|
14 |
16 |
0 |
|
15 |
18 |
0 |
|
16 |
26 |
0 |
|
17 |
64 |
3 |
|
18 |
121 |
4 |
|
19 |
182 |
10 |
|
20 |
247 |
13 |
|
21 |
292 |
16 |
|
22 |
344 |
37 |
|
23 |
367 |
60 |
|
24 |
355 |
132 |
|
25 |
276 |
190 |
|
26 |
225 |
177 |
|
27 |
147 |
136 |
|
28 |
90 |
97 |
|
29 |
57 |
54 |
|
30 |
26 |
33 |
|
31 |
24 |
26 |
|
32 |
18 |
13 |
|
33 |
10 |
9 |
|
34 |
12 |
7 |
|
35 |
5 |
6 |
|
36 |
10 |
5 |
|
37 |
6 |
2 |
|
38 |
3 |
4 |
|
39 |
2 |
2 |
|
40 |
2 |
3 |
|
41 |
0 |
1 |
|
42 |
0 |
2 |
|
43 |
0 |
2 |
|
44 |
0 |
1 |
|
45 |
0 |
1 |
|
46 |
0 |
1 |
|
47 |
0 |
0 |
|
48 |
0 |
0 |
TABLEAU 2. - MERLAN
|
Longueur |
Cul-de-chalut |
Poche extérieure |
|
cm |
||
|
10 |
0 |
0 |
|
11 |
0 |
0 |
|
12 |
0 |
3 |
|
13 |
0 |
7 |
|
14 |
0 |
21 |
|
15 |
0 |
27 |
|
16 |
0 |
23 |
|
17 |
0 |
19 |
|
18 |
2 |
20 |
|
19 |
3 |
83 |
|
20 |
23 |
219 |
|
21 |
68 |
302 |
|
22 |
89 |
256 |
|
23 |
79 |
154 |
|
24 |
115 |
55 |
|
25 |
106 |
42 |
|
26 |
87 |
18 |
|
27 |
63 |
7 |
|
28 |
55 |
0 |
|
29 |
38 |
2 |
|
30 |
34 |
0 |
|
31 |
11 |
0 |
|
32 |
11 |
0 |
|
33 |
6 |
0 |
|
34 |
9 |
0 |
|
35 |
5 |
0 |
|
36 |
2 |
0 |
|
37 |
1 |
0 |
|
38 |
1 |
0 |
|
39 |
0 |
0 |
b) Un cul-de-chalut de 74 mm doublé extérieurement d'une poche à petites mailles a capturé des merlans dont la répartition entre le cul et la poche est indiquée au tableau 2.
Tracer d'après ces données, les courbes de sélectivité pour la plie et le merlan et déterminer les longueurs moyennes de sélection lc. Noter que le nombre de poissons pénétrant dans le filet est pour a) (traits alternés) égal à la prise dans le filet à petites mailles, tandis que pour b) il est constitué par le total des poissons retenus dans le cul-de-chalut et de ceux qui sont sortis de celui-ci et se trouvent pris dans la poche extérieure. Calculer les facteurs de sélection.
2. Courbe approximative de recrutement pour la, pêcherie britannique de plies de la mer du Nord:
|
Longueur. (cm) |
20 |
21 |
22 |
23 |
24 |
25 |
26 |
27 |
28 |
29 |
30 |
31 |
32 |
33 |
34 |
|
Fraction recrutée |
0,02 |
0,05 |
0,08 |
0,16 |
0,27 |
0,38 |
0,48 |
0,62 |
0,75 |
0,86 |
0,94 |
0,97 |
0,99 |
1,00 |
1,00 |
Calculer la courbe de sélectivité résultante pour une maille de 112 mm (en utilisant les résultats de l'exercice 1), et, à partir; de là, la valeur de lc.
Si, au lieu de la courbe de recrutement ci-dessus, on constatait que le recrutement est effectivement «à arête vive» à 26 cm, quelle serait la valeur de lc avec une maille de 112 mm?
3. Convertir l'estimation ci-dessus de lc pour la plie et le merlan en âge (tc) en utilisant l'équation de von Bertalanffy et les valeurs suivantes des paramètres:
|
Plie |
K= 0,095 |
Merlan |
K= 0,32 |
|
|
L¥ = 68 cm |
|
L¥ = 45,5 cm |
|
|
t0 = - 0,8 année |
|
t0 = - 0,4 année |
4. Pourcentage des répartitions de longueur des cabillauds capturés aux filets maillants et aux sennes tournantes à Lofoten en 1952 (source des données: Rollefsen, 1952):
|
Longueur (cm) |
65- |
70- |
75- |
80- |
85- |
90- |
95- |
100- |
105- |
110- |
115- |
120+ |
|
Sennes tournantes |
2 |
4 |
8 |
13 |
20 |
28 |
35 |
34 |
25 |
15 |
8 |
5 |
|
Filets maillants.. |
0 |
2 |
5 |
15 |
30 |
50 |
45 |
27 |
15 |
5 |
2 |
1 |
En supposant que les sennes tournantes n'exercent pas de sélection, déterminer l'allure de la courbe de sélectivité des filets maillants. Quelle est la taille de poissons que ceux-ci capturent le mieux?
5. On a utilisé pour la pêche du saumon dans le Frazer, en 1947 et 1948, neuf maillages différents de filets maillants. Le tableau suivant donne le nombre de poissons de diverses longueurs capturés par chaque filet (avec corrections pour différents temps de pêche) (d'après Peterson, 1954).
|
Longueur à la fourche (cm) |
Maillage |
||||||||
|
cm |
|||||||||
|
13,5 |
14.0 |
14,8 |
15,4 |
15.9 |
16,6 |
17.8 |
19,0 |
20,6 |
|
|
50-51 |
6 |
4 |
|
|
|
|
|
|
|
|
52-53 |
52 |
11 |
1 |
1 |
- |
- |
- |
- |
- |
|
54-55 |
102 |
91 |
16 |
4 |
4 |
2 |
- |
2 |
2 |
|
56-57 |
295 |
232 |
131 |
61 |
17 |
13 |
3 |
1 |
2 |
|
58-59 |
309 |
318 |
362 |
243 |
95 |
26 |
4 |
2 |
1 |
|
60-61 |
118 |
173 |
326 |
342 |
199 |
100 |
10 |
8 |
1 |
|
62-63 |
79 |
87 |
191 |
239 |
202 |
201 |
39 |
10 |
6 |
|
64-65 |
27 |
48 |
111 |
143 |
133 |
185 |
72 |
16 |
3 |
|
66-67 |
14 |
17 |
44 |
51 |
52 |
122 |
74 |
27 |
3 |
|
68-69 |
8 |
6 |
14 |
23 |
25 |
59 |
65 |
51 |
1 |
|
70-71 |
7 |
3 |
8 |
14 |
15 |
16 |
34 |
22 |
2 |
|
72-73 |
|
3 |
1 |
2 |
5 |
4 |
6 |
10 |
1 |
|
74-75 |
|
|
|
|
|
|
|
1 |
|
a) Méthode graphique. En portant séparément, pour chaque catégorie de taille, le nombre des poissons péchés en fonction du maillage, estimer le nombre de poissons de chaque taille qui serait capturé par la maille la plus efficace pour cette taille. Exprimer ensuite la prise de chaque maille en pourcentage de la capture potentielle par la maille la plus efficace, autrement dit la sélectivité de cette maille pour cette taille de poissons.
Combiner ces courbes de sélectivité pour tous les maillages en exprimant la dimension de maille comme proportion de la maille la plus efficace pour la catégorie de longueurs en question. En tirer une courbe de sélectivité combinée unique des filets maillants utilisés pour la pêche du saumon.
b) Méthode analytique. Calculer le logarithme du rapport des captures de poissons d'une taille donnée effectuées par des paires de filets de maillages voisins. En supposant que pour chaque maille la courbe de sélectivité est normale et d'une variance constante, et que la longueur moyenne de sélection de chaque maille est proportionnelle au maillage, calculer la courbe de sélectivité de chaque maille.
![]()
![]()
![]()