Effects of climate change on plant pests
Case studies of individual pest species
Some pests have already expanded their host range or distribution, at least in part due to changes in climate. Examples of these pests, selected according to their relevance in different geographical areas, are summarized below. Examples of some likely effects of climate change on plant pests (insects, pathogens and weeds) in different climate zones are listed in Table 3.
Insects
1. Emerald ash borer (Agrilus planipennis) (Asia, Europe, North America)
The emerald ash borer, Agrilus planipennis, is a phloem-feeding beetle that infests ash trees (Fraxinus spp.) (EPPO, 2021b). The adult beetles feed on the ash foliage, but it is the extensive larval feeding in the phloem and cambium that disrupts translocation in the tree, girdling the tree (i.e. removing a ring of bark around the entire circumference of the branch of trunk) and resulting in its death.
Native to north-east China, the Korean peninsula and the east of the Russian Federation, the emerald ash borer has spread to other parts of Asia, North America (Canada and the United States of America) (Haack et al., 2002) and Europe (e.g. the western and southern parts of the Russian Federation, and Ukraine) (CABI, 2021b). It was probably introduced into North America, for example, in 2002 via wood packaging material, and dendrological studies indicate that it arrived on the continent about a decade before its detection. The subsequent spread of the beetle to various parts of the United States of America and Canada was probably facilitated by the movement of infested nursery stock, logs and firewood (Herms and McCullough, 2014; Ramsfield et al., 2016).
The impacts of the beetle are severe. Aukema et al. (2011) considered it to be the most destructive and costly invasive forest insect in the United States of America, with projections of economic losses from this insect up to 2020 exceeding USD 12.5 billion. The invasion of this beetle has also had important implications for biodiversity in the affected areas, as ash trees provide food, shelter and habitat for many species. Furthermore, it has been suggested that the invasion of the emerald ash borer and the consequent loss of trees has possible implications for human health (Donovan et al., 2013). Management strategies have focused on containment, for example by using quarantine zones, and on reducing population densities, such as through the introduction of biological control agents. Eradication was initially attempted and later abandoned (Herms and McCullough, 2014).
The distribution of ash trees is the main limitation on the range of the emerald ash borer, but climate is also thought to play an important role. In its native range, the emerald ash borer occurs in only a fraction of the range of the ash, but modelling by Liang and Fei (2014) has projected that climate change would result in a more northerly distribution of the beetle in North America, and a subsequent long-lasting risk to ash in those areas. However, it is expected that the southward invasion of the emerald ash borer in North America would be limited within a warming climate-change scenario, as the beetle requires strong seasonality with a long winter season. Research by Duan et al. (2020) on the overwintering survival of several introduced larval parasitoid species of the emerald ash borer after an extreme climate event (low winter temperatures) has also shown that extreme climate events associated with climate change could reduce the efficacy of biological control of the beetle.
2. Tephritid fruit flies (global)
Tephritids are a diverse family of insects, with more than 4 000 described species. Most of the species feed on plants and several can cause substantial economic damage, especially when their larvae develop in fruits of high market value. The family contains several invasive species, such as Bactrocera oleae (Figure 7 and Gutierrez et al., 2009), which feeds only on olive trees (and a few wild relatives), Bactrocera dorsalis, which feeds on several dozen fruit plant species, and the Mediterranean fruit fly, Ceratitis capitata, which feeds on a moderate number of tree crops.
Tephritids have been able to expand geographically from their original distribution to colonize both neighbouring areas and new regions owing to the expansion of cultivation of their hosts, international trade, and because climate change has allowed their winter survival and reproduction in habitats otherwise unsuitable for the species. Bactrocera oleae occurs in Africa, Europe and Asia and has invaded California and Mexico more recently (CABI, 2021c). However, Godefroid et al. (2015) concluded that the species may establish not only in the temperate Mediterranean-climate regions but also in the colder climates of northern latitudes in Europe, where olive trees are yet to be cultivated.
Bactrocera dorsalis is a pest of major concern throughout south-east Asia and further west through to Pakistan and north to southern China and Nepal; it has been reported in other areas, including most of Africa, the eastern United States of America, and several islands in the Pacific (EPPO, 2021c). As it has a wide host range, it is being intercepted often in international markets. Because the climatic range of B. dorsalis is primarily tropical and subtropical and it is considered to have rather complex requirements, the risk of direct economic losses from an incursion into temperate areas is low, but climate modification by global warming could allow a rapid increase in fly populations in mild seasons, with the flies spending the winter protected in fruits stored in sheltered places (EPPO, 2021c). This is also the case for C. capitata, which occurs in southern and central Europe, most of Africa and the Near East, Central and South America, and Western Australia, but can overwinter in colder regions as larvae, in fruits stored in warm places. It can spread through the international trade of oranges, mandarins and lemons (Fedchock et al., 2006).
3. Red palm weevil (Rhynchophorus ferrugineus) (Near East, Africa, Europe)
The red palm weevil, Rhynchophorus ferrugineus, is one of the most economically damaging insect pests of palm trees. Native to south-east Asia and Melanesia, larvae of this weevil feed within the apical growing point of the tree, causing extensive damage to the plant tissue, weakening the structure of the plant and in many cases resulting in tree death. In the Gulf region of the Near East, the annual losses due to death and removal of palms severely infested by red palm weevil have been estimated to range from USD 5.2 million to 25.9 million at 1 and 5 percent infestation, respectively (El-Sabea, Faleiro and Abo-El-Saad, 2009). Another estimate has considered the annual losses due to red palm weevil to be USD 15 million (Al-Ayedh, 2017).
The red palm weevil infests various palm species, including coconut and date palm (El-Mergawy and Al-Ajlan, 2011; FAO, 2020). It was first detected on date palms in the Near East in the mid-1980s, and subsequently spread to other countries in the Near East, and to Africa and Europe. In 2010, it was detected in California in the United States of America, where it was declared eradicated in 2015. Its global distribution has probably been assisted by movement of palm offshoots as planting material. Management strategies include the use of various cultural and phytosanitary measures, such as removal of infested trees, the application of insecticides and of nematodes pathogenic to insects, and the use of pheromone traps (FAO, 2020; Ge et al., 2015).
The distribution of the red palm weevil may expand because of climate change. Ge et al. (2015) predicted that in China the number of areas highly favourable to this pest would increase with climate change, resulting in the expansion of the insect into north China. Among the Rhynchophorus species, the red palm weevil is the only one that has significantly expanded its geographical range from its original home in south and south-east Asia (Wattanapongsiri, 1966). It has been reported in 45 countries and ecological-niche modelling predicts that it could expand its range even further (Fiaboe et al., 2012). The red palm weevil is still considered to be the major challenge for palm growers in the Near East, and despite all means of integrated control, its damaging effects are still causing great economic losses.
4. Fall armyworm (Spodoptera frugiperda) (Americas, Africa, Asia)
The fall armyworm (Spodoptera frugiperda) is a moth belonging to the family Noctuidae (Figure 7). It has a host range of hundreds of plant species, inflicting severe damage in grasses – particularly maize and sorghum, which are the preferred hosts – along with other crops, such as rice, cotton and soybean preferred by different species strains. It is native to tropical and-subtropical areas of the Americas and during summer it migrates into southern and northern temperate American regions. The pest was first reported in western Africa in 2016 (Goergen et al. 2016) and then throughout sub-Saharan Africa and Egypt in 2019. In 2018, it was reported in India, rapidly spreading all over southern and eastern Asia including China, the Republic of Korea, Japan and Pakistan. It has also been reported in Bangladesh, Indonesia, Myanmar, Sri Lanka, Thailand, the Philippines, Viet Nam and Yemen (EPPO, 2020a). In 2020, the fall armyworm was first detected on maize in Jordan and the United Arab Emirates (IPPC Secretariat, 2020a; 2020b) and in Israel (EPPO, 2020b). It has also spread across the Australian continent (IPPC Secretariat, 2021).
The fall armyworm is adapted to warm climates and not able to enter diapause, and its geographical distribution is closely dependent on climatic conditions. Adults can travel up to several kilometres in a single night and seasonal migrations can reach Canada from the southern United States of America. Ramirez-Cabral, Kumar and Shabani (2017) have highlighted the expansion of its geographical range in warmer climates because of its adaptability to different environments, its high dispersal capacity, the wide range of potential hosts and the intense international trade of commodities attacked by the larvae or pupae of the moth. They have also predicted a reduction or even partial disappearance of the species in the southern American hemisphere, due to the warmer and drier conditions expected there in the middle or end of this century in the north of the sub-continent. In the European Union, some warm areas in Spain, Italy and Greece could provide suitable climatic conditions for the establishment of the species, mainly from populations established in the North of Africa (Jeger et al., 2018).
5. Desert locust (Schistocerca gregaria) (Africa, western and southern Asia)
The desert locust (Schistocerca gregaria) is found mainly in Africa, through Arabia and western Asia, extending into parts of southern Asia (FAO, 2021a). It has been reported occasionally in south-west Europe. It swarms and voraciously feeds on key crops such as maize and sorghum, pastures, and any green vegetation that comes its way, thereby significantly affecting smallholder farmers and pastoralists (Kimathi et al., 2020).
The desert locust shows periodic changes in its body form and can change over generations, in response to environmental conditions, from a solitary, highly fecund, non-migratory form to a gregarious, migratory phase in which it may travel long distances, finally invading new areas. In general, the desert locust breeds extensively in semi-arid zones, extending from western Africa through the Near East to south-west Asia, threatening the livelihoods of people in over 65 countries. However, there is also a much less well-known subspecies, S. gregaria flaviventris, that occupies a limited area in southern Africa, and the potential of this subspecies to pose a threat in the future should be investigated (Meynard et al., 2017).
Large outbreaks of desert locust have been recorded over many centuries, and the FAO keeps a long-term, large-scale monitoring survey database on affected areas. Identifying the potential breeding sites of the pest is essential if cost-effective and timely preventive measures are to be carried out before the pest inflicts significant damage (Kimathi et al., 2020). Since the 1960s, outbreaks have been less frequent, but in 2019–2020 unprecedented locust breeding was observed in Eritrea, Somalia and Yemen, due to unusually heavy rainfall in the Horn of Africa. The current strategy for managing swarms of the locust is aerial spraying with chemical pesticides, which has high negative impacts on humans, livestock, the environment and biodiversity.
The behaviour, ecology and physiology of the desert locust changes in response to some climatic conditions. Attribution of a single event to climate change is difficult, but climatic changes such as increases in temperature and rainfall over desert areas, and the strong winds associated with tropical cyclones, provide a new favourable environment for pest breeding, development and migration. This suggests that global warming has played a role in providing the conditions required for the development, outbreak and survival of the locust. But the impact of climate change is complex and the FAO’s Commission for Controlling the Desert Locust in south-west Asia (FAO, 2021a) has therefore highlighted the need for international cooperation across affected countries to tackle the locust threat. Where it will fly next depends on wind direction, speed, and other weather parameters. Consequently, climate change may have an impact on future migration routes of the desert locust. The prediction of risk under different climate-change scenarios, however, may need to differentiate between different subspecies, because each of them may have different niche requirements.
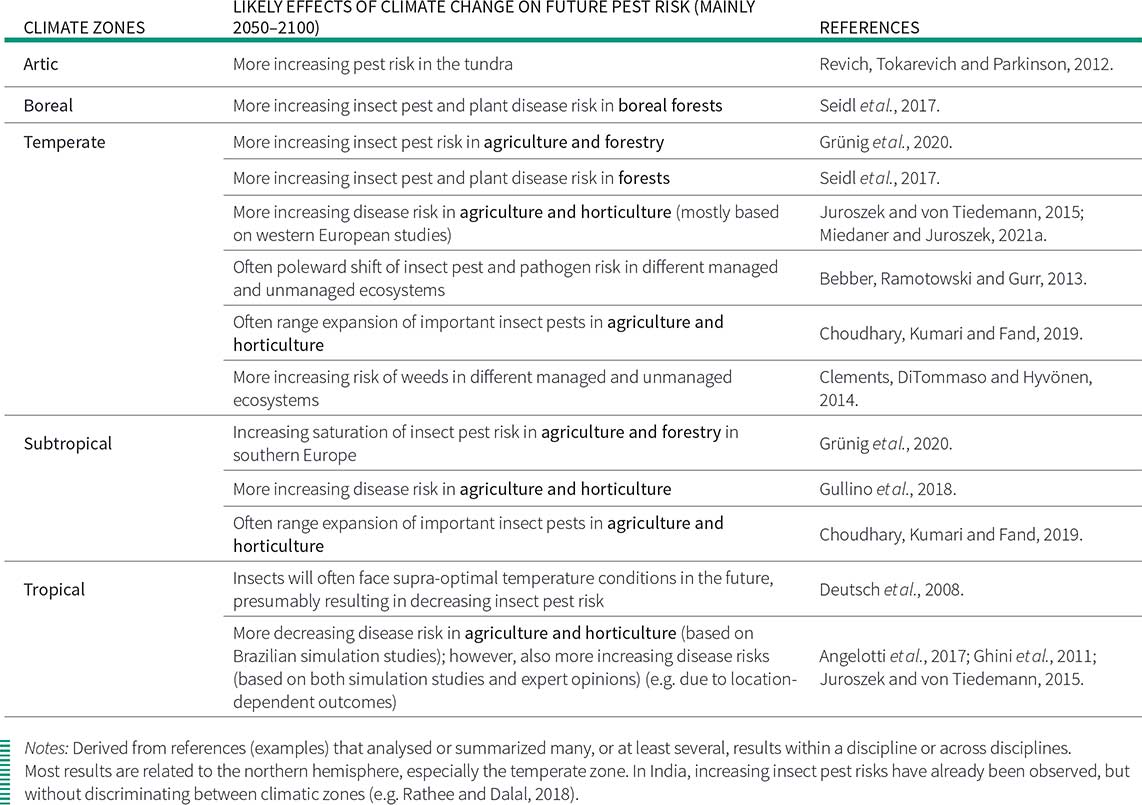
Table 3. Examples of some likely effects of climate change on plant pests (insects, pathogens and weeds) in different climate zones
Figure 7A

(a) Maize ear damaged by larvae; (b) larva and damage in whorl-stage maize.
Figure 7B

Plant pathogens
6. Coffee leaf rust (Hemileia vastatrix) (Africa, Asia, Latin America)
Coffee leaf rust, caused by Hemileia vastatrix, is one of the main factors limiting arabica coffee yield worldwide. Early and highly aggressive outbreaks of the disease have caused serious losses (up to 50–60 percent yield losses) in some Latin American countries, such as Colombia and Mexico, in the past few years.
Climate appears to play a role in the prevalence of the disease. One of the factors that promoted the occurrence of the rust epidemics in Central America was a reduction in the diurnal thermal amplitude, decreasing the latency period of the disease (Avelino et al., 2015). The shorter latency period promotes rapid increase of the pathogen population. Similarly, the pathogen’s incubation period may be reduced with global warming. The analysis by Ghini et al. (2011) on future climate-change scenarios in Brazil indicated a trend of reduction in the incubation period of H. vastatrix, meaning that more generations of this pathogen could develop within a growing season. Consequently, the risk of coffee leaf rust epidemics might increase in the future, unless other factors change to mitigate disease risk, such as a reduced ability of the pathogen to infect coffee plants. Less cold winters can increase the amount of inoculum, in anticipation of pathogen infection (Avelino et al., 2015), but cold temperatures may not present a problem for the pathogen, considering that in Africa the displacement of coffee production to cooler and higher altitude regions has not limited the occurrence of coffee leaf rust because it was already widespread (Iscaro, 2014) and can adapt to different climates (Avelino et al., 2015). Thus, coffee leaf rust has been, and still is, one of the greatest challenges to global coffee production, and it will require the development of new strategies to guarantee its management, particularly if climate change affects the biology of the pathogen in the ways indicated by these studies.
7. Banana Fusarium wilt (Fusarium oxysporum f. sp. cubense) TR4 (Australia, Mozambique, Colombia, Asia, Near East)
The soil-borne fungus Fusarium oxysporum f. sp. cubense causes Fusarium wilt on banana. The planting of ‘Cavendish’, a resistant cultivar, was the solution found for the devastation caused by race 1 of the pathogen (Ploetz, 2005; Stover, 1986). However, a new strain of F. oxysporum, tropical race 4 (TR4), was found in 1990 in eastern Asia, parts of south-east Asia, and northern Australia, attacking ‘Cavendish’ clones in the tropics (Ploetz, 2005; Ploetz and Pegg, 2000). Since 2010, this race has spread to south and south-east Asian countries (India, the Lao People’s Democratic Republic, Myanmar, Pakistan and Viet Nam), the Near East (Israel, Jordan, Lebanon and Oman), Africa (Mozambique) (Dita et al., 2018) and South America (Colombia) (García-Bastidas et al., 2019). This disease is a great threat to ‘Cavendish’ banana growers worldwide, irrespective of whether they farm on a large or small scale (Mostert et al., 2017). High temperatures, such as where temperatures rise from 24 to 34 °C, and extreme environmental events including cyclones and tropical storms, may increase the risk of the disease, particularly when ‘Cavendish’ banana plants suffer waterlogged soil (Pegg et al., 2019; Peng, Sivasithamparam and Turner, 1999). As there are still no banana cultivars resistant to TR4 and chemical control of the pathogen is not effective, preventive measures are the only option to manage the risk of Fusarium wilt disease (TR4). These include, for example, using disease-free ‘Cavendish’ banana planting material, detecting diseased plants early, and destroying diseased plants as soon as Fusarium wilt symptoms are observed (Pegg et al., 2019).
8. Xylella fastidiosa (Americas, southern Europe, Near East)
Xylella fastidiosa – a xylem-limited, Gram-negative bacterium – causes diseases on economically important crops, such as grapevine, citrus, olive, almond, peach and coffee, and in ornamental and forestry plants (Janse and Obradovic, 2010; Wells et al., 1987). It was reported in North and South America and Asia in the 1980s (Cornara et al., 2019). In 2013, X. fastidiosa subsp. pauca was reported on olive trees in southern Italy, causing severe losses and profoundly modifying the landscape typical of the local area by destroying centenary olive trees (Saponari et al., 2013). X. fastidiosa is transmitted by numerous species of sap-sucking hopper insects, including spittlebug and sharpshooter leafhoppers, mainly of the Aphrophoridae and Cicadellidae families (Almeida et al., 2005; Cornara et al., 2019).
Models of species bioclimatic distribution have shown that X. fastidiosa has the potential to expand beyond its current distribution and may reach other areas in Italy and elsewhere in Europe (Bosso et al. 2016; Godefroid et al., 2018). There are different subspecies identified for this bacterium, mainly fastidiosa, multiplex and pauca. According to the modelling predictions, the subspecies multiplex, and to a certain extent the subspecies fastidiosa, represent a threat to most of Europe, while the climatically suitable areas for the subspecies pauca are mostly limited to the Mediterranean countries (Godefroid et al., 2019). Through a predictive risk-ranking model, Frem et al. (2020) recently revealed that the Mediterranean basin, particularly Lebanon, runs the highest risk for establishment and spread of X. fastidiosa. Even though many Mediterranean countries are currently free of X. fastidiosa, in the near future they will be subject to a high risk of X. fastidiosa entry and establishment: Turkey is at the highest risk, followed by Greece, Morocco and Tunisia, which are ranked at high-risk level. Only three countries in the region (Bahrain, Libya and Yemen) are subject to the least risk level in terms of potential entry, establishment and spread of the bacterium. Notably, the problem is not limited to the Mediterranean. On the basis of disease symptoms and laboratory analysis, X. fastidiosa has been found associated with almond leaf scorch and Pierce’s disease in grapevine in several provinces of the Islamic Republic of Iran (Amanifar et al., 2014), indicating that it will start to spread to neighbouring countries in the Near East.
Bosso et al. (2016) have predicted that climate change will not further increase the risk of X. fastidiosa in the future in most of the Mediterranean region, but the complete “host plant–vector–bacterium” relationship should also be considered when predicting future risk. Fortunately, vector performance is likely to suffer due to supra-optimal temperature and suboptimal humidity conditions, as recently simulated by Godefroid et al. (2020).
Management of X. fastidiosa will rely on the development of efficient strategies for integrated pest management, including improvements in detection of the pathogen and insect vectors, agricultural practices, and – last but not least – effective quarantine treatments to control the spread of the pathogen.
9. Oomycetes, including Phytophthora infestans and Plasmopara viticola (global)
The potential displacement of oomycetes poleward because of climate change will present a challenge for plant protection, mainly in the northern hemisphere (Bebber, Ramotowski and Gurr, 2013). Phytophthora infestans, the oomycete that causes late blight in potato and tomato, has a great capacity to adapt to changing conditions, which is an important factor determining the risk of severe epidemics in the future. Indeed, several studies have suggested an increasing risk of P. infestans incidence in several countries (Hannukkala et al., 2007; Perez et al., 2010; Skelsey et al., 2016; Sparks et al., 2014), requiring the development of new strategies to control the disease and reduce its impact on food security, such as postponing the start of the potato growing season (Skelsey et al., 2016; Wu et al., 2020).
Studies in Egypt on the impact of climate change on tomato and potato late blight have shown how warmer winter weather affects their incidence and management (Fahim, Hassanein and Mostafa, 2003; Fahim et al., 2011). These have indicated that an epidemic of late blight on tomatoes occurring one to two weeks earlier would mean that two to three additional sprays would be needed to achieve sufficient control of the disease. Up to three additional fungicide sprays would therefore be needed each growing season in Egypt during the coming decades (2025–2100). As for potato late blight, caused by the same pathogen, comparison of weather conditions and disease occurrence in epidemic versus non-epidemic growing seasons has shown that wet and warm winter seasons promote potato late blight epidemics in Egypt. Favourable conditions in the winter allow a build-up of pathogen inoculum on early cultivars early in the growing season, leading to a tendency for the blight to appear in later-planted potato crops. It can therefore be expected that climate change will promote late blight epidemics in the future. However, there is a great need for further assessments of the impact of climate change on crop diseases in Egypt and other Near East countries (Fahim et al., 2011).
Downy mildew of grapevine, caused by the oomycete Plasmopara viticola, is another serious disease resulting in substantial production losses, varying from 5 percent yield losses up to 30–40 percent in most grape-growing regions. In the case of wine production, downy mildew also affects wine quality. As many of these regions have a temperate climate with temperatures that are suboptimal for the pathogen, an increase in air temperature will favour the occurrence of the disease. Studies considering future climate-change scenarios have therefore projected earlier disease outbreaks that require more treatments to control them (Angelotti et al., 2017; Salinari et al., 2006, 2007). Short-term studies carried out in phytotrons have also confirmed an increased severity of grape downy mildew under simulated climate-change conditions (Pugliese, Gullino and Garibaldi, 2010).
10. Fungi producing mycotoxins (global)
In general, climate change is expected to result in an increased presence of mycotoxins in crops, but the complexity of the fungal flora associated with each crop and its interaction with the environment means that it is difficult to draw conclusions without conducting specific studies. Nevertheless, there are many results available. For instance, the work carried out by Battilani et al. (2016) indicates that global warming could extend the northern limit of aflatoxin risk in maize in Europe, and Van der Fels-Klerx, Liu and Battilani (2016) have made quantitative estimations of the impacts of climate change on mycotoxin occurrence. Medina et al. (2017) reviewed the impacts of climate change on mycotoxigenic fungi, examining the impacts of the three-way interactions between elevated CO2 (350–400 vs 650–1200 ppm), temperature increases (+2–5 °C) and drought stress on growth and mycotoxin production by key spoilage fungi in cereals and nuts, including Alternaria, Aspergillus, Fusarium and Penicillium species. The growth of Aspergillus flavus, responsible for producing aflatoxin B1, appears to be unaffected by simulated climate-change scenarios. However, a significant stimulation of aflatoxin B1 production has been found both in vitro and in vivo in maize. In contrast, the behaviour of other Aspergillus species, responsible for ochratoxin A contamination of a range of commodities, and Fusarium verticillioides, producing fumonisins, suggests that some species are more resilient to climate change than others, especially in terms of mycotoxin production.
In addition to the effects of climate change on these commonly occurring fungi, climate change could also influence the mycotoxin production of emerging pathogens, such as the increases shown experimentally by Siciliano et al. (2017a, 2017b) in Alternaria and Myrothecium species. Furthermore, acclimatization of mycotoxigenic fungal pathogens to climate-change factors may result in increased disease and perhaps mycotoxin contamination of staple cereals as well as other crops. Thus, managing mycotoxin risks will remain a great challenge in the future (Juroszek and von Tiedemann, 2013b) as climate change could worsen the situation (Miedaner and Juroszek, 2021b).
Nematodes
11. Citrus lesion nematode (Pratylenchus coffeae) (global)
The citrus lesion nematode, Pratylenchus coffeae, is widely distributed in citrus orchards worldwide. It mainly infests the plant via the feeder roots, where motile stages of the pest penetrate the cortical tissue. The vascular tissue remains intact until invaded by other organisms in a secondary infection (Duncan, 2009). The nematode is known to reduce citrus root weights by as much as half, and experimental inoculation of young trees has shown growth reductions ranging from 49 to 80 percent, with a three- to 20-fold reduction in the numbers of fruits (O’Bannon and Tomerlin, 1973). Recent studies on current climate change in Egypt indicate that higher temperatures may aggravate the damage caused by the citrus lesion nematode to the citrus root system, because the nematode’s reproduction rate is highest when soil temperatures are relatively high (26–30 °C) (Abd-Elgawad, 2020). At such temperatures, the life cycle is completed in less than one month and the pathogen may reach density levels as high as 10 000 nematodes/g root; the nematode can also survive in roots in the soil for at least four months. Unfortunately, however, commercial rootstocks resistant to this nematode are not available yet (Abd-Elgawad, 2020).
12. Soybean cyst nematode (Heterodera glycines) (global)
The soybean cyst nematode (Heterodera glycines) is the most economically damaging pathogen of soybean (Glycine max) in the United States of America and Canada (Tylka and Marett, 2014). It also causes considerable yield losses in many other major soybean-producing countries, such as Argentina, Brazil and China. Thus, its potential for causing serious worldwide yield loss is high.
Global warming is likely to promote the northward (northern hemisphere) and southward (southern hemisphere) expansion of the nematode’s geographical range, and increase the number of nematode generations per soybean crop-growing season (St-Marseille et al., 2019) until supra-optimal temperature conditions for the nematode are reached.
To manage the pest, the most important strategies are the use of resistant cultivars (Shaibu et al., 2020) and crop rotation (Niblack, 2005). According to Niblack (2005), rotation includes at least three different aspects: ideally, cultivating soybean only once every five years in any one field (although the benefit of crop rotation may be less if weeds are present that can act as alternative hosts of the pest); using non-host crop plants, including cover or catch crops in a wide crop rotation; and planting different resistant or tolerant soybean cultivars in different years in the same field, in order to minimize the adaptation potential of the nematode populations.
13. Pine wilt nematode (Bursaphelenchus xylophilus) (North America and eastern Asia)
According to Jones et al. (2013), the pine wilt nematode, B. xylophilus, is native to North America, where it infests pine trees (Pinus species) but does not seriously damage them. However, in its non-native environment, including Asia (China, Republic of Korea, Japan and others) and Europe (a few occurrences in Portugal and Spain), it is a serious pest, killing millions of pine trees. The nematode is vectored by the adult stage of Monochamus beetles, which fly among pine trees and across longer distances. It is expected that pine wilt disease will be increasingly favoured by global warming because Monochamus beetles, like many other forest insects (Seidl et al., 2017), will benefit from increasing temperatures, particularly in temperate regions (Ikegami and Jenkins, 2018). A number of risk assessments carried out demonstrate that with increasing temperatures in temperate regions the mortality of conifers will increase. In the Mediterranean area, which is the most endangered area in Europe, high mortality of coniferous trees would have serious environmental consequences.
Weeds
14. Butterfly bush (Buddleja davidii) (global)
The geographical range of the invasive weed Buddleja davidii in Europe, North America and New Zealand is projected to expand by the end of the twenty-first century as growth limitations due to cold stress are reduced (Kriticos et al., 2011). In contrast, the range of this weed in Africa, Asia, South America, and Australia is projected to contract because of increased heat stress. Overall, the total area of land with suitable growing conditions for the weed is projected to decrease by 11 percent on average (8, 10 or 16 percent, dependent on the climate-change scenario used). Possible adaptation strategies include the identification of areas of increasing and decreasing invasion threat, so that management resources can be allocated appropriately to reduce further spread of the weed (Kriticos et al., 2011).
15. Serrated tussock grass (Nassella trichotoma) (global)
Under current climate conditions, the grass weed Nassella trichotoma has considerable potential to spread. In future, opportunities to invade new areas that are suitable will continue, but by the end of the twenty-first century, it is projected that the total area suitable will have decreased globally between 20 and 27 percent (depending on the climate-change scenario used), mostly as a result of a projected increase in heat stress (Watt et al., 2011). Possible management strategies include identifying high-risk areas of invasion, applying measures to reduce human-assisted dispersal of seeds, and applying weed-control measures to reduce the wind-borne dispersal of seeds (Watt et al., 2011).