![]()
![]()
![]()
3.1 Growth by length and weight
3.2 Conversion of length to age
3.3 Exercises
For population analysis it is desirable to express the growth of fish in a mathematical expression. The basic requirement is an expression which will give the size (in terms of length or weight) at any given age which agrees with the observed data of size at age, and which is in a mathematical form which can be incorporated reasonably easily in expressions for yield. Strictly, most population analysis is concerned more directly with growth rate, i.e. increase in weight or length per unit time, rather than with the size at various ages, because many problems in fishery assessment are essentially a matter of comparing weight gained by growth against that lost by natural mortality. Sometimes, for instance when considering the effect of an increase in size of first capture, it is particularly important to know the rate of growth over a comparatively short part of the total life span - that is, to know how long it will take a fish to grow from the original size of first capture to the new. There are therefore good reasons for preferring, other things being equal, a method of fitting equations to growth rate data, rather than simply size at age.
Other desirable features of a growth equation are that the computational work involved in fitting it to observed data should be small, that the number of constants used should be few, that as far as possible these constants should have some biological meaning, and that if extrapolated to ages beyond those used in fitting the equation, the equation should not lead to unreasonable results.
There exists a considerable and growing literature on growth equations, which covers a wide range of possible equations, none of which seems to be entirely satisfactory in all situations. In fact it is most unlikely that a simple formula would always be able to describe the growth of even a single fish through most of its life, in which there could be greatly different conditions of food supply, reproductive strain, etc. These notes do not attempt to give a complete survey of growth equations, but will be mainly concerned with one particular equation, that ascribed to von Bertalanffy (1938), which satisfies the two most important criteria - it fits most of the observed data of fish growth, and can be incorporated readily into stock assessment models.
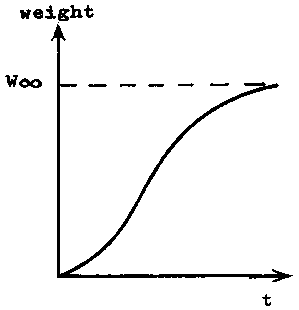
If the length of a fish, crustacean, or lamellibranch is plotted against age, the result is usually a curve of which the slope continuously de- creases with increasing age, and which approaches an upper asymptote parallel to the X-axis (see Figure 3.1.1). Curves of weight at age also approach an upper asymptote, but usually form an asymmetrical sigmoid, the inflexion occurring at a weight of about one third of the asymptotic weight (see Figure 3.1.2).
FIGURE 3.1.1. - Curve of growth by length.
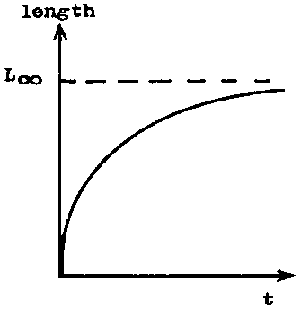
FIGURE 3.1.2. - Curve of growth by weight.
If the rate of growth in length is plotted against length, the result is often well fitted by a straight line, cutting the X-axis at a point L¥ beyond which the fish will not grow. This is of course the asymptote of the plot of length on age. If the rate of growth in length is linearly related to length, then in mathematical terms
................... (3.1)
where L¥ is the value of l for which the rate of growth is zero. This equation is in the general form of a straight line relation, y = ax + b, with a = - K, b = K L¥ . To integrate this differential equation, we write it in the form
(see section 2)
and therefore - log (L¥ - l) = Kt + constant
or L¥ - l = Kt + constantor l = L¥ - constant × e-Kt
Putting t0 as the theoretical age at which l = 0 we can obtain the value of the constant from the relation
then
................. (3.2)
Equation (3.1) is identical with that deduced by von Bertalanffy (1938) on physiological grounds. He considered that growth in weight was the resultant of the difference between anabolic and katabolic factors, taken as proportional to surface area and weight respectively
or, putting w µ l3, s µ l2
In fitting the growth curve the instantaneous rate of growth will not be known, but only lengths at certain times - often one year apart when they are derived from age determination, but at irregular intervals if derived from tagging data. If the times t1, t2 are close together, a close approximation to the instantaneous rate of growth is given by 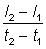 , where l1, l2 are the lengths at times t1, t2 respectively. If this growth rate is plotted against the mean length
, where l1, l2 are the lengths at times t1, t2 respectively. If this growth rate is plotted against the mean length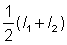 , then a plot corresponding to equation (3.1) is obtained; the intercept on the X-axis gives an estimate of L¥ , and the slope an estimate of - K.
, then a plot corresponding to equation (3.1) is obtained; the intercept on the X-axis gives an estimate of L¥ , and the slope an estimate of - K.
For data at equal time intervals of length T, from equation (3.2)
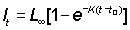
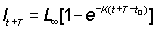
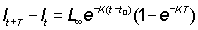 lt+T - lT = (L¥ - lt)(1 - e-KT) ............. (3.3)
lt+T - lT = (L¥ - lt)(1 - e-KT) ............. (3.3)
The plot of increment lt+T - lT, against initial length, lt, therefore gives a line, slope - (1 - e-KT), and an intercept, on the X-axis, of L¥ (see Figure 3.2.1). The important special curve is T = 1 year, when the slope = - (1 - e-K). An alternative form of equation (3.3) is
lt+T - lT = L¥ (1 - e-KT) + lte-KT ..............(3.4)
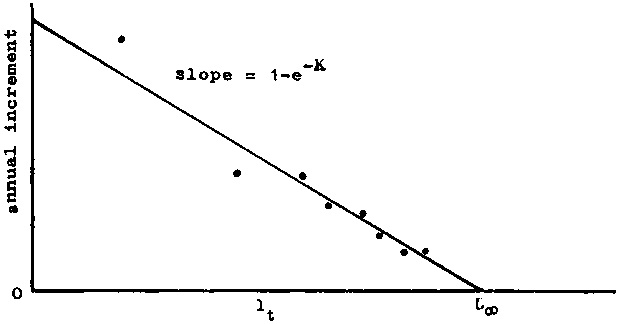
which, for T = 1, is the well-known Ford-Walford plot, of lt+i against lt, which gives a straight line, slope e-K, and an intercept on the 45° line, where lt = lt+1, of L¥ (see Figure 3.2.2). This plot is essentially the same as the plot of increment (lt+1 - lt) against initial length. The points will seem to fit the Ford-Walford line better, but because the intersection of the regression line and the 45° line is very oblique, L¥ is in fact estimated with equal precision by the two plots, and if the fitting is done graphically rather than by calculating regression lines, e.g. by least squares, greater errors in drawing are likely to be introduced in the plot of lt+1 against lt.
FIGURE 3.2.1. - Estimation of growth constants. Plot of length increment against initial length.
FIGURE 3.2.2. - Plot of length against length one year earlier (Ford-Walford plot).
L¥ and K can be determined directly from the lines fitted to these plots either by regression analysis, or by eye; then t0 can be estimated from equation (3.2) for any particular observation of length at age. For this, equation (3.2) is best rewritten as
or
............... (3.5)
While an estimate of t0 can be thus obtained for each age for which the mean length is known, these estimates will not be equally good. Those from old fish will be highly variable, because a small difference in lt makes a big difference to the estimate of t0 when lt is nearly equal to L¥ , while the mean length of the youngest fish may be biased because only the bigger fish of these ages appear in the catches. The best estimate of t0 would appear to be the mean of the estimates of t0 obtained from the younger, but fully recruited, age groups.
Alternatively, only L¥ is estimated from the plots of increment against initial length. From this 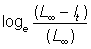 can be calculated, and plotted against t. From equation (3.5) this should give a straight line, slope - K intercept on the t-axis equal to t0.
can be calculated, and plotted against t. From equation (3.5) this should give a straight line, slope - K intercept on the t-axis equal to t0.
Data on length at annual intervals can be obtained and tabulated in various ways. Probably the most straightforward is the analysis of the growth of a single fish - the length at the end of each year of life being obtained from measurements along an axis of a scale, otolith or other hard structure. Alternatively, the growth of a single year-class can be followed over a period of years - i.e. for the 1956 year-class the increment in the fifth year of life is the difference between the mean length of five-year-old fish at the end of 1961 and four-year-old fish at the end of 1960. Both these methods are concerned with the growth of a group of fish during their life, the growth during each year of life providing one point in the plot. Thus the growth increments refer to different calendar years; for example for the 1956 year-class the increment in the third year of life is put on in 1959, possibly under conditions very different to those in 1961, the fifth year of life. Growth in a particular year can be studied by taking the growth of different year-classes - e.g. data of growth in 1960 are given by the growth of the 1956 year-class in its fifth year of life, of the 1957 year-class in its fourth year of life - each year-class provides one point in the plot. This is a particularly useful method for studying the effect of the environment - food, temperature, density, etc. on growth. Finally, growth can be estimated from a single year's data, e.g. from data in 1960 the increment in the fifth year of life can be estimated from the difference in lengths of the 1955 and 1956 year-classes, the fourth year's increment from the difference between the 1956 and 1957 year-classes, etc. These differences do not, in fact, correspond to the growth of any particular fish or group of fish, and the method should be used only when other methods cannot. An improvement is to take the average size at each age over a number of years, but in this case it is usually possible, and more satisfactory, to analyse the growth of the individual year-classes or in each calendar year.
The weight of a fish is usually closely proportional to the cube of its length, so that from equation (3.2)
................ (3.6)
where W¥ is the asymptotic weight corresponding to the asymptotic length L¥ .
Similarly, if weight is proportional to the nth power of the length, 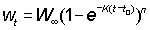 .
.
When growth data are given in terms of weight, fitting of growth curves is most easily done by using the cube-root of the weight as an index of length, fitting this to the equations of growth in length as above, and finally cubing to return to weight.
A more general growth equation has been given by Richards (1959); with a slight change from his notation this is
.............. (3.7)
For various values of m this equation becomes one or other of the common growth equations. Thus if 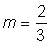 , equation (3.7) becomes the same as equation (3.2) for the von Bertalanffy curve. If m = 2 the equation, with some rearrangement, becomes the autocatalytic equation
, equation (3.7) becomes the same as equation (3.2) for the von Bertalanffy curve. If m = 2 the equation, with some rearrangement, becomes the autocatalytic equation
and for m = 0, the monomolecular equation
wt = W¥ (1 - ae-kt)
and it can be shown that in the limit as m ® 1, the equation becomes the Gompertz log wt = log W¥ (1 - ae-Kt). This curve satisfies a relation similar to that of equation (3.1), only in terms of log length rather than length, i.e.
The expressions above give the weight or length of a fish in terms of its age. Sometimes, the inverse procedure is required, i.e. it is desired to know the age of fish of a given length, e.g. selection data are normally in terms of length, but for incorporation in yield equations need to be expressed in terms of age.
If the seasonal growth pattern is very marked, and all fish of a particular age are nearly the same length, the actual mean age of fish of a given length may be quite different from the mean age determined from the average annual growth curve, particularly at the beginning and end of the season of rapid growth. If this is the case, the conversion of length to age is best made empirically from the observed age-length curve. Also, particularly if there is much individual variation in growth, the curve of mean length at a given age will be different from the curve of mean age at a given length (just as generally the regression line of y on x is different from the regression line of x on y). Usually, however, it is sufficient to convert length to age from a growth equation fitted to all the observed data of mean length at age, e.g. the von Bertalanffy equation
To obtain t in terms of l we divide both sides by L¥ , and subtract from unity, giving
Taking natural logs of both sides gives
and therefore
................. (3.8)
The span of age t2 - t1 between two lengths l1, l2 is therefore
................. (3.9)
1. The von Bertalanffy growth equation contains three parameters (L¥ , K and t0), which can therefore be estimated from three observations of length at age. From equation (3.2) write down the lengths at times t - 1, t, t + 1, and the increments in length between t - 1 and t and between t and t + 1. Hence determine an estimate for K from the ratio of these increments; by substituting this estimate for K in equation (3.2), or otherwise, find an estimate for L¥ , in terms of lt and the increments.
2. Kitahama (1955) gives the following size-at-age data for the Pacific herring, Clupea pallasii C. and V., caught off the western coast of Hokkaido from 1910-54. The lengths are averages over a forty-five year period; ages determined from scale rings.
|
Age |
Total length |
|
years |
cm |
|
3 |
25.70 |
|
4 |
28.40 |
|
S |
30.15 |
|
6 |
31.65 |
|
7 |
32.85 |
|
8 |
33.65 |
|
9 |
34.44 |
|
10 |
34.97 |
|
11 |
35.56 |
|
12 |
36.03 |
|
13 |
35.93 |
|
14 |
37.04 |
|
15 |
37.70 |
W = 7.8 × 103-3 L3 g(a) Construct a work-sheet and determine L¥ , W¥ , K and t0.
(b) Calculate the theoretical curves of growth in length and weight over the range 0-15 years.
3. Yokota (1951) gives the following data for growth of the shark Mustelus kanekonis (age determined from seasonal changes in length of claspers).
|
Age |
Weight |
|
groups |
g |
|
I ............... |
375 |
|
II ............... |
1 519 |
|
III ............... |
2 430 |
|
IV ............... |
3 247 |
W = 7.4 × 10-3 L3 g(a) Determine L¥ , W¥ , K and a working value for t0.
(b) Calculate curves of growth in length and weight up to age group VIII.
4. Posgay (1953) tagged sea scallops, and obtained the following data on growth during the ten months (approximately) between tagging and recapture.
|
|
SHELL DIAMETER | |||||||||
|
|
mm | |||||||||
|
At tagging ................ |
64 |
69 |
71 |
94 |
104 |
105 |
110 |
117 |
117 |
126 |
|
At recapture .............. |
98 |
102 |
93 |
115 |
120 |
126 |
125 |
127 |
136 |
138 |
(a) Calculate L¥ , and K.(b) Taking t0 = 0, calculate and plot the curve of growth in length up to six years.
5. Ochiai (1956) gives the following data for the growth of the soles Heteromycteris japonicus and Pardachirus pavoninus.
|
Age |
Standard length | |
|
|
H. japonicus |
P. pavoninus |
|
Years |
cm | |
|
1............... |
6.09 |
10.00 |
|
2............... |
8.10 |
14.80 |
|
3............... |
9.20 | |
(a) Determine arithmetically L¥ , K and t0 for each species (assume t0 = Q for P. pavoninus).(b) Demonstrate the patterns of growth in weight, assuming a cubic relation of weight to length.
6. Moiseev (1946) gives the following data for growth (total length in cm) of the flatfish Limanda aspera in Peter the Great bay, Japan sea.
|
Age
|
Sex
|
SAMPLING YEAR |
||||
|
1928 |
1930 |
1931 |
1932 |
1933 |
||
|
1
|
|
5.15 |
7.60 |
6.58 |
7.72 |
8.34 |
|
|
5.77 |
7.22 |
7.20 |
9.50 |
7.34 |
|
|
2
|
|
10.25 |
13.60 |
11.70 |
13.80 |
14.10 |
|
|
11.77 |
11.88 |
13.20 |
15.50 |
12.68 |
|
|
3
|
|
16.40 |
19.40 |
17.42 |
18.82 |
19.44 |
|
|
17.41 |
19.00 |
18.80 |
21.00 |
18.68 |
|
|
4
|
|
20.45 |
24.00 |
21.22 |
22.92 |
24.32 |
|
|
22.13 |
25.00 |
23.60 |
25.50 |
25.32 |
|
|
5
|
|
25.05 |
26.80 |
25.42 |
26.38 |
27.66 |
|
|
26.51 |
26.12 |
27.60 |
29.50 |
27.32 |
|
|
Index of total demersal catch..... |
1 |
1 |
5.5 |
8.0 |
7.7 |
|
(a) Analyse sexual differences in growth.(b) Bearing in mind that large-scale fishing was started in the bay only in the season 1929-30, and that L. aspera formed 80 percent of the demersal catch, examine the year-to-year variation in growth. The mean age composition by number, during 1931-33, was as follows:
|
Age |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
Percent |
0.6 |
4.7 |
16.1 |
33.3 |
27.9 |
11.3 |
3.4 |
1.7 |
0.9 |
0.2 |
(c) Estimate the total mortality coefficient and examine the growth data for bias in mean length at age, due to selection.(This part of the exercise is to be completed after sections 5 and 7.)
![]()
![]()
![]()
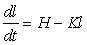
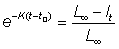
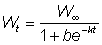
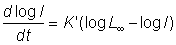
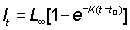
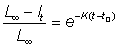
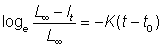
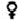
{kind=link}
{kind=link}