![]()
![]()
![]()
5.1 LONGUEUR AU DEBUT DE LA MATURITE
5.2 RAPPORT LONGUEUR-POIDS
5.3 REPRODUCTION
5.4 CROISSANCE
5.5 MORTALITE
A partir d'échantillons prélevés entre 1967 et 1969, Fontana et Pianet (1973) ont réuni des données sur la longueur des sardinelles au début de la maturité dans le secteur congolais. Ils ont constaté que toutes les femelles de S. aurita de plus de 21 cm et toutes les femelles de S. maderensis de plus de 20 cm ont à ce stade commencé leur cycle sexuel (troisième phase ovarienne). Les graphiques proposés montrent que 50 pour cent des femelles de S. aurita ont entamé leur troisième phase ovarienne à environ 19 cm et celles de S. maderensis à un peu plus de 18 cm.
Pour Fontana et Pianet, tout au moins en ce qui concerne S. maderensis, la preuve est faite qu'il existe, suivant la saison, une différence de longueur au début de la maturité.
Les échantillons ne portaient que sur une seule année complète et on ne dispose d'aucune donnée sur d'éventuelles variations de longueur susceptibles de se produire d'une année sur l'autre au début de la maturité.
Au large du Gabon, au nord du 2 30'S, S. aurita est beaucoup plus petite que plus au sud et 50 pour cent des femelles atteignent la maturité à environ 14 cm, ce qui laisse entendre qu'il existerait un stock distinct et limité de S. aurita dans ce secteur.
Le rapport longueur-poids n'a été établi que pour les sardinelles du secteur congolais (Fontana et Pianet, 1973). Les données recueillies portent sur les années 1965 et 1967 pour S. aurita et sur 1969 pour S. maderensis.
Les auteurs ont observé une forte variation saisonnière de l'exposant (b) de l'équation W = a x Lb (dans laquelle W représente le poids en grammes, L la longueur a la fourche en cm, et a et b sont des constantes). Toutefois, pour ces mêmes auteurs, les valeurs-clés longueur-poids calculées à l'aide des équations pour les différentes saisons sont extrêmement voisines et c'est pourquoi ils ont adopté un rapport généralisé pour chaque espèce, à savoir:
S. maderensis W = 5,94 x 10-6 x L 3,183Les remarques formulées par ces deux auteurs sur les variations annuelles du facteur conditionnel (le poids divisé par le cube de la longueur) pour chaque espèce (voir tableau ci-dessous) laissent supposer de probables variations de ce rapport d'une anné à l'autre.S. aurita W = 4,25 x 10-6 x L3,218
Variations saisonnières du facteur conditionnel
(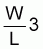 ).
).
Différence (en pourcentage du minimum) entre la valeur mensuelle la plus faible et la valeur mensuelle la plus élevée du facteur conditionnel pour chaque année (d'après Fontana et Pianet, 1973).
|
|
1967 |
1968 |
1969 |
1970 |
|
S. aurita |
30 |
23 |
12 |
36 |
|
S. maderensis |
24 |
14 |
5 |
15 |
5.3.1 Sardinella aurita
5.3.2 Sardinella maderensis
Divers auteurs et notamment Fontana (1969), Dessier et Pianet (1971) et Fontana et Pianet (1973), ont étudié la reproduction des sardinelles dans le secteur congolais. La question de la reproduction des sardinelles en Angola n'a fait l'objet d'aucune publication, bien qu'il semble que certaines données pertinentes aient été recueillies dans la région. Les deux espèces se reproduisent de manière très différente.
La conclusion de Fontana et Pianet (1973) est que cette espèce ne fraye qu'en saison froide, dans des eaux dans lesquelles la productivité a augmenté, en raison de phénomènes hydrographiques tels que le upwelling (remontée d'eau froide). En outre, de fortes variations de l'indice gonadosomatique des femelles (poids de l'ovaire divisé par le cube de la longueur) (cf. tableau 6) indiquent d'importantes fluctuations de l'activité sexuelle des stocks exploités par les senneurs de Pointe Noire (Fontana, 1969):
a) fluctuations saisonnières indiquant l'existence d'une courte période de frai pendant la “courte saison froide” (décembre - janvier) et d'une période de frai principale pendant la “longue saison froide” qui s'étend de juin à septembre;Comparant les captures enregistrées en 1970-1972 et l'indice gonadosomatique relatif aux saisons antérieures, Ghéno et Fontana (1973) concluent que variabilité du frai se traduit par une forte variabilité de l'abondance des classes d'âge constituant le recrutement du secteur congolais et, par voie de conséquence, des taux de capture de cette pêcherie.b) fortes fluctuations d'une année à l'autre: l'activité sexuelle, mesurée d'après l'indice gonadosomatique, a été intense en 1967, 1968 et 1971 et très faible en 1969 et 1972.
On notera que l'indice gonadosomatique n'est pas nécessairement un indice de la production totale d'oeufs, laquelle dépend à la fois de l'activité des femelles en matière de ponte et du nombre de femelles contenues dans le stock. Il conviendrait à cet égard d'étudier plus avant la question de savoir s'il existe une relation entre l'indice gonadosomatique et le recrutement.
L'inégalité de la répartition spatio-temporelle (saisonnière) du frai et confirmée par les observations de Dessier et Pianet (1971) sur la répartition des larves de sardinelles dans le secteur situé entre Libreville et le fleuve Congo, de mai à juillet 1968 (8 campagnes, 80 stations) (tableaux 7a et 7b).
La fécondité de S. aurita dans les eaux congolaises a été étudiée par Fontana et Pianet (1973) qui ont trouvé la relation suivante:
F = 436,415 W - 22076,437
dans laquelle F = fécondité (nombre d'oeufs par femelle) et W = poids en grammes.
On ne dispose d'aucune donnée quantitative sur la reproduction de S. aurita dans le secteur angolais. La présence de juvéniles de S. aurita dans ces eaux et la capture de femelles matures (Ghéno et Campos Rosado, 1972) montrent toutefois que cette espèce fraye également dans les eaux angolaises.
De l'analyse des données relatives à l'indice gonadosomatique (tableau 6b), des observations sur la répartition des larves (tableau 7a) et des données sur la capture des juvéniles, Fontana et Pianet (1973) concluent que cette espèce fraye tout au long de l'année dans toute la zone côtière qui va du Gabon à Cabinda, avec un maximum pendant la saison froide.
Les mêmes auteurs ont exprimé la fécondité de S. maderensis dans le secteur congolais par la formule suivante:
F = 418 W - 18974
dans laquelle F = nombre d'oeufs par femelle et W = poids en grammes.
On est en droit de penser que cette espèce se reproduit aussi dans les eaux congolaises, mais on ne dispose d'aucune donnée quantitative à l'appui de cette hypothèse.
5.4.1 Sardinella aurita
5.4.2 Sardinella maderensis
Ghéno (1968, 1975) a étudié la croissance de S. aurita à partir d'échantillons provenant de la pêcherie de Pointe Noire (Congo). Ces études sur la croissance reposent sur la lecture des anneaux de croissance sur les écailles ainsi que sur d'autres éléments tels que:
a) Les saisons de frai, qui ont été définies d'après l'état de développement des gonades. Comme il est indiqué à la section 4.5, les saisons de frai sont généralement faciles à distinguer chez S. aurita.Deux groupes d'individus ont été identifiés, à savoir: les individus nés au cours de la ''courte saison froide” (“naissance” au 1er janvier) et les individus nés au cours de la “longue saison froide” (“naissance” au 1er mai). On a utilisé l'équation de von Bertalanffy pour calculer le rapport longueur-âge de chaque groupe de poisson, ce qui donne les paramètres suivants:b) Analyse de la fréquence des longueurs.
c) Détermination du nombre de vertèbres permettant d'identifier les groupes d'individus nés à une même température de l'eau de mer.
|
“Naissance” |
1er janvier |
1er mai |
|
L¥ |
25,97 |
26,00 |
|
K |
1,023 |
1,210 |
|
to |
-0,08 |
-0,02 |
Ghéno et Le Guen (1968) ont étudié la croissance des S. maderensis capturées dans le secteur congolais en 1966 et 1967.
Pour ce faire, ces deux auteurs ont procédé à des lectures d'écailles et à l'identification de groupes de poissons nés dans des conditions hydrographiques analogues, comme en témoigne le nombre moyen de vertèbres. Cette analyse a été rendue compliquée par le fait que cette espèce n'a pas de périodes de frai caractéristiques.
L'équation de von Bertalanffy appliquée a ces données a donné les paramètres suivants:
L¥ = 24,92
K = 0,984
to = 0,287
Sardinella maderensis a donc les mêmes caractéristiques de croissance que S. aurita, a savoir un développement très rapide chez les jeunes et une croissance ralentie par la suite. Comme dans le cas de S. aurita, il se peut que la croissance soit différente selon le secteur et l'année considérée. Sans doute serait-il possible d'améliorer les études relatives a la croissance en analysant l'ensemble des données recueillies à Pointe Noire depuis 1968, époque a laquelle Ghéno et Le Guen ont procédé à leur analyse.
Il convient de noter que la nette différence de longueur des S. maderensis capturées d'une part dans les eaux congolaises et d'autre part dans le secteur angolais en 1974 (voir annexe 3) pourrait être partiellement imputable à des différences entre les normes de mesure (longueur a la fourche ou longueur totale); elle ne saurait donc être considérée comme l'indice d'une différence de taille entre les individus peuplant différents secteurs. Il se pourrait, toutefois, que S. maderensis des eaux congolaises et celle du secteur congolais aient une croissance différente, étant donné les différences de conditions hydrographiques .
5.5.1 Sardinella aurita
5.5.2 Sardinella maderensis
Ghéno (1972) a calculé le taux global de mortalité apparent (Z) de S. aurita dans le secteur congolais. Il s'est fondé sur le fait que la génération 1964 de cette espèce a été très peu abondante, ce qui a facilité la distinction entre les juvéniles et les individus nés en 1963 ou antérieurement. Ghéno a donc calculé la valeur Z en fonction de la diminution des cpue des individus de la génération 1963 ainsi que des générations antérieures pour la période 1964-1969. Il a utilise une méthode analogue pour étudier les générations 1968 et antérieures des captures de 1970 et 1971.
Ces deux analyses ont livré des estimations très voisines du taux global de mortalité soit respectivement 0,76 et 0,79. Il se pourrait néanmoins que la méthode utilisée ait produit des résultats faussés et ce, pour les raisons suivantes:
a) les données utilisées pour ces études provenaient de l'unique senneur opérant dans le secteur durant la période considérée et les prises de ce senneur ne donnaient pas nécessairement une représentation fidèle du stock de sardinelle du secteur en question;On ne dispose d'aucune estimation faite à partir de l'étude des poissons capturés dans le secteur angolais.b) il n'a été tenu aucun compte d'éventuelles modifications de la puissance de pêche du navire utilisé pour ces études;
c) la vulnérabilité des stocks ou de certaines classes d'âge a pu fluctuer d'une année à l'autre, en fonction des conditions hydrographiques;
d) certains doutes demeurent quant à l'âge des poissons étudiés, notamment des spécimens plus âgés.
A noter que pour les clupéidés, d'après Beverton et Holt (1959) et Cushing (1968), le rapport M/K a généralement une valeur proche de l'unité.
Cela signifie que, si l'on obtient une estimation de K voisine de l'unité, comme indiqué ci-dessus. M, qui est le taux de mortalité naturelle, devrait aussi être autour de 1.
Encore qu'il soit difficile de tirer des conclusions précises des renseignements dont on dispose, il semblerait, d'après les données recueillies, que le taux de mortalité due à la pêche de S. aurita a été faible dans le secteur au cours des périodes pour lesquelles on a pu faire des estimations de la mortalité.
Ghéno a calculé le taux global de mortalité apparent (Z) de S. maderensis dans la même étude que celle ou il parle de S. aurita (1972). Pour ce faire, il a utilisé deux méthodes différentes:
a) L'approximation donnée par Beverton et Holt (1956)
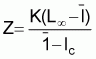
dans laquelle Z = taux global de mortalité apparent
L¥ et K = paramètres de l'équation de croissance de von BertalanffyGhéno a utilisé les valeurs suivantes, à savoir: L¥ =19,50 cm, K = 0,98 cm, l = 24,93 cm et lc =21,24 cm, ce qui lui a permis d'obtenir une valeur Z = 2,12. Le Groupe de travail ad hoc note que la valeur de Z ainsi obtenue est très sensible a des différences de la valeur l - lc, l'étude de la composition des captures par longueur laisse penser que lc pour - rait être plus proche de 16 cm que de la valeur utilisée, a savoir 21,24 cm. Par exemple, si l'on appliquait la valeur lc = 16 cm dans l'équation, on trouverait une estimation de Z = 0,70.
lc = longueur moyenne au recrutement c
l = longueur moyenne du poisson capturé
b) La conversion de la composition par longueur des captures de 1964-1969 en composition par âge au moyen de l'équation de croissance de von Bertalanffy établie pour l'espèce en question. La diminution moyenne des captures par groupe d'âge représente alors le taux global de mortalité apparent des individus du stock, estimé grâce à cette méthode à 1,93.
Plusieurs facteurs ont pu fausser cette estimation:
i) la conversion de la composition par longueur en composition par âge sur la base des données relatives à la croissance moyenne apparaît très difficile étant donné que les individus de plus de 20 cm de long se développent très lentement et que la longueur des poissons d'une même cohorte est très variable. Cette variabilité, tout comme la variabilité de la croissance selon les cohortes, est sans doute bien supérieure à la croissance annuelle théorique des individus âgés. Il en résulte également qu'il est difficile de répartir entre les diverses classes d'âge les individus d'une longueur supérieure à L¥.Il convient donc de tenir ces estimations de Z pour provisoires et de les utiliser avec circonspection.ii) L'estimation est vraisemblablement sensible à la valeur des paramètres de croissance utilisés pour convertir la composition par longueur en composition par âge, et il semblerait que les paramètres de croissance établis pour S. maderensis ne puissent être considérés comme très précis.
iii) La composition par longueur des captures peut constituer un échantillon erroné de la composition par longueur des stocks du secteur de Pointe Noire.
![]()
![]()
![]()