![]()
![]()
![]()
A. Lahlou-Kassi et R. BoukhliqDépartement de Reproduction Animale et d'Insémination Artificielle
Institut Agronomique et Vétérinaire Hassan II
B.P. 6202
Rabat-Instituts
Maroc
Resume
Summary
Introduction
Contraintes environnementales et comportement sexuel saisonnier
Variations saisonnières du comportement sexuel
Synchronisation des chaleurs
Modification de la période de lutte
Conclusions
References
Les variations saisonnières de l'activité sexuelle chez le mouton dépendent des variations de la durée du jour. L'expression oestrale peut être déprimée par les photopériodes longues. Le maintien d'une activité spermatogénétique élevée est possible par l'utilisation de cycles lumineux courts. La mélatonine semble nécessaire pour la perception de la durée du jour et ses variations: son utilisation chez des femelles en anoestrus permet d'avancer le début de la saison sexuelle mais une injection en pleine saison sexuelle n'étend pas celle-ci et la saison suivante n'est pas affectée. Chez le bélier la mélatonine, administrée pendant la période de longues journées, stimule la croissance des testicules de la même manière qu'en journées courtes mais quand administrée sur une circonférence scrotale maximale elle n'empêchera pas la régression des testicules quand la photopériode croît. Parmi les facteurs environnementaux non photopériodiques susceptibles d'affecter la saison sexuelle, "l'effet bélier" reste le moyen le plus simple et le plus économique pour avancer le début de la saison sexuelle et synchroniser les saillies sans faire appel à une thérapeutique hormonale.
Seasonal variation in reproductive activity of sheep is related to variations in daylength. Overt oestral activity may be depressed by long days. Active spermatogenesis can be maintained by using short duration light cycles. Melatonin appears to be necessary for the perception of daylength and its variations. Use of melatonin on anoestrus ewes results in an earlier breeding season but its injection in the middle of the season does not prolong it, nor does it affect the subsequent season. Melatonin injected into rams during the long day period stimulates testicular growth in the same way as it does when injected during short days but, if used when scrotal circumference is at a maximum, does not prevent regression when daylength increases. Among the environmental factors not related to daylength, the "ram effect" is the simplest and most economic method for advancing the breeding season and for synchronizing mating without having recourse to hormonal treatment.
Dans les systèmes traditionnels d'élevage des petite ruminants en Afrique, la permanence des béliers dans les troupeaux est la pratique dominante (Lahlou-Kassi, 1987; Faugère et al, 1988; Wilson, 1988). L'étalement des mises bas avec des pies d'agnelage plus ou moins marqués selon les latitudes et les écosystèmes observés n'est pas toujours souhaitable pour optimiser la productivité d'un troupeau.
La manipulation et le contrôle de la saison sexuelle sont souvent envisagés dans les programmes d'amélioration de la productivité des troupeaux ovins. C'est pour permettre d'élaborer un programme adapté de gestion de la reproduction que les recherches rapportées ici ont été menées.
Le choix de la saison de lutte et de sa durée est fonction de la variation saisonnière des paramètres d'environnement (température, pluviométrie), des facteurs de production (disponibilités alimentaires et des sous-produits agricoles), des maladies ou affections des jeunes agneaux et du comportement sexuel des races exploitées.
Le calendrier des saisons des disponibilités alimentaires et de reproduction diffère selon les régions (figure 1). Dans la région méditerranéenne le pic d'agnelage se situe en nov/déc, période froide et parfois pluvieuse. Les ressources fourragères sont très rares et la mortalité des jeunes agneaux est élevée (20 p.cent). La période de lutte principale (jun/jul) coïncide avec la période des récoltes (quand les chaumes et les sous-produits agricoles sont disponibles) et les fortes chaleurs estivales.
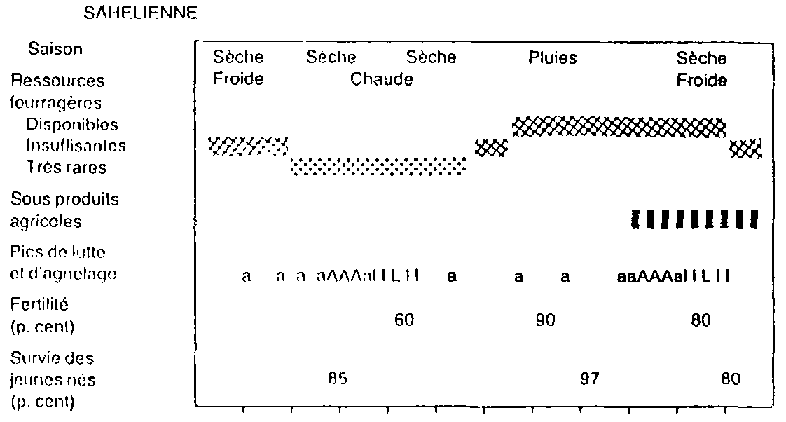
Figure 1. Calendrier des événements influant la reproduction en trois zones africaines. - SAHELIENNE
Figure 1. Calendrier des événements influant la reproduction en trois zones africaines. - SUBHUMIDE
Dans la région sahélienne on note 2 pics d'agnelage. Le 1er pic se situe en avr/mai, période sèche et chaude. Les ressources fourragères sont rares et la mortalité des jeunes est élevée (15-20 p.cent). Le 2ème pic a lieu en sep/oct, période de fin des pluies et début de saison sèche, à forte disponibilité de fourrages et de sous-produits agricoles. Ce dernier pic correspond à la lutte des mois d'avr/mai où le taux de fertilité des brebis est le plus bas (60 p.cent), le déficit alimentaire et l'anoestrus saisonnier étant certainement les causes.
Dans la zone subhumide les agnelages sont étalés tout au long de l'année, avec un léger pic en nov-déc. Ce pic a lieu à une période très favorable où l'alimentation est bonne (fourrages et sous-produits agricoles disponibles) et la température est clémente. La lutte principale a lieu, par contre, en début de saison des pluies où les brebis sont en mauvais état corporel et leur fertilité est réduite.
Pour un ensemble des races étudiées en Afrique du Nord et Sahélienne, il est évident qu'il y a une période de faible activité sexuelle et ovarienne centrée en mars (figure 2) sauf pour la race D'man qui montre un comportement sexuel constant tout au long de l'année. Pour les autres races méditerranéennes la période d'anoestrus saisonnier est de 3-4 mois (fév-mai) et les taux de fertilité enregistrés pour cette période sont très faibles (tableau 1). Pour la race sahélienne Peul et la race Djallonké de la zone subhumide, l'anoestrus saisonnier est moins marqué et est limité en mars et en avril.
Tableau 1. Taux (p.cent) de quelques paramètres de reproduction pour deux races de moutons marocains.
|
Race et paramètre |
Dates de lutte |
|||||
|
2.viii-10.xiii 1982 |
1.vi-20.vii 1983 |
15.xii-30.i 1983/4 |
29.ix-10.x 1984 |
15.xii-30.vi 1985 |
||
|
Sardi |
|
|
|
|
|
|
|
|
Fertilité |
100 |
75 |
45 |
96 |
64 |
|
|
Prolificité |
117 |
118 |
128 |
127 |
116 |
|
|
Agneaux/brebis mises en lutte |
1,17 |
0,80 |
0,50 |
1,21 |
0,70 |
|
|
D'man |
|
|
|
|
|
|
|
Fertilité |
- |
92 |
88 |
95 |
86 |
|
|
Prolificité |
- |
233 |
219 |
249 |
208 |
|
|
Agneaux/brebis mises en lutte |
- |
2,14 |
1,93 |
2,37 |
1,79 |
Il semble, de cette analyse, que la période favorable pour la naissance et le bon démarrage des jeunes est limitée à 2-3 mois et une synchronisation des saillies (et donc des agnelages) est susceptible d'améliorer la productivité des troupeaux ovins. Dans certaines situations, cette période favorable des saillies coïncide avec la période de baisse d'activité sexuelle, d'où l'application de méthodes d'induction d'ovulation et de synchronisation des chaleurs semblerait nécessaire.
Utilisation des progestagènes
Le traitement aux progestagènes (FGA - Acétate de fluorogestone, MAP - Acétate medronyprogestérone) combiné aux gonadotropines (PMSG, HCG, GnRH) est utilisé pour avancer la saison sexuelle, pour synchroniser les chaleurs ou améliorer la prolificité.
Chez les agnelles. Suite à un traitement à base d'éponges vaginales (60 mg MAP) et 500 ui d'HCG chez des antenaises de races Awassi, Karadi et Arabi, le pourcentage des femelles observées en chaleurs 47-51 h après le retrait des éponges est de 60 p.cent à 94 p.cent selon les races (Hamra et al, 1988). Le taux de gestation observé après saillie naturelle est d'environ 60 p.cent.
Chez les adultes. Le taux de femelles à oestrus synchronisés après le traitement à base de progestérone-gonadotropines est très variable selon les races traitées (tableau 2). Des pourcentages de réponse de 53 p.cent à 100 p.cent ont été observés chez la Sardi et la D'man. Cette variabilité entre races pourrait être liée à leur niveau de cyclicité au moment du traitement et à leur état corporel. L'adjonction de la PMSG au traitement progestérone améliore significativement le taux de femelles en oestrus chez les races saisonnières, telle la Beni Hsen.
Le taux d'agnelage après synchronisation varie de 20 p.cent à 70 p.cent et est nettement inférieur à celui obtenu chez les brebis non traitées. Le taux d'agnelage après insémination artificielle est beaucoup plus faible, de 3 p.cent à 40 p.cent.
Tableau 2. Résultats d'expériences de synchronisation des chaleurs par les progestagènes et les gonadotropines.
|
Race |
n |
Traitement |
Réponse (p .cent) |
Taux d'agnelage1) |
Références | |
|
Beni Hsen |
29 |
Progestérone |
69,9 |
55,0 |
S |
Dkhissi, 1977 |
|
Beni Hsen |
54 |
Progestérone + 500 ui PMSG |
87,5 |
56,5 |
S |
Dkhissi, 1977 |
|
Sardi |
34 |
FGA + PMSG |
53,0 |
3,0 |
I |
Manar et al, 1988 |
|
D'man |
30 |
FGA + PMSG |
76,0 |
10,0 |
I |
Manar et al, 1988 |
|
D'man |
23 |
FGA + 400 ui PMSG |
100,0 |
21,7 |
I |
Tibary et al, 1988 |
|
Timahdite |
23 |
FGA + 400 ui PMSG |
91,0 |
39,1 |
I |
Tibary et al, 1988 |
|
Timahdite |
50 |
FGA + 400 ui PMSG |
- |
20,0 |
S |
Tibary et al, 1988 |
|
Timahdite |
100 |
Témoin |
- |
70,0 |
S |
Tibary et al, 1988 |
|
Touabire |
10 |
FGA |
- |
75,0 |
S |
Mbaye, 1981 |
|
Touabire |
10 |
FGA + 100 ui PMSG |
- |
77,7 |
S |
Mbaye, 1981 |
|
Touabire |
10 |
FGA + 200 ui PMSG |
- |
50,0 |
S |
Mbaye, 1981 |
Note: 1. S - saillie naturelle, I - insémination artificielle.
Utilisation des prostaglandines
L'utilisation des prostaglandines pour la synchronisation des chaleurs se fait à travers 2 injections espacées de 9 j à 11 j chez les femelles cyclées mais la réponse à ce traitement est assez variable. Au Maroc, l'utilisation des PGF2-a chez des brebis D'man et Timahdite donne un pourcentage de réponse (oestrus) de 65 p.cent et 47 p.cent et un taux de fertilité de 35 p.cent et 13 p.cent après l'insémination à temps fixe pour les 2 races (Tibary et Manar, 1988). La variabilité de la réponse semble être liée à l'étalement chez les animaux ne répondant pas au traitement.
Les traitements hormonaux en vue, soit de synchroniser les agnelages, soit d'utiliser l'insémination artificielle dans un programme d'amélioration génétique, ne sont pas encore maitrisés. Des expérimentations plus élaborées sont nécessaires: celles-ci doivent tenir compte de la saisonnalité de l'activité ovarienne et de la réponse aux gonadotropines spécifiques à chaque race, mais aussi de l'état corporel des animaux au moment du traitement.
Traitement lumineux
La plupart des investigations concernant le rôle et l'utilisation de la photopériode ont concerné les ovins originaires des pays tempérés ayant une saison de reproduction limitée. Les données relatives aux ovins des régions intertropicales sont peu nombreuses.
Pour tester la sensibilité aux variations de la photopériode, certains régimes lumineux ont été appliqués sur des brebis au Maroc (Sardi) et en Egypte (Rahmani). Au Maroc, ceci a lieu à travers l'examen du comportement sexuel de brebis soumises à un régime lumineux artificiel simulant la photopériode à la latitude 56°N (Ecosse) ou à la photopériode naturelle (32°N). Chez la plupart des brebis du lot témoin, l'activité sexuelle commence en mai (figure 3) pour s'estomper progressivement en décembre. Chez les brebis du lot traité (régime lumineux écossais), le début de la saison sexuelle a été retardé de 2 mois (août au lieu de juin). Le pourcentage de femelles en oestrus ou en activité ovarienne est significativement plus faible dans le lot traité que dans le lot témoin en mai et en juin. Un protocole similaire, réalisé sur des brebis Rahmani, montre une légère diminution de l'activité oestrale et ovarienne pendant la période des jours croissants (jan-mai). Une décroissance graduelle de la photopériode imposée pendant cette période donne lieu A une augmentation du nombre de femelles en chaleur (Aboul Naga et al, 1988).
Figure 3. Variations saisonnières des brebis Sardi ovulant (a), ou en oestrus (b) sous photopériode naturelle (34°N - - - ) ou artificielle (56°N----).

Ces 2 expériences suggèrent que le début de la saison d'activité sexuelle peut être avancé ou retardé par l'application de régimes lumineux appropriés. L'application de cette méthode par les éleveurs nécessiterait un investissement en bâtiments d'élevage relativement couteux. C'est pourquoi des investigations concernant l'utilisation de la mélatonine en remplacement du traitement lumineux peuvent être envisagées.
Traitement à la mélatonine
Le message photopériodique qui stimule l'apparition ou l'arrêt de l'activité sexuelle est transmis à travers la glande pinéale qui le convertit en signal hormonal. Ce signal prend la forme d'un rythme circadien de sécrétion de la mélatonine (Karsh, 1984). L'utilisation de la mélatonine pour avancer le début de la saison sexuelle a été largement étudiée (Kennaway et al, 1982; Crocker et al, 1987) et a conduit A la fabrication d'une mélatonine synthétique (Regulin, Gene Link Australia Limited). La libération continue de mélatonine par des implants induit un effet des 'jours courts" tout A fait comparable A celui obtenu par un apport particulier de mélatonine dans la nourriture ou par des photo-périodes courtes (Arendt, 1988).
L'application répétée d'un implant sous-cutané de mélatonine (350 mg) tous les 10 j permet d'avancer le début de la saison sexuelle chez les brebis à condition qu'elles aient subi une période préalable de jours longs naturels ou artificiels (Johnston et al, 1988; Jordan et al, 1988;
Chemineau et al, sous presse). Dans la région méditerranéenne l'utilisation de la mélatonine pour améliorer la fertilité de la lutte de printemps (fév-avr) nécessite la mise des brebis sous régime lumineux artificiel simulant les jours longs pendant la période précédant le traitement.
"L'effet bélier"
"L'effet bélier" est utilisé, soit pour avancer le début de la saison sexuelle, soit pour rompre l'anoestrus postpartum, L'introduction de béliers entiers ou de leurs substituts (mâles castrés et traités aux stéroïdes, femelles traitées aux androgènes, ou odeur des béliers) dans un troupeau de femelles non cyclées induit l'ovulation chez la grande majorité des brebis (D R Lindsay et J Thimonier, comm. pers.). Cet effet est d'autant plus marqué que l'on se rapproche du début de la saison sexuelle. Le stimulus du mâle induit une accélération des décharges pulsatiles de LH conduisant à un pic préovulatoire et donnant lieu à l'ovulation 48 h environ après la mise au mâle.
Réponse à "l'effet bélier" en fonction de la saison. La mise au bélier de brebis Sardi en début mai (période où normalement le pourcentage des femelles cyclées est de 40 p.cent) a induit l'expression oestrale chez 80 p.cent des femelles (figure 2). Il y a donc induction de l'ovulation et de l'oestrus par "l'effet bélier" chez 67 p.cent des femelles non cyclées. L'analyse de la distribution des saillies des brebis Sardi révèle 2 pics de saillies centrés autour du 17 j et du 25 j après la mise au bélier. Des observations similaires ont été rapportées chez la race Barbarine (Khaldi, 1984), et pour cette race il a été démontré que les femelles en oestrus A 25 j avaient un cycle court (6-7 j), commençant 2 j à 3 j après la mise au bélier.
La suppression des cycles courts par l'administration de progestérone (20 mg) au moment de la mise au bélier permet une meilleure induction et synchronisation de l'ovulation et des saillies vers le jour 19 (D R Lindsay et J Thimonier, comm. pers.). Au milieu de la saison sexuelle, alors que l'ensemble des femelles sont cyclées, l'introduction du bélier n'entraîne aucune synchronisation des saillies. En effet, dans le lot de brebis Sardi, les saillies sont régulièrement réparties tout au long des premiers 17 j.
Effet de la race du bélier. Pour la lutte de mai/jun (début de saison sexuelle de brebis Sardi), le bélier D'man a induit l'ovulation et l'oestrus chez 90 p.cent des femelles Sardi jusqu'à 17 j. alors que ce pourcentage n'est que de 27 p.cent en présence d'un bélier Sardi (figure 4). Le bélier D'man produit un "effet mâle" significativement plus marqué que le bélier Sardi. Ce résultat pourrait être mis à profit en utilisant un bélier D'man vasectomisé comme "teaser" pour synchroniser les saillies des femelles Sardi.
Réduction de l'intervalle entre agnelages par "l'effet mâle". L'utilisation de "l'effet bélier" pour réduire l'anoestrus postpartum a été testée sur des brebis de race Barbarine (tableau 3). Les résultats obtenus sont encourageants puisque 90 p.cent à 100 p.cent d'ovulations induites ont été obtenues chez des femelles non cyclées mises aux béliers 25 j à 35 j après la mise bas (Khaldi, 1984). Cependant, à 15 j du part, l'introduction des béliers n'a induit que 70 p.cent d'ovulations, suivis dans 71 p.cent de cas par des cycles courts.
Figure 4. Distribution des saillies
Tableau 3. Réponse (p.cent) des brebis Barbarine postparturientes à l'effet bélier.
|
Paramètre |
Temps après agnelage (j) |
||
|
15 ± 2 |
25 ± 2 |
25 ± 2 |
|
|
Activité ovarienne avant l'introduction des brebis |
- |
40,0 |
40,0 |
|
Ovulations induites chez les femelles non cyclées (3 premiers jours) |
70,0 |
91,7 |
100,0 |
|
Fréquence des cycles courts |
71,4 |
27,3 |
25,0 |
|
Taux d'ovulation induite |
1,41 |
1,64 |
1,42 |
|
Femelles rentrant en anoestrus après l'introduction des béliers |
35,7 |
27,3 |
58,3 |
Le contrôle de la saison de lutte s'avère d'une grande importance pour mettre à profit les ressources alimentaires qui sont très saisonnières en Afrique. Ce contrôle doit passer d'abord par l'adoption, par les éleveurs, du principe de la lutte contrôlée, en retirant les mâles adultes du troupeau des femelles reproductrices. L'emploi de méthodes de synchronisation et d'induction des chaleurs faisant appel à des traitements lumineux ou hormonaux est prématuré. Des expérimentations complémentaires de ces techniques dans nos conditions vent encore nécessaires. Les résultats d'utilisation de "l'effet bélier" sont, par contre, plus prometteuses et cette méthode pourrait être expérimentée directement au niveau de l'éleveur pour les luttes à contre saison.
Aboul Naga A M, Aboul Ela M B. El Nakhla A et Mehrez A Z. 1988. Oestrus and ovarian activity of subtropical fat-tailed Rahmani sheep and their response to light treatment. Journal of Agricultural Science, Cambridge 108: 617-621.
Arendt J. 1988. Role of the pineal gland in seasonal reproductive function in mammals. Oxford Reviews on Reproductive Biology 8: 266-320.
Chemineau P. Pelletier J. Guerin Y. Colas G. Ravault J P. Touré G. Almeida G. Thimonier J et Ortavant R. sous presse. Photoperiodic and melatonin treatments for the control of seasonal reproduction in sheep and goats. Reproduction, Nutrition, Développement.
Crocker K, Dunstan E, Reeve J. Williams A, McPhee S. Ayton B. Wagg M, Pollard T. Parker J. Foote M, Ritar A et Staples L. 1987. Field studies of the use of melatonin implants to improve lambing percentage in spring joined Merino and crossbred ewes. Proceedings of the Australian Society of Reproductive Biology 19: 21.
Dkhissi A. 1977. Induction de l'oestrus et de l'ovulation à contre saison. Thèse Doctorat Vétérinaire. Institut Agronomique et Vétérinaire Hassan II, Rabat, Maroc.
Faugère O, Faugère B. Merlin P. Dockes C et Perrot C. 1988. L'élevage traditionnel des petite ruminants dans la zone de Kolda (Haute Casamance). Institut Sénégalais des Recherches Agricoles/Institut d'Elevage et de Médecine Vétérinaire des Pays Tropicaux, Dakar, Sénégal (miméo).
Hamra A H. Al Jalil Z F. Alkass J E et Al Hiti S M. 1988. Fertility in ewe lambs pretreated with progestagen intravaginal sponges and injected with HCG or GnRH. Proceedings of the 11th International Congress on Animal Reproduction and Artificial Insemination 1: 409.
Johnston J P J. Quirke J F. Boland M P et Roche J F. 1988. The effect of continuous or intermittent melatonin on seasonal breeding of ewes . Proceedings of the 11th International Congress on Animal Reproduction and Artificial Insemination 1: 408.
Jordan B. Hanrahan J P et Roche J F. 1988. The effect of melatonin implantation in January on the breeding season of ewes. Proceedings of the 11th International Congress on Animal Reproduction and Artificial Insemination 1: 410.
Karsh F J. 1984. Endocrine and environmental control of oestrus cyclicity in sheep. In: D R Lindsay and D T Pearce (eds.), Reproduction in sheep. Australian Academy of Science, Canberra, Australia.
Kennaway D J. Gilmore T A et Seamark R F. 1982. Effect of melatonin feeding on serum prolactin levels and the onset of seasonal oestrus cyclicity in sheep. Endocrinology 110: 1766-1772.
Khaldi G. 1984. Variations saisonnières de l'activité ovarienne de l'anoestrus postpartum des femelles ovines de race Barbarine: Influence du niveau alimentaire et de la présence du mâle. Thèse Doctorat d'Etat. Université des Sciences Techniques du Languedoc, Montpellier, France.
Lahlou-Kassi A. 1987. Improving the reproductive performance of sheep in Morocco. In: A W Qureshi and H A Fitzhugh (eds.), Small ruminants in the Near East. Animal Production and Health Paper No.54. Food and Agriculture Organisation, Rome, Italy.
Manar S. Tibary A, Boukhliq R et Adnani M. 1988. Synchronisation des chaleurs et essai d'insémination artificielle chez des brebis de race locale. Journées de l'association Nationale de la Production Animale, 10-11 Mars 1988, Rabat, Maroc.
Mbaye M. 1981. Le mouton Peul-Peul et Touabire. Rapport d'activité. Centre des Recherches Zootechniques, Dahra, Sénégal.
Tibary A et Manar S. 1988. Factors affecting oestrus synchronisation in two Moroccan breeds of sheep. Proceedings of the 11th International Congress on Animal Reproduction and Artificial Insemination 1: 462.
Wilson R T. 1988. Small ruminant production systems in tropical Africa. Small Ruminant Research 1: 305-325.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}