Chapter 4. Environmental, health and socio-economic impacts of soil pollution
Ecosystems impairment caused by soil pollution
Soil pollution affects above- and below-ground biodiversity in two main ways. The number of organisms can be reduced due to the toxicity caused by the contaminants, but also changes can be observed at community level in that tolerant or resistant organisms will benefit compared to those sensitive to the contaminants. It has also been observed that the exposure to some trace elements can induce resistance in soil microorganisms and also promote resistance to antimicrobials (Heydari, 2020). In addition, soil contaminants can enter the food chain and cause disease and mortality in soil-dwelling, terrestrial (including humans) and aquatic organisms. The loss of biodiversity and biomass therefore leads to a decrease in organic matter and changes in nutrient inputs and cycling. This affects the primary productivity of natural and agricultural ecosystems and leads to the overall loss of soil ecosystem services. Additionally, polluted soils in turn become a source of pollution for groundwater, through leaching of contaminants, and for fresh water and the marine environment, because contaminants can be transported off-site through wind and water erosion. These changes can be gradual or remain inert until an inflexion point when severe degradation occurs (Baudrot et al., 2018).
4.2.1. Soil degradation and loss of soil ecosystem services
According to Daily (1997), the conditions and processes provided by ecosystems components (both biotic and abiotic) to sustain and maintain human well-being are defined as ecosystem services. Due to increasing pressures on natural ecosystems, these services must be valued and considered in decision-making processes to ensure their continuity for future generations.
According to the Common International Classification of Ecosystem Services (CICES), three major groups of ecosystem services are provided by soils (Figure 3) (Haines-Young and Potschin, 2018).
Figure 3. Main ecosystem services provided by soils.

Supporting services (e.g. photosynthesis, primary production, formation of soil, nutrient and water cycling) are essential for ecosystem function; however, they are no longer considered in international frameworks due to their intermediate status (Potschin-Young et al., 2017; Smith et al., 2015). These supporting services operate alongside more basic ecological structures and processes to underpin the output of final services (Fisher and Kerry Turner, 2008) and must be also considered when assessing the impact of soil degradation in general, and soil pollution in particular.
Soil ecosystems services are provided by an interaction of abiotic soil components (soil organic carbon, mineral fractions, soil solution, and pore air) and living organisms (from genes to macro-organisms) present in soils. However, global soils are being depleted and degraded by the constant pressures of a growing population that demands more services and benefits (FAO and ITPS, 2015). Soils are also under threat from global challenges, such as by biodiversity loss, imbalances in biogeochemical cycles, chemical pollution, climate change and the hydrogeological cycle, and land use change (Rockström et al., 2009).
Many studies have been conducted on the role of soils in regulating and provisioning services for carbon, nutrients, water and climate (Daily, Matson and Vitousek, 1997; Eswaran, Van Den Berg and Reich, 1993; FAO and ITPS, 2015; Lal, 2016; Seneviratne et al., 2010; Smith et al., 2015; Stevenson and Cole, 1999; Várallyay, 2010). However, the filtering and buffering role of soils, despite being widely understood, is specifically quantified in only a few studies (Aslam et al., 2009; Blum, 2005; Burauel and Baßmann, 2005; Keesstra et al., 2012; Tedoldi et al., 2016; Yan et al., 2019).
Soils have the capacity to retain, buffer, filter and degrade certain contaminants depending on the soil properties and contaminant physicochemical properties. Total contaminant concentrations in soils do not have a direct relationship with the contaminant’s impact on living organisms but instead they are dependent on the contaminant bioavailability. Bioavailability is the amount of a contaminant that can move from the soil and cross cellular membranes and penetrate into living tissues (see Glossary). Freely dissolved forms and aqueous complexes are generally the most bioavailable. When contaminants are not physically or chemically retained in the soil matrix, but are dissolved or suspended in the soil solution, they can directly be absorbed by plant roots and interact with soil organisms. For example, in plants, contaminants can be actively or passively transported into the root cells, although the molecular mechanisms involved are often not fully elucidated. For example, arsenic is absorbed in arsenate form as an analogue of phosphate through P transporters (Danh et al., 2014). Translocation of trace element from roots to shoots is also mediated by transporter proteins (Shahid et al., 2015). These translocation processes may be part of a contaminant management mechanism developed by plants to better tolerate contaminants by storing them in less essential organs and tissues (Čiamporová, 2002; Gupta et al., 2019a).
Bioavailability is the portion of the total quantity of a chemical/substance that is ‘freely available’ to cross an organism’s cellular membrane from the medium the organism inhabits at a given time (Semple et al., 2004), and which can be metabolically active in a living organism (Klaassen, Casarett and Doull, 2013). ‘Freely available’ is defined operationally by different methods. Bioassays using plants or animals (Ng et al., 2015) are often time and resource-consuming and may comprise ethical issues. Indirect single-step or sequential chemical extractions using reagent(s) simulating the interaction of plant or human fluids/exudates with the contaminant(s) of interest are also employed (Cipullo et al., 2018). Some of these extractions have been standardized and taken into consideration in the legislation of some countries. Other extractions are undergoing standardization, a process necessary to make bioavailability a concept that can be included in legislation and policies for soil protection, and as a tool for risk assessment (Harmsen, 2007; Kim et al., 2015). In soil, contaminant bioavailability can be affected by soil properties (e.g., pH, redox potential, clay content, organic matter content, etc.) (Gupta et al., 2019b; Sinche et al., 2018), contaminant chemical properties (e.g., polarity, oxidation states, acidity, etc.), and/or environmental factors (e.g., moisture content, temperature, etc.). These properties are not static in time or place, thus it is important to understand the limitations of operationally-defined determinations of bioavailability when evaluating actual bioavailability at a given site temporally and spatially.
Soil consists of solid, liquid, gaseous and living phases, and therefore soil quality, unlike that of water or air, will depend not only on the existence and impact of pollution in each of these phases, but on the multitude of interactions between phases (Bünemann et al., 2018). Changes in soil properties that control the soil contaminant buffering and filtering capacity can cause a decrease in the protection mechanisms and instead act as a source, potentially releasing contaminants into other environmental compartments (water, air and organisms) (Biswas et al., 2018). Pollution induced soil degradation can become an iterative cycle of degradation (Figure 4) that can ultimately lead to the loss of ecosystem services (Figure 5).
Figure 4. Pollution causes a chain of degradation processes in soil, jeopardizing soil’s ability of providing key ecosystem services. SOC (soil organic carbon).
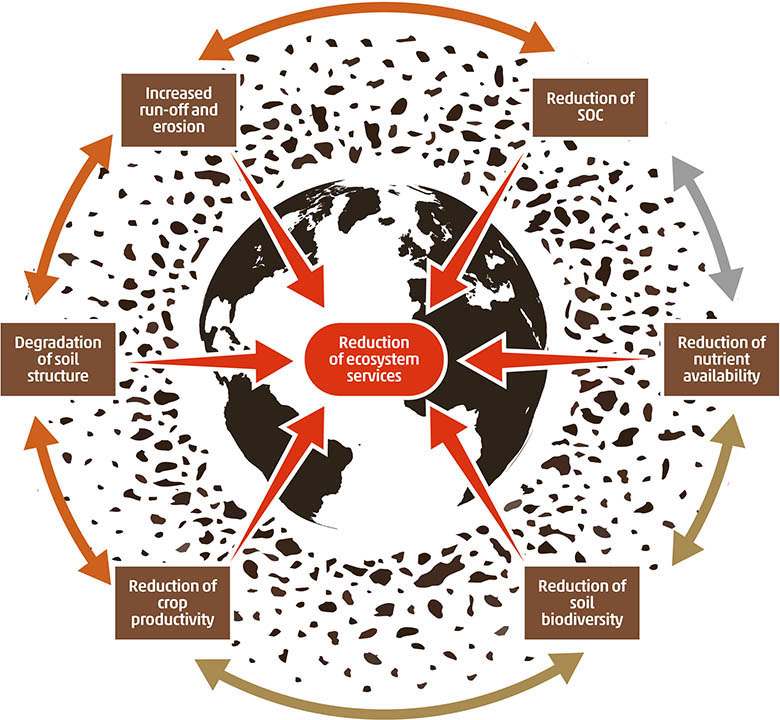
Figure 5. Impacts of soil pollution on soil ecosystem services.

4.2.1.1. Soil pollution as a driver of soil biodiversity loss
Soil biodiversity supports and performs many key ecosystem services and functions, from cycling of nutrients and controlling pests and diseases to building and maintaining soil structure (Wall et al., 2012). Consequently the loss of soil biodiversity or the alteration of soil communities will lead to the reduction of other ecosystems services.
Life on earth has been exposed to contaminants and toxic compounds, such as trace elements, natural radioactivity, ultraviolet light, microbial toxins and defensive compounds from plants and microorganisms, and has evolved to adapt to local conditions over millions of years. However, anthropogenic activities have released a new set of synthetic contaminants that have dispersed and accumulated putting great pressure on the evolution of populations. While some species are able to evolve relatively quickly- such as the emergence of weed resistance to herbicides or microbial resistance to antibiotics- others are not able to adapt or develop tolerance in a short period and can disappear from affected areas (Brady et al., 2017).
One of the first observed impacts of soil pollution are changes in above- and below-ground biodiversity (Carson, 1962). In general, soil pollution has clear impacts at the organism level, but is less evident at population level. Soil pollution may lead to changes in ecosystem structure and species distribution due to the replacement of the more sensitive species with more tolerant types. Low concentrations of soil contaminants and long-term exposure frequently lead to adaptation strategies through changes in physiology and feeding behaviours (Medina, Correa and Barata, 2007), but extreme soil pollution can lead to the effective loss of biodiversity above- and below-ground (Abbasian et al., 2016; Hernández and Pastor, 2008).
Although most of the literature on the relationship between soil pollution and soil organisms and plants has focused on the use of bacteria, fungi and plants to degrade or immobilize contaminants, studies on the impact of contaminants on soil biodiversity are reported (Boteva et al., 2016; Müller et al., 2002; Oliveira and Pampulha, 2006; Rajapaksha, Tobor-Kapłon and Bååth, 2004; Tyler et al., 1989). Soil contaminants can cause reduced biomass, growth and reproductive capacity and result in physiological changes that may be lethal for some soil-dwelling organisms and plants (Freedman et al., 1990; Šalamún et al., 2017). Sensitivity and resistance mechanisms, including avoidance, exclusion, immobilization, excretion, and enzymatic degradation vary widely among species and depend on the type and concentration of soil contaminants (Tyler et al., 1989), but all contribute to the changes in the ecological structure of a polluted soil environment. There is a lack of consistent information about long-term effects of soil pollution exposure on soil communities, especially for microbial communities, which varies according to the pollution gradient, time of exposure, edaphoclimatic conditions and type of vegetation (FAO et al., 2020). To develop the comprehensive knowledge needed to make informed-decisions, we will require a harmonized methodology for long-term monitoring soil communities in polluted soils (Liu et al., 2012b; Loria, Cristescu and Gonzalez, 2019).
Impact on soil microorganisms
Soil microorganisms include bacteria, archaea and fungi and represent the major biomass of soil biodiversity and are also involved in the provision of many key soil ecosystem services (FAO et al., 2020). Soil contaminants may reduce the biomass and certain enzymatic activities of soil microorganisms, inducing changes in soil communities (Kenarova et al., 2014; Smejkalova, Mikanova and Boruvka, 2003; Wang et al., 2007; Zhang et al., 2015).
Soil microbial communities are resilient to some extent to moderate levels of soil pollution due to their great diversity and complex interactions (Ventorino et al., 2018). This acquired resilience has been confirmed for functional groups (Brandt et al., 2010). However, analytical techniques are only now becoming able to reliably report resilience at the species level, so uncertainty is still high in this regard (Azarbad et al., 2015).
Some organisms show the capacity to tolerate soil contaminants, and some organic contaminants can even be used as a source of carbon and energy, with adapted communities proliferating over other more sensitive organisms (Boivin et al., 2002; Musilova et al., 2016). Ultimately, this may lead to an imbalance in functional diversity that may jeopardize other ecosystem services such as carbon and nutrient cycling. Soil microbial communities are characterized by a high level of functional redundancy and, therefore, although some species may be more severely affected by pollution, others may continue to perform without major changes in nutrient and carbon cycling in the soil (FAO et al., 2020). Changes in functional diversity, and hence imbalances in nutrient cycling, are only expected in case of severe toxicity or absence of communities redundancy (Boivin et al., 2002). Disturbed ecosystems such as agricultural or industrial soils where soil biodiversity has already been reduced before the occurrence of pollution are likely to be more prone to nutrient imbalances and more extensive changes in soil communities.
Genetic analysis through Illumina sequencing of 16s rRNA or phospholipid fatty acid (PLFA) analysis have sufficient resolution to differentiate changes in microbial communities. Using these techniques, Konopka et al. (1999), Liu et al. (2012b), Stemmer et al. (2007), and Ventorino et al. (2018), among others, have demonstrated shifts in microbial communities under various levels of stress caused by trace element and organic pollution. Müller et al. (2002) found significant changes in species diversity in a mercury-polluted soil, but no changes were observed in functional diversity, and that key biogeochemical functions (cycling of nutrients, degradation of organic matter, etc.) were carried out by the populations that replaced the pre-existing ones. Similarly, Li et al. (2017c) reported an increase in the cooperative interactions among bacterial and archaeal communities as an adaptive mechanism to soil pollution, which would mask the negative effects.
Changes in enzymatic activities of soil bacteria and fungi have also been widely reported in response to soil pollution. Soil microorganisms show the capacity to increase the production of phosphatase enzyme for the phosphorylation of trace elements and other contaminants, such as radionuclides and petroleum hydrocarbons, as a defence mechanism, while dehydrogenase activity is reduced (Boteva et al., 2016; Chao et al., 2016; Oliveira and Pampulha, 2006; Smejkalova, Mikanova and Boruvka, 2003; Zhang et al., 2015). As a result of this defensive strategy, carbon consumption is mostly directed towards the creation of these phosphorylated ligands and not to the production of organic and inorganic compounds that contribute to cell growth and division, thus increasing the basal respiration (measured by the release of CO2) without the development of microbial communities (Boteva et al., 2016; Smejkalova, Mikanova and Boruvka, 2003). Dehydrogenase activity is one of the most important enzymatic groups in soil, and are involved in the cycling of soil organic matter and thus also influencing the redox of the soil system. Dehydrogenase activity is noted as a good bioindicator of soil pollution due to inhibition by contaminants such as trace elements, fertilizers and pesticides (Wolińska and Stępniewska, 2012). Trace elements pollution can lead therefore to the shift in community structure towards trace-element resistant microbial populations (Gall, Boyd and Rajakaruna, 2015). In addition, nitrification enzymes are also good indicators of the effects of pesticides on soil microorganisms, since the response to the negative effects of contaminants is relatively strong because only a few species are involved in the nitrification processes (Jacobsen and Hjelmsø, 2014).
Changes in microbial activity lead to a lower decomposition rate of litter and fresh organic matter, affecting the biogeochemical cycles in the polluted areas (Kolesnikov et al., 2010; Stemmer et al., 2007; Zhang et al., 2015). The presence of contaminant mixtures can also have a significant effect on microbial populations as reported by Monteiro and Boxall (2009), who described the impact of sulphonamide-type veterinary drug on the degradation activity of other pharmaceuticals by soil microorganisms. However, very little published research on this exists.
Fungal communities are significantly affected by trace element pollution, although responses differ from those of bacteria because fungi are also dependent on associations with dominant vegetation (Bell et al., 2014; Rajapaksha, Tobor-Kapłon and Bååth, 2004). Soil fungi show several pollution tolerance mechanisms, such as absorbing contaminants into the cell walls, producing extracellular polysaccharides for immobilisation, or increasing spore production (Henderson et al., 2015). Several studies have reported an increase in hyphae and spore production by arbuscular mycorrhizal fungi as a mechanism of stress alleviation in polluted soils (Gao et al., 2011; Khade and Adholeya, 2009; Krishnamoorthy et al., 2015), but others have observed a reduction in biomass and diversity of fungi under long-term trace elements pollution with a significant shift from fungal to bacterial communities (Cotrufo et al., 1995; Liu et al., 2012b; Oliveira and Pampulha, 2006). The decrease in fungal mycelia and biomass leads to lower rates of litter decomposition than observed in uncontaminated soils (Cotrufo et al., 1995).
Certain organic contaminants, such as petroleum hydrocarbons, are frequently used by bacteria, fungi and cyanobacteria as a source of carbon and nutrients, and are degraded into simpler molecules (Chen et al., 2020; Vijayan and Abdulhameed, 2020). This degradation potential has been used for remediating polluted soils through the process known as bioremediation (see Chapter 7). However, not all microorganisms respond in the same way to the contaminants and the response depends on the soil conditions (availability of other available sources of carbon and nutrients), and the level of pollution (Bell et al., 2014; Chen et al., 2020; Kolesnikov et al., 2010). Petroleum hydrocarbons can also saturate soil pores and reduce the availability of oxygen and water, affecting plant roots, soil fauna and microorganisms (Figure 6) (Zhang et al., 2015). Despite a typical initial positive effect of the hydrocarbons as a source of carbon as confirmed by an increase in basal respiration; in general, bacterial and fungal activity is inhibited in soils severely polluted by crude oil (Bell et al., 2014; Zhang et al., 2015). Shifts in community structure are also observed, with a predominance of hydrocarbon-degrading species over nitrogen-cycle organisms (Liao et al., 2015). When the more labile contaminant forms have been reduced, only fungal communities with capacity to degrade recalcitrant forms are found in hydrocarbon polluted soils. Microorganisms rapidly consume nutrients released after the first stages of hydrocarbon degradation (Zhang et al., 2015), so at the medium and long term phases when release of nutrients is significantly reduced, carbon and nutrient concentrations in hydrocarbon polluted soils significantly decrease.
Figure 6. Oil spill in Peru. Oil collapse soil pores and impede life on soil. ©Achim Constantin.

The impact of pesticides and antibiotics on soil microorganisms is also important. Plant protection products (PPPs) have diverse effects on enzymatic activities, biomass and diversity of soil-dwelling organisms, varying from positive effects such as increasing certain enzyme activities, to dramatic reduction of microbial biomass, depending on the type of PPP (FAO and ITPS, 2017; Lo, 2010). One of the main and most significant effects of PPPs is the change in the microbial community structure. A loss of functionality may not necessarily occur, but a exposure may result in a shift to more resistant species or species with adaptation strategies for improved tolerance to conditions imposed by PPPs (FAO and ITPS, 2017). Megharaj et al. (2000), for example, observed significant changes in species and taxa in long-term polluted soils affected by different concentrations of DDT, in that algae species showed less tolerance to pollution than some species of fungi, which experienced major growth in medium-level polluted soils.
Studies on emerging contaminants and the impact on soil biodiversity are increasingly being published (Gomes et al., 2017). Lately, many researchers have focused on the role of antibiotics, antibiotic residues, antimicrobial resistant (AMR) genes and bacteria released in the soil environment due to agricultural and livestock practices (for example, use of sewage sludge as soil amendment and fertilizers, application of fresh manure, or irrigation with wastewater). As pointed out by Cycoń, Mrozik and Piotrowska-Seget (2019), antibiotic concentrations in soils range from “a few nanograms to milligrams per kg of soil”. Antibiotic half-life in soils varies from hours (amoxicillin) to more than nine years (ciprofloxacin) depending on the structure, physicochemical properties, and sorption binding mechanism to organic and mineral soil components (Cycoń, Mrozik and Piotrowska-Seget, 2019). Even low concentrations of antibiotics in soils (e.g. 0.2 mg/kg d.w. soil of ciprofloxacin inhibited respiration; 0.01 mg/ kg d.w. soil of Monensin inhibited nitrogen transformations) can lead to changes in enzymatic activities and community structure of soil microorganisms (Cycoń, Mrozik and Piotrowska-Seget, 2019). In soils with significant antibiotic residues (100 mg/kg or higher concentrations of ciprofloxacin, chlortetracycline, sulfadiazine, sulfamethoxazole), AMR bacteria will be selected over sensitive species (Fang et al., 2016; Liu et al., 2014, 2015, 2012a; Ma et al., 2014; Xu et al., 2016). Bacteria show high potential for horizontal transference1 of AMR genes, which leads to changes in the whole microbial community (Rodríguez Eugenio, McLaughlin and Pennock, 2018). Additionally, antibiotics may have similar effects of those caused by pesticides and other organic contaminants, reducing certain microbial activities and producing shifts in community structure and functionality (Cycoń, Mrozik and Piotrowska-Seget, 2019). Soil fungi usually benefit in the presence of antibiotics, being less sensitive than bacteria. Other emerging contaminants have been studied to a lesser extent and ecotoxicology effects are frequently assessed in laboratory conditions at concentrations much higher than those observed in the field (Gomes et al., 2017; Rillig and Bonkowski, 2018; Zhu et al., 2019). This limited data is available on the impact on soil organisms with more research needed.
According to the current knowledge, soil microorganisms are relatively sensitive to soil pollution and are good bioindicators of the presence and level of soil pollution. Soil microbial parameters are good indicators to be included in accurate risk assessment approaches as they are easier and less costly to analyse in the laboratory than the total and available content of many contaminants. Three parameters are needed for bioindicators to clearly indicate the impacts of soil pollution: microbial biomass, microbial activity (both respiration and enzymatic activity), and diversity of soil microbial communities (Gómez-Sagasti et al., 2012). It is important to note that microbial respiration is strongly dependent on environmental conditions, so it is recommended to take samples at the same time of the year and compare them with uncontaminated sites under the same conditions. Different contaminants affect specific enzymatic activities in different ways and hence the bioindicators should be selected in a case-by-case basis depending on the contaminants and their concentrations (Tang et al., 2019). For example, denitrifying enzymes and arylsulfatase have been observed to be particularly sensitive to the presence of trace elements (Utobo and Tewari, 2015), while dehydrogenases, acid phosphatase, alkaline phosphatase, and proteases are inhibited by PAHs (Bielińska et al., 2018). These parameters are also useful for evaluation of the success of remediation strategies (Shen et al., 2016).
Microplastics behave differently than trace elements and organic contaminants, as they can adsorb other contaminants but can also represent a microhabitat for soil microorganisms (Lu et al., 2019). Significant knowledge is emerging in the last years regarding the interactions and impacts of microplastics with soil microorganisms, but many uncertainties still remain and effects will depend on the type of plastic, the concentration in soils and the accompanying contaminants (Wiedner and Polifka, 2020; Yu et al., 2020). Yu and co-workers also suggest a competency between microplastics and soil microorganism for soil aggregate spaces, showing inhibition of microbial activities as well biomass due to a lower access to SOM in presence of microplastics (Yu et al., 2020)
Impacts on soil meso- and macroorganisms
The soil meso- and macroorganism categories (henceforth referred as collectively as macroorganisms) include those organisms from 100 micrometres to 2 millimetres (mesoorganisms), such as tardigrades, mites or collembolans, and from 2 mm to several centimetres (macroorganisms), such as earthworms, termites, ants, millipedes or woodlice (FAO et al., 2020). These species spend their entire life or part of their development cycle in the soil. Soil macroorganisms are key species for litter decomposition and nutrient cycling. Their detection and analysis are easier than microorganisms, and the percentage of known diversity is also higher. Soil-dwelling macroorganisms are also exposed to contaminants in soils and have therefore been frequently used as bioindicators of soil quality (Manu, 2017).
Soil macroorganisms present significant differences in sensitivity and habit among groups and species (van Straalen, 2004). Within the same ecological class, different metabolic and detoxification mechanisms may exist, which influences sensitivity. Soil organism sensitivity will also be conditioned by the type of contaminants, the bioavailability and the period of exposure to the source of pollution. For example, enchytraeids (potworms) are more sensitive to organic contaminants (specifically to herbicides) than earthworms, while earthworms accumulate higher concentrations of PAHs than arthropods (FAO and ITPS, 2017). Nematodes communities have been reported as sensitive and respond to the presence of copper in soil (van Straalen, 2004). Juveniles may be more sensitive than adults because body surface area – volume ratio is higher (Casquero et al., 2020).
The frequent direct effects of soil contaminants on soil macroorganisms are reduced growth and reproduction rate, shortened life cycle and mortality (Manu, 2017). Effects on prey-predator equilibria or changes on soil ecosystem properties also induce changes to soil macroorganisms communities.
Mobile soil fauna frequently use an avoidance strategy, reducing the first steps of litter breakdown and decomposition in highly polluted soils (Cuvillier-Hot et al., 2014). Avoidance of the heavily-polluted upper layer of soils has been frequently reported, leading to the accumulation of undecomposed litter and a reduction in burrowing in the first centimetres of the soil profile, where contaminants tend to accumulate (Figure 7) (Mombo et al., 2018). An inverse correlation between the invertebrates’ feeding activity and the distance to the source of soil contaminants has been observed (Filzek et al., 2004), with a sharp decrease in macroinvertebrate populations with distance from the source of pollution (Gillet and Ponge, 2002; Goix et al., 2015; Korkina and Vorobeichik, 2018). Simple techniques to assess changes in macroorganisms feeding rate such as bait-lamina-test or litterbag test have been successfully applied in ecotoxicological analysis and risk assessment studies (Boshoff et al., 2014; Kula and Römbke, 1998; Niemeyer et al., 2018). To better understand the impact of soil pollution on soil macroorganisms, indexes of species richness and diversity, number of individuals among species, changes in body size and in the structure of food webs, are also frequently used (van Straalen, 2004). However, some studies have reported an increase in surface soil macrofauna diversity in trace element-polluted soils, due to the reduction in microorganisms involved in litter decomposition and the consequent incremental increase in litter accumulation (Nahmani and Lavelle, 2002). Macroorganisms with a certain tolerance to soil contaminants frequently show an increase in abundance although the overall richness of the soil faunal community decreases (Fiera, 2009; Nahmani and Lavelle, 2002).
Figure 7. Soil meso and macrofauna frequently use avoidance strategies in polluted soils, where contaminated plant litter accumulates.

Avoidance strategies are relevant to assess risk when referring to point source pollution or soils affected by low mobility contaminants. However, little research has been conducted on the impact of diffuse pollution affecting large areas on soil fauna. Diffuse pollution is characterized by a high spatial variability of a mixture of contaminants, and heterogeneous exposure of soil-dwelling organisms to low or moderate contaminant concentration over long periods (van Straalen and van Gestel, 2008). Few studies have statistically quantified the impact of diffuse pollution on soil communities and changes in soil ecosystems (Kula and Römbke, 1998; van Straalen and van Gestel, 2008).
Ecotoxicology assessment frequently uses laboratory or mesocosm experimental data, in which soil macroorganisms are exposed to contaminants at higher concentrations than environmentally relevant for longer periods than expected in natural situations (Casquero et al., 2020; Kwak et al., 2017; Marinussen and van der Zee, 1996; Posthuma and Suter, 2011). This approach limits our understanding of the impact of contaminants present in soils on soil macrofauna. Kwak and co-workers (2017) conducted a review of the ecotoxicological effects on soil organisms of seven organic contaminants with known endocrine disrupting effects and identified significant gaps in knowledge due to the establishment of experiments using extremely high concentrations of contaminants unrelated to the concentrations normally identified in the environment, and the use of a limited number of species (Kwak et al., 2017). Kwak and co-workers (2020) also established a multispecies test to assess the impacts of perfluorooctanoic acid (PFOA), which has endocrine disrupting effects (Kwak et al., 2020). The authors observed chronic and acute responses at high concentrations but low risk at environmentally relevant concentrations (Kwak et al., 2020). Another reason why laboratory-exposed organisms could be more sensitive to contaminants is due to the lack of adaptation time to develop defence mechanisms and also because the contaminants have not undergone disturbance and degradation processes in the soil if applied fresh to a clean test soil. Laboratory studies are also run under conditions (usually warm and humid) which are optimal for organism survival, growth and reproduction and which also might be optimal conditions for contaminant bioavailability.
Few large field studies have focused on the effect of radionuclides on soil meso- and macrofauna. Radionuclides have a long persistence in soils and move down the soil profile facilitated by burrowing organisms. The release of radioactivity due to isotope decay damages the DNA and egg development and hatching, and can burn the skin of soil organisms (Zaitsev et al., 2014). Earthworms are especially sensitive to radionuclide pollution. For example, Zaitsev et al. (2014) reported changes in the entire soil food web, over different time periods, depending on feeding and living strategies, and radionuclide migration in the soil matrix. Recovery of soil biodiversity has not been observed until 25-40 years after a nuclear accident or deliberate release of radionuclides in moderately affected areas, whereas in areas severely affected a full recovery of the soil communities is not evident (Maksimova, 2005; Zaitsev et al., 2014). Belowground communities, in general, are affected for longer periods by soil contaminants than aboveground fauna and plants, and particularly by radionuclides, due to migration to deeper layers and longer persistence, due to adsorption by soil minerals and organic matter or entrapment within the soil matrix.
Not only are soil macroorganisms affected by direct exposure or ingestion of contaminants, but effects have also been observed in the gut microbiota of soil-dwelling organisms and these can be a more sensitive indicator of changes produced by contaminants than mortality or physical changes (Ma et al., 2020; Wang et al., 2019c). Xiang et al. (2019) observed significant changes in gut microbiota of collembola grown in a microplastic-polluted environment, including the predominance of biofilm-forming bacteria. When microplastics were combined with antibiotics, further changes in gut microbiota were observed. In both cases, the proliferation of antimicrobial resistant genes strongly related to changes in gut microbiota, with different resistant patterns depending on the presence or absence of antibiotic in the soil (Xiang et al., 2019). Similar increases of antimicrobial resistant genes have been observed in gut microbiota of meso- and macroorganisms exposed to other contaminants, such as trace elements (Hu et al., 2017; Li, Xia and Zhang, 2017; Wang et al., 2019a).
Impacts on soil megafauna
Soil megafauna refers to vertebrates that inhabit the soil environment (mammals, reptiles, and amphibians), and which are usually longer than 10 centimetres. These animals are mostly diggers and are adapted to life underground (Orgiazzi et al., 2016). They play a key role in redistributing soil organic matter within the soil profile and in building soil structure and porosity.
Soil vertebrates are higher on the soil food web. They can be dietary generalists or omnivorous, or more specialized, feeding on insects, other small vertebrates, and earthworms or plant roots and tubers. They can therefore accumulate higher concentrations of soil contaminants than their prey or food sources (De Lange et al., 2009). Carnivores, insectivores and omnivores are more prone to accumulation of soil contaminants than herbivores (Baudrot et al., 2018; Wijnhoven et al., 2007). It has also been observed that soil vertebrates change their feeding behaviour when soil pollution occurs. As reported in previous paragraphs, changes in soil communities and species richness is a consequence of the presence of soil contaminants, and hence, organisms at higher trophic levels have to adapt to food availability (Baudrot et al., 2018). In addition, vertebrates may have a selective behaviour depending on the chemical composition of their food. Ozaki et al. (2018) reported a decreased consumption of tolerant plants by wood mouse in trace element-polluted soils. Mammals are also exposed to soil contaminants through placental transference and breast milk (Smith et al., 2007).
Vertebrates are exposed to soil contaminants for longer periods due to longer life expectancy than micro and macroorganisms, resulting in a greater vulnerability than short-lived organisms, especially to persistent contaminants (Rowe, 2008). Additionally, soil-dwelling vertebrates can be exposed to soil contaminants through dermal contact, which is especially significant for amphibians and reptiles; and inhalation, particularly important for tunnelling organisms that mobilizes large amounts of soil particles in a poorly ventilated environment. However, data on these two exposure pathways are very limited (Smith et al., 2007). Vertebrates with low dispersive capacity and territorial behaviour are more prone to negative impacts of soil pollution than organisms that are more opportunistic and have high reproductive and dispersive capacities (De Lange et al., 2009).
Fate and toxicity greatly vary among and within species depending on physiological and development status once contaminants enter their bodies (Baudrot et al., 2018; Zarrintab and Mirzaei, 2017). Similarly to lower trophic levels, data on the toxicological effects of contaminants on soil vertebrates is limited and the majority of studies have focused on the body burden of contaminants (trace elements, radionuclides and organic contaminants) on different species (Beresford et al., 2020; Ishiniwa et al., 2019; Roumak et al., 2018; Wijnhoven et al., 2007). Frequently, trace elements are found predominantly in liver and kidneys, but fur and hair have also shown high concentrations (Haskins, Gogal and Tuberville, 2020; Zarrintab and Mirzaei, 2017). Pathologies related to inorganic and organic contaminants, are highly variable, causing a wide range of acute and sub-acute toxicity. For example, dioxin-like compounds cause alteration of the immune system, disruption of the thyroid activity, reduced reproduction capacity, neurological and growth retardation in vertebrates (White and Birnbaum, 2009); organophosphate pesticides are endocrine disruptors and have been related to alterations of the immune system, endocrine disruption, oxidative stress, reproductive toxicity and birth defects in mammals (Garcês, Pires and Rodrigues, 2020; Mitra and Maitra, 2018). Reproductive toxicity has also been reported for many other endocrine disruptors in different organisms (Bókony et al., 2018; Godfray et al., 2019; Verderame and Limatola, 2015). Phthalates and Bisphenol A are plastic additives with estrogenic activity, which cause an alteration of the endocrine system in vertebrates and some invertebrates, promoting a feminization of the populations (de Souza Machado et al., 2018a). Although these chemicals strongly interact with the soil matrix and are rapidly eliminated from the organism in vertebrates (Harrison and Hester, 2018), the ubiquitous presence in the environment has been demonstrated to cause reproductive alterations in terrestrial and aquatic organisms (Colborn, vom Saal and Soto, 1993; Kanda, 2019).
Evolutionary toxicology addresses the continuous exposure to contaminants over the lifetime of wildlife, and how this affects natural populations (Brady et al., 2017). The number of scientific papers on this subject has escalated since the late 1990s (according to Scopus search results). Toxicological effects on vertebrates are poorly understood, but changes observed range from alterations in enzymatic activities (Smits, Sanders and Charlebois, 2019) to genotoxic effects that lead to changes in population diversity and structure (Quina et al., 2019). Genetic changes can lead to a reduction of reproductive capacity and predominance of malformed embryos with a reduction in population size. DNA repair mechanisms can prevent such damage from being passed on from one generation to the next and allow the less affected organisms to reproduce more easily due to a reduced intra-specific competition. This results in the predominance of certain phenotypes and produces changes in genetic variability of the whole population (Obiakor et al., 2019).
4.2.1.2. Soil pollution as a driver of loss of soil organic carbon and nutrients
The carbon cycle starts in the soil compartment with the decay of litter and accumulation of dead organisms and organic residues (faeces, cast, eggshell, etc.) (Figure 8). The organic matter in the soil depends on the balance between primary productivity of terrestrial vegetation and other organic inputs, and the decomposition rates by soil organisms and mineralization by abiotic edaphoclimatic processes. Edaphoclimatic conditions2 are independent of soil pollution, and higher rates of organic matter storage in soils are observed under cold-wet climate and with higher contents of clay minerals, Fe/Al-oxides and no limitations in essential nutrients (N, P, Ca, Mg) (Post, 1993). Primary productivity and decomposer activity, however, are directly influenced by the presence of contaminants in soil.
Figure 8. Carbon cycle in soils.
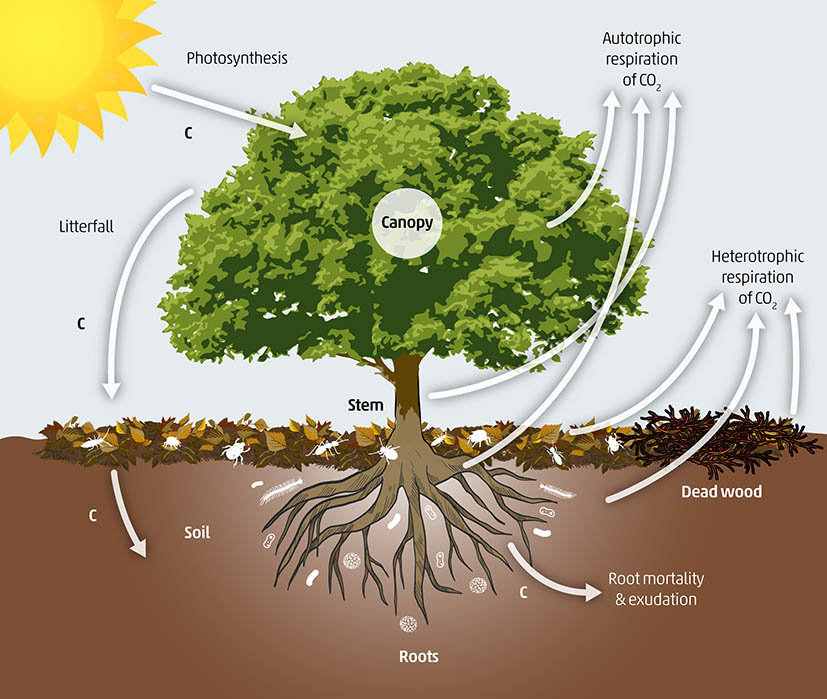
As mentioned in the previous paragraphs, heavily polluted soils show a significant reduction in plant biomass and changes in soil biodiversity, with consequent decrease in litter input from the soil surface, and breakdown and mineralization of organic matter. The carbon efficiency use decreases in polluted soils in comparison to unpolluted soils (Xu et al., 2018). In soils polluted with organic contaminants, despite an initial spike of microbial respiration due to the use of the contaminants as source of carbon, a rapid decrease in microbial activity and carbon and nitrogen use is detected due to inhibition of certain microbial enzymatic activities (Liang et al., 2016). Uptake of soil nutrients by plants is hampered by trace element contamination (Saha et al., 2017).
Litter decomposition can be reduced by 10 to 80 percent in severely polluted soil compared to uncontaminated soil (Kozlov and Zvereva, 2015 and references therein). Cotrufo and co-workers (1995) observed an 11 percent difference in litter mass loss between the trace element-polluted oak forest in Naples, Italy, and an uncontaminated control site outside the city (Cotrufo et al., 1995). Similar results were observed by Freitas and co-workers (2014), who observed a significant difference in litter decomposition rates between heavily polluted soils and unpolluted soils (Freitas et al., 2014). For slightly polluted soils, however, differences are not significant as environmental conditions play a major role on enhancing/inhibiting microbial decomposition activity.
The decrease in litter decomposition can also be observed when comparing soil organic matter fractions from polluted and unpolluted soils. Polluted soils show higher accumulation of fresher residues (fulvic acids with higher C/N ratio) in the larger SOC fraction (2 000 – 250 µm) than unpolluted soils (Zhou et al., 2018). Litter accumulates in highly polluted soils, in which no or very little detritivore activity is observed on organic topsoil (Freitas et al., 2014; Gillet and Ponge, 2002; Russell and Alberti, 1998). Korkina and Vorobeichik (2016, 2018) found significant differences on litter accumulation and zoogenic litter transformation on trace elements-polluted soils close to a smelter, with a clear impact along a pollution gradient. Kozlov and Zvereva (2015) observed a significant decline in saprotrophic bacteria and fungi associated with lower rates of litter decomposition in industrially polluted sites. A decrease in litter decomposition activity has also been reported in radionuclide-polluted soils near Chernobyl, where high levels of radiation lead to the accumulation of undecomposed litter (Mousseau et al., 2014). Higher C/N and C/P ratios are also expected in organic fractions of highly polluted soils and a lower decomposition rate of cellulose, as a signal of reduced mineralization (Li et al., 2017c; Vorobeichik and Pishchulin, 2011; Zhang et al., 2019; Zhou et al., 2018). Several studies indicate a lower plant N utilization in polluted soils with the lower nitrogen content in litter also impacting the biomass and activity of decomposers (Kozlov and Zvereva, 2015; Zhang et al., 2019; Zhou et al., 2018).
Additionally, the uptake and accumulation of soil contaminants into plant tissues, although differing among contaminants, alters litter quality (Lucisine et al., 2015). The presence of contaminants in litter and wood can lead to avoidance by decomposers, contributing to the accumulation of undecomposed organic matter on the surface and reducing the incorporation of soil organic carbon and nutrients within the soil profile (Gillet and Ponge, 2002; Zhou et al., 2018). The decomposition and mineralisation of contaminated litter will contribute to the cycling and redistribution of contaminants within the soil profile (Schroth et al., 2008; Sizmur et al., 2011; Udovic and Lestan, 2007).
4.2.1.3. Soil pollution as a driver of degradation of soil structure and increased soil erodibility
Degradation of soil physical status can occur due to the direct impact of infrastructure and human activities (compaction and sealing). The negative impact of soil contaminants on the diversity, activity and biomass of soil organisms has been mentioned in the previous sections. Since soil organisms are the main architects of soil structure and SOC is the major responsible for soil aggregates stability, it is clear that soil pollution leading to the loss of soil organic matter and reduced presence and activity of soil organisms will result in degraded physical structure and higher soil erodibility (Korkina and Vorobeichik, 2018).
But can contaminants per se cause changes in the physical properties of soils? Most of the existing literature refers to the influence of soil physical properties on the retention and bioavailability of contaminants in soil but very little information has been found on the opposite process. Nevertheless, some studies reveal that the physical structure of the soil can be degraded by the presence of certain contaminants, such as microplastics, nanomaterials or crude oils. These contaminants occupy soil pore space, alter bulk density and, if hydrophobic, will prevent water and air permeability. The changes in physical properties will vary depending on the contaminant, for example, crude oils increase soil bulk density while microplastics reduce it (Klamerus-Iwan et al., 2015; de Souza Machado et al., 2018b).
Some studies have reported the impact of certain soil contaminants on aggregates distribution and stability. Trace element pollution decrease the aggregates mean size and stability, mainly due to the toxic effect exerted on soil microorganisms (Yang et al., 2017). Microplastics also reduce aggregates water stability, but the overall effect varies among microplastic types and concentrations (Mbachu et al., 2021; de Souza Machado et al., 2018b).
On the other hand, the presence of organic contaminants, such as total petroleum hydrocarbons and PAHs, has a positive effect on the water stability of soil aggregates due to the associated increase in organic carbon but also produce major water repellency, which could have an impact on soil erodibility (Klamerus-Iwan et al., 2015; Vogeler, 2009). The misapplication3 of sewage sludge, manure or irrigation with wastewater with a high load of inorganic and organic contaminants can clog soil pores and reduce hydraulic conductivity (Mahmoud et al., 2010; Schacht and Marschner, 2015; Zhang and Liu, 2018). The clogging of the upper layer may produce the formation of a hydrophobic layer that leads to waterlogging. However, these soil amendments can also contribute to the stabilization of soil aggregates and to the immobilisation of inorganic contaminants such as trace elements (Li et al., 2011; Vogeler, 2009).
Pollution-derived changes in aggregate formation and aggregate water stability, SOC content, and in overall soil structure will have a direct impact on soil erodibility. Soil erodibility refers to the resistance of the soil to the impact of raindrops on the soil surface and to the shearing action of surface runoff (FAO, 2019), and it is determined by two parameters: critical shear stress (τc) and erodibility coefficient (kd). Contaminants such as trace elements or synthetic detergents have been experimentally shown to increase soil erodibility (Chuchkalov, Fadeev and Alekseev, 2020; Salah and Al-Madhhachi, 2016).
In addition, soil erosion contributes to the transport of soil contaminants to distant areas and other environmental compartments. As modelled by Breshears et al. (2012), the amount of contaminants transported by soil erosion, both by saltation motion and aerial transport of contaminated soil particles, may account for 10 percent of the total contaminants transported on a global scale under undisturbed conditions after 1 000 years, and can raise up to 25 percent if surface disturbances occur, such as fires or extreme climatic events (droughts) due to warmer and drier climate. This model is of particular relevance for radionuclides, for which dispersion can greatly increase after disturbances (Breshears et al., 2012). Resuspension and transport of radionuclides after forest fires affecting the Chernobyl exclusion zone were first tested experimentally (Yoschenko et al., 2006) and later demonstrated after real forest fires (Evangeliou et al., 2016, 2014). About 10.9 TBq of radiocesium-137 were mobilised from the 2015 wildfires in the Chernobyl exclusion zone, reaching distant areas in the Russia Federation, Finland, Latvia, Lithuania, Eastern and Central Europe, and the Balkans region (Evangeliou et al., 2016).
4.2.1.4. Soil pollution as a driver of acidification
Soil acidification can occur naturally due to weathering of acidic parent rocks or dissolution and washing away of calcium carbonates by rainfall in warm, humid environments. Anthropogenic pressures, however, greatly contribute to soil acidification (Filipek, 1994; Verheye, 2009). Overuse of nitrogen fertilizers is consider the major driver of soil acidification, coupled with industrial and transport emission and consequent acid deposition (see Chapter 5) (Meng et al., 2019; Tian and Niu, 2015). Soil acidification, like soil pollution, is another overlooked soil health threat, due to the reliance on the buffering capacity of soils. However, some research indicates that the soil may be reaching its limit for buffering capacity (Lockwood et al., 2003; Tian and Niu, 2015).
In parallel to the increase in atmospheric CO2, nitrogen oxides (NOx) and sulphur dioxide (SO2) species in the atmosphere have also increased as a result of industrial emissions, coal burning and traffic. These compounds are transformed into nitric and sulphuric acids by photochemical reactions, and return to soil as acid rain (Verheye, 2009). Although SO2 emissions have reduced drastically since the 1990s in some regions, such as Europe or North America (both by 70 percent) and South America (30 percent) (EEA, 2014; Ritchie and Roser, 2019), these emissions continue to represent an important source of air pollution and acidifying factor for soils at the global level (Smith et al., 2011). The NOx emissions also continue to climb (Ehhalt et al., 2001). Wet and dry deposition of atmospheric contaminants onto soils leads to direct acidification and alteration of the soil buffer capacity.
Soils have a natural buffering capacity and compensate acid additions by releasing base cations bonded on exchange sites (Filipek, 1994). This buffer capacity varies among soil parent materials and soil exchangeable cations. The H+ additions to the soil solution first produces leaching of Ca2+ and Mg2+ cations. If the acid addition is too high or prolonged over time, the cation exchange capacity is exhausted and leads to a strong decrease in pH and release of non-base cations, such as Al3+, Mn2+ or Fe3+ (Tian and Niu, 2015). Three degradation processes will therefore occur:
- non-base cations can be toxic to plants and soil organisms (Čiamporová, 2002; Graefe and Beylich, 2003; Illmer, Marschall and Schinner, 1995), and produce nutrient deficiencies in plants (Filipek, 1994);
- Al, Mn and Fe are essential structural components of soil, forming Al/Mn/Fe-oxides and part of the octahedral and tetrahedral structures of clays; these elements are replaced in the exchangeable positions by H+ at high concentrations leading to a destruction of the soil structure (Barshad, 1960; Paradelo, van Oort and Chenu, 2013);
- lower pH will increase the bioavailability of many trace element, accentuating the effects of soil pollution (see Section 3.1.1.1).
In agricultural areas, soil acidification is mainly caused by the application of mineral nitrogen fertilisers and livestock urine, and by the repeated harvesting of plant biomass (Tian and Niu, 2015). However, due to the reduction in crop yields caused by N-fertilisation acidification, these negative effects have historically been overcome with liming and addition of organic matter which reduce the acidification risk (Filipek, 1994).
The major forms of N-fertilisers are ammonium (NH4+), nitrate (NO3-), ammonia (NH3) and urea (CO(NH₂)₂). Ammonium salts show greater acidifying capacity than urea and anhydrous ammonia fertilisers (Verheye, 2009). Filipek, (1994) related an application rate of 70 kg N/ha in the form of ammonium sulphate with an acidification potential of 7 700 mol H+/ha in Polish agricultural soils. According to the latest report of UNEP and FAO on Environmental and health effects of pesticides and fertilisers (UNEP, 2021), the current application rates of N from inorganic fertilisers in countries with higher consumption (e.g. 367 kg N/ha in Egypt, 243 kg N/ha in the Netherlands, or 220 kg N/ha in China) could have a significant acidifying potential in those countries with low fertilizer use efficiency, where a significant amount of N is leached or emitted as NH3 (Elrys et al., 2019). Tian and Niu (2015) estimated that soil pH has reduced globally an average of 0.26 due to acid deposition and N additions.
When N-fertilisers are applied, N-reduced species are oxidised to nitrite (NO2-) and nitrate (NO3-) by nitrifying bacteria in soils, releasing H+ protons in the soil media. Both species are highly mobile in soils and can be leached to groundwater (Abdelwaheb et al., 2019; Babiker et al., 2004; Refsgaard et al., 1999). Nitrate and nitrite contaminated drinking water has severe health impacts, causing methemoglobinemia and reproductive/developmental effects (Bouchard, Williams and Surampalli, 1992). Additionally, both species can migrate to surface waters and into the marine environments causing eutrophication (Figure 9). Within the soil matrix as the continuation of the nitrogen cycle, denitrifying bacteria ultimately convert nitrate into elemental nitrogen (N2) with nitric and nitrous oxide (NO and N2O) as intermediate species. Both are highly volatile greenhouse gases that can escape from soils into the atmosphere before denitrification process is completed (Lehnert et al., 2015) depending on soil conditions. The NO and N2O represent significant emissions from soils since not all soil bacteria have the enzyme to complete the denitrification process. According to the IPCC report on Land and Climate Change, denitrifying bacteria are responsible for 80 percent of the emissions of N2O (Jia et al., 2019). N2O is a highly potent greenhouse gas with a global warming potential 256 times greater than CO2 (IPCC, 2014).
Figure 9. Nitrogen cycle.

4.2.2. Soil pollution impacts on terrestrial ecosystems and the food chain
Terrestrial ecosystems are under growing pressure due to climate change, overexploitation of natural resources, fragmentation and destruction of habitats, and chemical pollution. Ecosystems are somewhat resilient to these threats through changing community structure or avoidance strategies, which are observed at all levels of the trophic web (Kozlov and Zvereva, 2011). According to the meta-analysis conducted by Kozlov and Zvereva (2011) on the effects of pollution on terrestrial ecosystems (based on 206 point-source polluting activities located in 36 countries) ecotoxicological responses are highly variable and depend on the source of pollution, the main contaminants, the duration of the exposure, the trophic level and climate. In general, all trophic groups show reduced body size and survival with proximity to pollution source, while primary consumers are more resilient to pollution in terms of diversity and abundance than predators (Kozlov and Zvereva, 2011). However, responses to stress are site-specific and the generalization of local patterns to a global scale has high uncertainty.
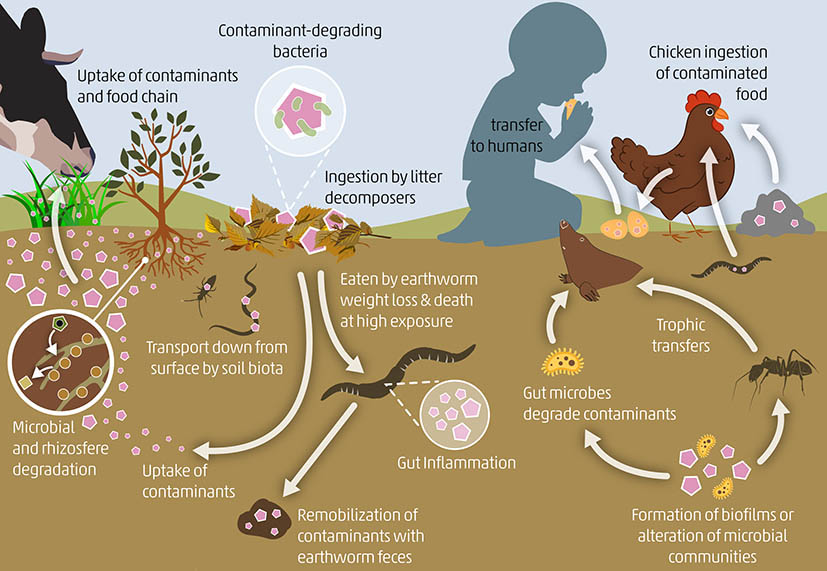
Pastures and crops can be affected by both diffuse and point source pollution either due to location near mining, industrial areas or waste deposits, or because they are subject to certain agricultural practices such as over-fertilisation, use of low-quality organic and inorganic fertilisers, irrigation with low-quality wastewater, or excessive or incorrect use of highly toxic pesticides (see Chapter 3). A wide range of contaminants are absorbed onto plant roots and translocated to edible tissues, and accumulated by soil-dwelling organisms (Abad-Valle et al., 2018; Huerta Lwanga et al., 2016; Lottermoser, 2011). Contaminated plants and soil organisms lead to potentially hazardous accumulations in animals higher in the food web such as grazing animals, birds, amphibians or mammals (Figure 10) (Huerta-Lwanga et al., 2017; Khan et al., 2020; Shen, Chi and Xiong, 2019).
Figure 10. Contaminant transfer into the terrestrial food web from the soil to pastures and crops, which are ingested by wildlife, livestock and humans, or from the soil to invertebrates, ingested by birds and poultry and ultimately transferred to humans.
Plants
In general, fewer effects have been observed in higher plant species than in soil micro- and macroorganisms (Domene et al., 2009; Hernández and Pastor, 2008), although changes in community structure and species richness have been reported (Dazy et al., 2009; Strandberg et al., 2006). In the past decades, plant tolerance to soil contaminants, such as trace elements and organic contaminants, has been studied and recognized (Tyler et al., 1989). Many plant species tolerate certain levels of soil contaminants (see Table 2 in Chapter 13) and have mechanisms to mitigate impact, for example, sequestration in the root system or immobilisation into the rhizosphere through the excretion of chelating exudates (for further details see phytoremediation options in Chapter 13).
Microorganisms present in the rhizosphere also contribute to the protection of plants against soil contaminants. Arbuscular mycorrizhal fungi (AMF) promote plant growth and contribute to the chelation of trace elements with fungal exudates and accumulation of contaminants in the rhizosphere, thereby avoiding transference to the shoots (Ferreira et al., 2018; Gao et al., 2011). AMF, bacterial symbionts, and other microorganisms colonizing the rhizosphere also contribute to the transformation and degradation of organic contaminants (Dubrovskaya et al., 2014; Fester et al., 2014). Microorganisms use organic contaminants as a source of carbon and energy and the additional supply of carbon from the plant root exudates can accelerate the biodegradation of contaminants by promoting microbial growth and activity. This win-win situation is exploited in enhanced phytoremediation processes (Gao et al., 2011; Xun et al., 2015) (see Chapter 13). However, in contrast, contaminants, when present as mixtures, can result in reduced degradation of organic contaminants and accumulation of contaminants in plant roots and translocation to shoots, as observed for the mixture pyrene/phenanthrene or trace elements/PAHs (Ekumankama, 2015; Gao et al., 2011). Adamczyk-Szabela et al. (2017) also observed increased trace element uptake and mobilization in above-ground plant biomass after soil fungicide treatments, which reduced soil AMF biomass.
Unlike many organic contaminants, which can serve as a source of carbon and energy for soil microorganisms, trace elements are not biodegradable and cannot be chemically eliminated. These contaminants can accumulate (bio-accumulation) and concentrate (biomagnification) in the food chain (Wu et al., 2010). Several trace elements have an essential role in plant growth and metabolism (e.g. iron, manganese, zinc, chromium, cobalt, copper, magnesium, molybdenum, and nickel), while others do not have a known function (e.g. arsenic, cadmium, lead, or mercury). Certain plant species are demonstrated hyperaccumulators of trace elements, absorbing through the roots and translocating to the shoots, contributing to the clean-up of the polluted soil (see Chapter 13). However, this translocation can also lead to the entrance of trace elements into the food chain when trace element-enriched parts are ingested by livestock, wild animals and humans (Corzo Remigio et al., 2020). In addition, hyperaccumulators can pose an environmental risk if harvested and residues not disposed properly, with the wider dispersion of the contaminants (Nascimento et al., 2015; Tian and Zhang, 2016).
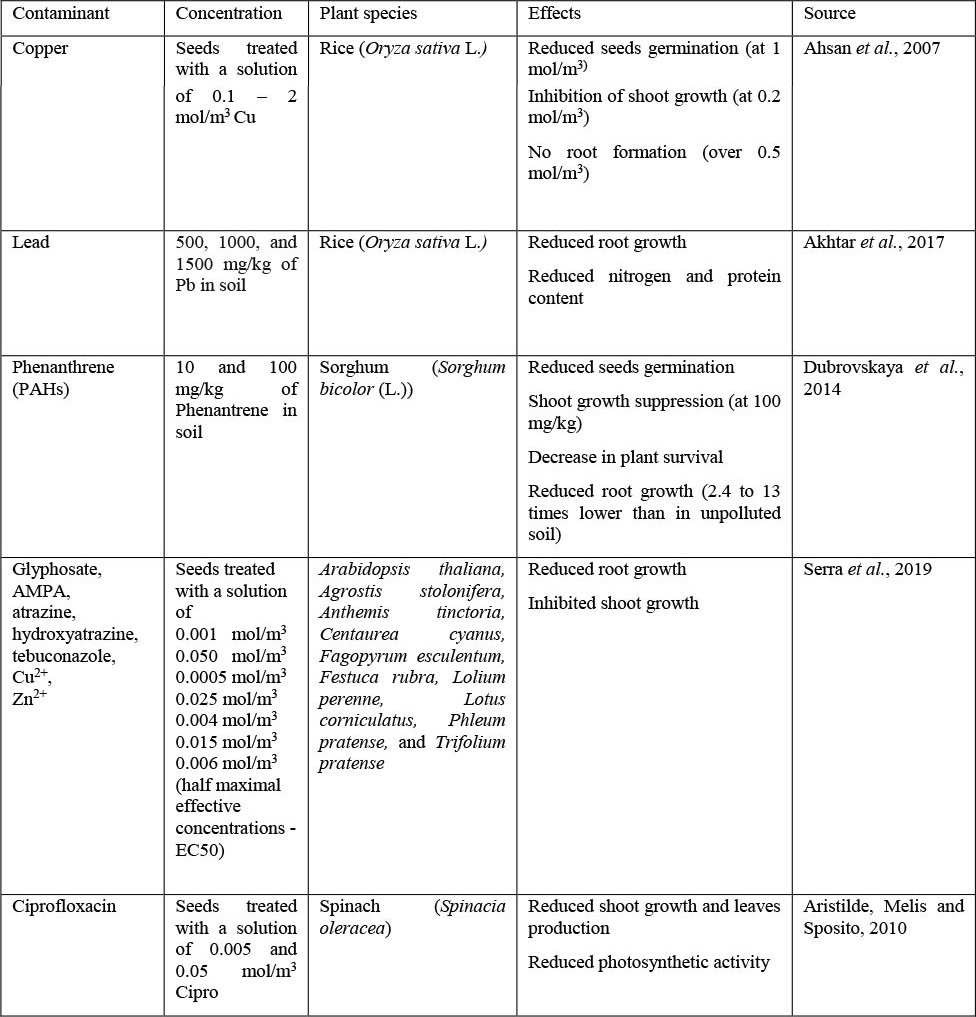
The uptake of trace elements and the exposure to organic contaminants (e.g. PAHs, pesticides, petroleum hydrocarbons, PPCPs) during plant germination and development stages can also produce growth inhibition and reduction of yields. Negative effects have been observed both in laboratory and field studies (Table 1) in seed germination, plant growth, root elongation, or photosynthesis (Ahsan et al., 2007; Akhtar et al., 2017; Aristilde, Melis and Sposito, 2010; Dubrovskaya et al., 2014; Serra et al., 2019; Tiecher et al., 2016). Root shortening and thickening can be a defence strategy to reduce the absorption of contaminants (Ferreira et al., 2018; Marques et al., 2018), but a general reduction in plant biomass is expected (Dubrovskaya et al., 2014; Ferreira et al., 2018). Higher expression of stress-related proteins has also been reported, for example in rice grown on copper-polluted soils (Ahsan et al., 2007). The decrease in plant biomass and photosynthesis leads to the reduction in primary production and crop yield, with economic impacts and threat to food security in countries where crops are grown in highly polluted agricultural soils.
Table 1. Some examples of contaminants-induced effects on higher plants
As previously mentioned, the decrease in litter decomposition and accumulation of non-decomposed organic material on the surface of polluted soils not only hampers soil organic carbon sequestration and nutrient cycling, but has shown an important impact on the physical status of terrestrial ecosystems and habitat availability (Köhler and Triebskorn, 2013). The accumulation of thick layers of litter on the surface prevents the germination of seed and the establishment of new plants, while also representing a reservoir of trace elements and other contaminants that will be released slowly due to lower rates of microbial and decomposer activity (Kozlov and Zvereva, 2015).
Terrestrial vertebrates
Thousands of contaminants present in terrestrial ecosystems have the potential to affect terrestrial wildlife and livestock. Normally, wildlife exposure to contaminants occurs at a low dose over lifetime span (chronic exposure), unless a large contaminant release is encountered (acute exposure). Soil contaminants can have neurotoxic, carcinogenic, teratogenic and endocrine disrupting effects in vertebrates (see Section 4.3.2). Soil contaminants exposure can cause changes in the expression of genes, proteins and hormones, or change organ histology or weight, decrease individual size, reduce reproductive output or cause premature death in cases of severe poisoning (Rhind et al., 2010). The effects on animal health and behaviour depend on the type of contaminant and the mix of contaminants to which animals are exposed. The living and dietary habits determine the time and intensity of exposure, as well as the development and immune status of the animal, in that immature or diseased organisms are more susceptible to negative effects or are less able to avoid polluted areas/food (Death, Griffiths and Story, 2019; French et al., 2017; Strebl and Tataruch, 2007). Herbivores also differ from omnivores in terms of exposure to contaminants, with the former accumulating higher concentrations of trace elements or radionuclides and the latter higher concentrations of lipophilic contaminants, such as PCBs or PFAS (Kowalczyk et al., 2018; Smith et al., 2007). Animals with smaller (potentially contaminated) feeding areas are at greater risk than those that can forage over larger areas (Smith et al., 2007). As observed in insects and other small soil organisms, terrestrial vertebrates use avoidance strategies and changes in food preferences in polluted areas can alter the whole terrestrial food web and prey-predator equilibrium (Ozaki et al., 2018). Reptiles and amphibians are particularly sensitive to hydrophilic and ionic contaminants which can pass through permeable egg walls, and result in dermal absorption (Sparling et al., 2010). Small mammals that burrow into the ground are exposed to soil pollution through inhalation, accidental ingestion of soil, dermal contact and direct ingestion of other contaminated organisms (Shore et al., 2014). Long-lived higher organisms are at risk of chronic exposure to soil contaminants from ingestion of these small mammals, although the changes may only be observed at the community rather than individual level (Gramss and Voigt, 2014).
Soil pollution can affect the food web in both natural ecosystems and in agricultural areas, with mammals and humans at the top of food webs being the most affected. Both livestock and wild animals can access croplands and grasslands where agrochemicals have been used, scavenge in open dumps or drink contaminated water (Ogbomida et al., 2018). According to scenarios to meet global food demand in 2030 developed by Zabel and co-workers, agricultural areas will expand and intensify, especially in tropical regions, threatening wildlife due to increased agrochemicals inputs and habitat fragmentation (Zabel et al., 2019).
The ingestion of polluted soil particles adhered to plant roots and direct ingestion of polluted soil while grazing are important pathways for contaminant intake in grazing livestock and wildlife (Gramss and Voigt, 2014; Loganathan, Hedley and Grace, 2008). Cattle can ingest as much as 1.5 kg of soil a day while grazing (Rychen et al., 2008). Accidental ingestion of soil particles by grazing sheep increases tenfold in winter months (Loganathan, Hedley and Grace, 2008). Soil can represent as much as 26 percent of the diet for root-grazing pigs or 60 percent for ground-digging birds (Gramss and Voigt, 2014). Soil ingestion was one of the main pathways for arsenic and lead intake for grazing animals in the vicinity of an abandoned mine in Spain (Abad-Valle et al., 2018). The transfer of cadmium and fluorine from phosphate-fertilised soils to grassing animals has also been reported in several countries (Loganathan, Hedley and Grace, 2008). Fluorine can cause chronic fluorosis in grazing animals and the consequently loss of productivity (Kim, Taylor and Drewry, 2016). The daily allowable intake of lead, cadmium, copper, manganese, zinc and uranium is surpassed by several multiples by grazing wildlife animals feeding from forage and rangeland plants and the associated polluted soil, in a metalliferous uranium mine in Germany (Gramss and Voigt, 2014).
The studies on ecotoxicological effects on wildlife increased significantly after the publication of Silent Spring (Carson, 1962). Most of the studies on wildlife have analysed the concentrations of contaminants in the tissues of dead organisms or in the edible organs of livestock and game which may pose a risk to human health through ingestion, but little is known about the effects of soil pollution at the developmental and behavioural level of the individual, nor the impact at the community or ecosystem level (Figure 11). Terrestrial wildlife is expected to be less exposed to soil contaminants than livestock. Natural ecosystems normally show low concentrations of soil contaminants but vary widely in terms of specific contaminants. The health effects of chronic low-dose exposure and the synergistic, summative or antagonistic effects of such mixtures have generally been neglected in terrestrial ecosystems (Death, Griffiths and Story, 2019). The following are some specific examples of health problems caused by single contaminants.
Figure 11. Ecotoxicological effects of soil contaminants on wildlife and terrestrial ecosystems.

The bio-availability of trace elements depends largely on soil properties (see Chapter 2), yet they are ubiquitous and persistent in the environment. Consequently, trace elements are among the most common contaminants detected in plant and animal tissues. However, few studies analyse the impact of exposure to trace elements on grazing animal health. Shen, Chi and Xiong (2019) reported diverse health problems, such as anaemia, anorexia and weakness, in sheep fed with cadmium and lead contaminated forage from pastures near a smelting facility in China. Oxidative stress, reduction of male reproductive function, bone mineralization and altered immune response have been reported in wild species in a mining area in south-central Spain where high levels of lead in blood and organs of grazing livestock were also observed (Pareja-Carrera, Mateo and Rodríguez-Estival, 2014 and references therein). According to animal experiments, arsenic toxicity causes diarrhoea at low doses and may have more severe effects on the gastrointestinal tract, kidney, thyroid and reproductive system at higher doses (JECFA, 2011b). High concentrations of arsenic, cadmium, mercury and lead were also found in the gizzard, liver, kidney and to less extent in muscles of free-range chickens, goats and sheep near gold mines in Ghana (Bortey-Sam et al., 2015). In amphibians, trace elements have been shown to alter thyroid function and delay the rate of development and metamorphosis (Thambirajah et al., 2019).
Mercury is ubiquitous in the environment in different oxidative states and has attracted much concern due to its impact on human health. Many mercury forms are highly volatile, and in soil mercury is mainly found in inorganic forms or organometallic form (methylmercury) which is produced through methylation by anaerobic bacteria. Methylmercury can easily pass the cell barrier and be transferred from soil to plants and to animals through the gastrointestinal track which represents more than 90 percent of mercury uptake by animals (Kalisinska, Lisowski and Kosik-Bogacka, 2012). Methylmercury is highly toxic to animals, capable of crossing the blood-brain barrier, and accumulates in tissues and organs, thus biomagnifying in the terrestrial food chain. The accumulation of methylmercury in insects can cause serious physiological, neurological, behavioural and reproductive disorders, and can be transferred to invertebrate predators, birds and mammals through the diet, which may cause central nervous system disorders (Kalisinska, Lisowski and Kosik-Bogacka, 2012; Yung et al., 2019). Methylmercury shows a wide range of health effects in wildlife, from neurotoxic effects to alterations of the reproductive system (Chételat et al., 2020). Mercury accumulation in the kidney and liver of cows and sheep has been reported as a result of transfer from plants and soils, in polluted sites near mining and industrial areas in Portugal, although the latter to a lesser extent (Henriques et al., 2013; Rodrigues et al., 2012). Maternal transference of mercury to the eggs or the foetus through placental transference has also been reported in wildlife (Chételat et al., 2020).
Other contaminants have also been reported to bioaccumulate in wild animals and livestock. For example, the accumulation of radiocaesium (137Cs) over long periods in wild animals living in the areas affected by the Chernobyl and Fukushima nuclear accidents has been reported, despite the radionuclides’ decay and concentration decrease in plants and soils over time (IAEA et al., 2009; Tagami, Howard and Uchida, 2016). In addition to direct exposure to radiation after these accidents, wildlife was exposed to radionuclides through dietary intake of contaminated plants and polluted soil particles which increased exposure to ionizing radiation by at least an order of magnitude above the ambient dose (Beresford et al., 2020). The assimilation by plants of the long-lived isotope of caesium, 137Cs, may stabilize at a certain level and even decrease as a result of sorption to the soil matrix. Different crops on the same soil bio-accumulate radiocesium to different extents in the following order buckwheat > beans, soybeans > peas > oats > wheat, rye > millet, barley > triticale > corn, with the accumulation of radiocaesium in buckwheat 18 times higher than in corn (Dutov and Yermolayev, 2013). European wild boar showed higher concentrations of 137Cs compared to other animals after the Chernobyl accident, which was related to the seasonal ingestion of mushrooms and roots (Strebl and Tataruch, 2007). Nemoto, Saito and Oomachi (2018) observed a positive relationship between the concentration of 137C in the soil and in the muscles of wild boars and bears in Fukushima Prefecture, with concentrations higher in the former due to their feeding habits that include the intake of roots and tubers. Exposure to radioactivity causes a reduction of body size, pathological changes of internal organs (liver, kidneys, or lungs), and necrosis leading to individual death, with resulting decrease in the general population (Sazykina and Kryshev, 2006). Taking into account that wild boars and bears are game prey, the risk of transmission of contaminants to humans should be assessed (Nemoto, Saito and Oomachi, 2018; Strebl and Tataruch, 2007).
Many organic contaminants, including pesticides, PCBs, some PFAS or phthalates, are neurotoxic, carcinogenic and endocrine disruptors (Stockholm Convention, 2021). Although pesticide use is now regulated in many countries, acute exposure of wildlife still occurs in others where pesticide use is less well regulated. Recent research has focused on the impact of long-term low-dose pesticide exposure on wildlife (Köhler and Triebskorn, 2013). Garcês, Pires and Rodrigues (2020), Gibbons, Morrissey and Mineau (2015), and Köhler and Triebskorn (2013) have summarized direct and indirect effects of pesticides on wildlife at environmental concentrations, which range from reduced growth and reproduction to behavioural disorders, teratogenic defects in offspring, and neurotoxic effects at all levels of organization, from invertebrates to mammals, birds and reptiles, and in individuals and populations.
Wild animals that feed on terrestrial plants and seeds or small mammals in areas surrounding agricultural areas are especially exposed to pesticides and other organic contaminants (Golden and Rattner, 2003). Rychen et al. (2008) reported a transference of organic contaminants from soils to ruminant, ranging from 5 to 90 percent for PCBs, 1 to 40 percent for PCDD/Fs and 0.5 to 8 percent for PAHs. Exposure to a mixture of organic and inorganic contaminants, such as that observed in soils treated with sewage sludge, produces an alteration in the behaviour of the ruminant offspring, reducing the ability to obtain food and compete with other individuals (Rhind et al., 2010). Soil-plant-vertebrate transference of contaminants has also been reported for PFOA and PFOS, for which liver accumulation has been observed in domestic pigs and wild boars (Kowalczyk et al., 2018).
Endocrine-disrupting contaminants are especially relevant for amphibians, since thyroid hormones regulate many steps of the metamorphosis process, causing changes in genes and proteins expressions to the reduction of metamorphosis, reproduction rates, and survival (Thambirajah et al., 2019).
Pharmaceuticals and veterinary medicines are widely present in the environment, especially in areas irrigated with wastewater or fertilized with treated-animal manure or sewage sludge, or in areas of intensive livestock production (Sukul and Spiteller, 2007; Van Doorslaer et al., 2014). Once in the soil matrix, antibiotics undergo several degradation processes, both by biotransformation and biodegradation mediated by fungi enzymes, and by abiotic processes through reactions with soil oxides or photodegradation (Sukul and Spiteller, 2007). However, some antibiotics, including fluoroquinolones, may persist for several months in sludge and manure amended soils (Albero et al., 2018; Peruchi, Fostier and Rath, 2015; Rusu, Hancu and Uivaroşi, 2015).
Environmental concentrations of antibiotics in soils irrigated with wastewater or in soils treated with manure or biosolids are generally low: average agricultural soil concentrations are in the range of tens of ng/kg to hundreds of µg/kg (An et al., 2015; Gothwal and Shashidhar, 2015; Li et al., 2015a). However, after long-term continuous application, soil concentrations may become environmentally relevant, with concentrations greater than 8 000 µg/ kg having been reported in greenhouse agricultural soils of China under continuous application of manure (Zhang et al., 2016). Several laboratory and field studies have demonstrated that plants can uptake antibiotics from the soil solution (Bártíková et al., 2015 and references therein; Sallach, 2015; Tasho and Cho, 2016). Long-term exposure may result in significant accumulation of antibiotics in plant tissues which may cause damage to organisms feeding on contaminated plants, despite no phytotoxic symptoms evident in the plants (Pan and Chu, 2017; Riaz et al., 2018). Faeces and urine from livestock treated with antibiotics can reach freshwater streams, ponds and lakes that supply wildlife. Wildlife can also be affected by feeding on carcasses of livestock treated with drugs, as occurred with vultures in India and Pakistan (Oaks and Watson, 2011). In a survey conducted in Spain, up to 92 percent of vultures analysed presented different concentrations of fluoroquinolones residues (veterinary antibiotic) originating from livestock (Blanco et al., 2016). There is strong evidence for the emergence of antimicrobial resistant bacteria in wildlife, especially those that forage close to agricultural lands or that share grazing areas with livestock (Allen et al., 2010; Dias et al., 2015; Rogers et al., 2018). This antibacterial resistance can be gained due to the exposure of wildlife gut bacteria to antibiotics and residues through the ingestion of contaminated plants, carcasses or contaminated water (Chowdhury, Langenkämper and Grote, 2016; Riaz et al., 2018) or due to the exposure to antimicrobial resistant bacteria present in sewage or manure treated soils ingested with soil while grazing (Chen et al., 2019b; Grenni, Ancona and Barra Caracciolo, 2018; Rogers et al., 2018; Tyrrell et al., 2019). The accidental consumption of pharmaceuticals and veterinary drugs by wildlife can cause alteration of natural gut microbiota or hypersensitivity reactions, among other developmental disorders such as alterations of cartilages or bones growth, together with the development of antimicrobial resistance of gut bacteria and exogenous bacteria (Blanco, Lemus and García-Montijano, 2011; Hruba et al., 2019).
Many countries have established biomonitoring programs to characterize changes in ecosystems using key bioindicator species (Golden and Rattner, 2003). Selecting the correct species to monitor for environmental pollution could support the improved understanding of contaminants in soils given the greater ease of analysing contaminants in animal samples than in soil. Furthermore, the total concentration of contaminants in soil does not provide sufficient information on the environmental impact due to the different bioavailability and interactions among contaminants, and between contaminants and the soil matrix. Several European countries have established specific systems to monitor environmental pollution caused by contaminants such as pesticides, PCBs or trace elements, which use raptors as sentinel organisms; however, these systems are not connected or harmonised (Espín et al., 2016; Gómez-Ramírez et al., 2014). In France, a toxicovigilance system for pesticide effects in wildlife, SAGIR, has been in place since late 1960s focused on acute poisoning and wildlife deaths associated to exposure to pesticides (Berny et al., 2010). For over two decades, the U.S. Fish and Wildlife Service has established a contaminant monitoring system for wildlife, named the National Contaminant Biomonitoring Program (NCBP) (LaRoe and US National Biological Service, 1995). Although this is mainly focused on fish monitoring, other terrestrial species have also been analysed. The monitoring system registered 207 separate bird and mammal mortality incidents related to organophosphorus or carbamate pesticides alone throughout the United States. The Canadian Government also has monitoring programmes in place for assessing contaminants levels and the impact on wildlife species, including birds and amphibians (Canada, 2013). These biomonitoring programmes are essential to rapidly detect environmental pollution, and support risk assessment tools for prioritization of intervention areas. Nevertheless, the biomonitoring programmes should focus not only on measuring contaminants concentrations in sentinel species but also on monitoring behavioural changes to detect sub-lethal effects that will threaten species resilience (Saaristo et al., 2018).
4.2.3. Soil pollution impacts on aquatic ecosystems
Precipitation, floods, snowmelt, and irrigation increases the water content in soil pores and, once soil is saturated, leads to waterlogging of flat areas and run-off from slopes. Dissolved organic matter and entrained contaminants, and fine soil particles with sorbed contaminants are carried by runoff water and can reach nearby wetlands, rivers and lakes, and ultimately be transported to seas and oceans (Shi and Schulin, 2018). This transport of soil particles increases water turbidity and can reduce channel depth. Borrelli et al. (2017) estimated an annual average of soil loss and mobilization due to erosion accounting for 35 Pg4 of topsoil per year. According to the calculations done by Kok, Papendick and Saxton (2009), one third of soil eroded from agricultural areas ends up in water bodies, which represents 12 Pg of agricultural topsoil and associated nutrients and contaminants being discharged in streams, rivers, lakes and coastal areas annually. Additionally, plants growing in polluted areas that accumulate contaminants into their tissues could also represent a pathway of contaminant dispersion to surrounding areas and neighbouring water bodies through litter dispersion (Maunoury-Danger et al., 2018).
On the other hand, soil pore water has also a vertical movement through soil pores favouring the transport of dissolved and particulate contaminants by drainage or leaching, reaching surface water through subsurface drainage network or the groundwater table, respectively.
In the marine environment, plastic and nutrient contamination are of primary interest, although other contaminants can also reach water bodies from land-based activities and affect the equilibrium and health of aquatic ecosystems. Land-based activities are responsible for over 80 percent of marine pollution (Cicin-Sain et al., 2011). A large body of scientific evidence is available that analyses the impacts of different contaminants on the health and performance of aquatic organisms. In fact, the ecotoxicology of fish and other marine organisms is much more developed than that for terrestrial organisms.
Most contaminants in aquatic ecosystems come from anthropogenic land-based sources. Therefore, soil management can have an enormous impact on water quality, including pollution. However, it is beyond the scope of this report to go into detail on the effects of each of the contaminants that could have a terrestrial origin on marine and aquatic organisms. Some reviews have recently been published that are recommended reading to complement the information in this report (Cuevas, Martins and Costa, 2018; Elizalde-Velázquez and Gómez-Oliván, 2020; Girones et al., 2020; Jacquin et al., 2020; Kallenborn et al., 2018; Kumari et al., 2017; Li et al., 2017a; Pereira et al., 2020; Zhang, Zhao and Fent, 2020).
Eutrophication
One of the main problems of global water pollution is eutrophication caused by land-based activities. As explained in Section 4.2.1.4 relative to acidification, agricultural soils with excessive nutrient loading leads to eutrophication of water bodies. FAO estimates that 109 million tonnes of nitrogen and 45 million tonnes of phosphorus were applied to agricultural soils worldwide in 2017 (FAOSTAT, 2020). With nutrient efficiencies ranging from 30 to 100 kg of harvest dry matter per kg N input and 60 to 700 kg harvest dry matter per kg P input, an average of 50 percent of nutrients added to soils are being leached or washed into rivers, lakes and ultimately seas and oceans (Bouwman et al., 2017; Bouwman, Beusen and Billen, 2010; Cosme, Mayorga and Hauschild, 2018).
Nitrite and nitrate are highly mobile in the soil matrix and leach easily to groundwater. Nitrate interacts with sulphates and iron in soils and sediments, contributing to phosphorous mobilization and therefore enhanced eutrophication (Smolders et al., 2010):
- Nitrate uses the iron disulfide (FeS2) in the pyritic rocks as an electron donor, releasing sulphate (SO42-) into the soil solution and groundwater (Hayakawa et al., 2013). Sulphates ions are highly efficient in oxidizing organic compounds (Rastogi, Al-Abed and Dionysiou, 2009), enhanced through a microbial-mediated process involved in the mineralization of organic matter. Oxidised sulfides increase H+ concentrations and the acidity is toxic to aquatic animals causing even massive deaths. Phosphorous is released during the mineralization of organic molecules in hypoxic or anoxic conditions (Brock and Schulz-Vogt, 2011; Sinkko et al., 2013);
- Sulphate ions can be reduced in aquatic ecosystems to sulfide, which binds again with dissolved reduced iron ions forming FeS2. Phosphate sorbed or bonded to the iron (hydr-)oxides is then released (Smolders et al., 2010);
Therefore, the higher the load of nitrates, the higher the eutrophication of waterbodies by the phosphate surplus (Smolders et al., 2010). Phosphate surplus is the major driver of freshwater eutrophication whereas in the marine environment eutrophication is mainly due to great inputs of nitrogen, which is the major limiting nutrient (Smith, Joye and Howarth, 2006). This feedback of nitrate and phosphate eutrophication cycles produces what are known as massive “dead zones” in many of the world’s major marine water bodies, such as the Gulf of Mexico, the Bay of Bengal, Hudson Bay, the Mediterranean Sea or the Baltic Sea (Cosme, Mayorga and Hauschild, 2018; Lehnert et al., 2015; Sinkko et al., 2013). Well known examples of freshwater eutrophication include Lake Erie in North America, Lake Winnipeg in Canada, or Lake Victoria in eastern Africa (Prepas and Charette, 2003). Excessive nutrients favour the growth of harmful and nuisance algal blooms (Wurtsbaugh, Paerl and Dodds, 2019). When these large amounts of algae die and sink to the lake/ocean floor, bacteria decomposing the algae consume significant amounts of oxygen. This reduction or complete elimination of oxygen in these regions of deeper, cooler waters (hypoxic or anoxic zones, respectively), popularly referred to as “dead zones”, result in avoidance by or death of aquatic organisms that normally reside in these areas. Some of the algae that make up these blooms, such as some species of cyanobacteria, also produce compounds that are highly toxic to aquatic organisms, wildlife, domestic animals, livestock and humans (Lehnert et al., 2015; Watson et al., 2016). In addition, nitrates in drinking water results in significant health effects both in animals and humans, causing alteration of the thyroid functioning and disruption of gonadal steroidogenesis (Guillette and Edwards, 2005). Nitrate has been observed to reduce progesterone production and reduce reproductive success in small mammals (Inoue et al., 2004). Excess nitrate (> 10 mg/L) in drinking water from contaminated wells also causes infant methemoglobinemia (“blue-baby syndrome”).
Micro-and nanoplastics
Plastics in terrestrial systems is itself a problem (see Chapter 2 and Chapter 3). It has been estimated that about 80 percent of the plastic entering the ocean has a land origin (Li, Tse and Fok, 2016). According to Jambeck et al. (2015), mismanaged waste disposal in coastal countries is responsible for 4.8 to 12.7 million metric tonnes of plastic entering the ocean by wind erosion, or transported via waterways or wastewater, while between 75 000 to 1.1 million tonnes are discharged in rivers from in-land activities (Figure 12) (Eunomia, 2016).
Figure 12. Mass concentration of surface plastics in marine, lake and river environments (g/km2).

Plastic items greatly vary in size and composition, from large items such as bottles, bags and package containers, to microbeads used in personal care products or microfibers used in clothing. Water treatment plants normally do not have the capacity to filter and retain those microfibers and micrometric pieces of plastics, so they end up being released into the environment (Okoffo et al., 2019). Once in the environment, plastics undergo processes of aging, breakdown and degradation (Barnes et al., 2009), accumulating in different habitats and affecting virtually all trophic levels (de Sá et al., 2018).
A significant proportion of those plastics are discharged directly into the aquatic environment (Alimi et al., 2018), but the majority of plastic debris of variable size accumulates in terrestrial environments. In agricultural soils alone, the microplastics content is estimated to be between 63 000 - 430 000 and 44 000 - 300 000 tonnes in Europe and North America, respectively (Nizzetto, Futter and Langaas, 2016). In agricultural land, plastics are sourced from direct application of mulching films and other agroplastics, or indirectly through irrigation with wastewater and application of sewage sludge. However, soils under other land uses also receive plastics from direct disposal in open dumpsites or poorly-managed landfills, from tire abrasion or from irrigation with wastewater or direct release of wastewater effluents on soils. Therefore, the amount of plastics accumulated in the world’s soils could be much greater than estimated (He et al., 2018).
Soils have the capacity of retain micro and nanoplastics, which sorb into organic matter, metallic oxides or clays, are occluded in aggregates, get trapped in small pores or are ingested and accumulated by soil organisms. Soil erosion by wind or water and floods can transport micro- and nanoplastics sorbed to soil particles, even to remote areas (Bläsing and Amelung, 2018) and the leaching of smaller particles into the groundwater can also be significant. However, as pointed out by He et al. (2018) the number of studies on plastic fate in soils is much lower than in other environmental compartments (152 papers about fate in soils versus 226 about fate in marine environments available in Scopus) and the uncertainties on plastic fate in soil environment is very high (Alimi et al., 2018). The lack of an efficient reliable method for detecting and quantifying microplastics in soil is the major limiting barrier in research. Degradation, retention, and leaching are conditioned by soil physicochemical and biological properties, plastics chemical composition, tridimensional structure and size, and soil pore water characteristics. Studies combining all these aspects are practically non-existent, which limits our understanding of the full extent of the problem. However, it seems clear that soils are important reservoirs of micro- and nano-plastics that contribute to the pollution of aquatic environments (Hurley and Nizzetto, 2018).
The presence of microplastics in the marine environment and the impact on aquatic species have been extensively studied (Andrady, 2011; Auta, Emenike and Fauziah, 2017; Wang, Ge and Yu, 2020). Despite the substantial increase in ecotoxicological studies over the last decade (Figure 13), there is still a lack of analysis at environmentally relevant concentrations covering all groups of organisms to better understand their impact and trophic transfer (Au et al., 2017). Once ingested, micro- and nano-plastics can have various effects on aquatic organisms, ranging from physiological alterations in phytoplankton and aquatic plants (alteration of chlorophyll production), to appetite inhibition, reduced growth or even collapse of the digestive system due to accumulation of plastics in vertebrates and consequent death (Wang et al., 2019b). Additionally, plastics adsorb other contaminants into their surfaces, such as trace elements, PAHs, pesticides, and other organic contaminants, contributing to their transfer to the food web and causing additional health effects on aquatic organisms (Alimi et al., 2018).
Figure 13. Number of scientific publications in Scopus database containing “ecotoxicology”, “plastics” and “aquatic organisms”.

Other contaminants
Other land-based contaminants can also reach the freshwater and marine environments with effects including food chain contamination, bio-accumulation and biomagnification (Saha, Ghosh and Basak, 2019). Some examples of water pollution with soil contaminants from land-based activities are presented below.
Trace elements are the most widely distributed contaminants in soils worldwide. Trace elements can be transported by erosion and runoff to nearby water bodies and, when bound to sediments, can be transported long distances. Apart from atmospheric long range transport and wet and dry deposition (see Chapter 5), trace elements mobilized from soils due to erosion represent an important source of coastal contamination (Häder et al., 2020; Kwasigroch et al., 2018; Polidoro et al., 2017). Mining and industrial activities in coastal areas represent the most important source of trace elements to the marine environment (Häder et al., 2020). Streams and rivers passing near agricultural areas, landfills and industrial sites are subject to contaminated sediment inputs that wash downstream to the coast (Polidoro et al., 2017; Wang et al., 2015). Mercury is a particularly concerning trace element affecting aquatic environments, due to its high toxicity and transference within the food web (Landrigan et al., 2020). Land-based erosion and leaching to groundwater are equally important sources of mercury than direct releases of effluents and atmospheric deposition (Kocman et al., 2017). Approximately 7 000-8 000 tons of mercury are dumped into soils each year from industrial and mining (both large and small-scale artisanal gold mining) activities, and these can be washed into watercourses and coastal areas (UNEP, 2019).
Organic contaminants can also be dissolved in runoff water or be transported with soil particles, colloids and even bounded to other contaminants, such as microplastics. Some pesticides have a high leaching potential and longer half-life in water than in soil. Metolachlor (one of the most widely used herbicides) has been found to have higher persistence in water (Sun et al., 2020) than bromacil or tebuthiuron (Gillespie, Czapar and Hager, 2011). Despite having been banned for two decades, organochlorine pesticides can still be detected in the marine environment, especially in areas close to major agricultural regions, such as Argentina and Brazil in South America (Girones et al., 2020), or the Baltic Sea (Sonne et al., 2020). Although usage is decreasing, areas in which these pesticides are still in use exist and represent major threats to the marine environment, such as in the Sea of Japan (Law, 2014; Tsygankov and Lukyanova, 2019). Pesticides in aquatic and marine environments can have adverse effects in non-targeted organisms, causing from endocrine disruption to neurological and behavioural alterations (Cuevas, Martins and Costa, 2018; Li et al., 2017a).
Per- and polyfluoroalkyl compounds (PFASs) are considered “forever” contaminants of emerging concern due to high persistence in the environment and high toxicity. PFASs are widely used industrial chemicals that are released locally to soils (Langberg, 2021). Diffuse PFAS contamination, leaching to groundwater, and erosion of PFASs-polluted soil and runoff are key pathways for PFASs entry into aquatic environments and for global circulation (Brumovský et al., 2016; Pistocchi and Loos, 2009; Yamashita et al., 2004), of which, marine ecosystems are the ultimate reservoir (Yamashita et al., 2008). Marine waters show a heterogeneous distribution of PFASs, with higher concentrations near coastal point sources of pollution and river deltas with higher upstream discharge of contaminants (Langberg, 2021; Pistocchi and Loos, 2009). PFASs are soluble and lipophilic and hence have been detected in the marine food chain, although the scarce ecotoxicological information is still inconclusive (Ali et al., 2021). Although the scarce ecotoxicological information is still inconclusive on the effects of PFASs on aquatic organisms, the evidence of the impact on human health is increasingly strong (Landrigan et al., 2020).
Pharmaceuticals and personal care products (PPCPs) also represent a significant risk to aquatic ecosystems. These compounds primarily enter the aquatic environment from direct discharge into water bodies from pharmaceutical manufacturing or from urban wastewater treatment plants (see Chapter 2). PPCPs can also reach rivers, lagoons and oceans when sorbed to soil particles suspended in surface water by erosion processes or leaching from soils treated with sewage sludge or manure, or irrigated with wastewater (Boxall et al., 2012; Felis et al., 2020; Klatte, Schaefer and Hempel, 2017). PPCPs can accumulate in aquatic organisms (Landrigan et al., 2020), where these compounds show endocrine disruption effects, alter sexual characteristics and reproductive effectiveness inter alia (Elizalde-Velázquez and Gómez-Oliván, 2020; Gworek et al., 2019; Jacquin et al., 2020; Majumder, Gupta and Gupta, 2019; Ohoro et al., 2019).
Aquatic ecosystems, like soils, are therefore exposed to complex mixtures of contaminants, and the available information on their fate, uptake by aquatic organisms and entry into the food chain is still limited (Landrigan et al., 2020). In addition, aquatic ecosystems are exposed to multiple environmental stressors such as increased acidification due to higher concentration of atmospheric CO2, rising temperatures, and anthropogenic overexploitation, which exacerbate the negative effects of pollution (Chapman, 2017; Gunderson, Armstrong and Stillman, 2016). Therefore, reducing the level of pollution in soils both by more sustainable soil management practices and by remediating already contaminated and polluted soils, coupled with stronger regulations and policies, will reduce the impact on aquatic environmental compartments (Häder et al., 2020).
- 1 Many of the antibiotic resistance genes carry plasmids, transposons or integrons that can act as vectors that transfer these genes to other “contemporary” neighboring members of the same bacterial species, as well as to bacteria of another genus or species (MSU, 2011).
- 2 Set of moisture and temperature conditions prevailing within a soil (see Glossary)
- 3 Misapplication refers to excessive application at one time or repeated applications over time, or application at the wrong time (e.g., before rain or on frozen fields) or manner (e.g., sprayed on the surface instead of incorporated into the shallow subsurface of the soil)
- 4 Pg= petagram = billion tonnes
- 5 This dataset is a compilation of all data produced by Oceaneye between 2011 and 2019 during several monitoring expeditions with the use of Manta Nets (mesh size = 330 um). The sampling and analysis methodology is described in Faure & al. 2015. The columns pc_num_km2 and pc_g_km2 feature respectively the surface plastic concentration in numbers of particles per km2 and the surface plastic concentration in mass per km2. Values are non-adjusted plastic concentration for waves height or wind.